
Evolution and Function of the Notch Signaling Pathway: An Invertebrate Perspective


 , and
, and
Abstract
1. Introduction
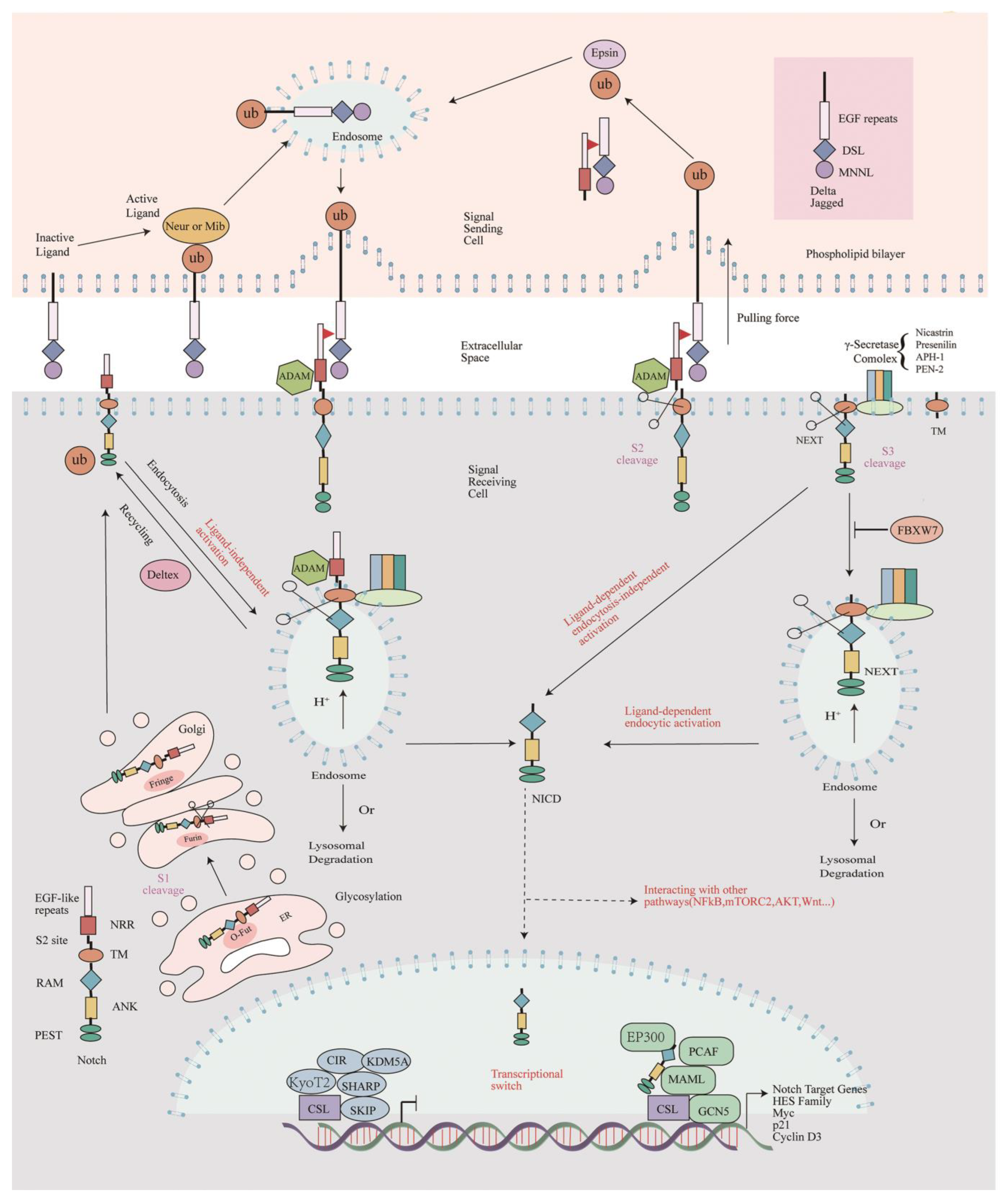
2. The Architecture of the Canonical Notch Signaling Pathway
2.1. Notch Receptor Processing and Signal Transduction
2.2. The Notch Cascade Is Negatively Affected by γ-Secretase Inhibitor
2.3. The Auxiliary Proteins That Affect Notch Signaling
2.3.1. Corepressors and Coactivators of Rbpj
2.3.2. Post-Translational Modifications
3. Functional Roles of Notch Signaling Pathway Members in Invertebrates
3.1. Choanozoa: Monosiga brevicollis (M. brevicollis)
3.2. Porifera/Spongia
3.3. Cnidaria
3.3.1. Hydrozoa
3.3.2. Anthozoa: N. vectensis
3.4. Platyhelminthes
3.4.1. Turbellaria: Dugesia japonica (D. japonica) and Schmidtea mediterranea (S. mediterranea)
3.4.2. Trematoda: Schistosoma mansoni (S. mansoni)
3.4.3. Cestoda: Echinococcus granulosus (E. granulosus)
3.5. Nemathelminthes, Nematoda: Caenorhabdits elegans (C. elegans)
3.6. Annelida
3.6.1. Earthworms: Perionyx excavates (P. excavatus)
3.6.2. Polychaeta: Platynereis dumerilii (P. dumerilii)
3.6.3. Clitellata: Helobdella robusta (H. robusta)
3.7. Mollusca
3.7.1. Gastropoda: Ilyanassa obsoleta (I. obsoleta)
3.7.2. Bivalvia (Pelecypoda): Crassostrea gigas (C. gigas)
3.7.3. Cephalopoda
3.8. Arthropoda
3.8.1. Uniramia: D. melanogaster, T. castaneum, and S. maritima
3.8.2. Chelicerata: Cupiennius salei (C. salei)
3.8.3. Crustacea: Penaeus vannamei (P. vannamei) and Litopenaeus vannamei (L. vannamei)
3.9. Echinodermata
3.9.1. Echinoidea/Sea Urchin
3.9.2. Asteroidea/Starfish: Patiria miniata (P. miniata)
3.9.3. Ophiuroidea/Brittle Stars
3.9.4. Holothuroidea/Sea Cucumbers
3.10. Chordata
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Mohr, O.L. Character Changes Caused by Mutation of an Entire Region of a Chromosome in Drosophila. Genetics 1919, 4, 275–282. [Google Scholar] [CrossRef]
- Metz, C.W.; Bridges, C.B. Incompatibility of Mutant Races in Drosophila. Proc. Natl. Acad. Sci. USA 1917, 3, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, H.; Yi, T.C.; Shen, J.; Zhang, J. Notch Signaling in Insect Development: A Simple Pathway with Diverse Functions. Int. J. Mol. Sci. 2023, 24, 14028. [Google Scholar] [CrossRef] [PubMed]
- Kopan, R.; Ilagan, M.X. The canonical Notch signaling pathway: Unfolding the activation mechanism. Cell 2009, 137, 216–233. [Google Scholar] [CrossRef] [PubMed]
- Nurmahdi, H.; Hasegawa, M.; Mujizah, E.Y.; Sasamura, T.; Inaki, M.; Yamamoto, S.; Yamakawa, T.; Matsuno, K. Notch Missense Mutations in Drosophila Reveal Functions of Specific EGF-like Repeats in Notch Folding, Trafficking, and Signaling. Biomolecules 2022, 12, 1752. [Google Scholar] [CrossRef]
- Zhou, B.; Lin, W.; Long, Y.; Yang, Y.; Zhang, H.; Wu, K.; Chu, Q. Notch signaling pathway: Architecture, disease, and therapeutics. Signal Transduct. Target. Ther. 2022, 7, 95. [Google Scholar] [CrossRef]
- Nandagopal, N.; Santat, L.A.; Elowitz, M.B. Cis-activation in the Notch signaling pathway. eLife 2019, 8, e37880. [Google Scholar] [CrossRef]
- Mumm, J.S.; Schroeter, E.H.; Saxena, M.T.; Griesemer, A.; Tian, X.; Pan, D.J.; Ray, W.J.; Kopan, R. A ligand-induced extracellular cleavage regulates gamma-secretase-like proteolytic activation of Notch1. Mol. Cell 2000, 5, 197–206. [Google Scholar] [CrossRef]
- Brou, C.; Logeat, F.; Gupta, N.; Bessia, C.; LeBail, O.; Doedens, J.R.; Cumano, A.; Roux, P.; Black, R.A.; Israel, A. A novel proteolytic cleavage involved in Notch signaling: The role of the disintegrin-metalloprotease TACE. Mol. Cell 2000, 5, 207–216. [Google Scholar] [CrossRef]
- Gazave, E.; Lapebie, P.; Richards, G.S.; Brunet, F.; Ereskovsky, A.V.; Degnan, B.M.; Borchiellini, C.; Vervoort, M.; Renard, E. Origin and evolution of the Notch signalling pathway: An overview from eukaryotic genomes. BMC Evol. Biol. 2009, 9, 249. [Google Scholar] [CrossRef]
- Murata, A.; Hayashi, S. Notch-Mediated Cell Adhesion. Biology 2016, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Gordon, W.R.; Zimmerman, B.; He, L.; Miles, L.J.; Huang, J.; Tiyanont, K.; McArthur, D.G.; Aster, J.C.; Perrimon, N.; Loparo, J.J.; et al. Mechanical Allostery: Evidence for a Force Requirement in the Proteolytic Activation of Notch. Dev. Cell 2015, 33, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Hsia, H.E.; Tushaus, J.; Brummer, T.; Zheng, Y.; Scilabra, S.D.; Lichtenthaler, S.F. Functions of ‘A disintegrin and metalloproteases (ADAMs)’ in the mammalian nervous system. Cell. Mol. Life Sci. 2019, 76, 3055–3081. [Google Scholar] [CrossRef] [PubMed]
- Canalis, E.; Zanotti, S. Hairy and Enhancer of Split-Related with YRPW Motif-Like (HeyL) Is Dispensable for Bone Remodeling in Mice. J. Cell. Biochem. 2017, 118, 1819–1826. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Lamarca, M.J.; Falo-Sanjuan, J.; Stojnic, R.; Abdul Rehman, S.; Muresan, L.; Jones, M.L.; Pillidge, Z.; Cerda-Moya, G.; Yuan, Z.; Baloul, S.; et al. Activation of the Notch Signaling Pathway In Vivo Elicits Changes in CSL Nuclear Dynamics. Dev. Cell 2018, 44, 611–623.e7. [Google Scholar] [CrossRef] [PubMed]
- Geling, A.; Steiner, H.; Willem, M.; Bally-Cuif, L.; Haass, C. A gamma-secretase inhibitor blocks Notch signaling in vivo and causes a severe neurogenic phenotype in zebrafish. EMBO Rep. 2002, 3, 688–694. [Google Scholar] [CrossRef]
- Gertsik, N.; Chiu, D.; Li, Y.M. Complex regulation of gamma-secretase: From obligatory to modulatory subunits. Front. Aging Neurosci. 2014, 6, 342. [Google Scholar] [CrossRef]
- Wolfe, M.S. Structure and Function of the gamma-Secretase Complex. Biochemistry 2019, 58, 2953–2966. [Google Scholar] [CrossRef]
- Brown, M.S.; Ye, J.; Rawson, R.B.; Goldstein, J.L. Regulated intramembrane proteolysis: A control mechanism conserved from bacteria to humans. Cell 2000, 100, 391–398. [Google Scholar] [CrossRef]
- Liefke, R.; Oswald, F.; Alvarado, C.; Ferres-Marco, D.; Mittler, G.; Rodriguez, P.; Dominguez, M.; Borggrefe, T. Histone demethylase KDM5A is an integral part of the core Notch-RBP-J repressor complex. Genes Dev. 2010, 24, 590–601. [Google Scholar] [CrossRef]
- Wang, H.; Zang, C.; Liu, X.S.; Aster, J.C. The role of Notch receptors in transcriptional regulation. J. Cell. Physiol. 2015, 230, 982–988. [Google Scholar] [CrossRef] [PubMed]
- Kurooka, H.; Honjo, T. Functional interaction between the mouse notch1 intracellular region and histone acetyltransferases PCAF and GCN5. J. Biol. Chem. 2000, 275, 17211–17220. [Google Scholar] [CrossRef] [PubMed]
- Oswald, F.; Tauber, B.; Dobner, T.; Bourteele, S.; Kostezka, U.; Adler, G.; Liptay, S.; Schmid, R.M. p300 acts as a transcriptional coactivator for mammalian Notch-1. Mol. Cell. Biol. 2001, 21, 7761–7774. [Google Scholar] [CrossRef]
- Coyle-Thompson, C.A.; Banerjee, U. The strawberry notch gene functions with Notch in common developmental pathways. Development 1993, 119, 377–395. [Google Scholar] [CrossRef] [PubMed]
- Antfolk, D.; Antila, C.; Kemppainen, K.; Landor, S.K.; Sahlgren, C. Decoding the PTM-switchboard of Notch. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 118507. [Google Scholar] [CrossRef]
- Okajima, T.; Matsuda, T. Roles of O-fucosyltransferase 1 and O-linked fucose in notch receptor function. Methods Enzymol. 2006, 417, 111–126. [Google Scholar] [CrossRef]
- Shi, S.; Stanley, P. Protein O-fucosyltransferase 1 is an essential component of Notch signaling pathways. Proc. Natl. Acad. Sci. USA 2003, 100, 5234–5239. [Google Scholar] [CrossRef]
- Saiki, W.; Ma, C.; Okajima, T.; Takeuchi, H. Current Views on the Roles of O-Glycosylation in Controlling Notch-Ligand Interactions. Biomolecules 2021, 11, 309. [Google Scholar] [CrossRef]
- Ferrante, F.; Giaimo, B.D.; Friedrich, T.; Sugino, T.; Mertens, D.; Kugler, S.; Gahr, B.M.; Just, S.; Pan, L.; Bartkuhn, M.; et al. Hydroxylation of the NOTCH1 intracellular domain regulates Notch signaling dynamics. Cell Death Dis. 2022, 13, 600. [Google Scholar] [CrossRef]
- Carrieri, F.A.; Murray, P.J.; Ditsova, D.; Ferris, M.A.; Davies, P.; Dale, J.K. CDK1 and CDK2 regulate NICD1 turnover and the periodicity of the segmentation clock. EMBO Rep. 2019, 20, e46436. [Google Scholar] [CrossRef]
- Koo, B.K.; Yoon, K.J.; Yoo, K.W.; Lim, H.S.; Song, R.; So, J.H.; Kim, C.H.; Kong, Y.Y. Mind bomb-2 is an E3 ligase for Notch ligand. J. Biol. Chem. 2005, 280, 22335–22342. [Google Scholar] [CrossRef] [PubMed]
