
Optical Intracranial Self-Stimulation (oICSS): A New Behavioral Model for Studying Drug Reward and Aversion in Rodents

Abstract
1. Introduction
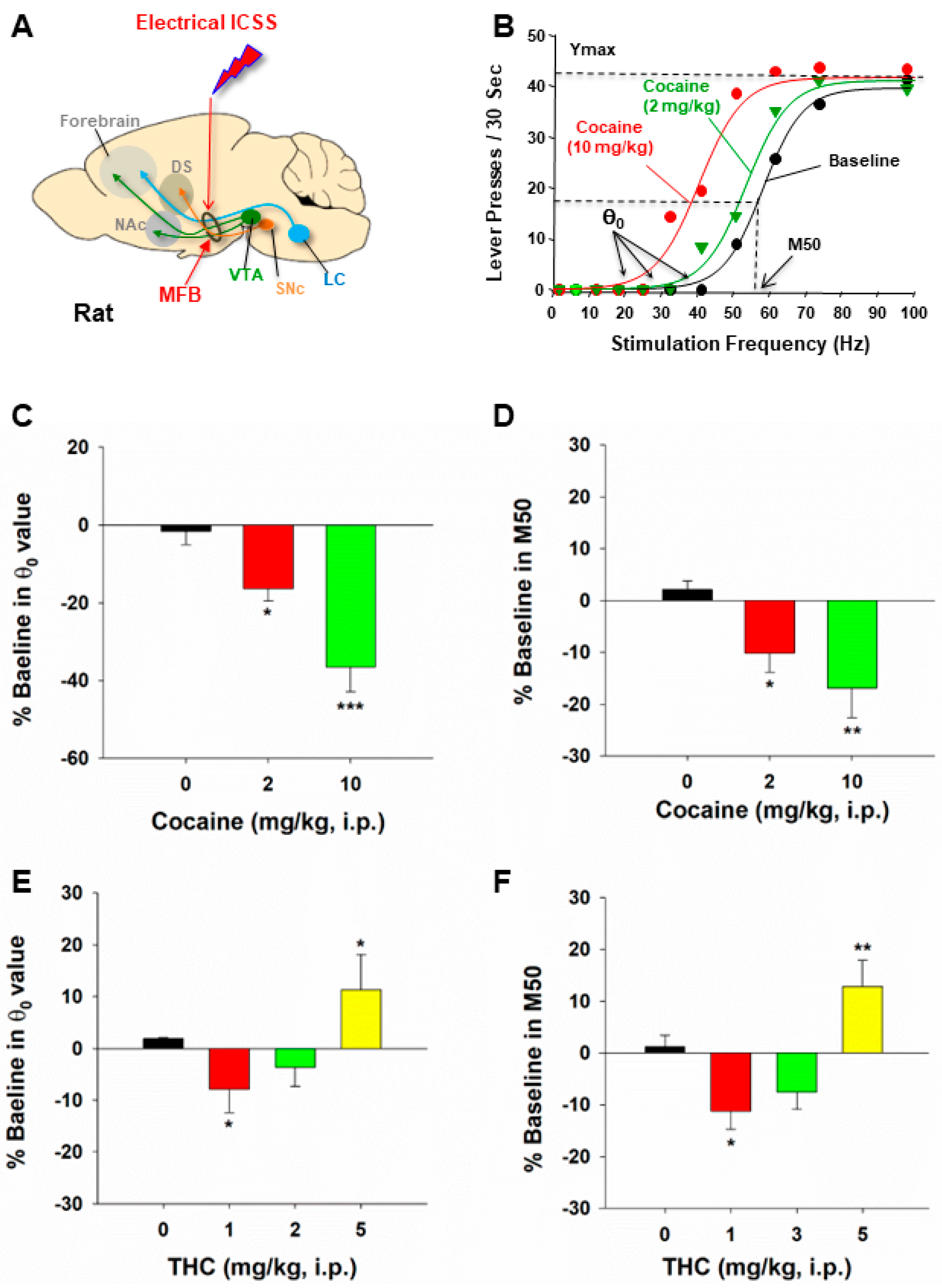
2. Principles of oICSS for Studying Drugs of Abuse
3. DA-Dependent oICSS and Its Implications in Studying Drug Reward versus Aversion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animals | Brain Region | Targeted Neurons | Opsins | Stim. Frequency | Finding | References |
|---|---|---|---|---|---|---|
Dopamine-dependent oICSS | ||||||
| TH-Cre rats | VTA | DA | ChR2 | 20 Hz | Produces oICSS | [4,48,54] |
| TH-Cre mice | VTA, SNc | DA | ChR2 | 25 Hz | Produces oICSS | [52,55] |
| TH-Cre mice | VTA | DA | ChR2 | 20 Hz | Produces oICSS | [56] |
| TH-Cre | VTA | DA | ChR2 | 20 Hz | Produces oICSS | [57,58,59,60,61] |
| DAT-Cre mice | VTA-NAc | DA terminals | ChR2 | 30 Hz | Produces oICSS | [62] |
| DAT-Cre | SNc | DA | ChR2 | 50 Hz | Produces oICSS | [63] |
| DAT-Cre mice | VTA | DA | ChR2 | 1, 5, 10, 25, 50, 100 Hz | Produces oICSS | [17,18,49,50,51] |
| DAT-Cre, Crhr1-, Cck-, mice | VTA | DA | ChR2 | 20 Hz | Produces oICSS | [53] |
| DAT-Cre mice | VTA | DA | ChR2 | 1, 5, 10, 20, 25, 50 Hz | Produces oICSS | [64] |
| DAT-Cre mice | VTA | DA | ChR2 | 1, 5, 10, 20, 25, 50, 65 Hz | Produces oICSS | [65] |
| DAT-Cre mice | VTA | DA | ChR2 | 20 Hz | Produces oICSS | [66] |
| DAT-Cre mice | NAc, PFC | DA terminals | ChR2 | 20 Hz | Produces oICSS | [66] |
| DAT-Cre | VTA | DA | ChR2 | 40 Hz | Produces oICSS | [67] |
| D1-Cre | Dentate gyrus | DA | Produces oICSS | |||
Glutamate-dependent oICSS | ||||||
| C57 WT mice | BLA-VTA | Glutamate | ChR2 | 20 Hz | Produces oICSS | [68] |
| C57 WT mice | vHipp-NAc | Glutamate | ChR2 | 20 Hz | Produces oICSS | [69] |
| VgluT2-Cre mice | VTA | Glutamate | ChR2 | 20 Hz | Produces oICSS | [35] |
| Thy1-ChR2-EYFP mice | NAc | Glutamate terminals | ChR2 | 20 Hz | Produces oICSS | [70] |
| VgluT2-Cre mice | Pedunculopontine | Glutamate | ChR2 | 10, 20, 30, 40 Hz | Produces oICSS | [71] |
| VgluT2-Cre mice | VTA | Glutamate | ChR2 | 1, 5, 10, 25, 50, 100 Hz | Produces oICSS | [14] |
| VgluT2-Cre mice | DMS, | Glutamate terminals | ChR2 | 20 Hz | Produces oICSS | [72] |
| VgluT2-Cre mice | NAc | Glutamate terminals | ChR2 | 40 Hz | Produces oICSS | [73] |
| VgluT2-Cre mice | VTA | Glutamate | ChR2 | 10, 20, 30, 40 Hz | Produces oICSS | [36] |
| VgluT2-Cre mice | VP, NAc, LHb | Glutamate terminals | ChR2 | 10, 20, 30, 40 Hz | Produces oICSS | [36] |
| VgluT2-Cre mice | RN | Glutamate | ChR2 | 20 Hz | Produces oICSS | [17] |
| VgluT2-Cre mice | VTA | Glutamate terminals | ChR2 | 20 Hz | Produces oICSS | [17] |
| VgluT2-Cre mice | Parabrachio-SNc | Glutamate terminals | ChR2 | 20 Hz | Produces oICSS | [74] |
GABA-dependent oICSS | ||||||
| C57 WT | NAc | GABA | ChR2 | 20 Hz | Produces oICSS | [69] |
| Vgat-Cre | SNr, | GABA | Halo | Constant, 20 s | Produces oICSS | [51] |
| Vgat-Cre | SNr | GABA | Arch3 | Constant, 3 s | Produces oICSS | [75] |
| D1-Cre mice | DS | D1-MSNs | ChR2 | Constant, 1 s | Produces oICSS | [76] |
| D1-Cre mice | DS | D1-MSNs | ChR2 | 40 Hz | Produces oICSS | [75] |
| D1-Cre mice | DS | D1-MSNs | ChR2 | 5 Hz | Produces oICSS | [77] |
| D1-Cre mice | NAc | D1-MSNs | ChR2 | 25 Hz | Produces oICSS | [78] |
Other substance-dependent oICSS | ||||||
| ePet-Cre mice | DRN | 5-HT | ChR2 | 5, 20 Hz | Produces oICSS | [79] |
| ePet-Cre mice | DRN | 5-HT | ChR2 | 20 Hz | Produces oICSS | [56] |
| SERT-Cre mice | DRN, 5-HT neurons | 5-HT | ChR2 | 40 Hz | Produces oICSS | [75] |
| Tac2-Cre mice | dMHb | Neurokinin-expressing | ChR2 | 20 Hz | Produces oICSS | [80] |
| D1-Cre | LC-DG | D1-expressing | ChR2 | 20 Hz | Produces oICSS | [81] |
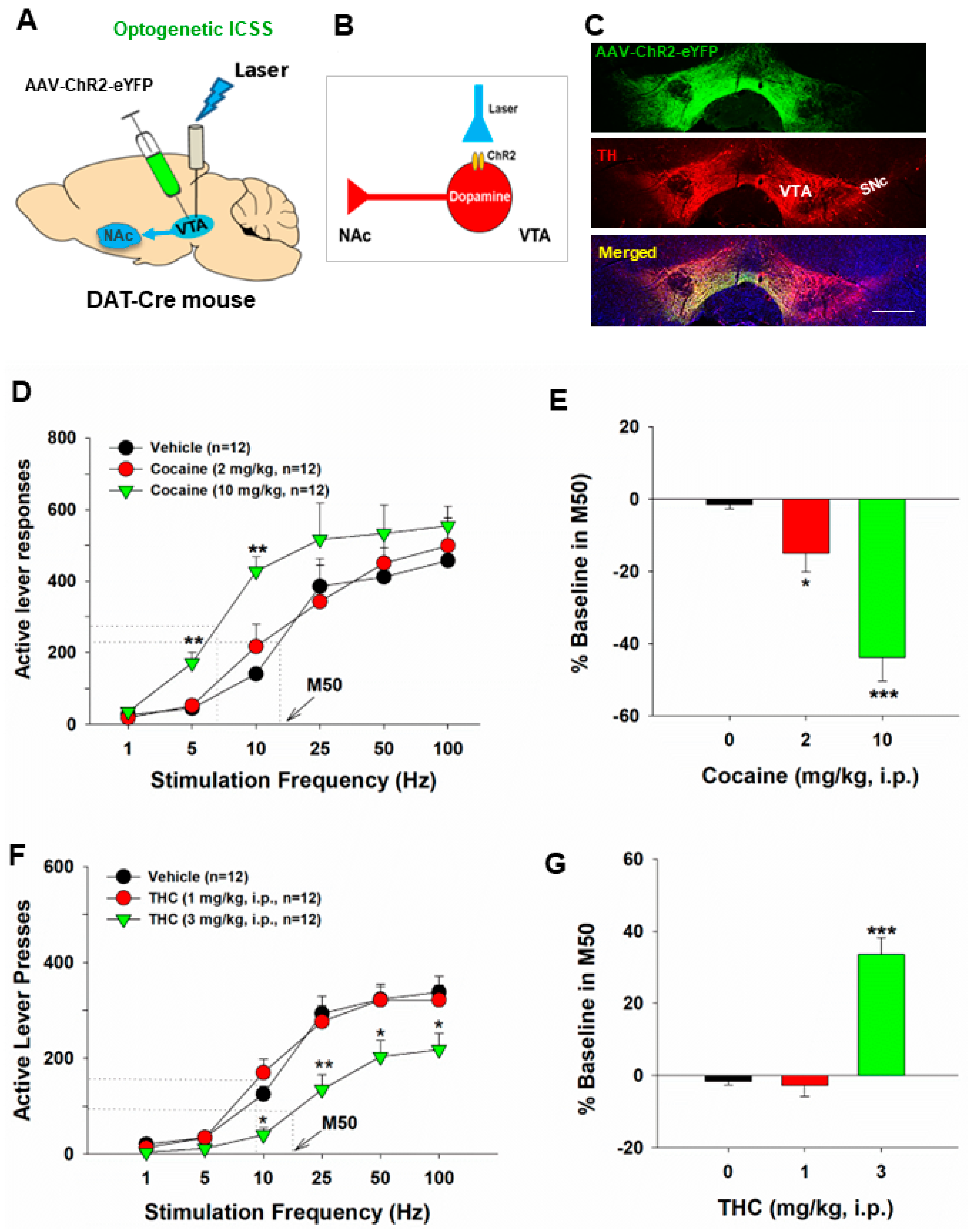
4. Glutamate-Dependent oICSS and Its Application in Studying Drug Reward and Addiction
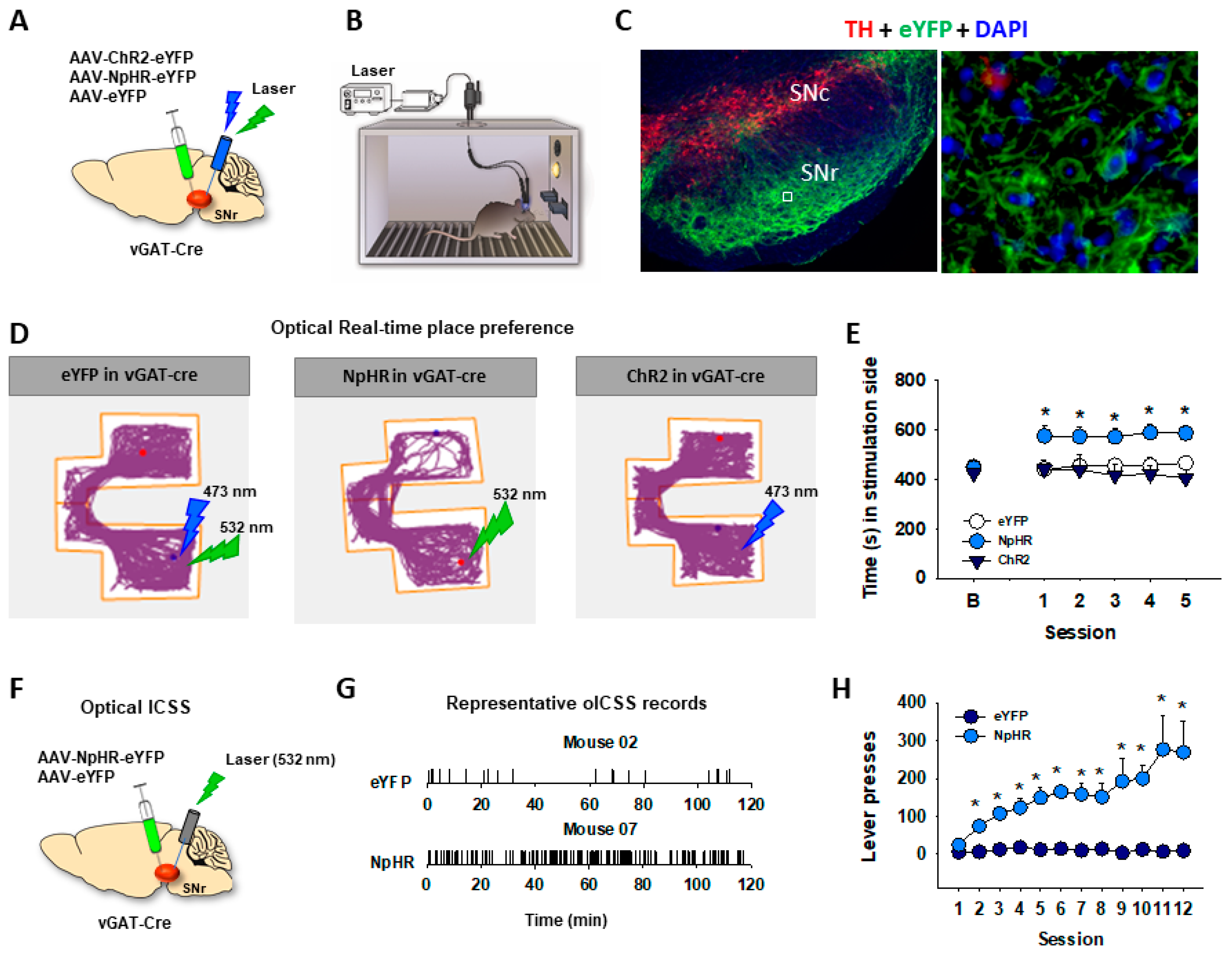
5. GABA-Dependent oICSS
6. Advantages and Limitations of oICSS
7. Potential Implications of oICSS in Human Studies
Funding
Conflicts of Interest
References
- Negus, S.S.; Miller, L.L. Intracranial self-stimulation to evaluate abuse potential of drugs. Pharmacol. Rev. 2014, 66, 869–917. [Google Scholar] [CrossRef] [PubMed]
- Gardner, E.L. Addiction and brain reward and antireward pathways. Adv. Psychosom. Med. 2011, 30, 22–60. [Google Scholar]
- Hernandez, G.; Haines, E.; Shizgal, P. Potentiation of intracranial self-stimulation during prolonged subcutaneous infusion of cocaine. J. Neurosci. Methods 2008, 175, 79–87. [Google Scholar] [CrossRef]
- Trujillo-Pisanty, I.; Conover, K.; Solis, P.; Palacios, D.; Shizgal, P. Dopamine neurons do not constitute an obligatory stage in the final common path for the evaluation and pursuit of brain stimulation reward. PLoS ONE 2020, 15, e0226722. [Google Scholar] [CrossRef]
- Olds, J.; Milner, P. Positive reinforcement produced by electrical stimulation of septal area and other regions of rat brain. J. Comp. Physiol. Psychol. 1954, 47, 419–427. [Google Scholar] [CrossRef]
- Wise, R.A. Addictive drugs and brain stimulation reward. Annu. Rev. Neurosci. 1996, 19, 319–340. [Google Scholar] [CrossRef]
