
A Review of the Use of Extracellular Vesicles in the Treatment of Neonatal Diseases: Current State and Problems with Translation to the Clinic

 , , ,
, , , 
Abstract
1. Introduction
1.1. Searching Criteria
1.2. Applying the ‘Therapeutic Unit’ Concept to Assess the Equivalence of EVs Concentrations
2. Biogenesis and Classification of EVs
3. The Cargoes of EVs
4. Mechanisms of EVs’ Interaction with Recipient Cell Membranes
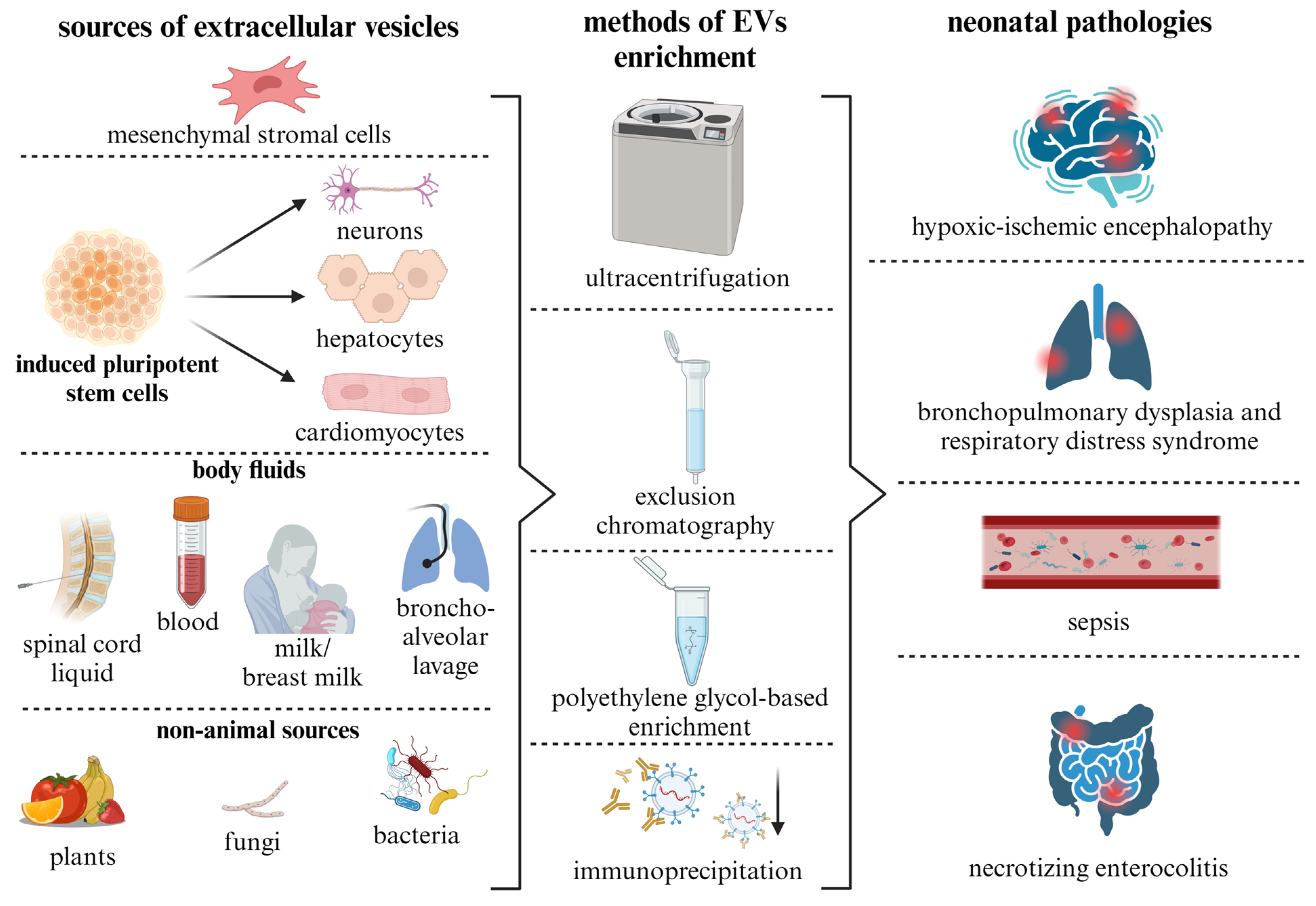
5. Sources of EVs and Methods for Their Isolation
6. Experimental Studies of EVs as Therapeutic Tools for Neonatal Pathologies
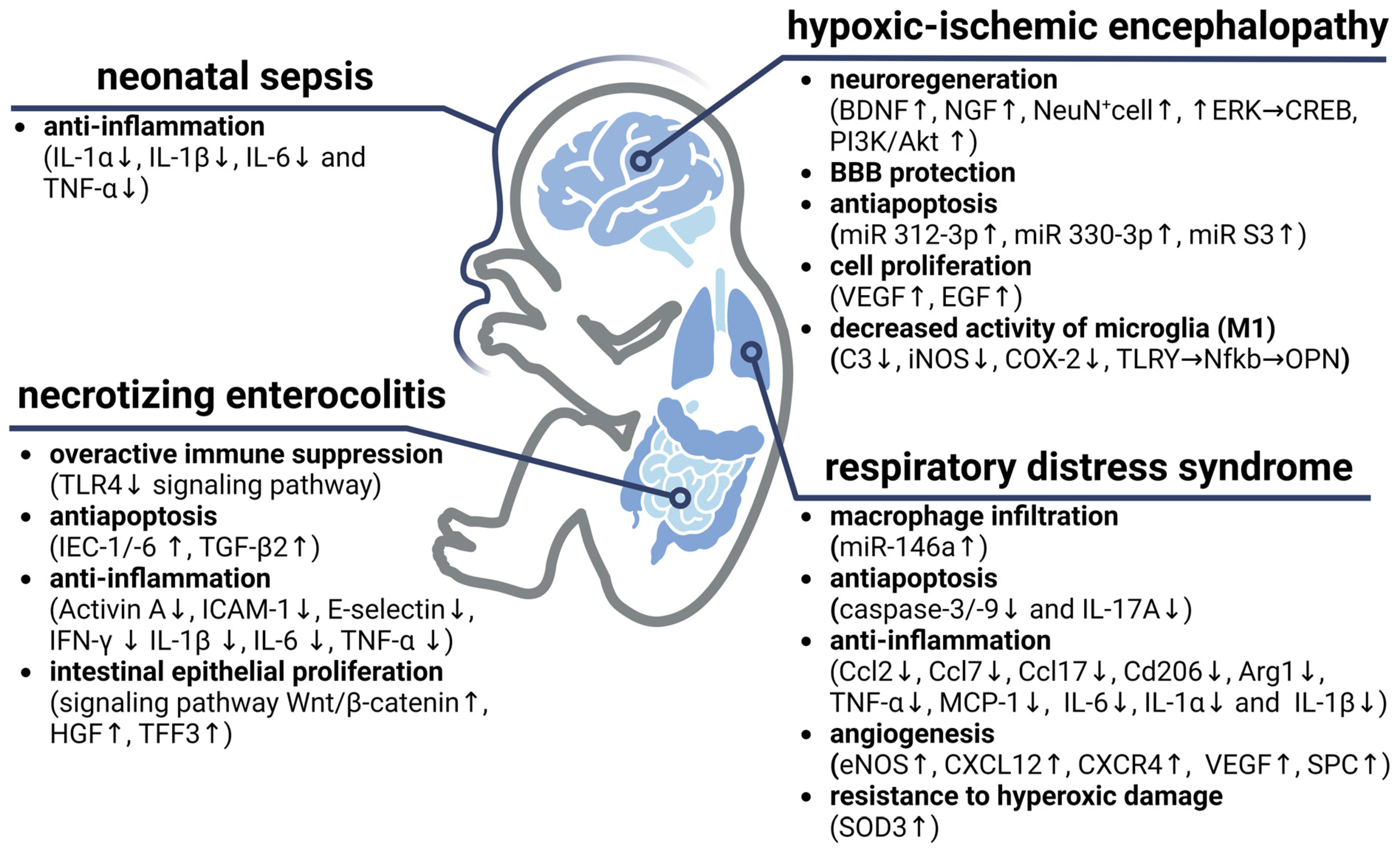
6.1. Hypoxic–Ischemic Encephalopathy
6.2. Respiratory Distress Syndrome and Bronchopulmonary Dysplasia
6.3. Neonatal Sepsis
6.4. Necrotizing Enterocolitis
7. Discussion and Challenges of Translating EVs into Clinical Practice
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vogel, J.P.; Chawanpaiboon, S.; Moller, A.-B.; Watananirun, K.; Bonet, M.; Lumbiganon, P. The Global Epidemiology of Preterm Birth. Best. Pract. Res. Clin. Obs. Gynaecol. 2018, 52, 3–12. [Google Scholar] [CrossRef]
- Belousova, V.; Svitich, O.; Timokhina, E.; Ignatko, I.; Bogomazova, I.; Pesegova, S.; Silaeva, T.; Kuzmina, T.; Skorobogatova, O. Caspase-3, Caspase-8 and XIAP Gene Expression in the Placenta: Exploring the Causes of Spontaneous Preterm Labour. Int. J. Mol. Sci. 2023, 24, 1692. [Google Scholar] [CrossRef]
- Ou, Z.; Yu, D.; Liang, Y.; He, H.; He, W.; Li, Y.; Zhang, M.; Gao, Y.; Wu, F.; Chen, Q. Global Trends in Incidence and Death of Neonatal Disorders and Its Specific Causes in 204 Countries/Territories during 1990–2019. BMC Public Health 2022, 22, 360. [Google Scholar] [CrossRef]
- Malhotra, A.; Thebaud, B.; Paton, M.C.B.; Fleiss, B.; Papagianis, P.; Baker, E.; Bennet, L.; Yawno, T.; Elwood, N.; Campbell, B.; et al. Advances in Neonatal Cell Therapies: Proceedings of the First Neonatal Cell Therapies Symposium (2022). Pediatr. Res. 2023, 94, 1631–1638. [Google Scholar] [CrossRef] [PubMed]
- Schiller, E.A.; Cohen, K.; Lin, X.; El-Khawam, R.; Hanna, N. Extracellular Vesicle-microRNAs as Diagnostic Biomarkers in Preterm Neonates. Int. J. Mol. Sci. 2023, 24, 2622. [Google Scholar] [CrossRef]
- Nitkin, C.R.; Rajasingh, J.; Pisano, C.; Besner, G.E.; Thébaud, B.; Sampath, V. Stem Cell Therapy for Preventing Neonatal Diseases in the 21st Century: Current Understanding and Challenges. Pediatr. Res. 2020, 87, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Leopoldino, R.D.; Santos, M.T.; Costa, T.X.; Martins, R.R.; Oliveira, A.G. Drug Related Problems in the Neonatal Intensive Care Unit: Incidence, Characterization and Clinical Relevance. BMC Pediatr. 2019, 19, 134. [Google Scholar] [CrossRef]
- Galderisi, U.; Peluso, G.; Di Bernardo, G. Clinical Trials Based on Mesenchymal Stromal Cells Are Exponentially Increasing: Where Are We in Recent Years? Stem Cell Rev. Rep. 2022, 18, 23–36. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Park, W.S.; Sung, S.I.; Chang, Y.S. Mesenchymal Stem Cell Therapy for Intractable Neonatal Disorders. Pediatr. Neonatol. 2021, 62 (Suppl. S1), S16–S21. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, V.V.; Patel, N.H.; Smith, E.E.; Barnes, C.L.; Gustafson, M.P.; Rao, R.R.; Samsonraj, R.M. Clinical Utility of Mesenchymal Stem/Stromal Cells in Regenerative Medicine and Cellular Therapy. J. Biol. Eng. 2023, 17, 44. [Google Scholar] [CrossRef] [PubMed]
- EL Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J.A. Extracellular Vesicles: Biology and Emerging Therapeutic Opportunities. Nat. Rev. Drug Discov. 2013, 12, 347–357. [Google Scholar] [CrossRef]
- Elsharkasy, O.M.; Nordin, J.Z.; Hagey, D.W.; de Jong, O.G.; Schiffelers, R.M.; Andaloussi, S.E.; Vader, P. Extracellular Vesicles as Drug Delivery Systems: Why and How? Adv. Drug Deliv. Rev. 2020, 159, 332–343. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Kordelas, L.; Rebmann, V.; Ludwig, A.-K.; Radtke, S.; Ruesing, J.; Doeppner, T.R.; Epple, M.; Horn, P.A.; Beelen, D.W.; Giebel, B. MSC-Derived Exosomes: A Novel Tool to Treat Therapy-Refractory Graft-versus-Host Disease. Leukemia 2014, 28, 970–973. [Google Scholar] [CrossRef] [PubMed]
- Wolf, P. The Nature and Significance of Platelet Products in Human Plasma. Br. J. Haematol. 1967, 13, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Amarasinghe, I.; Phillips, W.; Hill, A.F.; Cheng, L.; Helbig, K.J.; Willms, E.; Monson, E.A. Cellular Communication through Extracellular Vesicles and Lipid Droplets. J. Extracell. Biol. 2023, 2, e77. [Google Scholar] [CrossRef]
- Stahl, P.D.; Raposo, G. Extracellular Vesicles: Exosomes and Microvesicles, Integrators of Homeostasis. Physiology 2019, 34, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Gandham, S.; Su, X.; Wood, J.; Nocera, A.L.; Alli, S.C.; Milane, L.; Zimmerman, A.; Amiji, M.; Ivanov, A.R. Technologies and Standardization in Research on Extracellular Vesicles. Trends Biotechnol. 2020, 38, 1066–1098. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Samuel, M.; Mathivanan, S. Exomeres: A New Member of Extracellular Vesicles Family. Subcell. Biochem. 2021, 97, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Jeppesen, D.K.; Higginbotham, J.N.; Graves-Deal, R.; Trinh, V.Q.; Ramirez, M.A.; Sohn, Y.; Neininger, A.C.; Taneja, N.; McKinley, E.T.; et al. Supermeres Are Functional Extracellular Nanoparticles Replete with Disease Biomarkers and Therapeutic Targets. Nat. Cell Biol. 2021, 23, 1240–1254. [Google Scholar] [CrossRef]
- Laulagnier, K.; Motta, C.; Hamdi, S.; Roy, S.; Fauvelle, F.; Pageaux, J.-F.; Kobayashi, T.; Salles, J.-P.; Perret, B.; Bonnerot, C.; et al. Mast Cell- and Dendritic Cell-Derived Exosomes Display a Specific Lipid Composition and an Unusual Membrane Organization. Biochem. J. 2004, 380, 161–171. [Google Scholar] [CrossRef]
- Biró, É.; Akkerman, J.W.N.; Hoek, F.J.; Gorter, G.; Pronk, L.M.; Sturk, A.; Nieuwland, R. The Phospholipid Composition and Cholesterol Content of Platelet-Derived Microparticles: A Comparison with Platelet Membrane Fractions. J. Thromb. Haemost. 2005, 3, 2754–2763. [Google Scholar] [CrossRef]
- Zorova, L.D.; Kovalchuk, S.I.; Popkov, V.A.; Chernikov, V.P.; Zharikova, A.A.; Khutornenko, A.A.; Zorov, S.D.; Plokhikh, K.S.; Zinovkin, R.A.; Evtushenko, E.A.; et al. Do Extracellular Vesicles Derived from Mesenchymal Stem Cells Contain Functional Mitochondria? Int. J. Mol. Sci. 2022, 23, 7408. [Google Scholar] [CrossRef] [PubMed]
- Claridge, B.; Lozano, J.; Poh, Q.H.; Greening, D.W. Development of Extracellular Vesicle Therapeutics: Challenges, Considerations, and Opportunities. Front. Cell Dev. Biol. 2021, 9, 734720. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-K.; Kang, B.; Kim, O.Y.; Choi, D.; Lee, J.; Kim, S.R.; Go, G.; Yoon, Y.J.; Kim, J.H.; Jang, S.C.; et al. EVpedia: An Integrated Database of High-Throughput Data for Systemic Analyses of Extracellular Vesicles. J. Extracell. Vesicles 2013, 2, 20384. [Google Scholar] [CrossRef] [PubMed]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; et al. ExoCarta: A Web-Based Compendium of Exosomal Cargo. J. Mol. Biol. 2016, 428, 688–692. [Google Scholar] [CrossRef] [PubMed]
- Pathan, M.; Fonseka, P.; Chitti, S.V.; Kang, T.; Sanwlani, R.; Van Deun, J.; Hendrix, A.; Mathivanan, S. Vesiclepedia 2019: A Compendium of RNA, Proteins, Lipids and Metabolites in Extracellular Vesicles. Nucleic Acids Res. 2019, 47, D516–D519. [Google Scholar] [CrossRef]
- Anderson, J.D.; Johansson, H.J.; Graham, C.S.; Vesterlund, M.; Pham, M.T.; Bramlett, C.S.; Montgomery, E.N.; Mellema, M.S.; Bardini, R.L.; Contreras, Z.; et al. Comprehensive Proteomic Analysis of Mesenchymal Stem Cell Exosomes Reveals Modulation of Angiogenesis via Nuclear Factor-KappaB Signaling. Stem Cells 2016, 34, 601–613. [Google Scholar] [CrossRef]
- Qiu, G.; Zheng, G.; Ge, M.; Wang, J.; Huang, R.; Shu, Q.; Xu, J. Functional Proteins of Mesenchymal Stem Cell-Derived Extracellular Vesicles. Stem Cell Res. Ther. 2019, 10, 359. [Google Scholar] [CrossRef]
- Turovsky, E.A.; Golovicheva, V.V.; Varlamova, E.G.; Danilina, T.I.; Goryunov, K.V.; Shevtsova, Y.A.; Pevzner, I.B.; Zorova, L.D.; Babenko, V.A.; Evtushenko, E.A.; et al. Mesenchymal Stromal Cell-Derived Extracellular Vesicles Afford Neuroprotection by Modulating PI3K/AKT Pathway and Calcium Oscillations. Int. J. Biol. Sci. 2022, 18, 5345–5368. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Sung, D.K.; Kim, Y.E.; Sung, S.; Chang, Y.S.; Park, W.S. Brain-Derived Neurotropic Factor Mediates Neuroprotection of Mesenchymal Stem Cell-Derived Extracellular Vesicles against Severe Intraventricular Hemorrhage in Newborn Rats. Stem Cells Transl. Med. 2021, 10, 374–384. [Google Scholar] [CrossRef]
- Harrell, C.R.; Fellabaum, C.; Jovicic, N.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Molecular Mechanisms Responsible for Therapeutic Potential of Mesenchymal Stem Cell-Derived Secretome. Cells 2019, 8, 467. [Google Scholar] [CrossRef]
- Yari, H.; Mikhailova, M.V.; Mardasi, M.; Jafarzadehgharehziaaddin, M.; Shahrokh, S.; Thangavelu, L.; Ahmadi, H.; Shomali, N.; Yaghoubi, Y.; Zamani, M.; et al. Emerging Role of Mesenchymal Stromal Cells (MSCs)-Derived Exosome in Neurodegeneration-Associated Conditions: A Groundbreaking Cell-Free Approach. Stem Cell Res. Ther. 2022, 13, 423. [Google Scholar] [CrossRef] [PubMed]
- Kou, X.; Xu, X.; Chen, C.; Sanmillan, M.L.; Cai, T.; Zhou, Y.; Giraudo, C.; Le, A.; Shi, S. The Fas/Fap-1/Cav-1 Complex Regulates IL-1RA Secretion in Mesenchymal Stem Cells to Accelerate Wound Healing. Sci. Transl. Med. 2018, 10, eaai8524. [Google Scholar] [CrossRef]
- Riazifar, M.; Mohammadi, M.R.; Pone, E.J.; Yeri, A.; Lässer, C.; Segaliny, A.I.; McIntyre, L.L.; Shelke, G.V.; Hutchins, E.; Hamamoto, A.; et al. Stem Cell-Derived Exosomes as Nanotherapeutics for Autoimmune and Neurodegenerative Disorders. ACS Nano 2019, 13, 6670–6688. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Xiong, Y.; Li, Q.; Han, M.; Shan, D.; Yang, G.; Zhang, S.; Xin, D.; Zhao, R.; Wang, Z.; et al. Extracellular Vesicle-Mediated Transfer of miR-21-5p from Mesenchymal Stromal Cells to Neurons Alleviates Early Brain Injury to Improve Cognitive Function via the PTEN/Akt Pathway after Subarachnoid Hemorrhage. Cell Death Dis. 2020, 11, 363. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Li, Y.; Liu, Z.; Wang, X.; Shang, X.; Cui, Y.; Zhang, Z.G.; Chopp, M. MiR-133b Promotes Neural Plasticity and Functional Recovery after Treatment of Stroke with Multipotent Mesenchymal Stromal Cells in Rats via Transfer of Exosome-Enriched Extracellular Particles. Stem Cells 2013, 31, 2737–2746. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, J.; Su, M.; Wang, X.; Xie, C. Exosomal microRNA-22-3p Alleviates Cerebral Ischemic Injury by Modulating KDM6B/BMP2/BMF Axis. Stem Cell Res. Ther. 2021, 12, 111. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Li, J.; Che, Y. miR-31 from Adipose Stem Cell-Derived Extracellular Vesicles Promotes Recovery of Neurological Function after Ischemic Stroke by Inhibiting TRAF6 and IRF5. Exp. Neurol. 2021, 342, 113611. [Google Scholar] [CrossRef]