- Stunkel, W.; Campbell, R.M. Sirtuin 1 (SIRT1): The misunderstood HDAC. J. Biomol. Screen 2011, 16, 1153–1169. [Google Scholar] [CrossRef] [PubMed]
- Hein, K.; Mittler, G.; Cizelsky, W.; Kuhl, M.; Ferrante, F.; Liefke, R.; Berger, I.M.; Just, S.; Strang, J.E.; Kestler, H.A.; et al. Site-specific methylation of Notch1 controls the amplitude and duration of the Notch1 response. Sci. Signal. 2015, 8, ra30. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Xiao, W.; Chen, W.; Liu, X.; Wu, M.; Bo, Q.; Luo, Y.; Ye, S.; Cao, Y.; Liu, Y. MicroRNA-26a and -26b inhibit lens fibrosis and cataract by negatively regulating Jagged-1/Notch signaling pathway. Cell Death Differ. 2017, 24, 1431–1442. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.; Sharma, V.; Mutsuddi, M.; Mukherjee, A. Regulation of Notch signaling by E3 ubiquitin ligases. FEBS J. 2022, 289, 937–954. [Google Scholar] [CrossRef]
- Schweisguth, F. Regulation of notch signaling activity. Curr. Biol. 2004, 14, R129–R138. [Google Scholar] [CrossRef]
- Yamamoto, S.; Schulze, K.L.; Bellen, H.J. Introduction to Notch signaling. In Notch Signaling; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2014; Volume 1187, pp. 1–14. [Google Scholar] [CrossRef]
- Hartley, D.A.; Xu, T.A.; Artavanis-Tsakonas, S. The embryonic expression of the Notch locus of Drosophila melanogaster and the implications of point mutations in the extracellular EGF-like domain of the predicted protein. EMBO J. 1987, 6, 3407–3417. [Google Scholar] [CrossRef]
- Schiwitza, S.; Spruck, C.; Nitsche, F. Biogeographical distribution of Hartaetosiga strains (Choanoflagellatea, Craspedida, Salpingoecidae) including morphological and transcriptomic data from a transect across the Atlantic Ocean. J. Eukaryot. Microbiol. 2023, 70, e12933. [Google Scholar] [CrossRef]
- Schenkelaars, Q.; Pratlong, M.; Kodjabachian, L.; Fierro-Constain, L.; Vacelet, J.; Le Bivic, A.; Renard, E.; Borchiellini, C. Animal multicellularity and polarity without Wnt signaling. Sci. Rep. 2017, 7, 15383. [Google Scholar] [CrossRef]
- Richards, G.S.; Degnan, B.M. The expression of Delta ligands in the sponge Amphimedon queenslandica suggests an ancient role for Notch signaling in metazoan development. EvoDevo 2012, 3, 15. [Google Scholar] [CrossRef]
- Simionato, E.; Ledent, V.; Richards, G.; Thomas-Chollier, M.; Kerner, P.; Coornaert, D.; Degnan, B.M.; Vervoort, M. Origin and diversification of the basic helix-loop-helix gene family in metazoans: Insights from comparative genomics. BMC Evol. Biol. 2007, 7, 33. [Google Scholar] [CrossRef]
- Prexl, A.; Munder, S.; Loy, B.; Kremmer, E.; Tischer, S.; Bottger, A. The putative Notch ligand HyJagged is a transmembrane protein present in all cell types of adult Hydra and upregulated at the boundary between bud and parent. BMC Cell Biol. 2011, 12, 38. [Google Scholar] [CrossRef] [PubMed]
- Munder, S.; Tischer, S.; Grundhuber, M.; Buchels, N.; Bruckmeier, N.; Eckert, S.; Seefeldt, C.A.; Prexl, A.; Kasbauer, T.; Bottger, A. Notch-signalling is required for head regeneration and tentacle patterning in Hydra. Dev. Biol. 2013, 383, 146–157. [Google Scholar] [CrossRef]
- Gahan, J.M.; Schnitzler, C.E.; DuBuc, T.Q.; Doonan, L.B.; Kanska, J.; Gornik, S.G.; Barreira, S.; Thompson, K.; Schiffer, P.; Baxevanis, A.D.; et al. Functional studies on the role of Notch signaling in Hydractinia development. Dev. Biol. 2017, 428, 224–231. [Google Scholar] [CrossRef]
- Marlow, H.; Roettinger, E.; Boekhout, M.; Martindale, M.Q. Functional roles of Notch signaling in the cnidarian Nematostella vectensis. Dev. Biol. 2012, 362, 295–308. [Google Scholar] [CrossRef]
- Layden, M.J.; Martindale, M.Q. Non-canonical Notch signaling represents an ancestral mechanism to regulate neural differentiation. EvoDevo 2014, 5, 30. [Google Scholar] [CrossRef]
- DuBuc, T.Q.; Traylor-Knowles, N.; Martindale, M.Q. Initiating a regenerative response; cellular and molecular features of wound healing in the cnidarian Nematostella vectensis. BMC Biol. 2014, 12, 24. [Google Scholar] [CrossRef]
- Hemond, E.M.; Kaluziak, S.T.; Vollmer, S.V. The genetics of colony form and function in Caribbean Acropora corals. BMC Genom. 2014, 15, 1133. [Google Scholar] [CrossRef]
- Anderson, D.A.; Walz, M.E.; Weil, E.; Tonellato, P.; Smith, M.C. RNA-Seq of the Caribbean reef-building coral Orbicella faveolata (Scleractinia-Merulinidae) under bleaching and disease stress expands models of coral innate immunity. PeerJ 2016, 4, e1616. [Google Scholar] [CrossRef]
- Dong, Z.; Huo, J.; Liang, A.; Chen, J.; Chen, G.; Liu, D. Gamma-Secretase Inhibitor (DAPT), a potential therapeutic target drug, caused neurotoxicity in planarian regeneration by inhibiting Notch signaling pathway. Sci. Total Environ. 2021, 781, 146735. [Google Scholar] [CrossRef]
- Cowles, M.W.; Brown, D.D.; Nisperos, S.V.; Stanley, B.N.; Pearson, B.J.; Zayas, R.M. Genome-wide analysis of the bHLH gene family in planarians identifies factors required for adult neurogenesis and neuronal regeneration. Development 2013, 140, 4691–4702. [Google Scholar] [CrossRef]
- Sasidharan, V.; Marepally, S.; Elliott, S.A.; Baid, S.; Lakshmanan, V.; Nayyar, N.; Bansal, D.; Sanchez Alvarado, A.; Vemula, P.K.; Palakodeti, D. The miR-124 family of microRNAs is crucial for regeneration of the brain and visual system in the planarian Schmidtea mediterranea. Development 2017, 144, 3211–3223. [Google Scholar] [CrossRef]
- Magalhaes, L.G.; Morais, E.R.; Machado, C.B.; Gomes, M.S.; Cabral, F.J.; Souza, J.M.; Soares, C.S.; Sa, R.G.; Castro-Borges, W.; Rodrigues, V. Uncovering Notch pathway in the parasitic flatworm Schistosoma mansoni. Parasitol. Res. 2016, 115, 3951–3961. [Google Scholar] [CrossRef]
- Zheng, H.; Zhang, W.; Zhang, L.; Zhang, Z.; Li, J.; Lu, G.; Zhu, Y.; Wang, Y.; Huang, Y.; Liu, J.; et al. The genome of the hydatid tapeworm Echinococcus granulosus. Nat. Genet. 2013, 45, 1168–1175. [Google Scholar] [CrossRef]
- Dezaki, E.S.; Yaghoobi, M.M.; Taheri, E.; Almani, P.G.; Tohidi, F.; Gottstein, B.; Harandi, M.F. Differential Expression of Hox and Notch Genes in Larval and Adult Stages of Echinococcus granulosus. Korean J. Parasitol. 2016, 54, 653–658. [Google Scholar] [CrossRef]
- Liu, S.; Zhou, X.; Hao, L.; Piao, X.; Hou, N.; Chen, Q. Genome-Wide Transcriptome Analysis Reveals Extensive Alternative Splicing Events in the Protoscoleces of Echinococcus granulosus and Echinococcus multilocularis. Front. Microbiol. 2017, 8, 929. [Google Scholar] [CrossRef]
- Gao, J.; Zhou, X.; Liu, L.; Lv, G.; Hou, Q.; Zhang, X.; Shen, Y. Bioinformatics analysis and experimental verification of Notch signalling pathway-related miRNA-mRNA subnetwork in extracellular vesicles during Echinococcus granulosus encystation. Parasit. Vectors 2022, 15, 272. [Google Scholar] [CrossRef]
- Tax, F.E.; Thomas, J.H. Cell-cell interactions. Receiving signals in the nematode embryo. Curr. Biol. 1994, 4, 914–916. [Google Scholar] [CrossRef] [PubMed]
- Mango, S.E.; Thorpe, C.J.; Martin, P.R.; Chamberlain, S.H.; Bowerman, B. Two maternal genes, apx-1 and pie-1, are required to distinguish the fates of equivalent blastomeres in the early Caenorhabditis elegans embryo. Development 1994, 120, 2305–2315. [Google Scholar] [CrossRef]
- Mello, C.C.; Draper, B.W.; Priess, J.R. The maternal genes apx-1 and glp-1 and establishment of dorsal-ventral polarity in the early C. elegans embryo. Cell 1994, 77, 95–106. [Google Scholar] [CrossRef]
- Hoyos, E.; Kim, K.; Milloz, J.; Barkoulas, M.; Penigault, J.B.; Munro, E.; Felix, M.A. Quantitative variation in autocrine signaling and pathway crosstalk in the Caenorhabditis vulval network. Curr. Biol. 2011, 21, 527–538. [Google Scholar] [CrossRef]
- Fitzgerald, K.; Greenwald, I. Interchangeability of Caenorhabditis elegans DSL proteins and intrinsic signalling activity of their extracellular domains in vivo. Development 1995, 121, 4275–4282. [Google Scholar] [CrossRef]
- Wilkinson, H.A.; Fitzgerald, K.; Greenwald, I. Reciprocal changes in expression of the receptor lin-12 and its ligand lag-2 prior to commitment in a C. elegans cell fate decision. Cell 1994, 79, 1187–1198. [Google Scholar] [CrossRef]