- Der-Avakian, A.; Markou, A. The neurobiology of anhedonia and other reward-related deficits. Trends Neurosci. 2012, 35, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Carlezon, W.A., Jr.; Chartoff, E.H. Intracranial self-stimulation (ICSS) in rodents to study the neurobiology of motivation. Nat. Protoc. 2007, 2, 2987–2995. [Google Scholar] [CrossRef]
- Negus, S.S.; Moerke, M.J. Determinants of opioid abuse potential: Insights using intracranial self-stimulation. Peptides 2019, 112, 23–31. [Google Scholar] [CrossRef]
- Stuber, G.D.; Wise, R.A. Lateral hypothalamic circuits for feeding and reward. Nat. Neurosci. 2016, 19, 198–205. [Google Scholar] [CrossRef]
- Kornetsky, C.; Esposito, R.U.; McLean, S.; Jacobson, J.O. Intracranial self-stimulation thresholds: A model for the hedonic effects of drugs of abuse. Arch. Gen. Psychiatry 1979, 36, 289–292. [Google Scholar] [CrossRef]
- Vlachou, S.; Nomikos, G.G.; Panagis, G. CB1 cannabinoid receptor agonists increase intracranial self-stimulation thresholds in the rat. Psychopharmacology 2005, 179, 498–508. [Google Scholar] [CrossRef]
- Vlachou, S.; Nomikos, G.G.; Stephens, D.N.; Panagis, G. Lack of evidence for appetitive effects of Delta 9-tetrahydrocannabinol in the intracranial self-stimulation and conditioned place preference procedures in rodents. Behav. Pharmacol. 2007, 18, 311–319. [Google Scholar] [CrossRef]
- Han, X.; He, Y.; Bi, G.H.; Zhang, H.Y.; Song, R.; Liu, Q.R.; Egan, J.M.; Gardner, E.L.; Li, J.; Xi, Z.X. CB1 Receptor Activation on VgluT2-Expressing Glutamatergic Neurons Underlies Delta(9)-Tetrahydrocannabinol (Delta(9)-THC)-Induced Aversive Effects in Mice. Sci. Rep. 2017, 7, 12315. [Google Scholar] [CrossRef]
- Spiller, K.J.; Bi, G.H.; He, Y.; Galaj, E.; Gardner, E.L.; Xi, Z.X. Cannabinoid CB(1) and CB(2) receptor mechanisms underlie cannabis reward and aversion in rats. Br. J. Pharmacol. 2019, 176, 1268–1281. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.X.; Spiller, K.; Pak, A.C.; Gilbert, J.; Dillon, C.; Li, X.; Peng, X.Q.; Gardner, E.L. Cannabinoid CB1 receptor antagonists attenuate cocaine's rewarding effects: Experiments with self-administration and brain-stimulation reward in rats. Neuropsychopharmacology 2008, 33, 1735–1745. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Madeo, G.; Liang, Y.; Zhang, C.; Hempel, B.; Liu, X.; Mu, L.; Liu, S.; Bi, G.H.; Galaj, E.; et al. A red nucleus-VTA glutamate pathway underlies exercise reward and the therapeutic effect of exercise on cocaine use. Sci. Adv. 2022, 8, eabo1440. [Google Scholar] [CrossRef]
- Galaj, E.; Hempel, B.; Moore, A.; Klein, B.; Bi, G.H.; Gardner, E.L.; Seltzman, H.H.; Xi, Z.X. Therapeutic potential of PIMSR, a novel CB1 receptor neutral antagonist, for cocaine use disorder: Evidence from preclinical research. Transl. Psychiatry 2022, 12, 286. [Google Scholar] [CrossRef] [PubMed]
- Trujillo-Pisanty, I.; Conover, K.; Shizgal, P. A new view of the effect of dopamine receptor antagonism on operant performance for rewarding brain stimulation in the rat. Psychopharmacology 2013, 231, 1351–1364. [Google Scholar] [CrossRef]
- Breton, Y.A.; Conover, K.; Shizgal, P. The effect of probability discounting on reward seeking: A three-dimensional perspective. Front. Behav. Neurosci. 2014, 8, 284. [Google Scholar] [CrossRef]
- Hernandez, G.; Trujillo-Pisanty, I.; Cossette, M.P.; Conover, K.; Shizgal, P. Role of dopamine tone in the pursuit of brain stimulation reward. J. Neurosci. 2012, 32, 11032–11041. [Google Scholar] [CrossRef]
- Vlachou, S.; Markou, A. GABAB receptors in reward processes. Adv. Pharmacol. 2010, 58, 315–371. [Google Scholar]
- Stuber, G.D.; Britt, J.P.; Bonci, A. Optogenetic modulation of neural circuits that underlie reward seeking. Biol. Psychiatry 2012, 71, 1061–1067. [Google Scholar] [CrossRef]
- Ikemoto, S. Brain reward circuitry beyond the mesolimbic dopamine system: A neurobiological theory. Neurosci. Biobehav. Rev. 2010, 35, 129–150. [Google Scholar] [CrossRef] [PubMed]
- Yeomans, J.S.; Mathur, A.; Tampakeras, M. Rewarding brain stimulation: Role of tegmental cholinergic neurons that activate dopamine neurons. Behav. Neurosci. 1993, 107, 1077–1087. [Google Scholar] [CrossRef]
- Morales, M.; Margolis, E.B. Ventral tegmental area: Cellular heterogeneity, connectivity and behaviour. Nat. Rev. Neurosci. 2017, 18, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Boyden, E.S.; Zhang, F.; Bamberg, E.; Nagel, G.; Deisseroth, K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci. 2005, 8, 1263–1268. [Google Scholar] [CrossRef] [PubMed]
- Deisseroth, K. Optogenetics: 10 years of microbial opsins in neuroscience. Nat. Neurosci. 2015, 18, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Weidner, T.C.; Vincenz, D.; Brocka, M.; Tegtmeier, J.; Oelschlegel, A.M.; Ohl, F.W.; Goldschmidt, J.; Lippert, M.T. Matching stimulation paradigms resolve apparent differences between optogenetic and electrical VTA stimulation. Brain Stimul. 2020, 13, 363–371. [Google Scholar] [CrossRef]
- Qi, J.; Zhang, S.; Wang, H.L.; Wang, H.; de Jesus Aceves Buendia, J.; Hoffman, A.F.; Lupica, C.R.; Seal, R.P.; Morales, M. A glutamatergic reward input from the dorsal raphe to ventral tegmental area dopamine neurons. Nat. Commun. 2014, 5, 5390. [Google Scholar] [CrossRef]
