
Oxidative Stress Markers and Histopathological Changes in Selected Organs of Mice Infected with Murine Norovirus 1 (MNV-1)
 , , , , , ,
, , , , , ,
Abstract
1. Introduction
2. Results
2.1. Assessing the Effectiveness of Infection
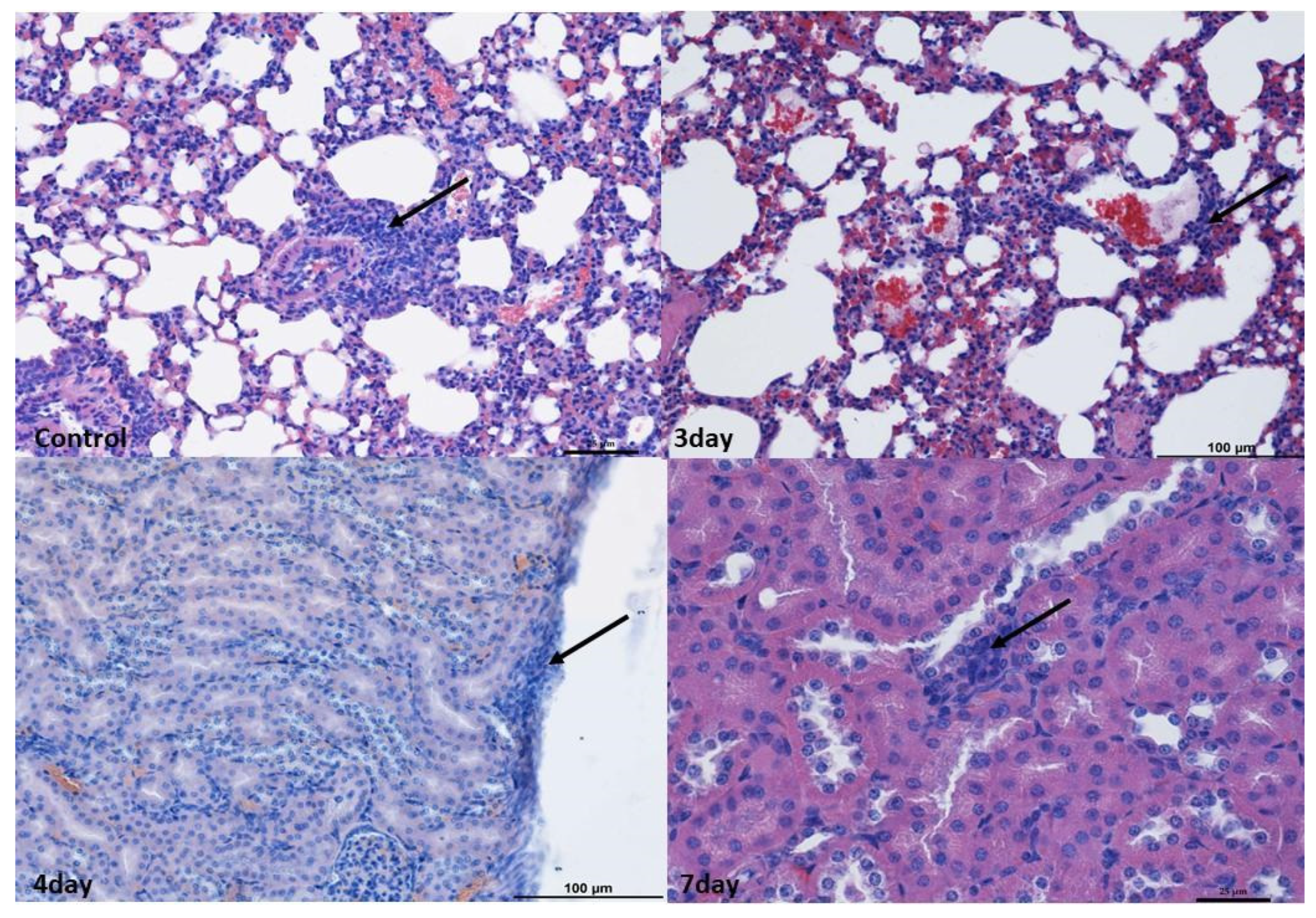
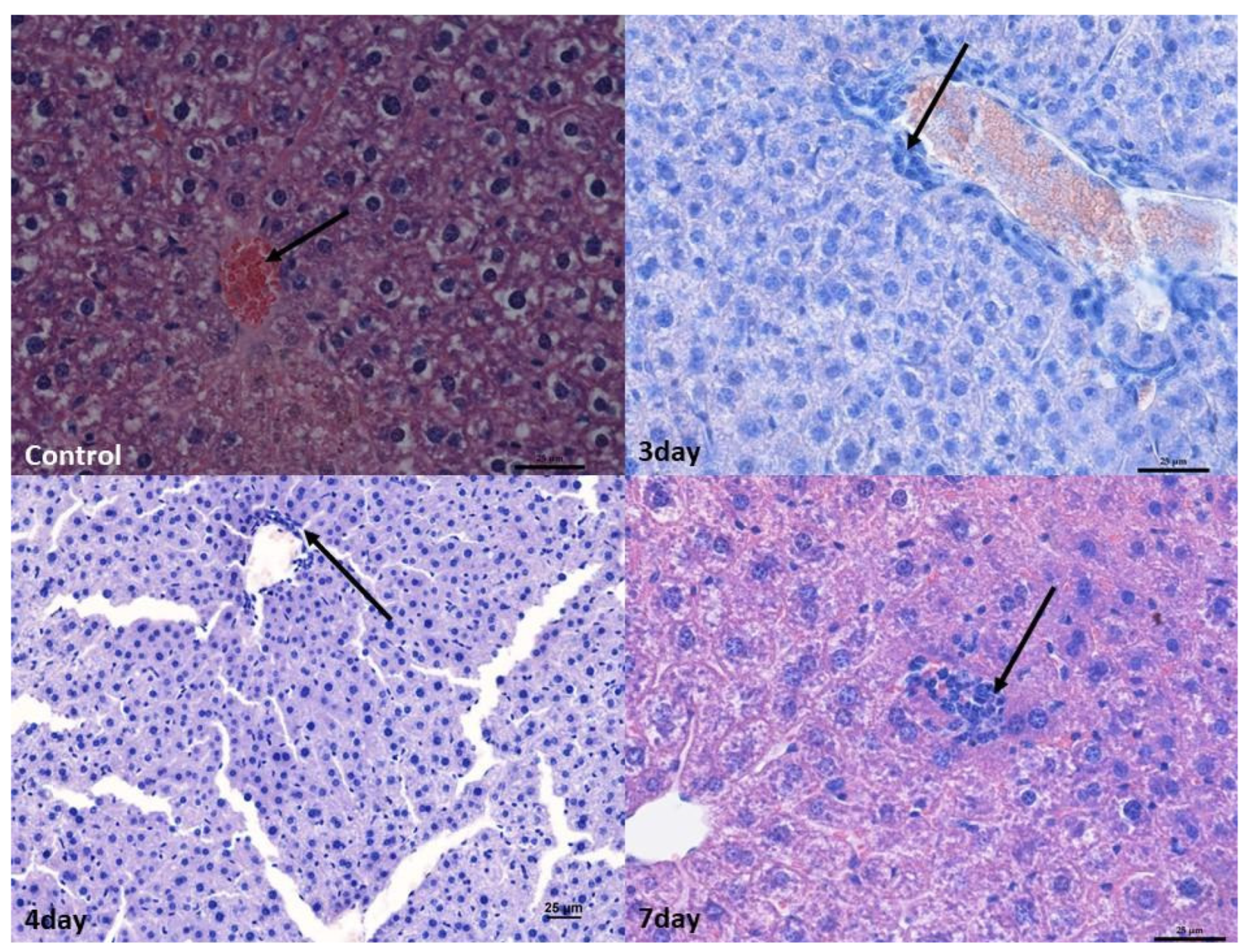
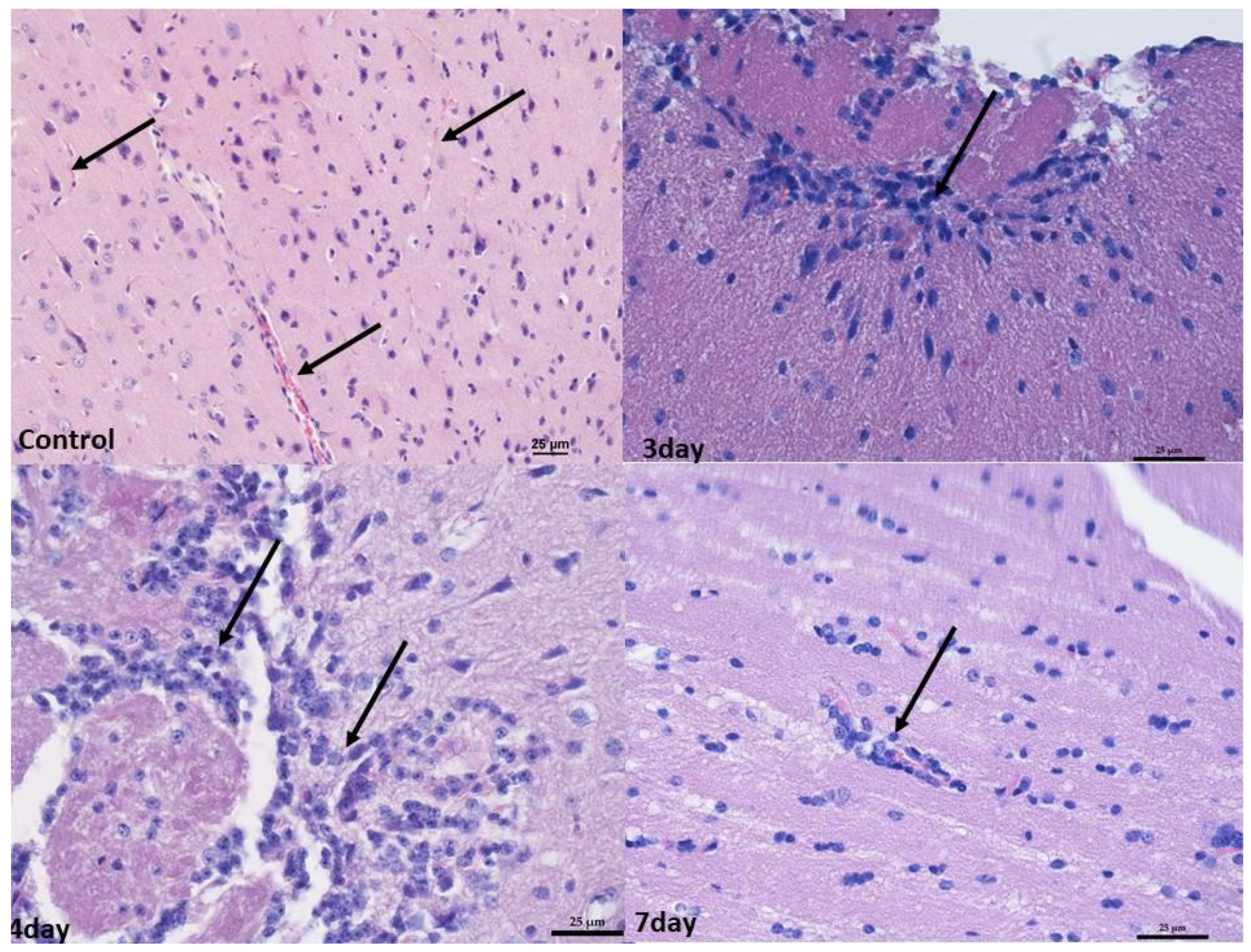
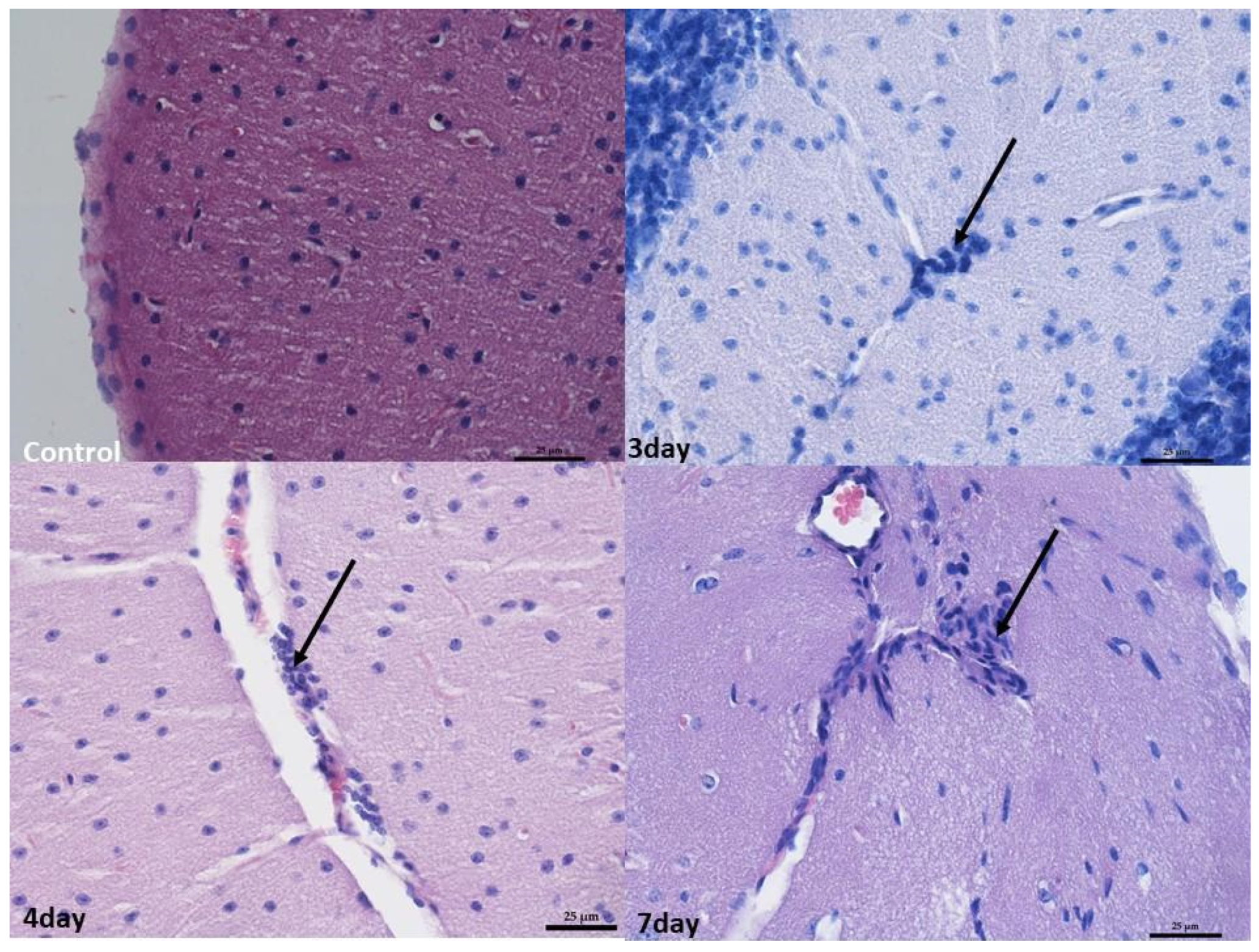
2.2. Monitoring Histopathological Changes in Organs of MNV-Infected Mice
2.3. Oxidative Stress Markers
3. Discussion
4. Materials and Methods
4.1. Mice
- Healthy controls (control);
- Euthanized 3 days after infection (3 days);
- Euthanized 4 days after infection (4 days);
- Euthanized 7 days after infection (7 days).
4.2. Monitoring the Kinetics of MNV-1 Infection in C57Bl/6J Mice
4.3. Cell Culture
4.4. Virus Propagation
4.5. Histological Examination
4.6. Oxidative Stress
4.6.1. Superoxide Dismutase (SOD) (EC 1.15.1.1) Activity
4.6.2. Catalase (CAT) Activity (EC 1.11.1.6)
4.6.3. Glutathione Peroxidase (GPx) Activity (EC 1.11.1.9)
4.6.4. Glutathione Reductase (GR) Activity (EC 1.8.1.7)
4.6.5. Glutathione-S Transferase (GST) Activity (EC 2.5.1.18)
4.6.6. Total Oxidant Status (TOS) and Total Antioxidant Capacity (TAC)
4.6.7. Malondialdehyde (MDA) Concentration
4.6.8. Statistical Analysis of Antioxidant Stress Markers
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Fernandes, I.G.; de Brito, C.A.; Dos Reis, V.M.S.; Sato, M.N.; Pereira, N.Z. SARS-CoV-2 and Other Respiratory Viruses: What Does Oxidative Stress Have to Do with It? Oxid. Med. Cell. Longev. 2020, 2020, 8844280. [Google Scholar] [CrossRef]
- Buesa, J.; Collado, B.; Lopez-Andujar, P.; Abu-Mallouh, R.; Rodriguez Diaz, J.; Garcia Diaz, A.; Prat, J.; Guix, S.; Llovet, T.; Prats, G.; et al. Molecular epidemiology of caliciviruses causing outbreaks and sporadic cases of acute gastroenteritis in Spain. J. Clin. Microbiol. 2002, 40, 2854–2859. [Google Scholar] [CrossRef]