- Singh, K.; Chao, M.Y.; Somers, G.A.; Komatsu, H.; Corkins, M.E.; Larkins-Ford, J.; Tucey, T.; Dionne, H.M.; Walsh, M.B.; Beaumont, E.K.; et al. C. elegans Notch signaling regulates adult chemosensory response and larval molting quiescence. Curr. Biol. 2011, 21, 825–834. [Google Scholar] [CrossRef]
- Yoo, A.S.; Bais, C.; Greenwald, I. Crosstalk between the EGFR and LIN-12/Notch pathways in C. elegans vulval development. Science 2004, 303, 663–666. [Google Scholar] [CrossRef]
- Cho, S.J.; Lee, M.S.; Tak, E.S.; Lee, E.; Koh, K.S.; Ahn, C.H.; Park, S.C. Gene expression profile in the anterior regeneration of the earthworm using expressed sequence tags. Biosci. Biotechnol. Biochem. 2009, 73, 29–34. [Google Scholar] [CrossRef]
- Yu, Y.S.; Kim, J.S.; Jimenez, B.I.M.; Kim, T.W.; Cho, S.J. Differential expression of primary pair-rule genes during bidirectional regeneration in Perionyx excavatus. Genes Genom. 2018, 40, 747–753. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Y.; Zhang, Y.; Kang, M.; Li, Y.; James, S.W.; Yang, Y.; Bi, Y.; Jiang, H.; Zhao, Y.; et al. Amynthas corticis genome reveals molecular mechanisms behind global distribution. Commun. Biol. 2021, 4, 135. [Google Scholar] [CrossRef]
- Hebrok, M.; Wertz, K.; Fuchtbauer, E.M. M-twist is an inhibitor of muscle differentiation. Dev. Biol. 1994, 165, 537–544. [Google Scholar] [CrossRef]
- Gazave, E.; Guillou, A.; Balavoine, G. History of a prolific family: The Hes/Hey-related genes of the annelid Platynereis. EvoDevo 2014, 5, 29. [Google Scholar] [CrossRef]
- Gazave, E.; Lemaitre, Q.I.; Balavoine, G. The Notch pathway in the annelid Platynereis: Insights into chaetogenesis and neurogenesis processes. Open Biol. 2017, 7, 160242. [Google Scholar] [CrossRef]
- Simionato, E.; Kerner, P.; Dray, N.; Le Gouar, M.; Ledent, V.; Arendt, D.; Vervoort, M. atonal- and achaete-scute-related genes in the annelid Platynereis dumerilii: Insights into the evolution of neural basic-Helix-Loop-Helix genes. BMC Evol. Biol. 2008, 8, 170. [Google Scholar] [CrossRef]
- Pfeifer, K.; Schaub, C.; Wolfstetter, G.; Dorresteijn, A. Identification and characterization of a twist ortholog in the polychaete annelid Platynereis dumerilii reveals mesodermal expression of Pdu-twist. Dev. Genes Evol. 2013, 223, 319–328. [Google Scholar] [CrossRef]
- Thamm, K.; Seaver, E.C. Notch signaling during larval and juvenile development in the polychaete annelid Capitella sp. I. Dev. Biol. 2008, 320, 304–318. [Google Scholar] [CrossRef]
- Meyer, N.P.; Seaver, E.C. Neurogenesis in an annelid: Characterization of brain neural precursors in the polychaete Capitella sp. I. Dev. Biol. 2009, 335, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Rivera, A.S.; Gonsalves, F.C.; Song, M.H.; Norris, B.J.; Weisblat, D.A. Characterization of Notch-class gene expression in segmentation stem cells and segment founder cells in Helobdella robusta (Lophotrochozoa; Annelida; Clitellata; Hirudinida; Glossiphoniidae). Evol. Dev. 2005, 7, 588–599. [Google Scholar] [CrossRef]
- Rivera, A.S.; Weisblat, D.A. And Lophotrochozoa makes three: Notch/Hes signaling in annelid segmentation. Dev. Genes Evol. 2009, 219, 37–43. [Google Scholar] [CrossRef]
- Gharbiah, M.; Nakamoto, A.; Johnson, A.B.; Lambert, J.D.; Nagy, L.M. Ilyanassa Notch signaling implicated in dynamic signaling between all three germ layers. Int. J. Dev. Biol. 2014, 58, 551–562. [Google Scholar] [CrossRef]
- Gibson, B.L. Cellular and ultrastructural features of the regenerating adult eye in the marine gastropod llyanassa obsoleta. J. Morphol. 1984, 180, 145–157. [Google Scholar] [CrossRef]
- Sun, W.; Song, X.; Dong, M.; Liu, Z.; Song, Y.; Wang, L.; Song, L. DNA binding protein CgIkaros-like regulates the proliferation of agranulocytes and granulocytes in oyster (Crassostrea gigas). Dev. Comp. Immunol. 2021, 124, 104201. [Google Scholar] [CrossRef]
- Feng, D.; Li, Q.; Yu, H.; Zhao, X.; Kong, L. Comparative Transcriptome Analysis of the Pacific Oyster Crassostrea gigas Characterized by Shell Colors: Identification of Genetic Bases Potentially Involved in Pigmentation. PLoS ONE 2015, 10, e0145257. [Google Scholar] [CrossRef]
- Wang, L.; Song, X.; Song, L. The oyster immunity. Dev. Comp. Immunol. 2018, 80, 99–118. [Google Scholar] [CrossRef] [PubMed]
- Yue, X.; Nie, Q.; Xiao, G.; Liu, B. Transcriptome analysis of shell color-related genes in the clam Meretrix meretrix. Mar. Biotechnol. 2015, 17, 364–374. [Google Scholar] [CrossRef]
- Dai, M.; Zhang, Y.; Jiao, Y.; Deng, Y.; Du, X.; Yang, C. Immunomodulatory effects of one novel microRNA miR-63 in pearl oyster Pinctada fucata martensii. Fish Shellfish Immunol. 2023, 140, 109002. [Google Scholar] [CrossRef]
- Auffret, P.; Le Luyer, J.; Sham Koua, M.; Quillien, V.; Ky, C.L. Tracing key genes associated with the Pinctada margaritifera albino phenotype from juvenile to cultured pearl harvest stages using multiple whole transcriptome sequencing. BMC Genom. 2020, 21, 662. [Google Scholar] [CrossRef]
- Koenig, K.M.; Sun, P.; Meyer, E.; Gross, J.M. Eye development and photoreceptor differentiation in the cephalopod Doryteuthis pealeii. Development 2016, 143, 3168–3181. [Google Scholar] [CrossRef]
- Napoli, F.R.; Daly, C.M.; Neal, S.; McCulloch, K.J.; Zaloga, A.R.; Liu, A.; Koenig, K.M. Cephalopod retinal development shows vertebrate-like mechanisms of neurogenesis. Curr. Biol. 2022, 32, 5045–5056.e3. [Google Scholar] [CrossRef]
- Fleming, R.J.; Scottgale, T.N.; Diederich, R.J.; Artavanis-Tsakonas, S. The gene Serrate encodes a putative EGF-like transmembrane protein essential for proper ectodermal development in Drosophila melanogaster. Genes Dev. 1990, 4, 2188–2201. [Google Scholar] [CrossRef]
- Thomas, U.; Speicher, S.A.; Knust, E. The Drosophila gene Serrate encodes an EGF-like transmembrane protein with a complex expression pattern in embryos and wing discs. Development 1991, 111, 749–761. [Google Scholar] [CrossRef]
- Vassin, H.; Bremer, K.A.; Knust, E.; Campos-Ortega, J.A. The neurogenic gene Delta of Drosophila melanogaster is expressed in neurogenic territories and encodes a putative transmembrane protein with EGF-like repeats. EMBO J. 1987, 6, 3431–3440. [Google Scholar] [CrossRef]
- Artavanis-Tsakonas, S.; Muskavitch, M.A.; Yedvobnick, B. Molecular cloning of Notch, a locus affecting neurogenesis in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1983, 80, 1977–1981. [Google Scholar] [CrossRef]
- de Celis, J.F.; Garcia-Bellido, A. Roles of the Notch gene in Drosophila wing morphogenesis. Mech. Dev. 1994, 46, 109–122. [Google Scholar] [CrossRef]
- Gho, M.; Lecourtois, M.; Geraud, G.; Posakony, J.W.; Schweisguth, F. Subcellular localization of Suppressor of Hairless in Drosophila sense organ cells during Notch signalling. Development 1996, 122, 1673–1682. [Google Scholar] [CrossRef]
- Ligoxygakis, P.; Yu, S.Y.; Delidakis, C.; Baker, N.E. A subset of notch functions during Drosophila eye development require Su(H) and the E(spl) gene complex. Development 1998, 125, 2893–2900. [Google Scholar] [CrossRef]
- Stempfle, D.; Kanwar, R.; Loewer, A.; Fortini, M.E.; Merdes, G. In vivo reconstitution of gamma-secretase in Drosophila results in substrate specificity. Mol. Cell. Biol. 2010, 30, 3165–3175. [Google Scholar] [CrossRef]
- Guo, Y.; Livne-Bar, I.; Zhou, L.; Boulianne, G.L. Drosophila presenilin is required for neuronal differentiation and affects notch subcellular localization and signaling. J. Neurosci. 1999, 19, 8435–8442. [Google Scholar] [CrossRef]
- Cooper, E.; Deng, W.M.; Chung, H.M. Aph-1 is required to regulate Presenilin-mediated gamma-secretase activity and cell survival in Drosophila wing development. Genesis 2009, 47, 169–174. [Google Scholar] [CrossRef][Green Version]
- Lopez-Schier, H.; St Johnston, D. Drosophila nicastrin is essential for the intramembranous cleavage of notch. Dev. Cell. 2002, 2, 79–89. [Google Scholar] [CrossRef]
- Kux, K.; Kiparaki, M.; Delidakis, C. The two Tribolium E(spl) genes show evolutionarily conserved expression and function during embryonic neurogenesis. Mech. Dev. 2013, 130, 207–225. [Google Scholar] [CrossRef]
- Baumer, D.; Strohlein, N.M.; Schoppmeier, M. Opposing effects of Notch-signaling in maintaining the proliferative state of follicle cells in the telotrophic ovary of the beetle Tribolium. Front. Zool. 2012, 9, 15. [Google Scholar] [CrossRef]
- Liu, W.; Singh, S.R.; Hou, S.X. JAK-STAT is restrained by Notch to control cell proliferation of the Drosophila intestinal stem cells. J. Cell. Biochem. 2010, 109, 992–999. [Google Scholar] [CrossRef]