- Tsai, H.C.; Zhang, F.; Adamantidis, A.; Stuber, G.D.; Bonci, A.; de Lecea, L.; Deisseroth, K. Phasic firing in dopaminergic neurons is sufficient for behavioral conditioning. Science 2009, 324, 1080–1084. [Google Scholar] [CrossRef] [PubMed]
- Adamantidis, A.R.; Tsai, H.C.; Boutrel, B.; Zhang, F.; Stuber, G.D.; Budygin, E.A.; Tourino, C.; Bonci, A.; Deisseroth, K.; de Lecea, L. Optogenetic interrogation of dopaminergic modulation of the multiple phases of reward-seeking behavior. J. Neurosci. 2011, 31, 10829–10835. [Google Scholar] [CrossRef]
- Gordon-Fennell, A.; Stuber, G.D. Illuminating subcortical GABAergic and glutamatergic circuits for reward and aversion. Neuropharmacology 2021, 198, 108725. [Google Scholar] [CrossRef]
- Root, D.H.; Barker, D.J.; Estrin, D.J.; Miranda-Barrientos, J.A.; Liu, B.; Zhang, S.; Wang, H.L.; Vautier, F.; Ramakrishnan, C.; Kim, Y.S.; et al. Distinct Signaling by Ventral Tegmental Area Glutamate, GABA, and Combinatorial Glutamate-GABA Neurons in Motivated Behavior. Cell Rep. 2020, 32, 108094. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Qi, J.; Zhang, S.; Wang, H.; Morales, M. Rewarding Effects of Optical Stimulation of Ventral Tegmental Area Glutamatergic Neurons. J. Neurosci. 2015, 35, 15948–15954. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.H.; Zell, V.; Gutierrez-Reed, N.; Wu, J.; Ressler, R.; Shenasa, M.A.; Johnson, A.B.; Fife, K.H.; Faget, L.; Hnasko, T.S. Ventral tegmental area glutamate neurons co-release GABA and promote positive reinforcement. Nat. Commun. 2016, 7, 13697. [Google Scholar] [CrossRef] [PubMed]
- Steidl, S.; Wang, H.; Ordonez, M.; Zhang, S.; Morales, M. Optogenetic excitation in the ventral tegmental area of glutamatergic or cholinergic inputs from the laterodorsal tegmental area drives reward. Eur. J. Neurosci. 2017, 45, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.X.; Kiyatkin, M.; Li, X.; Peng, X.Q.; Wiggins, A.; Spiller, K.; Li, J.; Gardner, E.L. N-acetylaspartylglutamate (NAAG) inhibits intravenous cocaine self-administration and cocaine-enhanced brain-stimulation reward in rats. Neuropharmacology 2010, 58, 304–313. [Google Scholar] [CrossRef]
- Vorel, S.R.; Ashby, C.R., Jr.; Paul, M.; Liu, X.; Hayes, R.; Hagan, J.J.; Middlemiss, D.N.; Stemp, G.; Gardner, E.L. Dopamine D3 receptor antagonism inhibits cocaine-seeking and cocaine-enhanced brain reward in rats. J. Neurosci. 2002, 22, 9595–9603. [Google Scholar] [CrossRef]
- Stoker, A.K.; Markou, A. Withdrawal from chronic cocaine administration induces deficits in brain reward function in C57BL/6J mice. Behav. Brain Res. 2011, 223, 176–181. [Google Scholar] [CrossRef]
- Kenny, P.J.; Markou, A. Neurobiology of the nicotine withdrawal syndrome. Pharmacol. Biochem. Behav. 2001, 70, 531–549. [Google Scholar] [CrossRef]
- Carter, F.; Cossette, M.P.; Trujillo-Pisanty, I.; Pallikaras, V.; Breton, Y.A.; Conover, K.; Caplan, J.; Solis, P.; Voisard, J.; Yaksich, A.; et al. Does phasic dopamine release cause policy updates? Eur. J. Neurosci. 2023. [Google Scholar] [CrossRef] [PubMed]
- Pallikaras, V.; Shizgal, P. Dopamine and Beyond: Implications of Psychophysical Studies of Intracranial Self-Stimulation for the Treatment of Depression. Brain Sci. 2022, 12, 1052. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.M.; Stuber, G.D. Optogenetic strategies to dissect the neural circuits that underlie reward and addiction. Cold Spring Harb. Perspect. Med. 2012, 2, a011924. [Google Scholar] [CrossRef] [PubMed]
- Bromberg-Martin, E.S.; Matsumoto, M.; Hikosaka, O. Dopamine in motivational control: Rewarding, aversive, and alerting. Neuron 2010, 68, 815–834. [Google Scholar] [CrossRef]
- Brischoux, F.; Chakraborty, S.; Brierley, D.I.; Ungless, M.A. Phasic excitation of dopamine neurons in ventral VTA by noxious stimuli. Proc. Natl. Acad. Sci. USA 2009, 106, 4894–4899. [Google Scholar] [CrossRef] [PubMed]
- Salamone, J.D.; Correa, M. The mysterious motivational functions of mesolimbic dopamine. Neuron 2012, 76, 470–485. [Google Scholar] [CrossRef]
- Witten, I.B.; Steinberg, E.E.; Lee, S.Y.; Davidson, T.J.; Zalocusky, K.A.; Brodsky, M.; Yizhar, O.; Cho, S.L.; Gong, S.; Ramakrishnan, C.; et al. Recombinase-driver rat lines: Tools, techniques, and optogenetic application to dopamine-mediated reinforcement. Neuron 2011, 72, 721–733. [Google Scholar] [CrossRef]
- Han, X.; Liang, Y.; Hempel, B.; Jordan, C.J.; Shen, H.; Bi, G.H.; Li, J.; Xi, Z.X. Cannabinoid CB1 Receptors Are Expressed in a Subset of Dopamine Neurons and Underlie Cannabinoid-Induced Aversion, Hypoactivity, and Anxiolytic Effects in Mice. J. Neurosci. 2023, 43, 373–385. [Google Scholar] [CrossRef]
- Humburg, B.A.; Jordan, C.J.; Zhang, H.Y.; Shen, H.; Han, X.; Bi, G.H.; Hempel, B.; Galaj, E.; Baumann, M.H.; Xi, Z.X. Optogenetic brain-stimulation reward: A new procedure to re-evaluate the rewarding versus aversive effects of cannabinoids in dopamine transporter-Cre mice. Addict. Biol. 2021, 26, e13005. [Google Scholar] [CrossRef]
- Galaj, E.; Han, X.; Shen, H.; Jordan, C.J.; He, Y.; Humburg, B.; Bi, G.H.; Xi, Z.X. Dissecting the Role of GABA Neurons in the VTA versus SNr in Opioid Reward. J. Neurosci. 2020, 40, 8853–8869. [Google Scholar] [CrossRef] [PubMed]