- Zhang, Z.; Zou, X.; Zhang, R.; Xie, Y.; Feng, Z.; Li, F.; Han, J.; Sun, H.; Ouyang, Q.; Hua, S.; et al. Human Umbilical Cord Mesenchymal Stem Cell-Derived Exosomal miR-146a-5p Reduces Microglial-Mediated Neuroinflammation via Suppression of the IRAK1/TRAF6 Signaling Pathway after Ischemic Stroke. Aging 2021, 13, 3060–3079. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological Properties of Extracellular Vesicles and Their Physiological Functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef]
- Lu, M.; Shao, W.; Xing, H.; Huang, Y. Extracellular Vesicle-Based Nucleic Acid Delivery. Interdiscip. Med. 2023, 1, e20220007. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The Biogenesis, Biology and Characterization of Circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef]
- Zhuo, C.-J.; Hou, W.-H.; Jiang, D.-G.; Tian, H.-J.; Wang, L.-N.; Jia, F.; Zhou, C.-H.; Zhu, J.-J. Circular RNAs in Early Brain Development and Their Influence and Clinical Significance in Neuropsychiatric Disorders. Neural Regen. Res. 2020, 15, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Chao, J.; Yao, H. Circular RNA and Its Mechanisms in Disease: From the Bench to the Clinic. Pharmacol. Ther. 2018, 187, 31–44. [Google Scholar] [CrossRef]
- Mahmoudi, E.; Fitzsimmons, C.; Geaghan, M.P.; Shannon Weickert, C.; Atkins, J.R.; Wang, X.; Cairns, M.J. Circular RNA Biogenesis Is Decreased in Postmortem Cortical Gray Matter in Schizophrenia and May Alter the Bioavailability of Associated miRNA. Neuropsychopharmacology 2019, 44, 1043–1054. [Google Scholar] [CrossRef]
- Wapinski, O.; Chang, H.Y. Long Noncoding RNAs and Human Disease. Trends Cell Biol. 2011, 21, 354–361. [Google Scholar] [CrossRef]
- Kino, T.; Hurt, D.E.; Ichijo, T.; Nader, N.; Chrousos, G.P. Noncoding RNA Gas5 Is a Growth Arrest- and Starvation-Associated Repressor of the Glucocorticoid Receptor. Sci. Signal. 2010, 3, ra8. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Gerin, I.; Miao, H.; Vu-Phan, D.; Johnson, C.N.; Xu, R.; Chen, X.-W.; Cawthorn, W.P.; MacDougald, O.A.; Koenig, R.J. Multiple Roles for the Non-Coding RNA SRA in Regulation of Adipogenesis and Insulin Sensitivity. PLoS ONE 2010, 5, e14199. [Google Scholar] [CrossRef]
- Ellis, B.C.; Graham, L.D.; Molloy, P.L. CRNDE, a Long Non-Coding RNA Responsive to Insulin/IGF Signaling, Regulates Genes Involved in Central Metabolism. Biochim. Biophys. Acta 2014, 1843, 372–386. [Google Scholar] [CrossRef]
- Tong, X.; Gu, P.; Xu, S.; Lin, X. Long Non-Coding RNA-DANCR in Human Circulating Monocytes: A Potential Biomarker Associated with Postmenopausal Osteoporosis. Biosci. Biotechnol. Biochem. 2015, 79, 732–737. [Google Scholar] [CrossRef]
- Patel, N.A.; Moss, L.D.; Lee, J.-Y.; Tajiri, N.; Acosta, S.; Hudson, C.; Parag, S.; Cooper, D.R.; Borlongan, C.V.; Bickford, P.C. Long Noncoding RNA MALAT1 in Exosomes Drives Regenerative Function and Modulates Inflammation-Linked Networks Following Traumatic Brain Injury. J. Neuroinflamm. 2018, 15, 204. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA Delivery by Extracellular Vesicles in Mammalian Cells and Its Applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef] [PubMed]
- De Luca, L.; Trino, S.; Laurenzana, I.; Simeon, V.; Calice, G.; Raimondo, S.; Podestà, M.; Santodirocco, M.; Di Mauro, L.; La Rocca, F.; et al. MiRNAs and piRNAs from Bone Marrow Mesenchymal Stem Cell Extracellular Vesicles Induce Cell Survival and Inhibit Cell Differentiation of Cord Blood Hematopoietic Stem Cells: A New Insight in Transplantation. Oncotarget 2016, 7, 6676–6692. [Google Scholar] [CrossRef] [PubMed]
- Wolin, S.L.; Steitz, J.A. The Ro Small Cytoplasmic Ribonucleoproteins: Identification of the Antigenic Protein and Its Binding Site on the Ro RNAs. Proc. Natl. Acad. Sci. USA 1984, 81, 1996–2000. [Google Scholar] [CrossRef] [PubMed]
- Valkov, N.; Das, S. Y RNAs: Biogenesis, Function and Implications for the Cardiovascular System. Adv. Exp. Med. Biol. 2020, 1229, 327–342. [Google Scholar] [CrossRef] [PubMed]
- Haga, H.; Yan, I.K.; Takahashi, K.; Matsuda, A.; Patel, T. Extracellular Vesicles from Bone Marrow-Derived Mesenchymal Stem Cells Improve Survival from Lethal Hepatic Failure in Mice. Stem Cells Transl. Med. 2017, 6, 1262–1272. [Google Scholar] [CrossRef]
- Cambier, L.; Giani, J.F.; Liu, W.; Ijichi, T.; Echavez, A.K.; Valle, J.; Marbán, E. Angiotensin II-Induced End-Organ Damage in Mice Is Attenuated by Human Exosomes and by an Exosomal Y RNA Fragment. Hypertension 2018, 72, 370–380. [Google Scholar] [CrossRef]
- Zakharova, L.; Svetlova, M.; Fomina, A.F. T Cell Exosomes Induce Cholesterol Accumulation in Human Monocytes via Phosphatidylserine Receptor. J. Cell. Physiol. 2007, 212, 174–181. [Google Scholar] [CrossRef]
- Record, M.; Carayon, K.; Poirot, M.; Silvente-Poirot, S. Exosomes as New Vesicular Lipid Transporters Involved in Cell-Cell Communication and Various Pathophysiologies. Biochim. Biophys. Acta 2014, 1841, 108–120. [Google Scholar] [CrossRef]
- Matsuo, H.; Chevallier, J.; Mayran, N.; Le Blanc, I.; Ferguson, C.; Fauré, J.; Blanc, N.S.; Matile, S.; Dubochet, J.; Sadoul, R.; et al. Role of LBPA and Alix in Multivesicular Liposome Formation and Endosome Organization. Science 2004, 303, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Morel, O.; Jesel, L.; Freyssinet, J.-M.; Toti, F. Cellular Mechanisms Underlying the Formation of Circulating Microparticles. Arter. Thromb. Vasc. Biol. 2011, 31, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Bianco, F.; Perrotta, C.; Novellino, L.; Francolini, M.; Riganti, L.; Menna, E.; Saglietti, L.; Schuchman, E.H.; Furlan, R.; Clementi, E.; et al. Acid Sphingomyelinase Activity Triggers Microparticle Release from Glial Cells. EMBO J. 2009, 28, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory Mechanisms and Intercellular Transfer of microRNAs in Living Cells. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar] [CrossRef]
- Ghossoub, R.; Lembo, F.; Rubio, A.; Gaillard, C.B.; Bouchet, J.; Vitale, N.; Slavík, J.; Machala, M.; Zimmermann, P. Syntenin-ALIX Exosome Biogenesis and Budding into Multivesicular Bodies Are Controlled by ARF6 and PLD2. Nat. Commun. 2014, 5, 3477. [Google Scholar] [CrossRef]
- Record, M.; Silvente-Poirot, S.; Poirot, M.; Wakelam, M.J.O. Extracellular Vesicles: Lipids as Key Components of Their Biogenesis and Functions. J. Lipid Res. 2018, 59, 1316–1324. [Google Scholar] [CrossRef]
- Ghadami, S.; Dellinger, K. The Lipid Composition of Extracellular Vesicles: Applications in Diagnostics and Therapeutic Delivery. Front. Mol. Biosci. 2023, 10, 1198044. [Google Scholar] [CrossRef]
- Lobasso, S.; Tanzarella, P.; Mannavola, F.; Tucci, M.; Silvestris, F.; Felici, C.; Ingrosso, C.; Corcelli, A.; Lopalco, P. A Lipidomic Approach to Identify Potential Biomarkers in Exosomes From Melanoma Cells With Different Metastatic Potential. Front. Physiol. 2021, 12, 748895. [Google Scholar] [CrossRef]
- Esser, J.; Gehrmann, U.; D’Alexandri, F.L.; Hidalgo-Estévez, A.M.; Wheelock, C.E.; Scheynius, A.; Gabrielsson, S.; Rådmark, O. Exosomes from Human Macrophages and Dendritic Cells Contain Enzymes for Leukotriene Biosynthesis and Promote Granulocyte Migration. J. Allergy Clin. Immunol. 2010, 126, 1032–1040.e4. [Google Scholar] [CrossRef]
- Holopainen, M.; Colas, R.A.; Valkonen, S.; Tigistu-Sahle, F.; Hyvärinen, K.; Mazzacuva, F.; Lehenkari, P.; Käkelä, R.; Dalli, J.; Kerkelä, E.; et al. Polyunsaturated Fatty Acids Modify the Extracellular Vesicle Membranes and Increase the Production of Proresolving Lipid Mediators of Human Mesenchymal Stromal Cells. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 1350–1362. [Google Scholar] [CrossRef]
- Pizzinat, N.; Ong-Meang, V.; Bourgailh-Tortosa, F.; Blanzat, M.; Perquis, L.; Cussac, D.; Parini, A.; Poinsot, V. Extracellular Vesicles of MSCs and Cardiomyoblasts Are Vehicles for Lipid Mediators. Biochimie 2020, 178, 69–80. [Google Scholar] [CrossRef]
- Gabrielli, M.; Battista, N.; Riganti, L.; Prada, I.; Antonucci, F.; Cantone, L.; Matteoli, M.; Maccarrone, M.; Verderio, C. Active Endocannabinoids Are Secreted on Extracellular Membrane Vesicles. EMBO Rep. 2015, 16, 213–220. [Google Scholar] [CrossRef]
- Nan, W.; Zhang, C.; Wang, H.; Chen, H.; Ji, S. Direct Modification of Extracellular Vesicles and Its Applications for Cancer Therapy: A Mini-Review. Front. Chem. 2022, 10, 910341. [Google Scholar] [CrossRef]
- Susa, F.; Limongi, T.; Borgione, F.; Peiretti, S.; Vallino, M.; Cauda, V.; Pisano, R. Comparative Studies of Different Preservation Methods and Relative Freeze-Drying Formulations for Extracellular Vesicle Pharmaceutical Applications. ACS Biomater. Sci. Eng. 2023, 9, 5871–5885. [Google Scholar] [CrossRef]
- Trenkenschuh, E.; Richter, M.; Heinrich, E.; Koch, M.; Fuhrmann, G.; Friess, W. Enhancing the Stabilization Potential of Lyophilization for Extracellular Vesicles. Adv. Healthc. Mater. 2022, 11, e2100538. [Google Scholar] [CrossRef]
- Gurung, S.; Perocheau, D.; Touramanidou, L.; Baruteau, J. The Exosome Journey: From Biogenesis to Uptake and Intracellular Signalling. Cell Commun. Signal. 2021, 19, 47. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, A.; Tahara, Y.; Sawada, S.-I.; Sasaki, Y.; Akiyoshi, K. Glycan Profiling Analysis Using Evanescent-Field Fluorescence-Assisted Lectin Array: Importance of Sugar Recognition for Cellular Uptake of Exosomes from Mesenchymal Stem Cells. Biochem. Biophys. Res. Commun. 2017, 491, 701–707. [Google Scholar] [CrossRef]
- Tkach, M.; Kowal, J.; Zucchetti, A.E.; Enserink, L.; Jouve, M.; Lankar, D.; Saitakis, M.; Martin-Jaular, L.; Théry, C. Qualitative Differences in T-Cell Activation by Dendritic Cell-Derived Extracellular Vesicle Subtypes. EMBO J. 2017, 36, 3012–3028. [Google Scholar] [CrossRef] [PubMed]