- Green, K.Y. Caliciviridae: The noroviruses. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins Health: Philadelphia, PA, USA, 2013; pp. 582–608. [Google Scholar]
- Karst, S.M.; Wobus, C.E.; Lay, M.; Davidson, J.; Virgin, H.W., 4th. STAT1-dependent innate immunity to a Norwalk-like virus. Science 2003, 299, 1575–1578. [Google Scholar] [CrossRef]
- Hsu, C.C.; Wobus, C.E.; Steffen, E.K.; Riley, L.K.; Livingston, R.S. Development of a microsphere-based serologic multiplexed fluorescent immunoassay and a reverse transcriptase PCR assay to detect murine norovirus 1 infection in mice. Clin. Vaccine Immunol. 2005, 12, 1145–1151. [Google Scholar] [CrossRef]
- Kahan, S.M.; Liu, G.; Reinhard, M.K.; Hsu, C.C.; Livingston, R.S.; Karst, S.M. Comparative murine norovirus studies reveal a lack of correlation between intestinal virus titers and enteric pathology. Virology 2011, 421, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Strong, D.W.; Thackray, L.B.; Smith, T.J.; Virgin, H.W. Protruding domain of capsid protein is necessary and sufficient to determine murine norovirus replication and pathogenesis in vivo. J. Virol. 2012, 86, 2950–2958. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.R.; Seok, S.H.; Kim, D.J.; Baek, M.W.; Na, Y.R.; Han, J.H.; Kim, T.H.; Park, J.H.; Turner, P.V.; Chung, D.H.; et al. Prevalence of murine norovirus infection in Korean laboratory animal facilities. J. Vet. Med. Sci. 2011, 73, 687–691. [Google Scholar] [CrossRef] [PubMed]
- Pritchett-Corning, K.R.; Cosentino, J.; Clifford, C.B. Contemporary prevalence of infectious agents in laboratory mice and rats. Lab. Anim. 2009, 43, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Muller, B.; Klemm, U.; Mas Marques, A.; Schreier, E. Genetic diversity and recombination of murine noroviruses in immunocompromised mice. Arch. Virol. 2007, 152, 1709–1719. [Google Scholar] [CrossRef] [PubMed]