- Ilango, A.; Kesner, A.J.; Keller, K.L.; Stuber, G.D.; Bonci, A.; Ikemoto, S. Similar roles of substantia nigra and ventral tegmental dopamine neurons in reward and aversion. J. Neurosci. 2014, 34, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Heymann, G.; Jo, Y.S.; Reichard, K.L.; McFarland, N.; Chavkin, C.; Palmiter, R.D.; Soden, M.E.; Zweifel, L.S. Synergy of Distinct Dopamine Projection Populations in Behavioral Reinforcement. Neuron 2020, 105, 909–920 e5. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, E.E.; Boivin, J.R.; Saunders, B.T.; Witten, I.B.; Deisseroth, K.; Janak, P.H. Positive reinforcement mediated by midbrain dopamine neurons requires D1 and D2 receptor activation in the nucleus accumbens. PLoS ONE 2014, 9, e94771. [Google Scholar] [CrossRef]
- Ilango, A.; Kesner, A.J.; Broker, C.J.; Wang, D.V.; Ikemoto, S. Phasic excitation of ventral tegmental dopamine neurons potentiates the initiation of conditioned approach behavior: Parametric and reinforcement-schedule analyses. Front. Behav. Neurosci. 2014, 8, 155. [Google Scholar] [CrossRef]
- McDevitt, R.A.; Tiran-Cappello, A.; Shen, H.; Balderas, I.; Britt, J.P.; Marino, R.A.M.; Chung, S.L.; Richie, C.T.; Harvey, B.K.; Bonci, A. Serotonergic versus nonserotonergic dorsal raphe projection neurons: Differential participation in reward circuitry. Cell Rep. 2014, 8, 1857–1869. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Simmler, L.D.; Van Zessen, R.; Flakowski, J.; Wan, J.X.; Deng, F.; Li, Y.L.; Nautiyal, K.M.; Pascoli, V.; Luscher, C. Synaptic mechanism underlying serotonin modulation of transition to cocaine addiction. Science 2021, 373, 1252–1256. [Google Scholar] [CrossRef]
- Pascoli, V.; Terrier, J.; Hiver, A.; Luscher, C. Sufficiency of Mesolimbic Dopamine Neuron Stimulation for the Progression to Addiction. Neuron 2015, 88, 1054–1066. [Google Scholar] [CrossRef]
- Harada, M.; Pascoli, V.; Hiver, A.; Flakowski, J.; Luscher, C. Corticostriatal Activity Driving Compulsive Reward Seeking. Biol. Psychiatry 2021, 90, 808–818. [Google Scholar] [CrossRef]
- Corre, J.; van Zessen, R.; Loureiro, M.; Patriarchi, T.; Tian, L.; Pascoli, V.; Luscher, C. Dopamine neurons projecting to medial shell of the nucleus accumbens drive heroin reinforcement. eLife 2018, 7, e39945. [Google Scholar] [CrossRef]
- Pascoli, V.; Hiver, A.; Van Zessen, R.; Loureiro, M.; Achargui, R.; Harada, M.; Flakowski, J.; Luscher, C. Stochastic synaptic plasticity underlying compulsion in a model of addiction. Nature 2018, 564, 366–371. [Google Scholar] [CrossRef]
- Berrios, J.; Stamatakis, A.M.; Kantak, P.A.; McElligott, Z.A.; Judson, M.C.; Aita, M.; Rougie, M.; Stuber, G.D.; Philpot, B.D. Loss of UBE3A from TH-expressing neurons suppresses GABA co-release and enhances VTA-NAc optical self-stimulation. Nat. Commun. 2016, 7, 10702. [Google Scholar] [CrossRef]
- Rossi, M.A.; Sukharnikova, T.; Hayrapetyan, V.Y.; Yang, L.; Yin, H.H. Operant self-stimulation of dopamine neurons in the substantia nigra. PLoS ONE 2013, 8, e65799. [Google Scholar] [CrossRef]
- Han, X.; Jing, M.Y.; Zhao, T.Y.; Wu, N.; Song, R.; Li, J. Role of dopamine projections from ventral tegmental area to nucleus accumbens and medial prefrontal cortex in reinforcement behaviors assessed using optogenetic manipulation. Metab. Brain Dis. 2017, 32, 1491–1502. [Google Scholar] [CrossRef]
- Jing, M.Y.; Han, X.; Zhao, T.Y.; Wang, Z.Y.; Lu, G.Y.; Wu, N.; Song, R.; Li, J. Re-examining the role of ventral tegmental area dopaminergic neurons in motor activity and reinforcement by chemogenetic and optogenetic manipulation in mice. Metab. Brain Dis. 2019, 34, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Jing, M.Y.; Ding, X.Y.; Han, X.; Zhao, T.Y.; Luo, M.M.; Wu, N.; Li, J.; Song, R. Activation of mesocorticolimbic dopamine projections initiates cue-induced reinstatement of reward seeking in mice. Acta Pharmacol. Sin. 2022, 43, 2276–2288. [Google Scholar] [CrossRef] [PubMed]
- Gnazzo, F.G.; Mourra, D.; Guevara, C.A.; Beeler, J.A. Chronic food restriction enhances dopamine-mediated intracranial self-stimulation. Neuroreport 2021, 32, 1128–1133. [Google Scholar] [CrossRef]
- Stuber, G.D.; Sparta, D.R.; Stamatakis, A.M.; van Leeuwen, W.A.; Hardjoprajitno, J.E.; Cho, S.; Tye, K.M.; Kempadoo, K.A.; Zhang, F.; Deisseroth, K.; et al. Excitatory transmission from the amygdala to nucleus accumbens facilitates reward seeking. Nature 2011, 475, 377–380. [Google Scholar] [CrossRef]
- Britt, J.P.; Benaliouad, F.; McDevitt, R.A.; Stuber, G.D.; Wise, R.A.; Bonci, A. Synaptic and behavioral profile of multiple glutamatergic inputs to the nucleus accumbens. Neuron 2012, 76, 790–803. [Google Scholar] [CrossRef]
- Prado, L.; Luis-Islas, J.; Sandoval, O.I.; Puron, L.; Gil, M.M.; Luna, A.; Arias-Garcia, M.A.; Galarraga, E.; Simon, S.A.; Gutierrez, R. Activation of Glutamatergic Fibers in the Anterior NAc Shell Modulates Reward Activity in the aNAcSh, the Lateral Hypothalamus, and Medial Prefrontal Cortex and Transiently Stops Feeding. J. Neurosci. 2016, 36, 12511–12529. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.H.; Zell, V.; Wu, J.; Punta, C.; Ramajayam, N.; Shen, X.; Faget, L.; Lilascharoen, V.; Lim, B.K.; Hnasko, T.S. Activation of Pedunculopontine Glutamate Neurons Is Reinforcing. J. Neurosci. 2017, 37, 38–46. [Google Scholar] [CrossRef]