- Munich, S.; Sobo-Vujanovic, A.; Buchser, W.J.; Beer-Stolz, D.; Vujanovic, N.L. Dendritic Cell Exosomes Directly Kill Tumor Cells and Activate Natural Killer Cells via TNF Superfamily Ligands. Oncoimmunology 2012, 1, 1074–1083. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J.A. Delivery of siRNA to the Mouse Brain by Systemic Injection of Targeted Exosomes. Nat. Biotechnol. 2011, 29, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.-H.; Guo, H.-D.; Li, H.; Zhai, Y.; Gong, Z.-B.; Wu, J.; Liu, J.-S.; Dong, Y.-R.; Hou, S.-X.; Liu, J.-R. RVG-Modified Exosomes Derived from Mesenchymal Stem Cells Rescue Memory Deficits by Regulating Inflammatory Responses in a Mouse Model of Alzheimer’s Disease. Immun. Ageing 2019, 16, 10. [Google Scholar] [CrossRef]
- Murphy, D.E.; de Jong, O.G.; Brouwer, M.; Wood, M.J.; Lavieu, G.; Schiffelers, R.M.; Vader, P. Extracellular Vesicle-Based Therapeutics: Natural versus Engineered Targeting and Trafficking. Exp. Mol. Med. 2019, 51, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, A.; Alini, M.; Baghaban Eslaminejad, M.; Hosseini, S. Engineering Strategies for Customizing Extracellular Vesicle Uptake in a Therapeutic Context. Stem Cell Res. Ther. 2022, 13, 129. [Google Scholar] [CrossRef] [PubMed]
- Yuana, Y.; Sturk, A.; Nieuwland, R. Extracellular Vesicles in Physiological and Pathological Conditions. Blood Rev. 2013, 27, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; He, B.; Wang, S.; Fletcher, S.; Niu, D.; Mitter, N.; Birch, P.R.J.; Jin, H. Message in a Bubble: Shuttling Small RNAs and Proteins Between Cells and Interacting Organisms Using Extracellular Vesicles. Annu. Rev. Plant Biol. 2021, 72, 497–524. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Cerione, R.A.; Antonyak, M.A. Isolation and Characterization of Extracellular Vesicles Produced by Cell Lines. STAR Protoc. 2021, 2, 100295. [Google Scholar] [CrossRef]
- Bost, J.P.; Saher, O.; Hagey, D.; Mamand, D.R.; Liang, X.; Zheng, W.; Corso, G.; Gustafsson, O.; Görgens, A.; Smith, C.E.; et al. Growth Media Conditions Influence the Secretion Route and Release Levels of Engineered Extracellular Vesicles. Adv. Healthc. Mater. 2022, 11, e2101658. [Google Scholar] [CrossRef]
- Tung, S.; Delavogia, E.; Fernandez-Gonzalez, A.; Mitsialis, S.A.; Kourembanas, S. Harnessing the Therapeutic Potential of the Stem Cell Secretome in Neonatal Diseases. Semin. Perinatol. 2023, 47, 151730. [Google Scholar] [CrossRef]
- Gowen, A.; Shahjin, F.; Chand, S.; Odegaard, K.E.; Yelamanchili, S.V. Mesenchymal Stem Cell-Derived Extracellular Vesicles: Challenges in Clinical Applications. Front. Cell Dev. Biol. 2020, 8, 149. [Google Scholar] [CrossRef]
- Sanz-Ros, J.; Mas-Bargues, C.; Romero-García, N.; Huete-Acevedo, J.; Dromant, M.; Borrás, C. Extracellular Vesicles as Therapeutic Resources in the Clinical Environment. Int. J. Mol. Sci. 2023, 24, 2344. [Google Scholar] [CrossRef]
- Bond, S.T.; Calkin, A.C.; Drew, B.G. Adipose-Derived Extracellular Vesicles: Systemic Messengers and Metabolic Regulators in Health and Disease. Front. Physiol. 2022, 13, 837001. [Google Scholar] [CrossRef] [PubMed]
- Willis, G.R.; Reis, M.; Gheinani, A.H.; Fernandez-Gonzalez, A.; Taglauer, E.S.; Yeung, V.; Liu, X.; Ericsson, M.; Haas, E.; Mitsialis, S.A.; et al. Extracellular Vesicles Protect the Neonatal Lung from Hyperoxic Injury through the Epigenetic and Transcriptomic Reprogramming of Myeloid Cells. Am. J. Respir. Crit. Care Med. 2021, 204, 1418–1432. [Google Scholar] [CrossRef] [PubMed]
- Thomi, G.; Surbek, D.; Haesler, V.; Joerger-Messerli, M.; Schoeberlein, A. Exosomes Derived from Umbilical Cord Mesenchymal Stem Cells Reduce Microglia-Mediated Neuroinflammation in Perinatal Brain Injury. Stem Cell Res. Ther. 2019, 10, 105. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Chen, Q.; Wang, F. Mesenchymal Stem Cells for the Treatment of Ulcerative Colitis: A Systematic Review and Meta-Analysis of Experimental and Clinical Studies. Stem Cell Res. Ther. 2019, 10, 266. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, J.; Liu, C.-Y.; Wang, I.-J.; Sieber, M.; Chang, J.; Jester, J.V.; Kao, W.W.Y. Cell Therapy of Congenital Corneal Diseases with Umbilical Mesenchymal Stem Cells: Lumican Null Mice. PLoS ONE 2010, 5, e10707. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, E.; van der Poll, T.; DelaRosa, O.; Dalemans, W. Mesenchymal Stem Cells as a Therapeutic Tool to Treat Sepsis. World J. Stem Cells 2015, 7, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Joerger-Messerli, M.S.; Oppliger, B.; Spinelli, M.; Thomi, G.; di Salvo, I.; Schneider, P.; Schoeberlein, A. Extracellular Vesicles Derived from Wharton’s Jelly Mesenchymal Stem Cells Prevent and Resolve Programmed Cell Death Mediated by Perinatal Hypoxia-Ischemia in Neuronal Cells. Cell Transpl. 2018, 27, 168–180. [Google Scholar] [CrossRef]
- Ma, Y.; Dong, L.; Zhou, D.; Li, L.; Zhang, W.; Zhen, Y.; Wang, T.; Su, J.; Chen, D.; Mao, C.; et al. Extracellular Vesicles from Human Umbilical Cord Mesenchymal Stem Cells Improve Nerve Regeneration after Sciatic Nerve Transection in Rats. J. Cell. Mol. Med. 2019, 23, 2822–2835. [Google Scholar] [CrossRef]
- Aiello, S.; Rocchetta, F.; Longaretti, L.; Faravelli, S.; Todeschini, M.; Cassis, L.; Pezzuto, F.; Tomasoni, S.; Azzollini, N.; Mister, M.; et al. Extracellular Vesicles Derived from T Regulatory Cells Suppress T Cell Proliferation and Prolong Allograft Survival. Sci. Rep. 2017, 7, 11518. [Google Scholar] [CrossRef]
- Alarcón-Vila, C.; Baroja-Mazo, A.; de Torre-Minguela, C.; Martínez, C.M.; Martínez-García, J.J.; Martínez-Banaclocha, H.; García-Palenciano, C.; Pelegrin, P. CD14 Release Induced by P2X7 Receptor Restricts Inflammation and Increases Survival during Sepsis. Elife 2020, 9, e60849. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, P.; Goodwin, A.J.; Cook, J.A.; Halushka, P.V.; Chang, E.; Fan, H. Exosomes from Endothelial Progenitor Cells Improve the Outcome of a Murine Model of Sepsis. Mol. Ther. 2018, 26, 1375–1384. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, C.V.; Liaw, N.Y.; Gunadasa-Rohling, M.; Matthaei, M.; Braga, L.; Kennedy, T.; Salinas, G.; Voigt, N.; Giacca, M.; Zimmermann, W.-H.; et al. Regenerative Potential of Epicardium-Derived Extracellular Vesicles Mediated by Conserved miRNA Transfer. Cardiovasc. Res. 2022, 118, 597–611. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Maimaitiaili, R.; Yao, J.; Xie, Y.; Qiang, S.; Hu, F.; Li, X.; Shi, C.; Jia, P.; Yang, H.; et al. Percutaneous Intracoronary Delivery of Plasma Extracellular Vesicles Protects the Myocardium Against Ischemia-Reperfusion Injury in Canis. Hypertension 2021, 78, 1541–1554. [Google Scholar] [CrossRef] [PubMed]
- Upadhya, R.; Madhu, L.N.; Attaluri, S.; Gitaí, D.L.G.; Pinson, M.R.; Kodali, M.; Shetty, G.; Zanirati, G.; Kumar, S.; Shuai, B.; et al. Extracellular Vesicles from Human iPSC-Derived Neural Stem Cells: miRNA and Protein Signatures, and Anti-Inflammatory and Neurogenic Properties. J. Extracell. Vesicles 2020, 9, 1809064. [Google Scholar] [CrossRef]
- Park, H.J.; Jeon, J.; Choi, J.; Kim, J.Y.; Kim, H.S.; Huh, J.Y.; Goldman, S.A.; Song, J. Human iPSC-Derived Neural Precursor Cells Differentiate into Multiple Cell Types to Delay Disease Progression Following Transplantation into YAC128 Huntington’s Disease Mouse Model. Cell Prolif. 2021, 54, e13082. [Google Scholar] [CrossRef]
- Attaluri, S.; Jaimes Gonzalez, J.; Kirmani, M.; Vogel, A.D.; Upadhya, R.; Kodali, M.; Madhu, L.N.; Rao, S.; Shuai, B.; Babu, R.S.; et al. Intranasally Administered Extracellular Vesicles from Human Induced Pluripotent Stem Cell-Derived Neural Stem Cells Quickly Incorporate into Neurons and Microglia in 5xFAD Mice. Front. Aging Neurosci. 2023, 15, 1200445. [Google Scholar] [CrossRef]
- McCulloh, C.J.; Olson, J.K.; Wang, Y.; Zhou, Y.; Tengberg, N.H.; Deshpande, S.; Besner, G.E. Treatment of Experimental Necrotizing Enterocolitis with Stem Cell-Derived Exosomes. J. Pediatr. Surg. 2018, 53, 1215–1220. [Google Scholar] [CrossRef]
- Lou, P.; Liu, S.; Wang, Y.; Lv, K.; Zhou, X.; Li, L.; Zhang, Y.; Chen, Y.; Cheng, J.; Lu, Y.; et al. Neonatal-Tissue-Derived Extracellular Vesicle Therapy (NEXT): A Potent Strategy for Precision Regenerative Medicine. Adv. Mater. 2023, 35, e2300602. [Google Scholar] [CrossRef]
- Babenko, V.A.; Silachev, D.N.; Danilina, T.I.; Goryunov, K.V.; Pevzner, I.B.; Zorova, L.D.; Popkov, V.A.; Chernikov, V.P.; Plotnikov, E.Y.; Sukhikh, G.T.; et al. Age-Related Changes in Bone-Marrow Mesenchymal Stem Cells. Cells 2021, 10, 1273. [Google Scholar] [CrossRef]
- Chutipongtanate, S.; Morrow, A.L.; Newburg, D.S. Human Milk Extracellular Vesicles: A Biological System with Clinical Implications. Cells 2022, 11, 2345. [Google Scholar] [CrossRef]
- Bellio, M.A.; Young, K.C.; Milberg, J.; Santos, I.; Abdullah, Z.; Stewart, D.; Arango, A.; Chen, P.; Huang, J.; Williams, K.; et al. Amniotic Fluid-Derived Extracellular Vesicles: Characterization and Therapeutic Efficacy in an Experimental Model of Bronchopulmonary Dysplasia. Cytotherapy 2021, 23, 1097–1107. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.-M.; Zhang, K.; Zhang, J.-H. Human Breast Milk-Derived Exosomal miR-148a-3p Protects Against Necrotizing Enterocolitis by Regulating P53 and Sirtuin 1. Inflammation 2022, 45, 1254–1268. [Google Scholar] [CrossRef] [PubMed]
- Iannotta, D.; Yang, M.; Celia, C.; Di Marzio, L.; Wolfram, J. Extracellular Vesicle Therapeutics from Plasma and Adipose Tissue. Nano Today 2021, 39, 101159. [Google Scholar] [CrossRef] [PubMed]
- Mitra, P.; Gupta, S.; Sharma, P. Extracellular Vesicles (EVs) as “A Window to the Brain”: Potential, Challenges and Future Perspectives. Indian J. Clin. Biochem. 2023, 38, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Brenna, S.; Krisp, C.; Altmeppen, H.C.; Magnus, T.; Puig, B. Brain-Derived Extracellular Vesicles in Health and Disease: A Methodological Perspective. Int. J. Mol. Sci. 2021, 22, 1365. [Google Scholar] [CrossRef]
- Qiu, G.; Fan, J.; Zheng, G.; He, J.; Lin, F.; Ge, M.; Huang, L.; Wang, J.; Xia, J.; Huang, R.; et al. Diagnostic Potential of Plasma Extracellular Vesicle miR-483-3p and Let-7d-3p for Sepsis. Front. Mol. Biosci. 2022, 9, 814240. [Google Scholar] [CrossRef]
- Igami, K.; Uchiumi, T.; Shiota, M.; Ueda, S.; Tsukahara, S.; Akimoto, M.; Eto, M.; Kang, D. Extracellular Vesicles Expressing CEACAM Proteins in the Urine of Bladder Cancer Patients. Cancer Sci. 2022, 113, 3120–3133. [Google Scholar] [CrossRef]
- Ghanam, J.; Chetty, V.K.; Barthel, L.; Reinhardt, D.; Hoyer, P.-F.; Thakur, B.K. DNA in Extracellular Vesicles: From Evolution to Its Current Application in Health and Disease. Cell Biosci. 2022, 12, 37. [Google Scholar] [CrossRef]
- Fang, Y.; Wang, Z.; Liu, X.; Tyler, B.M. Biogenesis and Biological Functions of Extracellular Vesicles in Cellular and Organismal Communication With Microbes. Front. Microbiol. 2022, 13, 817844. [Google Scholar] [CrossRef]