- Henderson, K.S. Murine norovirus, a recently discovered and highly prevalent viral agent of mice. Lab. Anim. 2008, 37, 314–320. [Google Scholar] [CrossRef]
- Kitajima, M.; Oka, T.; Tohya, Y.; Katayama, H.; Takeda, N.; Katayama, K. Development of a broadly reactive nested reverse transcription-PCR assay to detect murine noroviruses, and investigation of the prevalence of murine noroviruses in laboratory mice in Japan. Microbiol. Immunol. 2009, 53, 531–534. [Google Scholar] [CrossRef]
- Kelmenson, J.A.; Pomerleau, D.P.; Griffey, S.; Zhang, W.; Karolak, M.J.; Fahey, J.R. Kinetics of transmission, infectivity, and genome stability of two novel mouse norovirus isolates in breeding mice. Comp. Med. 2009, 59, 27–36. [Google Scholar]
- McFadden, N.; Bailey, D.; Carrara, G.; Benson, A.; Chaudhry, Y.; Shortland, A.; Heeney, J.; Yarovinsky, F.; Simmonds, P.; Macdonald, A.; et al. Norovirus regulation of the innate immune response and apoptosis occurs via the product of the alternative open reading frame 4. PLoS Pathog. 2011, 7, e1002413. [Google Scholar] [CrossRef]
- Basic, M.; Keubler, L.M.; Buettner, M.; Achard, M.; Breves, G.; Schröder, B.; Smoczek, A.; Jörns, A.; Wedekind, D.; Zschemisch, N.H.; et al. Norovirus triggered microbiota-driven mucosal inflammation in interleukin 10-deficient mice. Inflamm. Bowel Dis. 2014, 20, 431–443. [Google Scholar] [CrossRef]
- Cecchini, R.; Cecchini, A.L. SARS-CoV-2 infection pathogenesis is related to oxidative stress as a response to aggression. Med. Hypotheses 2020, 143, 110102. [Google Scholar] [CrossRef]
- Halliwell, B. Free radicals and antioxidants: Updating a personal view. Nutr. Rev. 2012, 70, 257–265. [Google Scholar] [CrossRef]