- Chipman, A.D.; Stollewerk, A. Specification of neural precursor identity in the geophilomorph centipede Strigamia maritima. Dev. Biol. 2006, 290, 337–350. [Google Scholar] [CrossRef]
- Chipman, A.D.; Ferrier, D.E.; Brena, C.; Qu, J.; Hughes, D.S.; Schroder, R.; Torres-Oliva, M.; Znassi, N.; Jiang, H.; Almeida, F.C.; et al. The first myriapod genome sequence reveals conservative arthropod gene content and genome organisation in the centipede Strigamia maritima. PLoS Biol. 2014, 12, e1002005. [Google Scholar] [CrossRef]
- Stollewerk, A. Recruitment of cell groups through Delta/Notch signalling during spider neurogenesis. Development 2002, 129, 5339–5348. [Google Scholar] [CrossRef]
- Schoppmeier, M.; Damen, W.G. Suppressor of Hairless and Presenilin phenotypes imply involvement of canonical Notch-signalling in segmentation of the spider Cupiennius salei. Dev. Biol. 2005, 280, 211–224. [Google Scholar] [CrossRef]
- Zhou, H.; Zhao, W.; Zheng, Z.; Aweya, J.J.; Zhang, Y.; Zhu, J.; Zhao, Y.; Chen, X.; Yao, D. The Notch receptor-ligand Delta is involved in the immune response of Penaeus vannamei. Dev. Comp. Immunol. 2021, 125, 104147. [Google Scholar] [CrossRef]
- Zhao, W.; Fang, Y.; Zheng, Z.; Lin, Z.; Zhao, Y.; Chen, X.; Yao, D.; Zhang, Y. The transcription factor CSL homolog in Penaeus vannamei positively regulates the transcription of the hemocyanin small subunit gene. Dev. Comp. Immunol. 2023, 145, 104723. [Google Scholar] [CrossRef]
- Zhao, W.; Zheng, Z.; Aweya, J.J.; Wang, F.; Li, S.; Tuan, T.N.; Yao, D.; Zhang, Y. Litopenaeus vannamei Notch interacts with COP9 signalosome complex subunit 1 (CNS1) to negatively regulate the NF-kappaB pathway. J. Proteom. 2021, 232, 104074. [Google Scholar] [CrossRef]
- Zhao, W.; Yu, Z.; Aweya, J.J.; Wang, F.; Yao, D.; Ma, H.; Lun, J.; Zhang, Y. Molecular cloning and functional characterization of a homolog of the transcriptional regulator CSL in Litopenaeus vannamei. Dev. Comp. Immunol. 2018, 88, 152–160. [Google Scholar] [CrossRef]
- Karasawa, K.; Sakamoto, N.; Fujita, K.; Ochiai, H.; Fujii, T.; Akasaka, K.; Yamamoto, T. Suppressor of Hairless (Su(H)) is required for foregut development in the sea urchin embryo. Zool. Sci. 2009, 26, 686–690. [Google Scholar] [CrossRef]
- Yaguchi, J.; Angerer, L.M.; Inaba, K.; Yaguchi, S. Zinc finger homeobox is required for the differentiation of serotonergic neurons in the sea urchin embryo. Dev. Biol. 2012, 363, 74–83. [Google Scholar] [CrossRef]
- Walton, K.D.; Croce, J.C.; Glenn, T.D.; Wu, S.Y.; McClay, D.R. Genomics and expression profiles of the Hedgehog and Notch signaling pathways in sea urchin development. Dev. Biol. 2006, 300, 153–164. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, Z.; Leng, X.; Jiang, H.; Liu, L.; Li, C.; Chang, Y. Transcriptome sequencing reveals phagocytosis as the main immune response in the pathogen-challenged sea urchin Strongylocentrotus intermedius. Fish Shellfish Immunol. 2019, 94, 780–791. [Google Scholar] [CrossRef]
- Erkenbrack, E.M.; Davidson, E.H.; Peter, I.S. Conserved regulatory state expression controlled by divergent developmental gene regulatory networks in echinoids. Development 2018, 145. [Google Scholar] [CrossRef]
- Mellott, D.O.; Thisdelle, J.; Burke, R.D. Notch signaling patterns neurogenic ectoderm and regulates the asymmetric division of neural progenitors in sea urchin embryos. Development 2017, 144, 3602–3611. [Google Scholar] [CrossRef]
- Konrad, K.D.; Song, J.L. microRNA-124 regulates Notch and NeuroD1 to mediate transition states of neuronal development. Dev. Neurobiol. 2023, 83, 3–27. [Google Scholar] [CrossRef] [PubMed]
- Burke, R.D.; Angerer, L.M.; Elphick, M.R.; Humphrey, G.W.; Yaguchi, S.; Kiyama, T.; Liang, S.; Mu, X.; Agca, C.; Klein, W.H.; et al. A genomic view of the sea urchin nervous system. Dev. Biol. 2006, 300, 434–460. [Google Scholar] [CrossRef] [PubMed]
- Peterson, R.E.; McClay, D.R. A Fringe-modified Notch signal affects specification of mesoderm and endoderm in the sea urchin embryo. Dev. Biol. 2005, 282, 126–137. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Slota, L.A.; McClay, D.R. Identification of neural transcription factors required for the differentiation of three neuronal subtypes in the sea urchin embryo. Dev. Biol. 2018, 435, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Reinardy, H.C.; Emerson, C.E.; Manley, J.M.; Bodnar, A.G. Tissue regeneration and biomineralization in sea urchins: Role of Notch signaling and presence of stem cell markers. PLoS ONE 2015, 10, e0133860. [Google Scholar] [CrossRef] [PubMed]
- Hinman, V.F.; Davidson, E.H. Evolutionary plasticity of developmental gene regulatory network architecture. Proc. Natl. Acad. Sci. USA 2007, 104, 19404–19409. [Google Scholar] [CrossRef]
- McCauley, B.S.; Akyar, E.; Saad, H.R.; Hinman, V.F. Dose-dependent nuclear beta-catenin response segregates endomesoderm along the sea star primary axis. Development 2015, 142, 207–217. [Google Scholar] [CrossRef]
- Perillo, M.; Swartz, S.Z.; Pieplow, C.; Wessel, G.M. Molecular mechanisms of tubulogenesis revealed in the sea star hydro-vascular organ. Nat. Commun. 2023, 14, 2402. [Google Scholar] [CrossRef] [PubMed]
- Mashanov, V.; Machado, D.J.; Reid, R.; Brouwer, C.; Kofsky, J.; Janies, D.A. Twinkle twinkle brittle star: The draft genome of Ophioderma brevispinum (Echinodermata: Ophiuroidea) as a resource for regeneration research. BMC Genom. 2022, 23, 574. [Google Scholar] [CrossRef] [PubMed]
- Mashanov, V.; Akiona, J.; Khoury, M.; Ferrier, J.; Reid, R.; Machado, D.J.; Zueva, O.; Janies, D. Active Notch signaling is required for arm regeneration in a brittle star. PLoS ONE 2020, 15, e0232981. [Google Scholar] [CrossRef]
- Xing, L.; Sun, L.; Liu, S.; Li, X.; Zhang, L.; Yang, H. De Novo assembly and comparative transcriptome analyses of purple and green morphs of Apostichopus japonicus during body wall pigmentation process. Comp. Biochem. Physiol. Part D Genom. Proteom. 2018, 28, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Yang, H.; Chen, M.; Ma, D.; Lin, C. RNA-Seq reveals dynamic changes of gene expression in key stages of intestine regeneration in the sea cucumber Apostichopus japonicus. PLoS ONE 2013, 8, e69441, Erratum in PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Zondag, L.E.; Rutherford, K.; Gemmell, N.J.; Wilson, M.J. Uncovering the pathways underlying whole body regeneration in a chordate model, Botrylloides leachi using de novo transcriptome analysis. BMC Genom. 2016, 17, 114. [Google Scholar] [CrossRef] [PubMed]
- Hamada, M.; Goricki, S.; Byerly, M.S.; Satoh, N.; Jeffery, W.R. Evolution of the chordate regeneration blastema: Differential gene expression and conserved role of notch signaling during siphon regeneration in the ascidian Ciona. Dev. Biol. 2015, 405, 304–315. [Google Scholar] [CrossRef]
- Chen, J.S.; Pedro, M.S.; Zeller, R.W. miR-124 function during Ciona intestinalis neuronal development includes extensive interaction with the Notch signaling pathway. Development 2011, 138, 4943–4953. [Google Scholar] [CrossRef]
- Hori, S.; Saitoh, T.; Matsumoto, M.; Makabe, K.W.; Nishida, H. Notch homologue from Halocynthia roretzi is preferentially expressed in the central nervous system during ascidian embryogenesis. Dev. Genes Evol. 1997, 207, 371–380. [Google Scholar] [CrossRef]
- Akanuma, T.; Hori, S.; Darras, S.; Nishida, H. Notch signaling is involved in nervous system formation in ascidian embryos. Dev. Genes Evol. 2002, 212, 459–472. [Google Scholar] [CrossRef]
- Karpov, S.A. Flagellar apparatus structure of choanoflagellates. Cilia 2016, 5, 11. [Google Scholar] [CrossRef]
- Leadbeater; Barry, S.C. Life-history and ultrastructure of a new marine species of Proterospongia (Choanoflagellida). J. Mar. Biol. Assoc. U. K. 1983, 63, 135–160. [Google Scholar] [CrossRef]
- King, N.; Westbrook, M.J.; Young, S.L.; Kuo, A.; Abedin, M.; Chapman, J.; Fairclough, S.; Hellsten, U.; Isogai, Y.; Letunic, I.; et al. The genome of the choanoflagellate Monosiga brevicollis and the origin of metazoans. Nature 2008, 451, 783–788. [Google Scholar] [CrossRef] [PubMed]
- Leys, S.P.; Rohksar, D.S.; Degnan, B.M. Sponges. Curr. Biol. 2005, 15, R114–R115. [Google Scholar] [CrossRef] [PubMed]