- Johnson, K.A.; Voyvodic, L.; Loewinger, G.C.; Mateo, Y.; Lovinger, D.M. Operant self-stimulation of thalamic terminals in the dorsomedial striatum is constrained by metabotropic glutamate receptor 2. Neuropsychopharmacology 2020, 45, 1454–1462. [Google Scholar] [CrossRef]
- Zell, V.; Steinkellner, T.; Hollon, N.G.; Warlow, S.M.; Souter, E.; Faget, L.; Hunker, A.C.; Jin, X.; Zweifel, L.S.; Hnasko, T.S. VTA Glutamate Neuron Activity Drives Positive Reinforcement Absent Dopamine Co-release. Neuron 2020, 107, 864–873.e4. [Google Scholar] [CrossRef]
- Han, W.; Tellez, L.A.; Perkins, M.H.; Perez, I.O.; Qu, T.; Ferreira, J.; Ferreira, T.L.; Quinn, D.; Liu, Z.W.; Gao, X.B.; et al. A Neural Circuit for Gut-Induced Reward. Cell 2018, 175, 887–888. [Google Scholar] [CrossRef]
- Lalive, A.L.; Lien, A.D.; Roseberry, T.K.; Donahue, C.H.; Kreitzer, A.C. Motor thalamus supports striatum-driven reinforcement. eLife 2018, 7, e34032. [Google Scholar] [CrossRef] [PubMed]
- Kravitz, A.V.; Tye, L.D.; Kreitzer, A.C. Distinct roles for direct and indirect pathway striatal neurons in reinforcement. Nat. Neurosci. 2012, 15, 816–818. [Google Scholar] [CrossRef]
- Vicente, A.M.; Galvao-Ferreira, P.; Tecuapetla, F.; Costa, R.M. Direct and indirect dorsolateral striatum pathways reinforce different action strategies. Curr. Biol. 2016, 26, R267–R269. [Google Scholar] [CrossRef]
- Cole, S.L.; Robinson, M.J.F.; Berridge, K.C. Optogenetic self-stimulation in the nucleus accumbens: D1 reward versus D2 ambivalence. PLoS ONE 2018, 13, e0207694. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhou, J.; Li, Y.; Hu, F.; Lu, Y.; Ma, M.; Feng, Q.; Zhang, J.E.; Wang, D.; Zeng, J.; et al. Dorsal raphe neurons signal reward through 5-HT and glutamate. Neuron 2014, 81, 1360–1374. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.W.; Morton, G.; Guy, E.G.; Wang, S.D.; Turner, E.E. Dorsal Medial Habenula Regulation of Mood-Related Behaviors and Primary Reinforcement by Tachykinin-Expressing Habenula Neurons. eNeuro 2016, 3. [Google Scholar] [CrossRef]
- Petter, E.A.; Fallon, I.P.; Hughes, R.N.; Watson, G.D.R.; Meck, W.H.; Ulloa Severino, F.P.; Yin, H.H. Elucidating a locus coeruleus-dentate gyrus dopamine pathway for operant reinforcement. eLife 2023, 12, e83600. [Google Scholar] [CrossRef] [PubMed]
- Hempel, B.; Crissman, M.; Pari, S.; Klein, B.; Bi, G.H.; Alton, H.; Xi, Z.X. PPARalpha and PPARgamma are expressed in midbrain dopamine neurons and modulate dopamine- and cannabinoid-mediated behavior in mice. Mol. Psychiatry 2023, 28, 4203–4214. [Google Scholar] [CrossRef]
- Jordan, C.J.; Feng, Z.W.; Galaj, E.; Bi, G.H.; Xue, Y.; Liang, Y.; McGuire, T.; Xie, X.Q.; Xi, Z.X. Xie2-64, a novel CB(2) receptor inverse agonist, reduces cocaine abuse-related behaviors in rodents. Neuropharmacology 2020, 176, 108241. [Google Scholar] [CrossRef]
- Gardner, E.L.; Paredes, W.; Smith, D.; Donner, A.; Milling, C.; Cohen, D.; Morrison, D. Facilitation of brain stimulation reward by delta 9-tetrahydrocannabinol. Psychopharmacology 1988, 96, 142–144. [Google Scholar] [CrossRef] [PubMed]
- Lepore, M.; Liu, X.; Savage, V.; Matalon, D.; Gardner, E.L. Genetic differences in delta 9-tetrahydrocannabinol-induced facilitation of brain stimulation reward as measured by a rate-frequency curve-shift electrical brain stimulation paradigm in three different rat strains. Life Sci. 1996, 58, PL365–PL372. [Google Scholar] [CrossRef] [PubMed]
- Katsidoni, V.; Kastellakis, A.; Panagis, G. Biphasic effects of Delta9-tetrahydrocannabinol on brain stimulation reward and motor activity. Int. J. Neuropsychopharmacol. 2013, 16, 2273–2284. [Google Scholar] [CrossRef] [PubMed]
- Kwilasz, A.J.; Negus, S.S. Dissociable effects of the cannabinoid receptor agonists Delta9-tetrahydrocannabinol and CP55940 on pain-stimulated versus pain-depressed behavior in rats. J. Pharmacol. Exp. Ther. 2012, 343, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Arnold, J.C.; Hunt, G.E.; McGregor, I.S. Effects of the cannabinoid receptor agonist CP 55,940 and the cannabinoid receptor antagonist SR 141716 on intracranial self-stimulation in Lewis rats. Life Sci. 2001, 70, 97–108. [Google Scholar] [CrossRef]
- Newman, A.H.; Cao, J.; Keighron, J.D.; Jordan, C.J.; Bi, G.H.; Liang, Y.; Abramyan, A.M.; Avelar, A.J.; Tschumi, C.W.; Beckstead, M.J.; et al. Translating the atypical dopamine uptake inhibitor hypothesis toward therapeutics for treatment of psychostimulant use disorders. Neuropsychopharmacology 2019, 44, 1435–1444. [Google Scholar] [CrossRef]
- Jordan, C.J.; Humburg, B.; Rice, M.; Bi, G.H.; You, Z.B.; Shaik, A.B.; Cao, J.; Bonifazi, A.; Gadiano, A.; Rais, R.; et al. The highly selective dopamine D(3)R antagonist, R-VK4-40 attenuates oxycodone reward and augments analgesia in rodents. Neuropharmacology 2019, 158, 107597. [Google Scholar] [CrossRef]
- Jordan, C.J.; He, Y.; Bi, G.H.; You, Z.B.; Cao, J.; Xi, Z.X.; Newman, A.H. (+/−)VK4-40, a novel dopamine D(3) receptor partial agonist, attenuates cocaine reward and relapse in rodents. Br. J. Pharmacol. 2020, 177, 4796–4807. [Google Scholar] [CrossRef] [PubMed]