- Urzì, O.; Gasparro, R.; Ganji, N.R.; Alessandro, R.; Raimondo, S. Plant-RNA in Extracellular Vesicles: The Secret of Cross-Kingdom Communication. Membranes 2022, 12, 352. [Google Scholar] [CrossRef]
- Gill, S.; Catchpole, R.; Forterre, P. Extracellular Membrane Vesicles in the Three Domains of Life and Beyond. FEMS Microbiol. Rev. 2019, 43, 273–303. [Google Scholar] [CrossRef]
- Silachev, D.N. Study of the Molecular Mechanisms of the Therapeutic Properties of Extracellular Vesicles. Int. J. Mol. Sci. 2023, 24, 7093. [Google Scholar] [CrossRef]
- Nemati, M.; Singh, B.; Mir, R.A.; Nemati, M.; Babaei, A.; Ahmadi, M.; Rasmi, Y.; Golezani, A.G.; Rezaie, J. Plant-Derived Extracellular Vesicles: A Novel Nanomedicine Approach with Advantages and Challenges. Cell Commun. Signal. 2022, 20, 69. [Google Scholar] [CrossRef]
- Freitas, M.S.; Bonato, V.L.D.; Pessoni, A.M.; Rodrigues, M.L.; Casadevall, A.; Almeida, F. Fungal Extracellular Vesicles as Potential Targets for Immune Interventions. mSphere 2019, 4, e00747-19. [Google Scholar] [CrossRef] [PubMed]
- Hosseini-Giv, N.; Basas, A.; Hicks, C.; El-Omar, E.; El-Assaad, F.; Hosseini-Beheshti, E. Bacterial Extracellular Vesicles and Their Novel Therapeutic Applications in Health and Cancer. Front. Cell. Infect. Microbiol. 2022, 12, 962216. [Google Scholar] [CrossRef] [PubMed]
- Lian, M.Q.; Chng, W.H.; Liang, J.; Yeo, H.Q.; Lee, C.K.; Belaid, M.; Tollemeto, M.; Wacker, M.G.; Czarny, B.; Pastorin, G. Plant-Derived Extracellular Vesicles: Recent Advancements and Current Challenges on Their Use for Biomedical Applications. J. Extracell. Vesicles 2022, 11, e12283. [Google Scholar] [CrossRef] [PubMed]
- Ñahui Palomino, R.A.; Vanpouille, C.; Costantini, P.E.; Margolis, L. Microbiota-Host Communications: Bacterial Extracellular Vesicles as a Common Language. PLoS Pathog. 2021, 17, e1009508. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Ren, Y.; Sayed, M.; Hu, X.; Lei, C.; Kumar, A.; Hutchins, E.; Mu, J.; Deng, Z.; Luo, C.; et al. Plant-Derived Exosomal MicroRNAs Shape the Gut Microbiota. Cell Host Microbe 2018, 24, 637–652. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, J.; Rodrigues, M.L.; Janbon, G. Extracellular Vesicles in Fungi: Past, Present, and Future Perspectives. Front. Cell. Infect. Microbiol. 2020, 10, 346. [Google Scholar] [CrossRef]
- Filip, R. An Update on the Role of Extracellular Vesicles in the Pathogenesis of Necrotizing Enterocolitis and Inflammatory Bowel Diseases. Cells 2021, 10, 3202. [Google Scholar] [CrossRef]
- Coughlan, C.; Bruce, K.D.; Burgy, O.; Boyd, T.D.; Michel, C.R.; Garcia-Perez, J.E.; Adame, V.; Anton, P.; Bettcher, B.M.; Chial, H.J.; et al. Exosome Isolation by Ultracentrifugation and Precipitation and Techniques for Downstream Analyses. Curr. Protoc. Cell Biol. 2020, 88, e110. [Google Scholar] [CrossRef]
- Weng, Y.; Sui, Z.; Shan, Y.; Hu, Y.; Chen, Y.; Zhang, L.; Zhang, Y. Effective Isolation of Exosomes with Polyethylene Glycol from Cell Culture Supernatant for In-Depth Proteome Profiling. Analyst 2016, 141, 4640–4646. [Google Scholar] [CrossRef] [PubMed]
- Akbar, A.; Malekian, F.; Baghban, N.; Kodam, S.P.; Ullah, M. Methodologies to Isolate and Purify Clinical Grade Extracellular Vesicles for Medical Applications. Cells 2022, 11, 186. [Google Scholar] [CrossRef] [PubMed]
- Kurinczuk, J.J.; White-Koning, M.; Badawi, N. Epidemiology of Neonatal Encephalopathy and Hypoxic-Ischaemic Encephalopathy. Early Hum. Dev. 2010, 86, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.A.; Brandon, D.H. Hypoxic Ischemic Encephalopathy: Pathophysiology and Experimental Treatments. Newborn Infant. Nurs. Rev. 2011, 11, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Silachev, D.N.; Plotnikov, E.Y.; Pevzner, I.B.; Zorova, L.D.; Balakireva, A.V.; Gulyaev, M.V.; Pirogov, Y.A.; Skulachev, V.P.; Zorov, D.B. Neuroprotective Effects of Mitochondria-Targeted Plastoquinone in a Rat Model of Neonatal Hypoxic-Ischemic Brain Injury. Molecules 2018, 23, 1871. [Google Scholar] [CrossRef] [PubMed]
- Robertsson Grossmann, K.; Eriksson Westblad, M.; Blennow, M.; Lindström, K. Outcome at Early School Age and Adolescence after Hypothermia-Treated Hypoxic-Ischaemic Encephalopathy: An Observational, Population-Based Study. Arch. Dis. Child. Fetal Neonatal Ed. 2023, 108, 295–301. [Google Scholar] [CrossRef]
- Shankaran, S.; Pappas, A.; McDonald, S.A.; Vohr, B.R.; Hintz, S.R.; Yolton, K.; Gustafson, K.E.; Leach, T.M.; Green, C.; Bara, R.; et al. Childhood Outcomes after Hypothermia for Neonatal Encephalopathy. N. Engl. J. Med. 2012, 366, 2085–2092. [Google Scholar] [CrossRef]
- Azzopardi, D.; Strohm, B.; Marlow, N.; Brocklehurst, P.; Deierl, A.; Eddama, O.; Goodwin, J.; Halliday, H.L.; Juszczak, E.; Kapellou, O.; et al. Effects of Hypothermia for Perinatal Asphyxia on Childhood Outcomes. N. Engl. J. Med. 2014, 371, 140–149. [Google Scholar] [CrossRef]
- Luo, B.-Y.; Zhou, H.-S.; Sun, Y.-F.; Xiao, Q.-X.; Chen, L.; She, H.-Q.; Wang, S.-F.; Yan, S.-S.; Chang, Q.-Y.; He, Y.-Q.; et al. The Fate and Prospects of Stem Cell Therapy in the Treatment of Hypoxic-Ischemic Encephalopathy. Eur. J. Neurosci. 2023, 58, 2384–2405. [Google Scholar] [CrossRef]
- Sukhikh, G.T.; Silachyov, D.N.; Pevzner, I.B.; Zorova, L.D.; Babenko, V.A.; Popkov, V.A.; Yankauskas, S.S.; Zubkov, V.V.; Zorov, D.B.; Plotnikov, E.Y. Prospects for Using Stem and Progenitor Cells in the Therapy of Consequences of Neonatal Hypoxic-Ischemic Encephalopathy. Obstet. Gynecol. 2016, 55–66. [Google Scholar] [CrossRef]
- Lee, J.A.; Kim, B.I.; Jo, C.H.; Choi, C.W.; Kim, E.-K.; Kim, H.-S.; Yoon, K.-S.; Choi, J.-H. Mesenchymal Stem-Cell Transplantation for Hypoxic-Ischemic Brain Injury in Neonatal Rat Model. Pediatr. Res. 2010, 67, 42–46. [Google Scholar] [CrossRef] [PubMed]
- van Velthoven, C.T.J.; van de Looij, Y.; Kavelaars, A.; Zijlstra, J.; van Bel, F.; Huppi, P.S.; Sizonenko, S.; Heijnen, C.J. Mesenchymal Stem Cells Restore Cortical Rewiring after Neonatal Ischemia in Mice. Ann. Neurol. 2012, 71, 785–796. [Google Scholar] [CrossRef] [PubMed]
- van Velthoven, C.T.J.; Kavelaars, A.; van Bel, F.; Heijnen, C.J. Mesenchymal Stem Cell Treatment after Neonatal Hypoxic-Ischemic Brain Injury Improves Behavioral Outcome and Induces Neuronal and Oligodendrocyte Regeneration. Brain Behav. Immun. 2010, 24, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.S.; Ahn, S.Y.; Im, G.H.; Sung, D.K.; Park, Y.R.; Choi, S.H.; Choi, S.J.; Chang, Y.S.; Oh, W.; Lee, J.H.; et al. Human Umbilical Cord Blood-Derived Mesenchymal Stem Cell Transplantation Attenuates Severe Brain Injury by Permanent Middle Cerebral Artery Occlusion in Newborn Rats. Pediatr. Res. 2012, 72, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.-C.; Shyu, W.-C.; Chiang, M.-F.; Lin, S.-Z.; Chang, Y.-C.; Wang, H.-J.; Su, C.-Y.; Li, H. Enhancement of Neuroplasticity through Upregulation of Beta1-Integrin in Human Umbilical Cord-Derived Stromal Cell Implanted Stroke Model. Neurobiol. Dis. 2007, 27, 339–353. [Google Scholar] [CrossRef]
- Ding, H.-F.; Zhang, H.; Ding, H.-F.; Li, D.; Yi, X.-H.; Gao, X.-Y.; Mou, W.-W.; Ju, X.-L. Therapeutic Effect of Placenta-Derived Mesenchymal Stem Cells on Hypoxic-Ischemic Brain Damage in Rats. World J. Pediatr. 2015, 11, 74–82. [Google Scholar] [CrossRef]
- Donega, V.; van Velthoven, C.T.J.; Nijboer, C.H.; van Bel, F.; Kas, M.J.H.; Kavelaars, A.; Heijnen, C.J. Intranasal Mesenchymal Stem Cell Treatment for Neonatal Brain Damage: Long-Term Cognitive and Sensorimotor Improvement. PLoS ONE 2013, 8, e51253. [Google Scholar] [CrossRef]
- Allan, D.; Tieu, A.; Lalu, M.; Burger, D. Mesenchymal Stromal Cell-Derived Extracellular Vesicles for Regenerative Therapy and Immune Modulation: Progress and Challenges toward Clinical Application. Stem Cells Transl. Med. 2020, 9, 39–46. [Google Scholar] [CrossRef]
- Akyurekli, C.; Le, Y.; Richardson, R.B.; Fergusson, D.; Tay, J.; Allan, D.S. A Systematic Review of Preclinical Studies on the Therapeutic Potential of Mesenchymal Stromal Cell-Derived Microvesicles. Stem Cell Rev. Rep. 2015, 11, 150–160. [Google Scholar] [CrossRef]
- Sisa, C.; Kholia, S.; Naylor, J.; Herrera Sanchez, M.B.; Bruno, S.; Deregibus, M.C.; Camussi, G.; Inal, J.M.; Lange, S.; Hristova, M. Mesenchymal Stromal Cell Derived Extracellular Vesicles Reduce Hypoxia-Ischaemia Induced Perinatal Brain Injury. Front. Physiol. 2019, 10, 282. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, N.; Köster, C.; Mouloud, Y.; Börger, V.; Felderhoff-Müser, U.; Bendix, I.; Giebel, B.; Herz, J. Mesenchymal Stromal Cell-Derived Extracellular Vesicles Reduce Neuroinflammation, Promote Neural Cell Proliferation and Improve Oligodendrocyte Maturation in Neonatal Hypoxic-Ischemic Brain Injury. Front. Cell. Neurosci. 2020, 14, 601176. [Google Scholar] [CrossRef] [PubMed]
- Shu, J.; Jiang, L.; Wang, M.; Wang, R.; Wang, X.; Gao, C.; Xia, Z. Human Bone Marrow Mesenchymal Stem Cells-Derived Exosomes Protect against Nerve Injury via Regulating Immune Microenvironment in Neonatal Hypoxic-Ischemic Brain Damage Model. Immunobiology 2022, 227, 152178. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Yuan, Y.; Zhang, X.; Lu, Y.; Dong, N.; Jiang, X.; Xu, J.; Zheng, D. Human Umbilical Cord Mesenchymal Stem Cell-Derived Exosomes Attenuate Oxygen-Glucose Deprivation/Reperfusion-Induced Microglial Pyroptosis by Promoting FOXO3a-Dependent Mitophagy. Oxid. Med. Cell. Longev. 2021, 2021, 6219715. [Google Scholar] [CrossRef] [PubMed]
- Cristante, E.; McArthur, S.; Mauro, C.; Maggioli, E.; Romero, I.A.; Wylezinska-Arridge, M.; Couraud, P.O.; Lopez-Tremoleda, J.; Christian, H.C.; Weksler, B.B.; et al. Identification of an Essential Endogenous Regulator of Blood-Brain Barrier Integrity, and Its Pathological and Therapeutic Implications. Proc. Natl. Acad. Sci. USA 2013, 110, 832–841. [Google Scholar] [CrossRef] [PubMed]
- Gussenhoven, R.; Klein, L.; Ophelders, D.R.M.G.; Habets, D.H.J.; Giebel, B.; Kramer, B.W.; Schurgers, L.J.; Reutelingsperger, C.P.M.; Wolfs, T.G.A.M. Annexin A1 as Neuroprotective Determinant for Blood-Brain Barrier Integrity in Neonatal Hypoxic-Ischemic Encephalopathy. J. Clin. Med. 2019, 8, 137. [Google Scholar] [CrossRef]
- Yang, Y.; Cai, Y.; Zhang, Y.; Liu, J.; Xu, Z. Exosomes Secreted by Adipose-Derived Stem Cells Contribute to Angiogenesis of Brain Microvascular Endothelial Cells Following Oxygen-Glucose Deprivation In Vitro Through MicroRNA-181b/TRPM7 Axis. J. Mol. Neurosci. 2018, 65, 74–83. [Google Scholar] [CrossRef]
- Geng, W.; Tang, H.; Luo, S.; Lv, Y.; Liang, D.; Kang, X.; Hong, W. Exosomes from miRNA-126-Modified ADSCs Promotes Functional Recovery after Stroke in Rats by Improving Neurogenesis and Suppressing Microglia Activation. Am. J. Transl. Res. 2019, 11, 780–792. [Google Scholar]