- Richards, G.S.; Simionato, E.; Perron, M.; Adamska, M.; Vervoort, M.; Degnan, B.M. Sponge genes provide new insight into the evolutionary origin of the neurogenic circuit. Curr. Biol. 2008, 18, 1156–1161. [Google Scholar] [CrossRef] [PubMed]
- Six, E.; Ndiaye, D.; Laabi, Y.; Brou, C.; Gupta-Rossi, N.; Israel, A.; Logeat, F. The Notch ligand Delta1 is sequentially cleaved by an ADAM protease and gamma-secretase. Proc. Natl. Acad. Sci. USA 2003, 100, 7638–7643. [Google Scholar] [CrossRef] [PubMed]
- Leys, S.P.; Degnan, B.M. Cytological basis of photoresponsive behavior in a sponge larva. Biol. Bull. 2001, 201, 323–338. [Google Scholar] [CrossRef] [PubMed]
- Renard, E.; Vacelet, J.; Gazave, E.; Lapebie, P.; Borchiellini, C.; Ereskovsky, A.V. Origin of the neuro-sensory system: New and expected insights from sponges. Integr. Zool. 2009, 4, 294–308. [Google Scholar] [CrossRef] [PubMed]
- Sakarya, O.; Armstrong, K.A.; Adamska, M.; Adamski, M.; Wang, I.F.; Tidor, B.; Degnan, B.M.; Oakley, T.H.; Kosik, K.S. A post-synaptic scaffold at the origin of the animal kingdom. PLoS ONE 2007, 2, e506. [Google Scholar] [CrossRef]
- Srivastava, M.; Larroux, C.; Lu, D.R.; Mohanty, K.; Chapman, J.; Degnan, B.M.; Rokhsar, D.S. Early evolution of the LIM homeobox gene family. BMC Biol. 2010, 8, 4. [Google Scholar] [CrossRef]
- Kayal, E.; Roure, B.; Philippe, H.; Collins, A.G.; Lavrov, D.V. Cnidarian phylogenetic relationships as revealed by mitogenomics. BMC Evol. Biol. 2013, 13, 5. [Google Scholar] [CrossRef]
- Abrams, M.J.; Tan, F.H.; Li, Y.; Basinger, T.; Heithe, M.L.; Sarma, A.; Lee, I.T.; Condiotte, Z.J.; Raffiee, M.; Dabiri, J.O.; et al. A conserved strategy for inducing appendage regeneration in moon jellyfish, Drosophila, and mice. eLife 2021, 10, e65092. [Google Scholar] [CrossRef]
- Traylor-Knowles, N.; Emery, M. Analysis of Spatial Gene Expression at the Cellular Level in Stony Corals. In Whole-Body Regeneration: Methods and Protocols; Methods in Molecular Biology; Springer: New York, NY, USA, 2022; Volume 2450, pp. 359–371. [Google Scholar] [CrossRef]
- Havrilak, J.A.; Al-Shaer, L.; Baban, N.; Akinci, N.; Layden, M.J. Characterization of the dynamics and variability of neuronal subtype responses during growth, degrowth, and regeneration of Nematostella vectensis. BMC Biol. 2021, 19, 104. [Google Scholar] [CrossRef]
- Bode, H.R. Head regeneration in Hydra. Dev. Dyn. 2003, 226, 225–236. [Google Scholar] [CrossRef]
- Galliot, B.; Quiquand, M.; Ghila, L.; de Rosa, R.; Miljkovic-Licina, M.; Chera, S. Origins of neurogenesis, a cnidarian view. Dev. Biol. 2009, 332, 2–24. [Google Scholar] [CrossRef]
- Marlow, H.Q.; Srivastava, M.; Matus, D.Q.; Rokhsar, D.; Martindale, M.Q. Anatomy and development of the nervous system of Nematostella vectensis, an anthozoan cnidarian. Dev. Neurobiol. 2009, 69, 235–254. [Google Scholar] [CrossRef]
- Putnam, N.H.; Srivastava, M.; Hellsten, U.; Dirks, B.; Chapman, J.; Salamov, A.; Terry, A.; Shapiro, H.; Lindquist, E.; Kapitonov, V.V.; et al. Sea anemone genome reveals ancestral eumetazoan gene repertoire and genomic organization. Science 2007, 317, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Burnett, A.L.; Diehl, N.A. The Nervous System of Hydra. I. Types, Distribution and Origin of Nerve Elements. J. Exp. Zool. 1964, 157, 217–226. [Google Scholar] [CrossRef]
- Lentz, T.L.; Barrnett, R.J. Fine Structure of the Nervous System of Hydra. Am. Zool. 1965, 5, 341–356. [Google Scholar] [CrossRef]
- Hanson, A. On being a Hydra with, and without, a nervous system: What do neurons add? Anim. Cogn. 2023, 26, 1799–1816. [Google Scholar] [CrossRef]
- Bossert, P.E.; Dunn, M.P.; Thomsen, G.H. A staging system for the regeneration of a polyp from the aboral physa of the anthozoan Cnidarian Nematostella vectensis. Dev. Dyn. 2013, 242, 1320–1331. [Google Scholar] [CrossRef]
- Passamaneck, Y.J.; Martindale, M.Q. Cell proliferation is necessary for the regeneration of oral structures in the anthozoan cnidarian Nematostella vectensis. BMC Dev. Biol. 2012, 12, 34. [Google Scholar] [CrossRef]
- Egger, B.; Lapraz, F.; Tomiczek, B.; Muller, S.; Dessimoz, C.; Girstmair, J.; Skunca, N.; Rawlinson, K.A.; Cameron, C.B.; Beli, E.; et al. A transcriptomic-phylogenomic analysis of the evolutionary relationships of flatworms. Curr. Biol. 2015, 25, 1347–1353. [Google Scholar] [CrossRef]
- Reuter, M.; Gustafsson, M.K. The flatworm nervous system: Pattern and phylogeny. In The Nervous Systems of Invertebrates: An Evolutionary and Comparative Approach; Experientia Supplementum; Birkhäuser: Basel, Switzerland, 1995; Volume 72, pp. 25–59. [Google Scholar] [CrossRef]
- Collins, J.J., 3rd. Platyhelminthes. Curr. Biol. 2017, 27, R252–R256. [Google Scholar] [CrossRef]
- Roberts-Galbraith, R.H.; Newmark, P.A. On the organ trail: Insights into organ regeneration in the planarian. Curr. Opin. Genet. Dev. 2015, 32, 37–46. [Google Scholar] [CrossRef]
- Ross, K.G.; Currie, K.W.; Pearson, B.J.; Zayas, R.M. Nervous system development and regeneration in freshwater planarians. Wiley Interdiscip. Rev. Dev. Biol. 2017, 6, e266. [Google Scholar] [CrossRef]
- Schadt, T.; Prantl, V.; Grosbusch, A.L.; Bertemes, P.; Egger, B. Regeneration of the flatworm Prosthiostomum siphunculus (Polycladida, Platyhelminthes). Cell Tissue Res. 2021, 383, 1025–1041. [Google Scholar] [CrossRef] [PubMed]
- LoVerde, P.T. Schistosomiasis. Adv. Exp. Med. Biol. 2019, 1154, 45–70. [Google Scholar] [CrossRef] [PubMed]
- Rollinson, D.; Knopp, S.; Levitz, S.; Stothard, J.R.; Tchuem Tchuente, L.A.; Garba, A.; Mohammed, K.A.; Schur, N.; Person, B.; Colley, D.G.; et al. Time to set the agenda for schistosomiasis elimination. Acta Trop. 2013, 128, 423–440. [Google Scholar] [CrossRef] [PubMed]
- Buddenborg, S.K.; Lu, Z.; Sankaranarayan, G.; Doyle, S.R.; Berriman, M. The stage- and sex-specific transcriptome of the human parasite Schistosoma mansoni. Sci. Data 2023, 10, 775. [Google Scholar] [CrossRef]
- Senft, A.W.; Weller, T.H. Growth and regeneration of Schistosoma mansoni in vitro. Proc. Soc. Exp. Biol. Med. 1956, 93, 16–19. [Google Scholar] [CrossRef]
- Reda, E.S.; El-Shabasy, E.A.; Said, A.E.; Mansour, M.F.; Saleh, M.A. Cholinergic components of nervous system of Schistosoma mansoni and S. haematobium (Digenea: Schistosomatidae). Parasitol. Res. 2016, 115, 3127–3137. [Google Scholar] [CrossRef]
- Budke, C.M.; Casulli, A.; Kern, P.; Vuitton, D.A. Cystic and alveolar echinococcosis: Successes and continuing challenges. PLoS Negl. Trop. Dis. 2017, 11, e0005477. [Google Scholar] [CrossRef] [PubMed]
- El Bejjani, R.; Hammarlund, M. Neural regeneration in Caenorhabditis elegans. Annu. Rev. Genet. 2012, 46, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Oikonomou, G.; Shaham, S. The glia of Caenorhabditis elegans. Glia 2011, 59, 1253–1263. [Google Scholar] [CrossRef]
- Barriere, A.; Bertrand, V. Neuronal specification in C. elegans: Combining lineage inheritance with intercellular signaling. J. Neurogenet. 2020, 34, 273–281. [Google Scholar] [CrossRef]
- Chao, M.Y.; Larkins-Ford, J.; Tucey, T.M.; Hart, A.C. lin-12 Notch functions in the adult nervous system of C. elegans. BMC Neurosci. 2005, 6, 45. [Google Scholar] [CrossRef]
- El Bejjani, R.; Hammarlund, M. Notch signaling inhibits axon regeneration. Neuron 2012, 73, 268–278. [Google Scholar] [CrossRef]
- Greenwald, I.; Kovall, R. Notch signaling: Genetics and structure. In WormBook: The Online Review of C. elegans Biology; WormBook: Pasadena, CA, USA, 2013; pp. 1–28. [Google Scholar] [CrossRef]
- Li, X.; Greenwald, I. HOP-1, a Caenorhabditis elegans presenilin, appears to be functionally redundant with SEL-12 presenilin and to facilitate LIN-12 and GLP-1 signaling. Proc. Natl. Acad. Sci. USA 1997, 94, 12204–12209. [Google Scholar] [CrossRef]
- Westlund, B.; Parry, D.; Clover, R.; Basson, M.; Johnson, C.D. Reverse genetic analysis of Caenorhabditis elegans presenilins reveals redundant but unequal roles for sel-12 and hop-1 in Notch-pathway signaling. Proc. Natl. Acad. Sci. USA 1999, 96, 2497–2502. [Google Scholar] [CrossRef]
- Yu, G.; Nishimura, M.; Arawaka, S.; Levitan, D.; Zhang, L.; Tandon, A.; Song, Y.Q.; Rogaeva, E.; Chen, F.; Kawarai, T.; et al. Nicastrin modulates presenilin-mediated notch/glp-1 signal transduction and betaAPP processing. Nature 2000, 407, 48–54. [Google Scholar] [CrossRef]