- Newman, A.H.; Ku, T.; Jordan, C.J.; Bonifazi, A.; Xi, Z.X. New Drugs, Old Targets: Tweaking the Dopamine System to Treat Psychostimulant Use Disorders. Annu. Rev. Pharmacol. Toxicol. 2021, 61, 609–628. [Google Scholar] [CrossRef] [PubMed]
- Jordan, C.J.; Xi, Z.X. Progress in brain cannabinoid CB(2) receptor research: From genes to behavior. Neurosci. Biobehav. Rev. 2019, 98, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, F.; Garcia-Gutierrez, M.S.; Manzanares, J. Pharmacological regulation of cannabinoid CB2 receptor modulates the reinforcing and motivational actions of ethanol. Biochem. Pharmacol. 2018, 157, 227–234. [Google Scholar] [CrossRef]
- He, Y.; Galaj, E.; Bi, G.H.; Wang, X.F.; Gardner, E.; Xi, Z.X. beta-Caryophyllene, a dietary terpenoid, inhibits nicotine taking and nicotine seeking in rodents. Br. J. Pharmacol. 2020, 177, 2058–2072. [Google Scholar] [CrossRef] [PubMed]
- He, X.H.; Galaj, E.; Bi, G.H.; He, Y.; Hempel, B.; Wang, Y.L.; Gardner, E.L.; Xi, Z.X. beta-caryophyllene, an FDA-Approved Food Additive, Inhibits Methamphetamine-Taking and Methamphetamine-Seeking Behaviors Possibly via CB2 and Non-CB2 Receptor Mechanisms. Front. Pharmacol. 2021, 12, 722476. [Google Scholar] [CrossRef] [PubMed]
- Galaj, E.; Bi, G.H.; Moore, A.; Chen, K.; He, Y.; Gardner, E.; Xi, Z.X. Beta-caryophyllene inhibits cocaine addiction-related behavior by activation of PPARalpha and PPARgamma: Repurposing a FDA-approved food additive for cocaine use disorder. Neuropsychopharmacology 2021, 46, 860–870. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Y.; Gao, M.; Liu, Q.R.; Bi, G.H.; Li, X.; Yang, H.J.; Gardner, E.L.; Wu, J.; Xi, Z.X. Cannabinoid CB2 receptors modulate midbrain dopamine neuronal activity and dopamine-related behavior in mice. Proc. Natl. Acad. Sci. USA 2014, 111, E5007–E5015. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Gao, M.; Shen, H.; Bi, G.H.; Yang, H.J.; Liu, Q.R.; Wu, J.; Gardner, E.L.; Bonci, A.; Xi, Z.X. Expression of functional cannabinoid CB(2) receptor in VTA dopamine neurons in rats. Addict. Biol. 2017, 22, 752–765. [Google Scholar] [CrossRef]
- Cai, J.; Tong, Q. Anatomy and Function of Ventral Tegmental Area Glutamate Neurons. Front. Neural Circuits 2022, 16, 867053. [Google Scholar] [CrossRef]
- Yu, J.; Ishikawa, M.; Wang, J.; Schluter, O.M.; Sesack, S.R.; Dong, Y. Ventral Tegmental Area Projection Regulates Glutamatergic Transmission in Nucleus Accumbens. Sci. Rep. 2019, 9, 18451. [Google Scholar] [CrossRef]
- Barbano, M.F.; Wang, H.L.; Zhang, S.; Miranda-Barrientos, J.; Estrin, D.J.; Figueroa-Gonzalez, A.; Liu, B.; Barker, D.J.; Morales, M. VTA Glutamatergic Neurons Mediate Innate Defensive Behaviors. Neuron 2020, 107, 368–382.e8. [Google Scholar] [CrossRef]
- Takamori, S.; Rhee, J.S.; Rosenmund, C.; Jahn, R. Identification of a vesicular glutamate transporter that defines a glutamatergic phenotype in neurons. Nature 2000, 407, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Sheen, W.; Morales, M. Glutamatergic neurons are present in the rat ventral tegmental area. Eur. J. Neurosci. 2007, 25, 106–118. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Wang, H.L.; Li, X.; Ng, T.H.; Morales, M. Mesocorticolimbic glutamatergic pathway. J. Neurosci. 2011, 31, 8476–8490. [Google Scholar] [CrossRef]
- Stuber, G.D.; Hnasko, T.S.; Britt, J.P.; Edwards, R.H.; Bonci, A. Dopaminergic terminals in the nucleus accumbens but not the dorsal striatum corelease glutamate. J. Neurosci. 2010, 30, 8229–8233. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zhang, Y.; Kim, H.; Grayson, V.S.; Jovasevic, V.; Ren, W.; Centeno, M.V.; Guedea, A.L.; Meyer, M.A.A.; Wu, Y.; et al. Excitatory VTA to DH projections provide a valence signal to memory circuits. Nat. Commun. 2020, 11, 1466. [Google Scholar] [CrossRef] [PubMed]
- Lammel, S.; Lim, B.K.; Malenka, R.C. Reward and aversion in a heterogeneous midbrain dopamine system. Neuropharmacology 2014, 76 Pt B, 351–359. [Google Scholar] [CrossRef]
- Stopper, C.M.; Floresco, S.B. What's better for me? Fundamental role for lateral habenula in promoting subjective decision biases. Nat. Neurosci. 2014, 17, 33–35. [Google Scholar] [CrossRef]
- Root, D.H.; Mejias-Aponte, C.A.; Qi, J.; Morales, M. Role of glutamatergic projections from ventral tegmental area to lateral habenula in aversive conditioning. J. Neurosci. 2014, 34, 13906–13910. [Google Scholar] [CrossRef]
- Soler-Cedeno, O.; Xi, Z.X. Neutral CB1 Receptor Antagonists as Pharmacotherapies for Substance Use Disorders: Rationale, Evidence, and Challenge. Cells 2022, 11, 3262. [Google Scholar] [CrossRef]
- Fields, H.L.; Margolis, E.B. Understanding opioid reward. Trends Neurosci. 2015, 38, 217–225. [Google Scholar] [CrossRef]
- Matsui, A.; Jarvie, B.C.; Robinson, B.G.; Hentges, S.T.; Williams, J.T. Separate GABA afferents to dopamine neurons mediate acute action of opioids, development of tolerance, and expression of withdrawal. Neuron 2014, 82, 1346–1356. [Google Scholar] [CrossRef]