- Ntyonga-Pono, M.P. COVID-19 infection and oxidative stress: An under-explored approach for prevention and treatment? Pan Afr. Med. J. 2020, 35 (Suppl. S2), 12. [Google Scholar] [CrossRef]
- Kazura, W.; Michalczyk, K.; Skrzep-Poloczek, B.; Chełmecka, E.; Zalejska-Fiolka, J.; Michalski, M.; Kukla, M.; Jochem, J.; Rutkowski, J.; Stygar, D. Liver Oxidative Status, Serum Lipids Levels after Bariatric Surgery and High-Fat, High-Sugar Diet in Animal Model of Induced Obesity. Int. J. Mol. Sci. 2023, 24, 16535. [Google Scholar] [CrossRef]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2015, 15, 71. [Google Scholar] [CrossRef] [PubMed]
- Maloney, N.S.; Thackray, L.B.; Goel, G.; Hwang, S.; Duan, E.; Vachharajani, P.; Xavier, R.; Virgin, H.W. Essential cell-autonomous role for interferon (IFN) regulatory factor 1 in IFN-γ-mediated inhibition of norovirus replication in macrophages. J. Virol. 2012, 86, 12655–12664. [Google Scholar] [CrossRef]
- Balmus, I.M.; Ciobica, A.; Antioch, I.; Dobrin, R.; Timofte, D. Oxidative stress implications in the affective disorders: Main biomarkers, animal models relevance, genetic perspectives, and antioxidant approaches. Oxidative Med. Cell. Longev. 2016, 2016, 25. [Google Scholar] [CrossRef]
- Abedi, A.; Ghobadi, H.; Sharghi, A.; Iranpour, S.; Fazlzadeh, M.; Aslani, M.R. Effect of saffron supplementation on oxidative stress markers (MDA, TAC, TOS, GPx, SOD, and pro-oxidant/antioxidant balance): An updated systematic review and meta-analysis of randomized placebo-controlled trials. Front. Med. 2023, 10, 1071514. [Google Scholar] [CrossRef]
- Mumphrey, S.M.; Changotra, H.; Moore, T.N.; Heimann-Nichols, E.R.; Wobus, C.E.; Reilly, M.J.; Moghadamfalahi, M.; Shukla, D.; Karst, S.M. Murine norovirus 1 infection is associated with histopathological changes in immunocompetent hosts, but clinical disease is prevented by STAT1-dependent interferon responses. J. Virol. 2007, 81, 3251–3263. [Google Scholar] [CrossRef]