- Budnik, V.; Ruiz-Cañada, C.; Wendler, F. Extracellular Vesicles Round off Communication in the Nervous System. Nat. Rev. Neurosci. 2016, 17, 160–172. [Google Scholar] [CrossRef]
- Song, Y.; Li, Z.; He, T.; Qu, M.; Jiang, L.; Li, W.; Shi, X.; Pan, J.; Zhang, L.; Wang, Y.; et al. M2 Microglia-Derived Exosomes Protect the Mouse Brain from Ischemia-Reperfusion Injury via Exosomal miR-124. Theranostics 2019, 9, 2910–2923. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Ye, G.; Liu, Y.; Huang, D.; Luo, Q.; Chen, W.; Qi, Z. miR-150-3p Enhances Neuroprotective Effects of Neural Stem Cell Exosomes after Hypoxic-Ischemic Brain Injury by Targeting CASP2. Neurosci. Lett. 2022, 779, 136635. [Google Scholar] [CrossRef]
- Holme, N.; Chetcuti, P. The Pathophysiology of Respiratory Distress Syndrome in Neonates. Paediatr. Child. Health 2012, 22, 507–512. [Google Scholar] [CrossRef]
- Schmidt, A.R.; Ramamoorthy, C. Bronchopulmonary Dysplasia. Paediatr. Anaesth. 2022, 32, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Principi, N.; Di Pietro, G.M.; Esposito, S. Bronchopulmonary Dysplasia: Clinical Aspects and Preventive and Therapeutic Strategies. J. Transl. Med. 2018, 16, 36. [Google Scholar] [CrossRef] [PubMed]
- Geetha, O.; Rajadurai, V.S.; Anand, A.J.; Dela Puerta, R.; Huey Quek, B.; Khoo, P.C.; Chua, M.C.; Agarwal, P. New BPD-Prevalence and Risk Factors for Bronchopulmonary Dysplasia/Mortality in Extremely Low Gestational Age Infants ≤28 Weeks. J. Perinatol. 2021, 41, 1943–1950. [Google Scholar] [CrossRef] [PubMed]
- Pierro, M.; Ionescu, L.; Montemurro, T.; Vadivel, A.; Weissmann, G.; Oudit, G.; Emery, D.; Bodiga, S.; Eaton, F.; Péault, B.; et al. Short-Term, Long-Term and Paracrine Effect of Human Umbilical Cord-Derived Stem Cells in Lung Injury Prevention and Repair in Experimental Bronchopulmonary Dysplasia. Thorax 2013, 68, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Willis, G.R.; Fernandez-Gonzalez, A.; Anastas, J.; Vitali, S.H.; Liu, X.; Ericsson, M.; Kwong, A.; Mitsialis, S.A.; Kourembanas, S. Mesenchymal Stromal Cell Exosomes Ameliorate Experimental Bronchopulmonary Dysplasia and Restore Lung Function through Macrophage Immunomodulation. Am. J. Respir. Crit. Care Med. 2018, 197, 104–116. [Google Scholar] [CrossRef]
- Wang, J.; Dong, W. Oxidative Stress and Bronchopulmonary Dysplasia. Gene 2018, 678, 177–183. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, A.; Huang, F.; Xu, J.; Zhao, M. MSC-EXO and Tempol Ameliorate Bronchopulmonary Dysplasia in Newborn Rats by Activating HIF-1α. Pediatr. Pulmonol. 2023, 58, 1367–1379. [Google Scholar] [CrossRef] [PubMed]
- Chiaradia, E.; Tancini, B.; Emiliani, C.; Delo, F.; Pellegrino, R.M.; Tognoloni, A.; Urbanelli, L.; Buratta, S. Extracellular Vesicles under Oxidative Stress Conditions: Biological Properties and Physiological Roles. Cells 2021, 10, 1763. [Google Scholar] [CrossRef]
- Sharma, M.; Bellio, M.A.; Benny, M.; Kulandavelu, S.; Chen, P.; Janjindamai, C.; Han, C.; Chang, L.; Sterling, S.; Williams, K.; et al. Mesenchymal Stem Cell-Derived Extracellular Vesicles Prevent Experimental Bronchopulmonary Dysplasia Complicated By Pulmonary Hypertension. Stem Cells Transl. Med. 2022, 11, 828–840. [Google Scholar] [CrossRef]
- Ai, D.; Shen, J.; Sun, J.; Zhu, Z.; Gao, R.; Du, Y.; Yuan, L.; Chen, C.; Zhou, J. Mesenchymal Stem Cell-Derived Extracellular Vesicles Suppress Hyperoxia-Induced Transdifferentiation of Rat Alveolar Type 2 Epithelial Cells. Stem Cells Dev. 2022, 31, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Mitsialis, S.A.; Aslam, M.; Vitali, S.H.; Vergadi, E.; Konstantinou, G.; Sdrimas, K.; Fernandez-Gonzalez, A.; Kourembanas, S. Exosomes Mediate the Cytoprotective Action of Mesenchymal Stromal Cells on Hypoxia-Induced Pulmonary Hypertension. Circulation 2012, 126, 2601–2611. [Google Scholar] [CrossRef] [PubMed]
- Reis, M.; Willis, G.R.; Fernandez-Gonzalez, A.; Yeung, V.; Taglauer, E.; Magaletta, M.; Parsons, T.; Derr, A.; Liu, X.; Maehr, R.; et al. Mesenchymal Stromal Cell-Derived Extracellular Vesicles Restore Thymic Architecture and T Cell Function Disrupted by Neonatal Hyperoxia. Front. Immunol. 2021, 12, 640595. [Google Scholar] [CrossRef] [PubMed]
- Chaubey, S.; Thueson, S.; Ponnalagu, D.; Alam, M.A.; Gheorghe, C.P.; Aghai, Z.; Singh, H.; Bhandari, V. Early Gestational Mesenchymal Stem Cell Secretome Attenuates Experimental Bronchopulmonary Dysplasia in Part via Exosome-Associated Factor TSG-6. Stem Cell Res. Ther. 2018, 9, 173. [Google Scholar] [CrossRef]
- Braun, R.K.; Chetty, C.; Balasubramaniam, V.; Centanni, R.; Haraldsdottir, K.; Hematti, P.; Eldridge, M.W. Intraperitoneal Injection of MSC-Derived Exosomes Prevent Experimental Bronchopulmonary Dysplasia. Biochem. Biophys. Res. Commun. 2018, 503, 2653–2658. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Park, W.S.; Kim, Y.E.; Sung, D.K.; Sung, S.I.; Ahn, J.Y.; Chang, Y.S. Vascular Endothelial Growth Factor Mediates the Therapeutic Efficacy of Mesenchymal Stem Cell-Derived Extracellular Vesicles against Neonatal Hyperoxic Lung Injury. Exp. Mol. Med. 2018, 50, 1–12. [Google Scholar] [CrossRef]
- Hou, A.; Fu, J.; Yang, H.; Zhu, Y.; Pan, Y.; Xu, S.; Xue, X. Hyperoxia Stimulates the Transdifferentiation of Type II Alveolar Epithelial Cells in Newborn Rats. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 308, L861–L872. [Google Scholar] [CrossRef]
- Lithopoulos, M.A.; Strueby, L.; O’Reilly, M.; Zhong, S.; Möbius, M.A.; Eaton, F.; Fung, M.; Hurskainen, M.; Cyr-Depauw, C.; Suen, C.; et al. Pulmonary and Neurologic Effects of Mesenchymal Stromal Cell Extracellular Vesicles in a Multifactorial Lung Injury Model. Am. J. Respir. Crit. Care Med. 2022, 205, 1186–1201. [Google Scholar] [CrossRef]
- You, J.; Zhou, O.; Liu, J.; Zou, W.; Zhang, L.; Tian, D.; Dai, J.; Luo, Z.; Liu, E.; Fu, Z.; et al. Human Umbilical Cord Mesenchymal Stem Cell-Derived Small Extracellular Vesicles Alleviate Lung Injury in Rat Model of Bronchopulmonary Dysplasia by Affecting Cell Survival and Angiogenesis. Stem Cells Dev. 2020, 29, 1520–1532. [Google Scholar] [CrossRef]
- Wu, Y.; Li, J.; Yuan, R.; Deng, Z.; Wu, X. Bone Marrow Mesenchymal Stem Cell-Derived Exosomes Alleviate Hyperoxia-Induced Lung Injury via the Manipulation of microRNA-425. Arch. Biochem. Biophys. 2021, 697, 108712. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Krause, M.; Yawno, T.; Kusuma, G.D.; Schwab, R.; Barabadi, M.; Maleken, A.S.; Chan, S.T.; Hunt, R.; Greening, D.; et al. Assessing the Impact of Gestational Age of Donors on the Efficacy of Amniotic Epithelial Cell-Derived Extracellular Vesicles in Experimental Bronchopulmonary Dysplasia. Stem Cell Res. Ther. 2022, 13, 196. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.R.; Sills, W.S.; Hanrahan, K.; Ziegler, A.; Tidd, K.M.; Cook, E.; Sannes, P.L. Expression of WNT5A in Idiopathic Pulmonary Fibrosis and Its Control by TGF-β and WNT7B in Human Lung Fibroblasts. J. Histochem. Cytochem. 2016, 64, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Edmond, K.; Zaidi, A. New Approaches to Preventing, Diagnosing, and Treating Neonatal Sepsis. PLoS Med. 2010, 7, e1000213. [Google Scholar] [CrossRef] [PubMed]
- Shane, A.L.; Sánchez, P.J.; Stoll, B.J. Neonatal Sepsis. Lancet 2017, 390, 1770–1780. [Google Scholar] [CrossRef]
- Li, J.; Shen, L.; Qian, K. Global, Regional, and National Incidence and Mortality of Neonatal Sepsis and Other Neonatal Infections, 1990–2019. Front. Public Health 2023, 11, 1139832. [Google Scholar] [CrossRef]
- Burgelman, M.; Vandendriessche, C.; Vandenbroucke, R.E. Extracellular Vesicles: A Double-Edged Sword in Sepsis. Pharmaceuticals 2021, 14, 829. [Google Scholar] [CrossRef]
- Kronstadt, S.M.; Pottash, A.E.; Levy, D.; Wang, S.; Chao, W.; Jay, S.M. Therapeutic Potential of Extracellular Vesicles for Sepsis Treatment. Adv. Ther. 2021, 4, 2000259. [Google Scholar] [CrossRef]
- Vergnano, S.; Sharland, M.; Kazembe, P.; Mwansambo, C.; Heath, P.T. Neonatal Sepsis: An International Perspective. Arch. Dis. Child. Fetal Neonatal Ed. 2005, 90, F220–F224. [Google Scholar] [CrossRef]
- Kim, Y.-E.; Ahn, S.-Y.; Park, W.-S.; Sung, D.-K.; Sung, S.-I.; Yang, M.-S.; Chang, Y.-S. Mesenchymal-Stem-Cell-Derived Extracellular Vesicles Attenuate Brain Injury in Escherichia Coli Meningitis in Newborn Rats. Life 2022, 12, 1030. [Google Scholar] [CrossRef]
- Alganabi, M.; Lee, C.; Bindi, E.; Li, B.; Pierro, A. Recent Advances in Understanding Necrotizing Enterocolitis. F1000Research 2019, 8, 1–8. [Google Scholar] [CrossRef]
- Bazacliu, C.; Neu, J. Necrotizing Enterocolitis: Long Term Complications. Curr. Pediatr. Rev. 2019, 15, 115–124. [Google Scholar] [CrossRef]
- Hackam, D.J.; Sodhi, C.P.; Good, M. New Insights into Necrotizing Enterocolitis: From Laboratory Observation to Personalized Prevention and Treatment. J. Pediatr. Surg. 2019, 54, 398–404. [Google Scholar] [CrossRef]
- Li, B.; Lee, C.; O’Connell, J.S.; Antounians, L.; Ganji, N.; Alganabi, M.; Cadete, M.; Nascimben, F.; Koike, Y.; Hock, A.; et al. Activation of Wnt Signaling by Amniotic Fluid Stem Cell-Derived Extracellular Vesicles Attenuates Intestinal Injury in Experimental Necrotizing Enterocolitis. Cell Death Dis. 2020, 11, 750. [Google Scholar] [CrossRef]
- Drucker, N.A.; Te Winkel, J.P.; Shelley, W.C.; Olson, K.R.; Markel, T.A. Inhibiting Hydrogen Sulfide Production in Umbilical Stem Cells Reduces Their Protective Effects during Experimental Necrotizing Enterocolitis. J. Pediatr. Surg. 2019, 54, 1168–1173. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, J.S.; Li, B.; Zito, A.; Ahmed, A.; Cadete, M.; Ganji, N.; Lau, E.; Alganabi, M.; Farhat, N.; Lee, C.; et al. Treatment of Necrotizing Enterocolitis by Conditioned Medium Derived from Human Amniotic Fluid Stem Cells. PLoS ONE 2021, 16, e0260522. [Google Scholar] [CrossRef] [PubMed]
- Rager, T.M.; Olson, J.K.; Zhou, Y.; Wang, Y.; Besner, G.E. Exosomes Secreted from Bone Marrow-Derived Mesenchymal Stem Cells Protect the Intestines from Experimental Necrotizing Enterocolitis. J. Pediatr. Surg. 2016, 51, 942–947. [Google Scholar] [CrossRef] [PubMed]
- Claud, E.C. Neonatal Necrotizing Enterocolitis-Inflammation and Intestinal Immaturity. Antiinflamm Antiallergy Agents Med. Chem. 2009, 8, 248–259. [Google Scholar] [CrossRef]
- Heidari, N.; Abbasi-Kenarsari, H.; Namaki, S.; Baghaei, K.; Zali, M.R.; Ghaffari Khaligh, S.; Hashemi, S.M. Adipose-Derived Mesenchymal Stem Cell-Secreted Exosome Alleviates Dextran Sulfate Sodium-Induced Acute Colitis by Treg Cell Induction and Inflammatory Cytokine Reduction. J. Cell. Physiol. 2021, 236, 5906–5920. [Google Scholar] [CrossRef]