- McGovern, D.J.; Polter, A.M.; Prevost, E.D.; Ly, A.; McNulty, C.J.; Rubinstein, B.; Root, D.H. Ventral tegmental area glutamate neurons establish a mu-opioid receptor gated circuit to mesolimbic dopamine neurons and regulate opioid-seeking behavior. Neuropsychopharmacology 2023, 48, 1889–1900. [Google Scholar] [CrossRef]
- Reeves, K.C.; Kube, M.J.; Grecco, G.G.; Fritz, B.M.; Munoz, B.; Yin, F.; Gao, Y.; Haggerty, D.L.; Hoffman, H.J.; Atwood, B.K. Mu opioid receptors on vGluT2-expressing glutamatergic neurons modulate opioid reward. Addict. Biol. 2021, 26, e12942. [Google Scholar] [CrossRef]
- Jalabert, M.; Bourdy, R.; Courtin, J.; Veinante, P.; Manzoni, O.J.; Barrot, M.; Georges, F. Neuronal circuits underlying acute morphine action on dopamine neurons. Proc. Natl. Acad. Sci. USA 2011, 108, 16446–16450. [Google Scholar] [CrossRef]
- Chen, M.; Zhao, Y.; Yang, H.; Luan, W.; Song, J.; Cui, D.; Dong, Y.; Lai, B.; Ma, L.; Zheng, P. Morphine disinhibits glutamatergic input to VTA dopamine neurons and promotes dopamine neuron excitation. eLife 2015, 4, e09275. [Google Scholar] [CrossRef]
- van Zessen, R.; Phillips, J.L.; Budygin, E.A.; Stuber, G.D. Activation of VTA GABA neurons disrupts reward consumption. Neuron 2012, 73, 1184–1194. [Google Scholar] [CrossRef] [PubMed]
- Bouarab, C.; Thompson, B.; Polter, A.M. VTA GABA Neurons at the Interface of Stress and Reward. Front. Neural Circuits 2019, 13, 78. [Google Scholar] [CrossRef] [PubMed]
- Beier, K.T.; Gao, X.J.; Xie, S.; DeLoach, K.E.; Malenka, R.C.; Luo, L. Topological Organization of Ventral Tegmental Area Connectivity Revealed by Viral-Genetic Dissection of Input-Output Relations. Cell Rep. 2019, 26, 159–167 e6. [Google Scholar] [CrossRef] [PubMed]
- Creed, M.C.; Ntamati, N.R.; Tan, K.R. VTA GABA neurons modulate specific learning behaviors through the control of dopamine and cholinergic systems. Front. Behav. Neurosci. 2014, 8, 8. [Google Scholar] [CrossRef]
- Brown, M.T.; Tan, K.R.; O'Connor, E.C.; Nikonenko, I.; Muller, D.; Luscher, C. Ventral tegmental area GABA projections pause accumbal cholinergic interneurons to enhance associative learning. Nature 2012, 492, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Omelchenko, N.; Sesack, S.R. Ultrastructural analysis of local collaterals of rat ventral tegmental area neurons: GABA phenotype and synapses onto dopamine and GABA cells. Synapse 2009, 63, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Matsui, A.; Williams, J.T. Opioid-sensitive GABA inputs from rostromedial tegmental nucleus synapse onto midbrain dopamine neurons. J. Neurosci. 2011, 31, 17729–17735. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Ba, W.; Zhao, G.; Ma, Y.; Harding, E.C.; Yin, L.; Wang, D.; Li, H.; Zhang, P.; Shi, Y.; et al. Dysfunction of ventral tegmental area GABA neurons causes mania-like behavior. Mol. Psychiatry 2021, 26, 5213–5228. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Matsubara, T.; Miyazaki, T.; Ono, D.; Fukatsu, N.; Abe, M.; Sakimura, K.; Sudo, Y.; Yamanaka, A. GABA neurons in the ventral tegmental area regulate non-rapid eye movement sleep in mice. eLife 2019, 8, e44928. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.R.; Yvon, C.; Turiault, M.; Mirzabekov, J.J.; Doehner, J.; Labouebe, G.; Deisseroth, K.; Tye, K.M.; Luscher, C. GABA neurons of the VTA drive conditioned place aversion. Neuron 2012, 73, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Graveland, G.A.; DiFiglia, M. The frequency and distribution of medium-sized neurons with indented nuclei in the primate and rodent neostriatum. Brain Res. 1985, 327, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Surmeier, D.J.; Ding, J.; Day, M.; Wang, Z.; Shen, W. D1 and D2 dopamine-receptor modulation of striatal glutamatergic signaling in striatal medium spiny neurons. Trends Neurosci. 2007, 30, 228–235. [Google Scholar] [CrossRef]
- Gadziola, M.A.; Stetzik, L.A.; Wright, K.N.; Milton, A.J.; Arakawa, K.; Del Mar Cortijo, M.; Wesson, D.W. A Neural System that Represents the Association of Odors with Rewarded Outcomes and Promotes Behavioral Engagement. Cell Rep. 2020, 32, 107919. [Google Scholar] [CrossRef]
- Lippert, M.T.; Takagaki, K.; Weidner, T.; Brocka, M.; Tegtmeier, J.; Ohl, F.W. Chapter 17—Optogenetic Intracranial Self-Stimulation as a Method to Study the Plasticity-Inducing Effects of Dopamine. In Handbook of In Vivo Neural Plasticity Techniques; Academic Press: Cambridge, MA, USA, 2018; Volume 28, pp. 311–326. [Google Scholar]
- Tye, K.M.; Deisseroth, K. Optogenetic investigation of neural circuits underlying brain disease in animal models. Nat. Rev. Neurosci. 2012, 13, 251–266. [Google Scholar] [CrossRef]
- Carter, L.P.; Griffiths, R.R. Principles of laboratory assessment of drug abuse liability and implications for clinical development. Drug Alcohol Depend. 2009, 105 (Suppl. S1), S14–S25. [Google Scholar] [CrossRef]
- Horton, D.B.; Potter, D.M.; Mead, A.N. A translational pharmacology approach to understanding the predictive value of abuse potential assessments. Behav. Pharmacol. 2013, 24, 410–436. [Google Scholar] [CrossRef]
- Food and Drug Administration. Assessment of Abuse Potential of Drugs, Guidance for Industry. January 2017. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/assessment-abuse-potential-drugs (accessed on 16 March 2024).