- Roth, A.N.; Helm, E.W.; Mirabelli, C.; Kirsche, E.; Smith, J.C.; Eurell, L.B.; Ghosh, S.; Altan-Bonnet, N.; Wobus, C.E.; Karst, S.M. Norovirus infection causes acute self-resolving diarrhea in wild-type neonatal mice. Nat. Commun. 2020, 11, 2968. [Google Scholar] [CrossRef]
- Dycke, J.V.; Neyts, J.; Rocha-Pereira, J. Assessing the Efficacy of Small Molecule Inhibitors in a Mouse Model of Persistent Norovirus Infection. Bio-Protocol 2018, 8, e2831. [Google Scholar] [CrossRef]
- Taube, S.; Kolawole, A.O.; Höhne, M.; Wilkinson, J.E.; Handley, S.A.; Perry, J.W.; Thackray, L.B.; Akkina, R.; Wobus, C.E. A mouse model for human norovirus. mBio 2013, 4, e00450-13. [Google Scholar] [CrossRef]
- Peiper, A.M.; Helm, E.W.; Nguyen, Q.; Phillips, M.; Williams, C.G.; Shah, D.; Tatum, S.; Iyer, N.; Grodzki, M.; Eurell, L.B.; et al. Infection of neonatal mice with the murine norovirus strain WU23 is a robust model to study norovirus pathogenesis. Lab. Anim 2023, 52, 119–129. [Google Scholar] [CrossRef]
- Hsu, C.C.; Piotrowski, S.L.; Meeker, S.M.; Smith, K.D.; Maggio-Price, L.; Treuting, P.M. histologic Lesions Induced by Murine Norovirus Infection in Laboratory Mice. Vet. Pathol. 2016, 53, 754–763. [Google Scholar] [CrossRef]
- Wobus, C.E.; Thackray, L.B.; Virgin IV, H.W. Murine Norovirus: A Model System to Study Norovirus, Biology and Pathogenesis. J. Virol. 2006, 80, 5104–5112. [Google Scholar] [CrossRef]
- Bohmwald, K.; Andrade, C.A.; Gálvez, N.M.S.; Mora, V.P.; Muñoz, J.T.; Kalergis, A.M. The Causes and Long-Term Consequences of Viral Encephalitis. Front. Cell. Neurosci. 2021, 15, 755875. [Google Scholar] [CrossRef]
- Marrocco, I.; Altieri, F.; Peluso, I. Measurement and Clinical Significance of Biomarkers of Oxidative Stress in Humans. Oxid. Med. Cell. Longev. 2017, 2017, 6501046. [Google Scholar] [CrossRef]