- Yang, S.; Liang, X.; Song, J.; Li, C.; Liu, A.; Luo, Y.; Ma, H.; Tan, Y.; Zhang, X. A Novel Therapeutic Approach for Inflammatory Bowel Disease by Exosomes Derived from Human Umbilical Cord Mesenchymal Stem Cells to Repair Intestinal Barrier via TSG-6. Stem Cell Res. Ther. 2021, 12, 315. [Google Scholar] [CrossRef]
- Yan, Y.; Li, K.; Jiang, J.; Jiang, L.; Ma, X.; Ai, F.; Qiu, S.; Si, W. Perinatal Tissue-Derived Exosomes Ameliorate Colitis in Mice by Regulating the Foxp3 + Treg Cells and Gut Microbiota. Stem Cell Res. Ther. 2023, 14, 43. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Zhang, Z.-Y.; Yuan, J.-T.; Ocansey, D.K.W.; Tu, Q.; Zhang, X.; Qian, H.; Xu, W.-R.; Qiu, W.; Mao, F. hucMSC-Derived Exosomes Attenuate Colitis by Regulating Macrophage Pyroptosis via the miR-378a-5p/NLRP3 Axis. Stem Cell Res. Ther. 2021, 12, 416. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Shen, Y.; Guo, D.; Yang, D.; Liu, J.; Fei, X.; Yang, Y.; Zhang, B.; Lin, Z.; Yang, F.; et al. EpCAM-Dependent Extracellular Vesicles from Intestinal Epithelial Cells Maintain Intestinal Tract Immune Balance. Nat. Commun. 2016, 7, 13045. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Huang, H.; Zhao, X.; Zhou, M.; Chen, S.; Wang, C.; Han, Z.; Han, Z.-C.; Guo, Z.; Li, Z.; et al. Extracellular Vesicles Derived from Human Placental Mesenchymal Stem Cells Alleviate Experimental Colitis in Mice by Inhibiting Inflammation and Oxidative Stress. Int. J. Mol. Med. 2020, 46, 1551–1561. [Google Scholar] [CrossRef]
- Yu, H.; Yang, X.; Xiao, X.; Xu, M.; Yang, Y.; Xue, C.; Li, X.; Wang, S.; Zhao, R.C. Human Adipose Mesenchymal Stem Cell-Derived Exosomes Protect Mice from DSS-Induced Inflammatory Bowel Disease by Promoting Intestinal-Stem-Cell and Epithelial Regeneration. Aging Dis. 2021, 12, 1423–1437. [Google Scholar] [CrossRef]
- Chen, T.; Xie, M.-Y.; Sun, J.-J.; Ye, R.-S.; Cheng, X.; Sun, R.-P.; Wei, L.-M.; Li, M.; Lin, D.-L.; Jiang, Q.-Y.; et al. Porcine Milk-Derived Exosomes Promote Proliferation of Intestinal Epithelial Cells. Sci. Rep. 2016, 6, 33862. [Google Scholar] [CrossRef]
- Hock, A.; Miyake, H.; Li, B.; Lee, C.; Ermini, L.; Koike, Y.; Chen, Y.; Määttänen, P.; Zani, A.; Pierro, A. Breast Milk-Derived Exosomes Promote Intestinal Epithelial Cell Growth. J. Pediatr. Surg. 2017, 52, 755–759. [Google Scholar] [CrossRef]
- Accarie, A.; l’Homme, B.; Benadjaoud, M.A.; Lim, S.K.; Guha, C.; Benderitter, M.; Tamarat, R.; Sémont, A. Extracellular Vesicles Derived from Mesenchymal Stromal Cells Mitigate Intestinal Toxicity in a Mouse Model of Acute Radiation Syndrome. Stem Cell Res. Ther. 2020, 11, 371. [Google Scholar] [CrossRef]
- Borewicz, K.; Gu, F.; Saccenti, E.; Arts, I.C.W.; Penders, J.; Thijs, C.; van Leeuwen, S.S.; Lindner, C.; Nauta, A.; van Leusen, E.; et al. Correlating Infant Fecal Microbiota Composition and Human Milk Oligosaccharide Consumption by Microbiota of 1-Month-Old Breastfed Infants. Mol. Nutr. Food Res. 2019, 63, e1801214. [Google Scholar] [CrossRef]
- van Herwijnen, M.J.C.; Zonneveld, M.I.; Goerdayal, S.; Nolte-’t Hoen, E.N.M.; Garssen, J.; Stahl, B.; Maarten Altelaar, A.F.; Redegeld, F.A.; Wauben, M.H.M. Comprehensive Proteomic Analysis of Human Milk-Derived Extracellular Vesicles Unveils a Novel Functional Proteome Distinct from Other Milk Components. Mol. Cell. Proteom. 2016, 15, 3412–3423. [Google Scholar] [CrossRef]
- Walker, W.A.; Iyengar, R.S. Breast Milk, Microbiota, and Intestinal Immune Homeostasis. Pediatr. Res. 2015, 77, 220–228. [Google Scholar] [CrossRef]
- Hu, Y.; Hell, L.; Kendlbacher, R.A.; Hajji, N.; Hau, C.; van Dam, A.; Berckmans, R.J.; Wisgrill, L.; Ay, C.; Pabinger, I.; et al. Human Milk Triggers Coagulation via Tissue Factor-Exposing Extracellular Vesicles. Blood Adv. 2020, 4, 6274–6282. [Google Scholar] [CrossRef]
- Kersin, S.G.; Özek, E. Breast Milk Stem Cells: Are They Magic Bullets in Neonatology? Turk. Arch. Pediatr. 2021, 56, 187–191. [Google Scholar] [CrossRef]
- Shackleton, M.; Vaillant, F.; Simpson, K.J.; Stingl, J.; Smyth, G.K.; Asselin-Labat, M.-L.; Wu, L.; Lindeman, G.J.; Visvader, J.E. Generation of a Functional Mammary Gland from a Single Stem Cell. Nature 2006, 439, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Cacho, N.T.; Lawrence, R.M. Innate Immunity and Breast Milk. Front. Immunol. 2017, 8, 584. [Google Scholar] [CrossRef] [PubMed]
- Maghraby, M.K.; Li, B.; Chi, L.; Ling, C.; Benmoussa, A.; Provost, P.; Postmus, A.C.; Abdi, A.; Pierro, A.; Bourdon, C.; et al. Extracellular Vesicles Isolated from Milk Can Improve Gut Barrier Dysfunction Induced by Malnutrition. Sci. Rep. 2021, 11, 7635. [Google Scholar] [CrossRef]
- van Herwijnen, M.J.C.; Driedonks, T.A.P.; Snoek, B.L.; Kroon, A.M.T.; Kleinjan, M.; Jorritsma, R.; Pieterse, C.M.J.; Nolte-’t Hoen, E.N.M.; Wauben, M.H.M. Abundantly Present miRNAs in Milk-Derived Extracellular Vesicles Are Conserved Between Mammals. Front. Nutr. 2018, 5, 81. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhan, Z.; Xu, L.; Ma, F.; Li, D.; Guo, Z.; Li, N.; Cao, X. MicroRNA-148/152 Impair Innate Response and Antigen Presentation of TLR-Triggered Dendritic Cells by Targeting CaMKIIα. J. Immunol. 2010, 185, 7244–7251. [Google Scholar] [CrossRef]
- Wang, X.; Yan, X.; Zhang, L.; Cai, J.; Zhou, Y.; Liu, H.; Hu, Y.; Chen, W.; Xu, S.; Liu, P.; et al. Identification and Peptidomic Profiling of Exosomes in Preterm Human Milk: Insights Into Necrotizing Enterocolitis Prevention. Mol. Nutr. Food Res. 2019, 63, e1801247. [Google Scholar] [CrossRef]
- Cheyuo, C.; Aziz, M.; Yang, W.-L.; Jacob, A.; Zhou, M.; Wang, P. Milk Fat Globule-EGF Factor VIII Attenuates CNS Injury by Promoting Neural Stem Cell Proliferation and Migration after Cerebral Ischemia. PLoS ONE 2015, 10, e0122833. [Google Scholar] [CrossRef]
- Bu, H.-F.; Zuo, X.-L.; Wang, X.; Ensslin, M.A.; Koti, V.; Hsueh, W.; Raymond, A.S.; Shur, B.D.; Tan, X.-D. Milk Fat Globule-EGF Factor 8/Lactadherin Plays a Crucial Role in Maintenance and Repair of Murine Intestinal Epithelium. J. Clin. Investig. 2007, 117, 3673–3683. [Google Scholar] [CrossRef]
- Tobita, K.; Kawahara, T.; Otani, H. Bovine Beta-Casein (1-28), a Casein Phosphopeptide, Enhances Proliferation and IL-6 Expression of Mouse CD19+ Cells via Toll-like Receptor 4. J. Agric. Food Chem. 2006, 54, 8013–8017. [Google Scholar] [CrossRef]
- Oh, N.S.; Joung, J.Y.; Lee, J.Y.; Kim, Y.; Kim, S.H. Enhancement of Antioxidative and Intestinal Anti-Inflammatory Activities of Glycated Milk Casein after Fermentation with Lactobacillus Rhamnosus 4B15. J. Agric. Food Chem. 2017, 65, 4744–4754. [Google Scholar] [CrossRef]
- Caccavo, D.; Pellegrino, N.M.; Altamura, M.; Rigon, A.; Amati, L.; Amoroso, A.; Jirillo, E. Antimicrobial and Immunoregulatory Functions of Lactoferrin and Its Potential Therapeutic Application. J. Endotoxin Res. 2002, 8, 403–417. [Google Scholar] [CrossRef]
- Nichols, B.L.; McKee, K.; Putman, M.; Henry, J.F.; Nichols, V.N. Human Lactoferrin Supplementation of Infant Formulas Increases Thymidine Incorporation into the DNA of Rat Crypt Cells. J. Pediatr. Gastroenterol. Nutr. 1989, 8, 102–109. [Google Scholar] [CrossRef]
- Sanwlani, R.; Fonseka, P.; Chitti, S.V.; Mathivanan, S. Milk-Derived Extracellular Vesicles in Inter-Organism, Cross-Species Communication and Drug Delivery. Proteomes 2020, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Pisano, C.; Galley, J.; Elbahrawy, M.; Wang, Y.; Farrell, A.; Brigstock, D.; Besner, G.E. Human Breast Milk-Derived Extracellular Vesicles in the Protection Against Experimental Necrotizing Enterocolitis. J. Pediatr. Surg. 2020, 55, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Zhao, Y.; Wang, K.; Nan, X.; Chen, R.; Xiong, B. Effects of Milk-Derived Extracellular Vesicles on the Colonic Transcriptome and Proteome in Murine Model. Nutrients 2022, 14, 3057. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Wang, K.; Zhao, Y.; Nan, X.; Chen, R.; Quan, S.; Xiong, B. Supplementation with Milk-Derived Extracellular Vesicles Shapes the Gut Microbiota and Regulates the Transcriptomic Landscape in Experimental Colitis. Nutrients 2022, 14, 1808. [Google Scholar] [CrossRef] [PubMed]
- Iliopoulos, D.; Hirsch, H.A.; Struhl, K. An Epigenetic Switch Involving NF-κB, Lin28, Let-7 MicroRNA, and IL6 Links Inflammation to Cell Transformation. Cell 2009, 139, 693–706. [Google Scholar] [CrossRef]
- Li, B.; Hock, A.; Wu, R.Y.; Minich, A.; Botts, S.R.; Lee, C.; Antounians, L.; Miyake, H.; Koike, Y.; Chen, Y.; et al. Bovine Milk-Derived Exosomes Enhance Goblet Cell Activity and Prevent the Development of Experimental Necrotizing Enterocolitis. PLoS ONE 2019, 14, e0211431. [Google Scholar] [CrossRef]
- Liu, S.; Zhao, W.; Lan, P.; Mou, X. The Microbiome in Inflammatory Bowel Diseases: From Pathogenesis to Therapy. Protein Cell 2021, 12, 331–345. [Google Scholar] [CrossRef]
- Mazmanian, S.K. Capsular Polysaccharides of Symbiotic Bacteria Modulate Immune Responses during Experimental Colitis. J. Pediatr. Gastroenterol. Nutr. 2008, 46 (Suppl. S1), E11–E12. [Google Scholar] [CrossRef]
- Shen, Y.; Giardino Torchia, M.L.; Lawson, G.W.; Karp, C.L.; Ashwell, J.D.; Mazmanian, S.K. Outer Membrane Vesicles of a Human Commensal Mediate Immune Regulation and Disease Protection. Cell Host Microbe 2012, 12, 509–520. [Google Scholar] [CrossRef]
- Wang, X.; Lin, S.; Wang, L.; Cao, Z.; Zhang, M.; Zhang, Y.; Liu, R.; Liu, J. Versatility of Bacterial Outer Membrane Vesicles in Regulating Intestinal Homeostasis. Sci. Adv. 2023, 9, eade5079. [Google Scholar] [CrossRef]
- Kang, C.-S.; Ban, M.; Choi, E.-J.; Moon, H.-G.; Jeon, J.-S.; Kim, D.-K.; Park, S.-K.; Jeon, S.G.; Roh, T.-Y.; Myung, S.-J.; et al. Extracellular Vesicles Derived from Gut Microbiota, Especially Akkermansia Muciniphila, Protect the Progression of Dextran Sulfate Sodium-Induced Colitis. PLoS ONE 2013, 8, e76520. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Yu, F.; Hu, X.; Liu, Y.; Jin, Y.; Ren, H.; Lu, S.; Guo, J.; Chang, J.; Li, Y.; et al. Protective Effect of Lactiplantibacillus Plantarum Subsp. Plantarum SC-5 on Dextran Sulfate Sodium—Induced Colitis in Mice. Foods 2023, 12, 897. [Google Scholar] [CrossRef]
- Ma, L.; Lyu, W.; Song, Y.; Chen, K.; Lv, L.; Yang, H.; Wang, W.; Xiao, Y. Anti-Inflammatory Effect of Clostridium Butyricum-Derived Extracellular Vesicles in Ulcerative Colitis: Impact on Host microRNAs Expressions and Gut Microbiome Profiles. Mol. Nutr. Food Res. 2023, 67, e2200884. [Google Scholar] [CrossRef] [PubMed]
- Rui, T.; Wang, K.; Xiang, A.; Guo, J.; Tang, N.; Jin, X.; Lin, Y.; Liu, J.; Zhang, X. Serum Exosome-Derived piRNAs Could Be Promising Biomarkers for HCC Diagnosis. Int. J. Nanomed. 2023, 18, 1989–2001. [Google Scholar] [CrossRef] [PubMed]