- Priess, J.R. Notch signaling in the C. elegans embryo. In WormBook: The Online Review of C. elegans Biology; WormBook: Pasadena, CA, USA, 2005; pp. 1–16. [Google Scholar] [CrossRef]
- Iwatsubo, T. The gamma-secretase complex: Machinery for intramembrane proteolysis. Curr. Opin. Neurobiol. 2004, 14, 379–383. [Google Scholar] [CrossRef]
- Wood, W.B.; Edgar, L.G. Patterning in the C. elegans embryo. Trends Genet. 1994, 10, 49–54. [Google Scholar] [CrossRef]
- Lambie, E.J.; Kimble, J. Two homologous regulatory genes, lin-12 and glp-1, have overlapping functions. Development 1991, 112, 231–240. [Google Scholar] [CrossRef]
- Fitzgerald, K.; Wilkinson, H.A.; Greenwald, I. glp-1 can substitute for lin-12 in specifying cell fate decisions in Caenorhabditis elegans. Development 1993, 119, 1019–1027. [Google Scholar] [CrossRef]
- Goutte, C.; Hepler, W.; Mickey, K.M.; Priess, J.R. aph-2 encodes a novel extracellular protein required for GLP-1-mediated signaling. Development 2000, 127, 2481–2492. [Google Scholar] [CrossRef] [PubMed]
- Levitan, D.; Yu, G.; St George Hyslop, P.; Goutte, C. APH-2/nicastrin functions in LIN-12/Notch signaling in the Caenorhabditis elegans somatic gonad. Dev. Biol. 2001, 240, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Goutte, C.; Tsunozaki, M.; Hale, V.A.; Priess, J.R. APH-1 is a multipass membrane protein essential for the Notch signaling pathway in Caenorhabditis elegans embryos. Proc. Natl. Acad. Sci. USA 2002, 99, 775–779. [Google Scholar] [CrossRef] [PubMed]
- Priess, J.R.; Thomson, J.N. Cellular interactions in early C. elegans embryos. Cell 1987, 48, 241–250. [Google Scholar] [CrossRef]
- Christensen, S.; Kodoyianni, V.; Bosenberg, M.; Friedman, L.; Kimble, J. lag-1, a gene required for lin-12 and glp-1 signaling in Caenorhabditis elegans, is homologous to human CBF1 and Drosophila Su(H). Development 1996, 122, 1373–1383. [Google Scholar] [CrossRef]
- Petcherski, A.G.; Kimble, J. LAG-3 is a putative transcriptional activator in the C. elegans Notch pathway. Nature 2000, 405, 364–368. [Google Scholar] [CrossRef]
- Austin, J.; Kimble, J. glp-1 is required in the germ line for regulation of the decision between mitosis and meiosis in C. elegans. Cell 1987, 51, 589–599. [Google Scholar] [CrossRef]
- Greenwald, I.S.; Sternberg, P.W.; Horvitz, H.R. The lin-12 locus specifies cell fates in Caenorhabditis elegans. Cell 1983, 34, 435–444. [Google Scholar] [CrossRef]
- Rebeiz, M.; Reeves, N.L.; Posakony, J.W. SCORE: A computational approach to the identification of cis-regulatory modules and target genes in whole-genome sequence data. Site clustering over random expectation. Proc. Natl. Acad. Sci. USA 2002, 99, 9888–9893. [Google Scholar] [CrossRef]
- Fujiwara, M.; Sengupta, P.; McIntire, S.L. Regulation of body size and behavioral state of C. elegans by sensory perception and the EGL-4 cGMP-dependent protein kinase. Neuron 2002, 36, 1091–1102. [Google Scholar] [CrossRef] [PubMed]
- Cuppen, E.; van der Linden, A.M.; Jansen, G.; Plasterk, R.H. Proteins interacting with Caenorhabditis elegans Galpha subunits. Comp. Funct. Genom. 2003, 4, 479–491. [Google Scholar] [CrossRef] [PubMed]
- L’Etoile, N.D.; Coburn, C.M.; Eastham, J.; Kistler, A.; Gallegos, G.; Bargmann, C.I. The cyclic GMP-dependent protein kinase EGL-4 regulates olfactory adaptation in C. elegans. Neuron 2002, 36, 1079–1089. [Google Scholar] [CrossRef] [PubMed]
- Raizen, D.M.; Cullison, K.M.; Pack, A.I.; Sundaram, M.V. A novel gain-of-function mutant of the cyclic GMP-dependent protein kinase egl-4 affects multiple physiological processes in Caenorhabditis elegans. Genetics 2006, 173, 177–187. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hukema, R.K.; Rademakers, S.; Dekkers, M.P.; Burghoorn, J.; Jansen, G. Antagonistic sensory cues generate gustatory plasticity in Caenorhabditis elegans. EMBO J. 2006, 25, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.C.; Westheide, W. Structure of the nervous system of Myzostoma cirriferum (Annelida) as revealed by immunohistochemistry and cLSM analyses. J. Morphol. 2000, 245, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Bely, A.E. Early events in annelid regeneration: A cellular perspective. Integr. Comp. Biol. 2014, 54, 688–699. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.C. Nerve development, growth and differentiation during regeneration in Enchytraeus fragmentosus and Stylaria lacustris (Oligochaeta). Dev. Growth Differ. 2004, 46, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Bodo, K.; Kellermayer, Z.; Laszlo, Z.; Boros, A.; Kokhanyuk, B.; Nemeth, P.; Engelmann, P. Injury-Induced Innate Immune Response during Segment Regeneration of the Earthworm, Eisenia andrei. Int. J. Mol. Sci. 2021, 22, 2363. [Google Scholar] [CrossRef] [PubMed]
- Kostyuchenko, R.P.; Kozin, V.V. Comparative Aspects of Annelid Regeneration: Towards Understanding the Mechanisms of Regeneration. Genes 2021, 12, 1148. [Google Scholar] [CrossRef] [PubMed]
- Giani, V.C., Jr.; Yamaguchi, E.; Boyle, M.J.; Seaver, E.C. Somatic and germline expression of piwi during development and regeneration in the marine polychaete annelid Capitella teleta. EvoDevo 2011, 2, 10. [Google Scholar] [CrossRef]
- Meyer, N.P.; Carrillo-Baltodano, A.; Moore, R.E.; Seaver, E.C. Nervous system development in lecithotrophic larval and juvenile stages of the annelid Capitella teleta. Front. Zool. 2015, 12, 15. [Google Scholar] [CrossRef]
- Macagno, E.R.; Gaasterland, T.; Edsall, L.; Bafna, V.; Soares, M.B.; Scheetz, T.; Casavant, T.; Da Silva, C.; Wincker, P.; Tasiemski, A.; et al. Construction of a medicinal leech transcriptome database and its application to the identification of leech homologs of neural and innate immune genes. BMC Genom. 2010, 11, 407. [Google Scholar] [CrossRef]
- Macagno, E.R. Number and distribution of neurons in leech segmental ganglia. J. Comp. Neurol. 1980, 190, 283–302. [Google Scholar] [CrossRef]
- Williams, S.T. Molluscan shell colour. Biol. Rev. Camb. Philos. Soc. 2017, 92, 1039–1058. [Google Scholar] [CrossRef]
- Neff, J.M. Ultrastructural studies of periostracum formation in the hard shelled clam Mercenaria mercenaria (L). Tissue Cell. 1972, 4, 311–326. [Google Scholar] [CrossRef]
- Baratte, S.; Bonnaud, L. Evidence of early nervous differentiation and early catecholaminergic sensory system during Sepia officinalis embryogenesis. J. Comp. Neurol. 2009, 517, 539–549. [Google Scholar] [CrossRef]
- Matsuo, R.; Ito, E. Spontaneous regeneration of the central nervous system in gastropods. Biol. Bull. 2011, 221, 35–42. [Google Scholar] [CrossRef]
- Callaghan, N.I.; Capaz, J.C.; Lamarre, S.G.; Bourloutski, E.; Oliveira, A.R.; MacCormack, T.J.; Driedzic, W.R.; Sykes, A.V. Reversion to developmental pathways underlies rapid arm regeneration in juvenile European cuttlefish, Sepia officinalis (Linnaeus 1758). J. Exp. Zool. B Mol. Dev. Evol. 2019, 332, 113–120. [Google Scholar] [CrossRef]
- Lambert, J.D. Chapter 5. Patterning the spiralian embryo: Insights from Ilyanassa. Curr. Top. Dev. Biol. 2009, 86, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.; He, P.; Zhang, X.; Li, W.; Zhang, L.; Guan, J.; Chen, X.; Lin, Y.; Zhuo, X.; Li, Q.; et al. Identification and characterization of microRNAs in the gonads of Crassostrea hongkongensis using high-throughput sequencing. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 31, 100606. [Google Scholar] [CrossRef]
- Jiao, Y.; Yang, S.; Cao, Y.; Zheng, Z.; Deng, Y.; Wang, Q.; Huang, R.; Du, X. Genome and transcriptome analyses providing insight into the immune response of pearl oysters after allograft and xenograft transplantations. Fish Shellfish Immunol. 2019, 90, 109–117. [Google Scholar] [CrossRef]
- Pungor, J.R.; Niell, C.M. The neural basis of visual processing and behavior in cephalopods. Curr. Biol. 2023, 33, R1106–R1118. [Google Scholar] [CrossRef] [PubMed]
- Imperadore, P.; Parazzoli, D.; Oldani, A.; Duebbert, M.; Buschges, A.; Fiorito, G. From injury to full repair: Nerve regeneration and functional recovery in the common octopus, Octopus vulgaris. J. Exp. Biol. 2019, 222, jeb209965. [Google Scholar] [CrossRef] [PubMed]
- Dingerkus, G.; Santoro, E.D. Cornea regeneration in the Pacific giant octopus, Octopus dofleini, and the common octopus, O. vulgaris. Experientia 1981, 37, 368–369. [Google Scholar] [CrossRef]