- Garofalo, R.P.; Kolli, D.; Casola, A. Respiratory syncytial virus infection: Mechanisms of redox control and novel therapeutic opportunities. Antioxid. Redox Signal. 2013, 18, 186–217. [Google Scholar] [CrossRef]
- Erel, O. A novel automated direct measurement mettivehod for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin. Biochem. 2004, 37, 277–285. [Google Scholar] [CrossRef]
- Karkhanei, B.; Talebi Ghane, E.; Mehri, F. Evaluation of oxidative stress level: Total antioxidant capacity, total oxidant status and glutathione activity in patients with COVID-19. New Microbes New Infect. 2021, 42, 100897. [Google Scholar] [CrossRef]
- Oyanagui, Y. Reevaluation of assay methods and establishment of kit for superoxide dismutase activity. Anal. Biochem. 1984, 142, 290–296. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Mannervik, B. Glutathione peroxidase. Methods Enzymol. 1985, 113, 490–495. [Google Scholar] [CrossRef]
- Carlberg, I.; Mannervik, B. Glutathione reductase. Methods Enzymol. 1985, 113, 484–490. [Google Scholar] [CrossRef]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem 2005, 38, 1103–1111. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oxidative Stress Marker | Group | Brain | Cerebellum | Liver | Lungs | Kidney | p |
|---|---|---|---|---|---|---|---|
| SOD (NU/mg protein) | control | 11.1 (9.0–14.1) | 15.0 (11.6–18.9) | 3.6 (3.3–4.8) | 4.1 (2.5–5.7) | 5.4 (4.8–6.8) | <0.001 |
| norovirus | 12.0 (11.2–13.4) | 15.0 (11.5–15.5) | 2.7 (2.4–2.8) | 4.9 (3.2–6.3) | 5.9 (5.0–6.1) | <0.001 | |
| pcontrol vs. norovirus | 0.443 | 0.443 | <0.01 | 0.523 | 0.701 | ||
| MnSOD (NU/mg protein) | control | 3.1 (2.1–4.1) | 2.5 (1.4–3.7) | 2.4 (2.2–2.9) | 2.6 (2.3–3.0) | 1.9 (1.8–2.0) | <0.05 |
| norovirus | 2.7 (2.6–3.0) | 2.6 (2.3–2.8) | 2.0 (1.9–2.2) | 2.6 (2.2–2.8) | 2.2 (2.1–2.4) | <0.01 | |