| Tested Drugs | Mice | oICSS | Major Findings | References |
|---|---|---|---|---|
Drugs of abuse | ||||
| Cocaine (5, 10, 15, 20 mg/kg) | DAT-Cre | 20 Hz | ↓ oICSS by 20 Hz laser | [58] |
| Heroin (1, 2, 4, 8, 16, 32 mg/kg) | DAT-Cre | 20 Hz | ↓ oICSS by 20 Hz laser | [60] |
| Cocaine (2, 10 mg/kg) | DAT-Cre | F-R curve | ↑ oICSS, Shift the F-R curve to the left | [83,91] |
| Oxycodone (0.3, 1, 3 mg/kg) | DAT-Cre | F-R curve | ↑ oICSS at low doses, ↓ oICSS at high doses, Shift the F-R curve upward or downward | [90] |
DAT inhibitors | ||||
| JJC8-088 (DAT inhibitor) | DAT-Cre | F-R curve | ↑ oICSS, Shift the F-R curve upward | [89] |
| JJC8-091 (Atypical DAT inhibitor) | DAT-Cre | F-R curve | ↓ oICSS, Shift the F-R curve downward | [89] |
Dopamine D3 receptor ligands | ||||
| (±)-VK4-40 (D3 antagonist) | DAT-Cre | F-R curve | ↓ oICSS, Shift the F-R curve downward | [91] |
| R-VK4-40 (D3 antagonist) | DAT-Cre | F-R curve | ↓ oICSS, Shift the F-R curve downward | [90] |
| S-VK4-40 (D3 partial agonist) | DAT-Cre | F-R curve | ↓ oICSS, Shift the F-R curve downward | [92] |
Cannabinoid receptor agonists | ||||
| THC | DAT-Cre | F-R curve | ↓ oICSS, Shift the F-R curve downward | [49] |
| THC | VgluT2-Cre | F-R curve | ↓ oICSS, Shift the F-R curve downward | [50] |
| WIN55,212-2 | DAT-Cre | F-R curve | ↓ oICSS, Shift the F-R curve downward | [50] |
| AM-2201 | DAT-Cre | F-R curve | ↓ oICSS, Shift the F-R curve downward | [50] |
| CBD | DAT-Cre | F-R curve | No effect on oICSS | [50] |
| ACEA (CB1 agonist) | DAT-Cre | F-R curve | ↓ oICSS, Shift the F-R curve downward | [50] |
| JWH133 (CB2 agonist) | DAT-Cre | F-R curve | ↓ oICSS, Shift the F-R curve downward | [49] |
| Xie2-64 (CB2 inverse agonist) | DAT-Cre | F-R curve | ↓ oICSS, Shift the F-R curve downward | [83] |
| BCP (CB2 agonist) | DAT-Cre | F-R curve | ↓ oICSS, Shift the F-R curve downward | [95,96,97] |
| GW7647 (PPARa agonist) | DAT-Cre | F-R curve | No effect | [82] |
| Pioglitazone (PPARg agonist) | DAT-Cre | F-R curve | ↓ oICSS, Shift the F-R curve downward | [82] |
Cannabinoid receptor antagonists | ||||
| PIMSR (Neutral CB1 antagonist) | DAT-Cre | F-R curve | ↓ oICSS, Shift the F-R curve downward | [18] |
| SR144528 (CB2 antagonist) | DAT-Cre | F-R curve | No effect on oICSS | [83] |
| GW6471 (PPARa antagonist) | DAT-Cre | F-R curve | ↓ oICSS, Shift the F-R curve downward | [82] |
| GW9662 (PPARg antagonist) | DAT-Cre | F-R curve | No effect on oICSS | [82] |
| Self-Administration | CPP/CPA | ICSS | |
|---|---|---|---|
| Reward assessment | Measures the reinforcing properties of drugs | Directly measures reward-associative learning Indirectly measures drug reward | Measures the rewarding properties of direct brain stimulation |
| Advantages | Mimics human drug-taking Measures motivation for drugs Allows for the assessment of drug intake patterns over time | Non-invasive Easy to implement | Brain region or cell type specificity Isolates reward circuits Provides real-time measurement of reward-related behavior |
| Disadvantages | Invasive procedure Does not provide information regarding brain regions or neural circuitry involved in drug reward | Does not provide information regarding brain regions or neural circuitry involved in this behavior | Invasive procedure Surgery, AAV, and electrical stimulation may cause brain tissue damage Technically more complicated |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, R.; Soler-Cedeño, O.; Xi, Z.-X. Optical Intracranial Self-Stimulation (oICSS): A New Behavioral Model for Studying Drug Reward and Aversion in Rodents. Int. J. Mol. Sci. 2024, 25, 3455. https://doi.org/10.3390/ijms25063455
Song R, Soler-Cedeño O, Xi Z-X. Optical Intracranial Self-Stimulation (oICSS): A New Behavioral Model for Studying Drug Reward and Aversion in Rodents. International Journal of Molecular Sciences. 2024; 25(6):3455. https://doi.org/10.3390/ijms25063455
Chicago/Turabian StyleSong, Rui, Omar Soler-Cedeño, and Zheng-Xiong Xi. 2024. "Optical Intracranial Self-Stimulation (oICSS): A New Behavioral Model for Studying Drug Reward and Aversion in Rodents" International Journal of Molecular Sciences 25, no. 6: 3455. https://doi.org/10.3390/ijms25063455
APA StyleSong, R., Soler-Cedeño, O., & Xi, Z.-X. (2024). Optical Intracranial Self-Stimulation (oICSS): A New Behavioral Model for Studying Drug Reward and Aversion in Rodents. International Journal of Molecular Sciences, 25(6), 3455. https://doi.org/10.3390/ijms25063455