- Legg, D.A.; Sutton, M.D.; Edgecombe, G.D. Arthropod fossil data increase congruence of morphological and molecular phylogenies. Nat. Commun. 2013, 4, 2485. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, E.; Bruce, H.S.; Patel, N.H. Evolving specialization of the arthropod nervous system. Proc. Natl. Acad. Sci. USA 2012, 109 (Suppl. S1), 10634–10639. [Google Scholar] [CrossRef]
- Suzuki, Y.; Chou, J.; Garvey, S.L.; Wang, V.R.; Yanes, K.O. Evolution and Regulation of Limb Regeneration in Arthropods. Results Probl. Cell Differ. 2019, 68, 419–454. [Google Scholar] [CrossRef] [PubMed]
- Prpic, N.M.; Damen, W.G. Notch-mediated segmentation of the appendages is a molecular phylotypic trait of the arthropods. Dev. Biol. 2009, 326, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Cordoba, S.; Estella, C. Role of Notch Signaling in Leg Development in Drosophila melanogaster. Adv. Exp. Med. Biol. 2020, 1218, 103–127. [Google Scholar] [CrossRef]
- Allan, D.W.; Thor, S. Transcriptional selectors, masters, and combinatorial codes: Regulatory principles of neural subtype specification. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 505–528. [Google Scholar] [CrossRef] [PubMed]
- Skeath, J.B.; Thor, S. Genetic control of Drosophila nerve cord development. Curr. Opin. Neurobiol. 2003, 13, 8–15. [Google Scholar] [CrossRef]
- Kato, K.; Forero, M.G.; Fenton, J.C.; Hidalgo, A. The glial regenerative response to central nervous system injury is enabled by pros-notch and pros-NFkappaB feedback. PLoS Biol. 2011, 9, e1001133. [Google Scholar] [CrossRef]
- Zhou, H.; Ma, Z.; Wang, Z.; Yan, S.; Wang, D.; Shen, J. Hedgehog signaling regulates regenerative patterning and growth in Harmonia axyridis leg. Cell. Mol. Life Sci. 2021, 78, 2185–2197. [Google Scholar] [CrossRef]
- Freas, C.A.; Cheng, K. Neuroecology beyond the brain: Learning in Echinodermata. Learn. Behav. 2022, 50, 20–36. [Google Scholar] [CrossRef]
- Hoekstra, L.A.; Moroz, L.L.; Heyland, A. Novel insights into the echinoderm nervous system from histaminergic and FMRFaminergic-like cells in the sea cucumber Leptosynapta clarki. PLoS ONE 2012, 7, e44220. [Google Scholar] [CrossRef]
- Pentreath, V.W.; Cobb, J.L. Neurobiology of echinodermata. Biol. Rev. Camb. Philos. Soc. 1972, 47, 363–392. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Akasaka, K. Regeneration in crinoids. Dev. Growth Differ. 2010, 52, 57–68. [Google Scholar] [CrossRef]
- Ferrario, C.; Ben Khadra, Y.; Sugni, M.; Candia Carnevali, M.D.; Martinez, P.; Bonasoro, F. Studying Echinodermata Arm Explant Regeneration Using Echinaster sepositus. Methods Mol. Biol. 2022, 2450, 263–291. [Google Scholar] [CrossRef]
- Hinman, V.F.; Burke, R.D. Embryonic neurogenesis in echinoderms. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e316. [Google Scholar] [CrossRef]
- Cary, G.A.; McCauley, B.S.; Zueva, O.; Pattinato, J.; Longabaugh, W.; Hinman, V.F. Systematic comparison of sea urchin and sea star developmental gene regulatory networks explains how novelty is incorporated in early development. Nat. Commun. 2020, 11, 6235. [Google Scholar] [CrossRef] [PubMed]
- Bronchain, O.; Philippe-Caraty, L.; Anquetil, V.; Ciapa, B. Precise regulation of presenilin expression is required for sea urchin early development. J. Cell Sci. 2021, 134, jcs258382. [Google Scholar] [CrossRef]
- Wood, N.J.; Mattiello, T.; Rowe, M.L.; Ward, L.; Perillo, M.; Arnone, M.I.; Elphick, M.R.; Oliveri, P. Neuropeptidergic Systems in Pluteus Larvae of the Sea Urchin Strongylocentrotus purpuratus: Neurochemical Complexity in a “Simple” Nervous System. Front. Endocrinol. 2018, 9, 628. [Google Scholar] [CrossRef] [PubMed]
- Zueva, O.; Khoury, M.; Heinzeller, T.; Mashanova, D.; Mashanov, V. The complex simplicity of the brittle star nervous system. Front. Zool. 2018, 15, 1. [Google Scholar] [CrossRef]
- Sun, L.; Xu, D.; Xu, Q.; Sun, J.; Xing, L.; Zhang, L.; Yang, H. iTRAQ reveals proteomic changes during intestine regeneration in the sea cucumber Apostichopus japonicus. Comp. Biochem. Physiol. Part D Genom. Proteom. 2017, 22, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Antoniadou, C.; Gerovasileiou, V.; Bailly, N. Ascidiacea (Chordata: Tunicata) of Greece: An updated checklist. Biodivers. Data J. 2016, 4, e9273. [Google Scholar] [CrossRef]
- Kassmer, S.H.; Rodriguez, D.; De Tomaso, A.W. Colonial ascidians as model organisms for the study of germ cells, fertility, whole body regeneration, vascular biology and aging. Curr. Opin. Genet. Dev. 2016, 39, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Kassmer, S.H.; Nourizadeh, S.; De Tomaso, A.W. Cellular and molecular mechanisms of regeneration in colonial and solitary Ascidians. Dev. Biol. 2019, 448, 271–278. [Google Scholar] [CrossRef]
- Jeffery, W.R. Closing the wounds: One hundred and twenty five years of regenerative biology in the ascidian Ciona intestinalis. Genesis 2015, 53, 48–65. [Google Scholar] [CrossRef]
- Whittaker, J.R. Siphon regeneration in Ciona. Nature 1975, 255, 224–225. [Google Scholar] [CrossRef]
- Radtke, F.; Fasnacht, N.; Macdonald, H.R. Notch signaling in the immune system. Immunity 2010, 32, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Liao, B.K.; Oates, A.C. Delta-Notch signalling in segmentation. Arthropod. Struct. Dev. 2017, 46, 429–447. [Google Scholar] [CrossRef]
- Liang, S.J.; Li, X.G.; Wang, X.Q. Notch Signaling in Mammalian Intestinal Stem Cells: Determining Cell Fate and Maintaining Homeostasis. Curr. Stem Cell Res. Ther. 2019, 14, 583–590. [Google Scholar] [CrossRef]
- Rottinger, E. Nematostella vectensis, an Emerging Model for Deciphering the Molecular and Cellular Mechanisms Underlying Whole-Body Regeneration. Cells 2021, 10, 2692. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species Name | Expression | Function | Ref. |
|---|---|---|---|
| Demospongea (A. queenslandica) | |||
| AmDelta1-5 | Embryos and larvae | Embryonic development and neurodevelopment | [40,41,42] |
| AmNotch | Globular cells, anterior polar cells, ciliated cells | ||
| AmHLH1 | |||
| Hexactinellida (A. vastus) | |||
| AvADAM10/17 AvFurinL AvDelta AvNotch AvSuH AvPEN2 AvAPH1 AvPSEN2 AvNCSTN AvNCoRL | [40,41] | ||
| Hexactinellida (O. minuta) | |||
| OmADAM10/17 OmFurinL OmDelta OmNotch OmSuH OmPEN2 OmAPH1 OmPSEN2 OmNCSTN OmNCoRL | [40,41] | ||
| Homoscleromorpha (O. carmela) | |||
| HsADAM10/15/17 HsFurin HsJag1/Jag2 HsDDL1/DDL3/DDL4 HsNotch1 HsSuH | [40,41] | ||
| Cnidaria | |||
| Hydrozoa (H. vulgaris) | |||
| HyJagged | Boundary between bud and parent | Regeneration and development | [43,44,45] |
| HvNotch | Ectodermal and the endodermal | ||
| HvSu(H) | |||
| HyHes-1 | Regenerating heads and tentacle buds | ||
| Hvpen-2 | |||
| Anthozoa (N. vectensis) | |||
| NvDelta/NvJagged | Oral ectoderm and endoderm of the planula stage | Neurodevelopment and regeneration | [46,47,48] |
| NvNotch | Neural structure, pharyngeal endoderm, gastrula and early planula | ||
| NvSuH | Endodermal plate and planula | ||
| NvHes1 | Ectoderm of the gastrula stage | ||
| NvHes2 | Oral ectodermal domain | ||
| NvHes3 | Aboral ectoderm of the gastrula and early planula | ||
| NvHes4 | |||
| Anthozoa (A. cervicornis and A. palmata) | |||
| Delta/Delta-like/Jagged Notch Hairy/E(Spl)/Su(H) E3 ubiquitin ligase MIB Numb | Growth | [49] | |
| Anthozoa(O. faveolata) | |||
| Delta/Jagged ADA NOTCH DTX PEN NICA PSN APH EP300 RBPJL HAIRLESS HES | Innate immune defense | [50] | |
| Platyhelminthes | |||
| Turbellaria (D. japonica) | |||
| DjNotch1–6 | Parenchymal tissues and blastema site | Neurodevelopment and regeneration, eye point development | [51] |
| DjRbpj | |||
| DjHes | |||
| Turbellaria (S. mediterranea) | |||
| SmedNotch1, SmedNotch2, SmedNotch4 | All notch components are expressed in discrete neural populations throughout the brain and in regenerating tissues. | Parenchymal tissues and the blastema site, eye point development. | [52,53] |
| Su(H) | |||
| Atoh | |||
| Coe | |||
| Fer3l-1 | |||
| Hes (Hesl-3) | |||
| Sim | |||
| Trematoda (S. mansoni) | |||
| SmJagged/SmSerrate SmNotch SmSu(H) SmHes SmSmart SmGroucho SmSkip smPresenilin SmNicastrin SmAph-1 SmPen-2 SmFurin SmAdam 17/SmKuzbanian Notchless Numb Dishevelled WWP1 | All identified Notch signaling components are present in all life cycles of the S. mansoni | Oogenesis and embryogenesis | [54] |
| Cestoda (E. granulosus) | |||