| pcontrol vs. norovirus | 0.609 | 0.609 | <0.05 | 0.701 | <0.05 | ||
| GR (IU/g protein) | control | 26.6 (25.7–28.0) | 26.8 (24.9–27.5) | 5.7 (5.4–7.4) | 13.5 (12.4–15.9) | 1.8 (1.5–2.4) | <0.001 |
| norovirus | 15.4 (15.0–17.8) | 21.9 (19.3–22.6) | 0.3 (0.2–0.3) | 18.3 (17.8–18.4) | 3.6 (2.6–4.7) | <0.001 | |
| pcontrol vs. norovirus | <0.01 | <0.01 | <0.01 | <0.01 | <0.05 | ||
| CAT (IU/g protein) | control | 184.1 (141.2–187.7) | 133.2 (108.1–147.1) | 396.4 (382.7–437.5) | 140.3 (133.4–144.3) | 290.8 (264.3–298.1) | <0.001 |
| norovirus | 154.0 (117.6–179.8) | 193.5 (186.3–238.9) | 225.6 (204.1–247.3) | 180.8 (166.1–187.7) | 358.2 (355.0–374.5) | <0.001 | |
| pcontrol vs. norovirus | 0.250 | <0.01 | <0.01 | <0.01 | <0.01 | ||
| GPx (IU/g protein) | control | 446 (257–555) | 488 (413–522) | 65 (60–69) | 496 (485–530) | 270 (208–401) | <0.001 |
| norovirus | 309 (271–358) | 256 (205–373) | 135 (113–150) | 389 (384–399) | 250 (245–270) | <0.001 | |
| pcontrol vs. norovirus | 0.201 | <0.01 | <0.01 | <0.01 | 0.443 | ||
| GST (IU/g protein) | control | 6.4 (6.2–7.0) | 8.1 (6.8–8.8) | 3.9 (3.1–4.6) | 6.2 (5.2–7.6) | 5.6 (5.2–6.6) | <0.001 |
| norovirus | 6.5 (5.4–6.8) | 7.5 (6.4–9.6) | 5.9 (4.7–7.5) | 6.4 (8.9–7.0) | 5.1 (5.1–5.2) | <0.05 | |
| pcontrol vs. norovirus | 0.609 | 1.0 | <0.01 | 0.798 | 0.196 | ||
| TAC (mmol/g protein) | control | 0.34 (0.32–0.38) | 0.41 (0.38–0.46) | 0.32 (0.31–0.36) | 0.26 (0.22–0.27) | 0.33 (0.32–0.44) | <0.001 |
| norovirus | 0.43 (0.41–0.53) | 0.16 (0.14–0.18) | 0.18 (0.11–0.24) | 0.25 (0.21–0.34) | 0.47 (0.44–0.48) | <0.001 | |
| pcontrol vs. norovirus | <0.01 | <0.01 | <0.01 | 0.256 | 0.070 | ||
| TOS (IU/g protein) | control | 0.45 (0.29–0.53) | 0.46 (0.43–0.56) | 0.40 (0.38–0.46) | 2.48 (2.20–2.76) | 0.62 (0.60–0.71) | <0.001 |
| norovirus | 0.74 (0.67–0.90) | 0.55 (0.51–0.59) | 0.87 (0.86–1.07) | 1.97 (1.54–2.14) | 0.43 (0.40–0.46) | <0.001 | |