- Matei, A.C.; Antounians, L.; Zani, A. Extracellular Vesicles as a Potential Therapy for Neonatal Conditions: State of the Art and Challenges in Clinical Translation. Pharmaceutics 2019, 11, 404. [Google Scholar] [CrossRef] [PubMed]
- Thakur, B.K.; Zhang, H.; Becker, A.; Matei, I.; Huang, Y.; Costa-Silva, B.; Zheng, Y.; Hoshino, A.; Brazier, H.; Xiang, J.; et al. Double-Stranded DNA in Exosomes: A Novel Biomarker in Cancer Detection. Cell Res. 2014, 24, 766–769. [Google Scholar] [CrossRef]
- Whitford, W.; Guterstam, P. Exosome Manufacturing Status. Future Med. Chem. 2019, 11, 1225–1236. [Google Scholar] [CrossRef]
- Pachler, K.; Lener, T.; Streif, D.; Dunai, Z.A.; Desgeorges, A.; Feichtner, M.; Öller, M.; Schallmoser, K.; Rohde, E.; Gimona, M. A Good Manufacturing Practice-Grade Standard Protocol for Exclusively Human Mesenchymal Stromal Cell-Derived Extracellular Vesicles. Cytotherapy 2017, 19, 458–472. [Google Scholar] [CrossRef]
- Willis, G.R.; Kourembanas, S.; Mitsialis, S.A. Toward Exosome-Based Therapeutics: Isolation, Heterogeneity, and Fit-for-Purpose Potency. Front. Cardiovasc. Med. 2017, 4, 63. [Google Scholar] [CrossRef]
- Ophelders, D.R.M.G.; Wolfs, T.G.A.M.; Jellema, R.K.; Zwanenburg, A.; Andriessen, P.; Delhaas, T.; Ludwig, A.-K.; Radtke, S.; Peters, V.; Janssen, L.; et al. Mesenchymal Stromal Cell-Derived Extracellular Vesicles Protect the Fetal Brain After Hypoxia-Ischemia. Stem Cells Transl. Med. 2016, 5, 754–763. [Google Scholar] [CrossRef]
- Chu, X.; Liu, D.; Li, T.; Ke, H.; Xin, D.; Wang, S.; Cao, Y.; Xue, H.; Wang, Z. Hydrogen Sulfide-Modified Extracellular Vesicles from Mesenchymal Stem Cells for Treatment of Hypoxic-Ischemic Brain Injury. J. Control. Release 2020, 328, 13–27. [Google Scholar] [CrossRef]
- Xin, D.; Li, T.; Chu, X.; Ke, H.; Liu, D.; Wang, Z. MSCs-Extracellular Vesicles Attenuated Neuroinflammation, Synapse Damage and Microglial Phagocytosis after Hypoxia-Ischemia Injury by Preventing Osteopontin Expression. Pharmacol. Res. 2021, 164, 105322. [Google Scholar] [CrossRef] [PubMed]
- Pathipati, P.; Lecuyer, M.; Faustino, J.; Strivelli, J.; Phinney, D.G.; Vexler, Z.S. Mesenchymal Stem Cell (MSC)-Derived Extracellular Vesicles Protect from Neonatal Stroke by Interacting with Microglial Cells. Neurotherapeutics 2021, 18, 1939–1952. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Jiang, Y.; Sun, Y. Astrocyte-Derived Exosomes Carry microRNA-17-5p to Protect Neonatal Rats from Hypoxic-Ischemic Brain Damage via Inhibiting BNIP-2 Expression. NeuroToxicology 2021, 83, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Lawson, A.; Snyder, W.; Peeples, E.S. Intranasal Administration of Extracellular Vesicles Mitigates Apoptosis in a Mouse Model of Neonatal Hypoxic-Ischemic Brain Injury. Neonatology 2022, 119, 345–353. [Google Scholar] [CrossRef]
- Luo, H.; Huang, F.; Huang, Z.; Huang, H.; Liu, C.; Feng, Y.; Qi, Z. microRNA-93 Packaged in Extracellular Vesicles from Mesenchymal Stem Cells Reduce Neonatal Hypoxic-Ischemic Brain Injury. Brain Res. 2022, 1794, 148042. [Google Scholar] [CrossRef]
- Labusek, N.; Mouloud, Y.; Köster, C.; Diesterbeck, E.; Tertel, T.; Wiek, C.; Hanenberg, H.; Horn, P.A.; Felderhoff-Müser, U.; Bendix, I.; et al. Extracellular Vesicles from Immortalized Mesenchymal Stromal Cells Protect against Neonatal Hypoxic-Ischemic Brain Injury. Inflamm. Regen. 2023, 43, 24. [Google Scholar] [CrossRef]
- Porzionato, A.; Zaramella, P.; Dedja, A.; Guidolin, D.; Van Wemmel, K.; Macchi, V.; Jurga, M.; Perilongo, G.; De Caro, R.; Baraldi, E.; et al. Intratracheal Administration of Clinical-Grade Mesenchymal Stem Cell-Derived Extracellular Vesicles Reduces Lung Injury in a Rat Model of Bronchopulmonary Dysplasia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 316, L6–L19. [Google Scholar] [CrossRef] [PubMed]
- Willis, G.R.; Fernandez-Gonzalez, A.; Reis, M.; Yeung, V.; Liu, X.; Ericsson, M.; Andrews, N.A.; Mitsialis, S.A.; Kourembanas, S. Mesenchymal Stromal Cell-Derived Small Extracellular Vesicles Restore Lung Architecture and Improve Exercise Capacity in a Model of Neonatal Hyperoxia-Induced Lung Injury. J. Extracell. Vesicles 2020, 9, 1790874. [Google Scholar] [CrossRef]
- Abele, A.N.; Taglauer, E.S.; Almeda, M.; Wilson, N.; Abikoye, A.; Seedorf, G.J.; Mitsialis, S.A.; Kourembanas, S.; Abman, S.H. Antenatal Mesenchymal Stromal Cell Extracellular Vesicle Treatment Preserves Lung Development in a Model of Bronchopulmonary Dysplasia Due to Chorioamnionitis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2022, 322, L179–L190. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, Y.; Xu, G.; Liu, L.; Li, H.; Li, Y.; Yin, J.; Wang, X.; Yu, Z. Human Breast Milk-Derived Exosomes through Inhibiting AT II Cell Apoptosis to Prevent Bronchopulmonary Dysplasia in Rat Lung. J. Cell. Mol. Med. 2022, 26, 4169–4182. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.-Q.; Wang, D.; Chen, S.; Zheng, J.; Hao, T.-F.; Li, X.-H.; Luo, L.-H.; Gu, J.; Lian, C.-Y.; Li, X.-S.; et al. Umbilical Cord Blood-Derived Exosomes from Healthy Term Pregnancies Protect against Hyperoxia-Induced Lung Injury in Mice. Clin. Transl. Sci. 2023, 16, 966–977. [Google Scholar] [CrossRef]
- Barlow, B.; Santulli, T.V.; Heird, W.C.; Pitt, J.; Blanc, W.A.; Schullinger, J.N. An Experimental Study of Acute Neonatal Enterocolitis--the Importance of Breast Milk. J. Pediatr. Surg. 1974, 9, 587–595. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, R.; Liang, H.; An, J.; Yang, Y.; Huo, J.; Chen, Z.; Quan, W.; Jiang, L.; Li, C.; et al. Comparison and Investigation of Exosomes from Human Amniotic Fluid Stem Cells and Human Breast Milk in Alleviating Neonatal Necrotizing Enterocolitis. Stem Cell Rev. Rep. 2023, 19, 754–766. [Google Scholar] [CrossRef]
- Silachev, D.N.; Goryunov, K.V.; Shpilyuk, M.A.; Beznoschenko, O.S.; Morozova, N.Y.; Kraevaya, E.E.; Popkov, V.A.; Pevzner, I.B.; Zorova, L.D.; Evtushenko, E.A.; et al. Effect of MSCs and MSC-Derived Extracellular Vesicles on Human Blood Coagulation. Cells 2019, 8, 258. [Google Scholar] [CrossRef]
- Guillamat-Prats, R. Role of Mesenchymal Stem/Stromal Cells in Coagulation. Int. J. Mol. Sci. 2022, 23, 10393. [Google Scholar] [CrossRef]
- Moll, G.; Ankrum, J.A.; Olson, S.D.; Nolta, J.A. Improved MSC Minimal Criteria to Maximize Patient Safety: A Call to Embrace Tissue Factor and Hemocompatibility Assessment of MSC Products. Stem Cells Transl. Med. 2022, 11, 2–13. [Google Scholar] [CrossRef]
- Unar, A.; Bertolino, L.; Patauner, F.; Gallo, R.; Durante-Mangoni, E. Pathophysiology of Disseminated Intravascular Coagulation in Sepsis: A Clinically Focused Overview. Cells 2023, 12, 2120. [Google Scholar] [CrossRef]
- Foley, J.H.; Conway, E.M. Cross Talk Pathways Between Coagulation and Inflammation. Circ. Res. 2016, 118, 1392–1408. [Google Scholar] [CrossRef] [PubMed]
- Unar, A.; Bertolino, L.; Patauner, F.; Gallo, R.; Durante-Mangoni, E. Decoding Sepsis-Induced Disseminated Intravascular Coagulation: A Comprehensive Review of Existing and Emerging Therapies. J. Clin. Med. 2023, 12, 6128. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.K.; Wallace, E.M.; Davis, P.G.; Malhotra, A.; Jacobs, S.E.; Hooper, S.B.; Lim, R. A Protocol for Cell Therapy Infusion in Neonates. Stem Cells Transl. Med. 2021, 10, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Herz, J.; Köster, C.; Reinboth, B.S.; Dzietko, M.; Hansen, W.; Sabir, H.; van Velthoven, C.; Bendix, I.; Felderhoff-Müser, U. Interaction between Hypothermia and Delayed Mesenchymal Stem Cell Therapy in Neonatal Hypoxic-Ischemic Brain Injury. Brain Behav. Immun. 2018, 70, 118–130. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| References | Sources of EVs | Model | Protocol of Administration | Dosing | Therapeutic Effects | Elucidated Mechanisms |
|---|---|---|---|---|---|---|
| HYPOXIC–ISCHEMIC ENCEPHALOPATHY | ||||||
| [245] 2016 | human bone marrow MSCs | the Rice–Vannucci model | total dose—2.4 × 1010 EVs route of administration—intravenous administrations per day—2 days—1 | 1 unit * | ↓ total number and duration of seizures ↓ pathological fluctuations of blood pressure | ↑ baroreflex-mediated heart rate response |
| [156] 2019 | human bone marrow MSCs | the Rice–Vannucci model | total dose—2.4 × 1010 EVs route of administration—intravenous administrations per day—2 days—1 | 1 unit | ↓ permeability of blood–brain barrier | ↑ Annexin A1/FPR in neonatal brain endothelial cells and microglia |
| [151] 2019 | human bone marrow MSCs | the Rice–Vannucci model | total dose—1.25 × 109 EVs route of administration—intranasal administrations per day—1 days—1 | 0.1 units | ↓ of tissue loss ↓ % of cell death ↓ microglial activation ↑ behavioral outcomes (negative geotaxis test) | |
| [93] 2019 | human Wharton’s jelly MSCs | the Rice–Vannucci model + intraperitoneal injection of LPS | total dose—325 µg of EV protein per animal route of administration—intranasal administrations per day—1 days—1 | 0.3 units | ↓ microgliosis ↓ neuroinflammation | ↓ LPS/TLR4 signaling in microglia |
| [246] 2020 | rat bone marrow MSCs (H2S preconditioning) | the Rice–Vannucci model | total dose—1.5 × 108 EVs route of administration—intracardial injection administrations per day—1 days—1 | 0.06 units | ↓ water content and infarct volume of the brain ↓ % of cell apoptosis ↑polarization toward the anti-inflammatory M2 phenotype ↑ memory function | ↑ miR-7b-5p ↓ FOS → ↓ Iba1+ in microglia |
| [152] 2020 | human bone marrow MSCs | the Rice–Vannucci model | total dose—2.7 × 108 EVs route of administration—intraperitoneal administrations per day—1 days—3 (1,3,5 post HI) | 0.03 units | ↓ striatal tissue loss ↓ M1 micro- and A1 astroglia activation ↑ neurogenensis and angiogenesis ↑ myelination | |
| [247] 2021 | mice bone marrow MSCs | the Rice–Vannucci model | total dose—100 µg of EV protein route of administration—intracardial injection administrations per day—1 days—1 | 0.1 units | ↓ HI-induced edema, infarction, infiltrating monocytes ↓ phagocytosis of viable neurons ↑ synaptic densities | ↓ p-NF-κB → ↓OPN → ↓ Iba1 in M1 microglia |
| [248] 2021 | mice bone marrow MSCs | the Rice–Vannucci model | total dose—5 µg of EV protein route of administration—intranasal administrations per day—1 days—1 | 0.005 units | ↓ injury volumes ↓ microglial activation ↓ neuroinflammation | ↓ Iba1 → ↓ Casp3 in microglia |
| [249] 2021 | rat primary astrocytes (P1) | the Rice–Vannucci model | total dose—2.5 µg of EVs protein route of administration—intraperitoneal administrations per day—1 days—1 | 0.0024 units | ↓ the area of cerebral infarction ↓ HIBD-induced neuronal apoptosis ↓ oxidative stress ↓ neuroinflammation ↑ body weight ↑ cognitive functions(grip test, negative geotaxis test | ↑ miR-17-5p → ↓ BNIP → ↓ Bax in brain tissue |