| Delta Jagged Notch Presenilin1 Aph1 Pen-2 Su(H) Dishevelled Numb Metalloproteinase domain-containing protein E3 ubiquitin-protein ligase RNF167 ADAM 17-like protease E1A/CREB-binding protei SNW domain-containing protein 1 C-terminal-binding protein Groucho Histone deacetylase 1/2 Furin-like protease 1 | They are expressed at all developmental stages. | Mitotic cell division and proliferation | [55,56,57,58] |
| Nemathelminthes | |||
| Nematoda (C. elegans) | |||
| Lag-2(Sel-3)/Apx-1/Arg-1/Dsl-1 | IL2 neurons | Neurogenesis, embryonic development, normal spawning and vulva development | [59,60,61,62,63,64,65,66] |
| Lin-12/Glp-1 | Larval RIG neurons, somatic gonadal, and vulvar profiles | ||
| Sel-12/Hop-1 | Early blastomeres | ||
| Aph-1/Aph-2 | |||
| Pen-2 | |||
| Lag-1/Sel-8(Lag-3) | |||
| Annelida | |||
| Earthworms (P. excavates) | |||
| Pex-DeltaD/Pex-Serrate | Early bidirectional regeneration, regeneration inducing | [67,68,69] | |
| Pex-Notch | Anterior regeneration | ||
| Pex-Hes1A | |||
| Pex-Hes4A | Regenerated head and tail tissues | ||
| Pex-Hes1B | Intact tissues and throughout bidirectional regeneration | ||
| Pex-Hey | |||
| Polychaeta (P. dumerilii) | |||
| Pdu-Deltatv1/Pdu-Deltatv2 | Chaetal sacs and brain cells | Formation of chaetal sacs, proneural and neuronal specification, larval development, posterior growth, and caudal regeneration | [70,71,72,73,74] |
| Pdu-Jagged | Mesodermal posterior stem cells | ||
| Pdu-Notch | Chaetal sacs and brain cells | ||
| Pdu-SuH | Chaetal sacs and brain cells | ||
| Pdu-Presenilin | |||
| Pdu-Hes1-15 | Central and peripheral NS | ||
| Pdu-Twist | Posterior growth zone and in mesodermal anlagen and developing muscles | ||
| Pdu-Nrarp | |||
| Pdu-Numb | |||
| Pdu-Fringe | |||
| Polychaeta (C. teleta) | |||
| CapI-Delta | Terminal growth zone, ventral nerve cord, chaetal sacs | Development of hair follicles, neurogenesis, segmentation | [75,76] |
| CapI-Notch | Chaetal sacs, terminal growth zone, and ventral nerve cord | ||
| CapI-hes1 | Mesoderm | ||
| CapI-hes2 | Brain, pharynx, ventral nerve cord, foregut, and segmental tissue | ||
| CapI-hes3 | Brain, pharynx, ventral nerve cord, foregut, and segmental tissue | ||
| CapI-hesr1 | |||
| CapI-hesr2 | |||
| Clitellata (H. robusta) | |||
| Hro-notch | Both expressed in various cells of the early embryo | Ganglion morphogenesis, posterior elongation and segmentation | [77,78] |
| Hro-hes/Csa-hairy | |||
| Mollusca | |||
| Gastropoda (I. obsoleta) | |||
| IoDelta | Transcripts are widely expressed at the early cleavage stage | embryonic development | [79,80] |
| IoNotch | |||
| IoSuH | |||
| Bivalvia (C. gigas) | |||
| CgDelta/CgJagged CgNotch CgHes1 | Haematopoiesis, transplantation immunity | [81,82,83] | |
| Bivalvia (M. meretrix) | |||
| Delta | Mantle | Shell pigmentation, color patterning | [84] |
| Notch | |||
| Bivalvia (P. f. martensii) | |||
| NOTCH1-3 DTX TBCE | Regulates transplantation immune | [85] | |
| Bivalvia(P. margaritifera) | |||
| Delta Jagged-2 E3 ubiquitin-protein ligase | Pigmentation | [86] | |
| Cephalopoda (D. pealeii) | |||
| Delta/Jagged | Apical side of the retinal epithelium | Essential for maintaining retinal progenitor cell identity | [87,88] |
| DpNotch | Ventral side of the placode and surrounding extraocular tissue | ||
| DpHes-1 | Entire placode | ||
| Arthropoda | |||
| Uniramia (D. melanogaste) | |||
| Serrate/Delta | Ventral nerve cord and within the supraesophogeal ganglia (brain hemispheres) neurogenic regions and intestine | Wing development, embryonic development, regeneration and neurodevelopment | [89,90,91,92,93,94,95,96,97,98,99] |
| Notch | |||
| Psen | Neurons | ||
| Nct | |||
| Aph-1 | |||
| Pen-2 | |||
| Su(H) | Pre-blastoderm embryos | ||
| E(spl) | Embryonic/Larval/pupal eye disk/pupal CNS/Gut/Ovary/pupal leg disk/Larval trachea/pupal wing disk | ||
| Uniramia (T. castaneum) | |||
| E(spl)1/E(spl)3 | Embryonic cephalic and trunk neuroectoderm | Immune defense | [100,101,102] |
| Uniramia (S. maritima) | |||
| StmDelta | Ventral neural ectoderm and the posterior disc | Neural precursor specification and segmented | [103,104] |
| StmNotch | Lateral region of the ventral neural ectoderm | ||
| StmHes1-4 | Posterior disc | ||
| Chelicerata (C. salei) | |||
| CsDelta1/CsDelta2 | Ventral nerve ectodermal neuronal and fragment | Development of ventral neural ectoderm and body segment formation | [105,106] |
| CsNotch | Ventral nerve ectodermal neuronal and fragment | ||
| CsPsn | Prosomal, opisthosomal segments, and growth zone | ||
| CsSu(H)-1/CsSu(H)-2 | Prosomal and opisthosomal segments and growth zone | ||
| CsHairy | Ventral nerve ectodermal neuronal and fragment | ||
| Crustacea (P. vannamei) | |||
| PvDelta | Universal expression | Immune defense | [107,108] |
| PvNotch | |||
| PvCsl | |||
| PvHey2 | |||
| Crustacea (L. vannamei) | |||
| LvNotch | Immune responses and regulates hemocyte proliferation | [109,110] | |
| LvCSL | Universal expression | ||
| LvPsen2 | |||
| LvAph-1 | |||
| LvPsen1 | |||
| LvCtBP | |||
| LvTACE | |||
| Echinodermata | |||
| Echinoidea (H. pulcherrimus) | |||
| HpDelta | Micromere descendants | Specification of SMCs, neurogenesis, involved in sea urchin foregut development, non-skeletogenic mesoderm specification | [111,112] |
| HpNotch | Veg2 blastomeres | ||
| HpSu(H) | Vegetal plate region, the archenteron | ||
| Numb | |||
| Fringe | |||
| Echinoidea (S. purpuratus) | |||
| SpDelta16128/SpSerrate22053 | Embryonic development, neural control Limiting the number of neural progenitor cells and neurons, non-skeletogenic mesoderm (NSM) specification, pigment cell formation, and phagocytosis | [113,114,115,116,117,118] | |
| SpNotch14131 | |||
| SpNotchlike27006/SpNotchlike19810 | |||
| SpPsen | Uniformly in unfertilized eggs, endodermal and mesodermal regions, around the digestive system, neurogenic regions | ||
| SpAph-1 | |||
| SpPen-2 | |||
| SpNicastrin19033 | |||
| SpHes | Mesenchyme blastula stage and the vegetal hemisphere | ||
| Echinoidea (L. variegatus) | |||
| LvDelta | Endoderm, neural progenitor cells of the oral ectoderm, and apical organs | Development of the mesoderm and endoderm, regeneration of tube feet and spines | [119,120,121] |
| LvFringe | Universal expression during germ layer development | ||
| Asteroidea (P. miniata) | |||
| Delta | Mesodermal progenitors of the central vegetal plate, ectoderm | Development of mesoderm and endoderm | [122,123,124] |
| Ophiuroidea (O. brevispina) | |||
| Delta/Serrate ADAM 10/17 Notch RBPJ Nct Psen1 Hes Deltex Notchless | Arm regeneration | [125,126] | |
| Holothuroidea (A. japonicus) | |||
| Serrate/Delta Deltex Fringe Notch MAML Psen Csl CtBP | All expressed in melanocytes | Participation in the pigmentation process of sea cucumbers | [127,128] |
| Chordates | |||
| Ascidiacea (C. intestinalis) | |||
| Delta and Jagged Fringe Notch Neuralized CiSu(H) Hes | Neurodevelopment, regeneration of the oral siphon (OS) | [129,130,131] | |
| Ascidiacea (B. leachii) | |||
| Delta Notch Hairy/E(Spl) Blgroucho1 Blgroucho2 | Whole-body regeneration | [129] | |
| Ascidiacea (H. roretzi) | |||
| HrDelta | Precursor cells of peripheral neurons and palp | Fate determinations of palps and peripheral neurons | [132,133] |
| HrNotch | Epidermal and neural cells |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, Y.; Pang, X.; Cao, Z.; Song, C.; Liu, B.; Wu, W.; Pang, Q. Evolution and Function of the Notch Signaling Pathway: An Invertebrate Perspective. Int. J. Mol. Sci. 2024, 25, 3322. https://doi.org/10.3390/ijms25063322
Lv Y, Pang X, Cao Z, Song C, Liu B, Wu W, Pang Q. Evolution and Function of the Notch Signaling Pathway: An Invertebrate Perspective. International Journal of Molecular Sciences. 2024; 25(6):3322. https://doi.org/10.3390/ijms25063322
Chicago/Turabian StyleLv, Yan, Xuan Pang, Zhonghong Cao, Changping Song, Baohua Liu, Weiwei Wu, and Qiuxiang Pang. 2024. "Evolution and Function of the Notch Signaling Pathway: An Invertebrate Perspective" International Journal of Molecular Sciences 25, no. 6: 3322. https://doi.org/10.3390/ijms25063322
APA StyleLv, Y., Pang, X., Cao, Z., Song, C., Liu, B., Wu, W., & Pang, Q. (2024). Evolution and Function of the Notch Signaling Pathway: An Invertebrate Perspective. International Journal of Molecular Sciences, 25(6), 3322. https://doi.org/10.3390/ijms25063322