| pcontrol vs. norovirus | <0.01 | 0.097 | <0.01 | 0.055 | <0.01 | ||
| MDA (IU/g protein) | control | 7.1 (6.5–8.2) | 14.3 (13.5–15.1) | 2.9 (2.8–3.6) | 3.3 (3.1–3.8) | 2.7 (2.1–3.2) | <0.001 |
| norovirus | 7.2 (6.7–8.4) | 12.1 (11.2–12.3) | 2.4 (2.1–2.6) | 3.7 (3.2–4.6) | 3.1 (2.8–3.8) | <0.001 | |
| pcontrol vs. norovirus | 0.898 | 0.055 | <0.05 | 0.523 | 0.246 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janicka, P.; Stygar, D.; Chełmecka, E.; Kuropka, P.; Miążek, A.; Studzińska, A.; Pogorzelska, A.; Pala, K.; Bażanów, B. Oxidative Stress Markers and Histopathological Changes in Selected Organs of Mice Infected with Murine Norovirus 1 (MNV-1). Int. J. Mol. Sci. 2024, 25, 3614. https://doi.org/10.3390/ijms25073614
Janicka P, Stygar D, Chełmecka E, Kuropka P, Miążek A, Studzińska A, Pogorzelska A, Pala K, Bażanów B. Oxidative Stress Markers and Histopathological Changes in Selected Organs of Mice Infected with Murine Norovirus 1 (MNV-1). International Journal of Molecular Sciences. 2024; 25(7):3614. https://doi.org/10.3390/ijms25073614
Chicago/Turabian StyleJanicka, Paulina, Dominika Stygar, Elżbieta Chełmecka, Piotr Kuropka, Arkadiusz Miążek, Aleksandra Studzińska, Aleksandra Pogorzelska, Katarzyna Pala, and Barbara Bażanów. 2024. "Oxidative Stress Markers and Histopathological Changes in Selected Organs of Mice Infected with Murine Norovirus 1 (MNV-1)" International Journal of Molecular Sciences 25, no. 7: 3614. https://doi.org/10.3390/ijms25073614
APA StyleJanicka, P., Stygar, D., Chełmecka, E., Kuropka, P., Miążek, A., Studzińska, A., Pogorzelska, A., Pala, K., & Bażanów, B. (2024). Oxidative Stress Markers and Histopathological Changes in Selected Organs of Mice Infected with Murine Norovirus 1 (MNV-1). International Journal of Molecular Sciences, 25(7), 3614. https://doi.org/10.3390/ijms25073614