| [250] 2022 | brain tissues of neonatal mice (P9) after HI | the Rice–Vannucci model | total dose—8 × 109 EVs route of administration—intranasal administrations per day—2 days—1 | 0.066 units | ↓ infarct size ↓ Casp3 expression | ↑ miR-342-3p and miR-330-3p in brain tissue |
| [251] 2022 | mice bone marrow MSCs | the Rice–Vannucci model | total dose—2 × 109 EVs route of administration—intranasal administrations per day—1 days—1 | 0.2 units | ↑ animal survival ↓ infarct volume of brain ↓ % of apoptosis cells ↓ neuroinflammation ↑ proprioceptive function | ↑ miR-93 → ↓ JMJD3 → ↑ KLF2 → ↓ Casp3,Bax in neurons |
| [252] 2023 | immortalized human bone marrow MSCs | the Rice–Vannucci model | total dose—2.7 × 108 EVs route of administration—intranasal administrations per day—1 days—3 (1,3,5 post HI) | 0.03 units | ↑ neurogenesis and angiogenesis ↓ monocyte infiltration ↓ astrogliosis and microgliosis | |
| BRONCHOPULMONARY DYSPLASIA | ||||||
| [167] 2018 | human Wharton’s jelly MSCs | hyperoxia (HYRX)-induced BPD mice model (P1–P7 75% O2) | total dose—0.9 µg of EV protein route of administration—intravenous administrations per day—1 days—1 | 0.001 units | ↑ lung architecture ↓ lung fibrosis ↓ peripheral pulmonary arterial remodeling | |
| [175] 2018 | preterm human Wharton’s jelly MSCs | HYRX-induced BPD mice model (c P1–P4 95% O2) | total dose—4.5 × 108 EVs route of administration—intraperitoneal administrations per day—1 days—2 (P2 and P4) | 0.038 units | ↑ lung architecture ↓ infiltration of neutrophils ↓ pulmonary hypertension ↓ alveolar-capillary leak | ↑ TSG-6 signaling in lung tissue |
| [253] 2018 | human Wharton’s jelly MSCs | HYRX-induced BPD rat model (P1–P14 60% O2) | total dose—0.213 × 1010 EVs route of administration—intratracheal administrations per day—1 days—3 (P3, P7 and P10) | 0.27 units | ↑ alveolar development ↓ pulmonary vascular remodeling | |
| [176] 2018 | rat bone marrow MSCs | HYRX-induced BPD rat model (P0–P14 85% O2) | total dose—3.4 × 109 EVs route of administration—intraperitoneal administrations per day—1 days—14 (P1–P15) | 1.96 units | ↑ alveolar growth ↑ lung blood vessel density ↓ pulmonary hypertension | ↑ VEGF signaling in lung tissue |
| [177] 2018 | human umbilical cord blood MSCs | HYRX-induced BPD rat model (P1–P14 90% O2) | total dose—20 µg of EV protein route of administration—intratracheal administrations per day—1 Days—1 (P5) | 0.019 units | ↑ alveolarization and angiogenesis | ↑ VEGF signaling in lung tissue |
| [254] 2020 | human Wharton’s jelly MSCs | HYRX-induced BPD mice model (P0–P14 75% O2) | total dose—6 × 108 EVs route of administration—intravenous administrations per day—1 days—1 (PN4) | 0.025 units | ↓ alveolar simplification ↓ septal collagen disposition ↑ blood vessel count ↓ pulmonary hypertension ↑ functional exercise capacity | |
| [255] 2021 | human bone marrow MSCs. | In utero induced BPD rat model (antenatal injection of E. coli endotoxin e20) | total dose—0.25 × 106 EVs route of administration— intra-amniotic administrations per day—10 per pregnant rat days—1 (e20) | 0.17 units | ↓ lung simplification ↑ vascularization ↓ pulmonary hypertension ↑ lung mechanical function | |
| [174] 2021 | human Wharton’s jelly MSCs | HYRX-induced BPD mice model (P–P7 75% O2) | total dose—6 × 108 EVs route of administration—intravenous administrations per day—1 days—1 (PN4) | 0.025 units | ↑ thymic development ↑ proportion of CD4+FoxP3+ regulatory T cells ↓ alveolar simplification | |
| [253] 2021 | human umbilical cord blood MSCs | HYRX-induced BPD rat model (P1–P14 60% O2) | total dose—0.64 × 1010 EVs route of administration—intratracheal administrations per day—1 days—4 (P3, P7, P10 and P21) | 0.27 units | ↑ alveolar development ↓ deposition of fibrous tissue ↑ density of M2 macrophages ↓ pulmonary hypertension | |
| [111] 2021 | amniotic fluid-derived EVs (full-term cesarean sections) | HYRX-induced BPD rat model (P1–P14 85% O2) | total dose—1 × 1010 EVs route of administration—intratracheal administrations per day—1 days—1 (P3) | 0.42 units | ↑ alveolar development ↓ pulmonary hypertension | |
| [256] 2022 | human breast milk-derived EVs | HYRX-induced BPD rat model (P1–P7 85% O2) | total dose—140 µg of EV protein route of administration—intragastric administrations per day—1 days—1 (PN7) | 0.133 units | ↓ lung tissue collapse ↓ cleaved caspase 3 | ↓ IL-17/↓ FADD in Type II alveolar epitheliocytes |
| [171] 2022 | human Wharton’s jelly MSCs | HYRX-induced BPD rat model (P1–P14 85% O2) | total dose—96 × 108 EVs route of administration—intratracheal administrations per day—1 days—1 (PN3) | 0.04 units | ↑ lung vascular density and alveolar structure ↓ lung inflammation ↓ pulmonary hypertension | ↑ VEGF/eNOS in lung tissue |
| [182] 2022 | human amniotic epithelial cells (term birth after caesarean sections) | in utero induced BPD mice model (injection of LPS e16) + (P3.5–P28 65% O2) | total dose—10 µg of EV protein route of administration—intravenous administrations per day—1 days—1 (PN4) | 0.01 units | ↑ lung tissue-to-air space ratio ↓ lung inflammation ↑ type II alveolar epithelial cell ↓ pulmonary hypertension ↑ lung tissue elasticity | |
| [179] 2022 | human Wharton’s jelly MSCs | HYRX-induced BPD mice model (injection of LPS P7/P8) + 40% O2 P10 | total dose—1 × 106 EVs route of administration—intratracheal administrations per day—1 days—1 (PN9) | 0.0002 units | ↑ lung architecture ↑ blood vessel density ↑ mRNA of antiinflammatory cytokines in lung tissue | |
| [257] 2023 | human umbilical cord blood MSCs | HYRX-induced BPD mice model (P1–P14 85% O2) | total dose—15 × 105 EVs route of administration—intraperitoneal administrations per day—1 days—3 (P4–P6) | 0.000063 units | ↓ lung fibrosis ↑ vascular development | ↑ miR-185-5p →↓ CDK6 → ↑ angiogenesis in lung tissue |
| [169] 2023 | human bone marrow MSCs | hypoxia—induced BPD rat model (10 min 40% O2 + 2 min 1% O2 12 times daily P1–P14) | total dose—2 × 105 EVs route of administration—intraperitoneal administrations per day—1 days—14 (P1–P14) | 0.00012 units | ↓ simplified alveolar structure ↓ pulmonary hypertension ↑ capillary distribution ↑ respiratory efficiency ↓ oxidative stress | ↑ PI3K/AKT → ↑ SOD in lung tisue |
| NECROTIZING ENTEROCOLITIS | ||||||
| [197] 2016 | mice bone marrow MSCs | NEC—induced preterm mice model, Barlow et al. [258] (21e) + 90 s 100% N2 + 4 °C 10 min twice daily (P1–P4) | total dose—2.5 × 109 EVs route of administration—intraperitoneal administrations per day—1 days—1 (prior NEC) | 0.1 units | ↓ the overall incidence of NEC ↑ gut barrier function | |
| [107] 2018 | neonatal mice enteric neuronal stem cells | NEC—induced preterm rat model, Barlow et al. [258] (21e) + (90 s 100% N2 + 4 °C 10 min every 8 h + LPS every 4 h (P1–P4) | total dose—4 × 108 EVs route of administration—intraperitoneal administrations per day—1 days—1 (prior NEC) | 0.017 units | ↓ intestine villus destruction ↓ the overall incidence of NEC | |
| [231] 2019 | bovine milk-derived EVs | NEC—induced preterm mice model, Barlow et al. [258] (10 min 5% O2 3 times between P5–P9 + LPS 4 times between P6 and P7) | total dose—1.2 mg of EV protein route of administration—intragastric via gavage administrations per day—3 days—5 (P5–P9) | 1.14 units | ↑ intestine villus destruction ↑ number of goblet cells ↓ intestinal mucosal inflammation ↓ oxidative stress | |
| [227] 2019 | human breast milk-derived EVs | NEC—induced preterm rat model, Barlow et al. [258] (21e) 90 s 1.5% O2 + 4 °C 10 min 3 times daily P1–P4 + LPS 1 time P1 | total dose—2.4 × 1010 EVs route of administration—intragastric via gavage administrations per day—6 days—4 (P1–P4) | 1 unit | ↓ villus destruction ↓ the overall incidence of NEC | |
| [194] 2020 | rat amniotic fluid CD117 stem cells (e14.5) | NEC—induced preterm mice model, Barlow et al. [258] (10 min 5% O2 3 times between P5–P9 + LPS 4 times between P6–P7) | total dose—3.5 × 108 EVs route of administration—intraperitoneal administrations per day—1 days—2 (P6–P7) | 0.015 units | ↑ gut epithelial regeneration ↓ intestinal inflammation | ↑ Wnt/β-catenin → increased intestinal epithelial proliferation |
| [259] 2022 | human breast milk-derived EVs | NEC—induced preterm mice model, Barlow et al. [258] (1 min 100% N2 + 4 °C 5 min twice a day P6–P10) | total dose—30 µg of EV protein route of administration—intragastric via gavage administrations per day—3 days—3 (P8–P10) | 0.04 units | ↑ ileal crypts number ↓ inflammatory microenvironment | |
| NEONATAL MENINGITIS | ||||||
| [190] 2022 | human Wharton’s jelly-derived MSCs after thrombin preconditioning | Escherichia coli—induced meningitis in newborn rats (P11) | total dose—2.6 × 107 EVs route of administration—intraventricular administrations per day—1 days—1 (P11) | 0.001 units | ↓ neural cell death ↓ number of active microglia ↓ levels of inflammatory cytokines in brain tissues | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goryunov, K.; Ivanov, M.; Kulikov, A.; Shevtsova, Y.; Burov, A.; Podurovskaya, Y.; Zubkov, V.; Degtyarev, D.; Sukhikh, G.; Silachev, D. A Review of the Use of Extracellular Vesicles in the Treatment of Neonatal Diseases: Current State and Problems with Translation to the Clinic. Int. J. Mol. Sci. 2024, 25, 2879. https://doi.org/10.3390/ijms25052879
Goryunov K, Ivanov M, Kulikov A, Shevtsova Y, Burov A, Podurovskaya Y, Zubkov V, Degtyarev D, Sukhikh G, Silachev D. A Review of the Use of Extracellular Vesicles in the Treatment of Neonatal Diseases: Current State and Problems with Translation to the Clinic. International Journal of Molecular Sciences. 2024; 25(5):2879. https://doi.org/10.3390/ijms25052879
Chicago/Turabian StyleGoryunov, Kirill, Mikhail Ivanov, Andrey Kulikov, Yulia Shevtsova, Artem Burov, Yulia Podurovskaya, Victor Zubkov, Dmitry Degtyarev, Gennady Sukhikh, and Denis Silachev. 2024. "A Review of the Use of Extracellular Vesicles in the Treatment of Neonatal Diseases: Current State and Problems with Translation to the Clinic" International Journal of Molecular Sciences 25, no. 5: 2879. https://doi.org/10.3390/ijms25052879
APA StyleGoryunov, K., Ivanov, M., Kulikov, A., Shevtsova, Y., Burov, A., Podurovskaya, Y., Zubkov, V., Degtyarev, D., Sukhikh, G., & Silachev, D. (2024). A Review of the Use of Extracellular Vesicles in the Treatment of Neonatal Diseases: Current State and Problems with Translation to the Clinic. International Journal of Molecular Sciences, 25(5), 2879. https://doi.org/10.3390/ijms25052879