
Effects of Potassium-Containing Fertilizers on Sugar and Organic Acid Metabolism in Grape Fruits

 , and
, and
Abstract
1. Introduction
2. Results
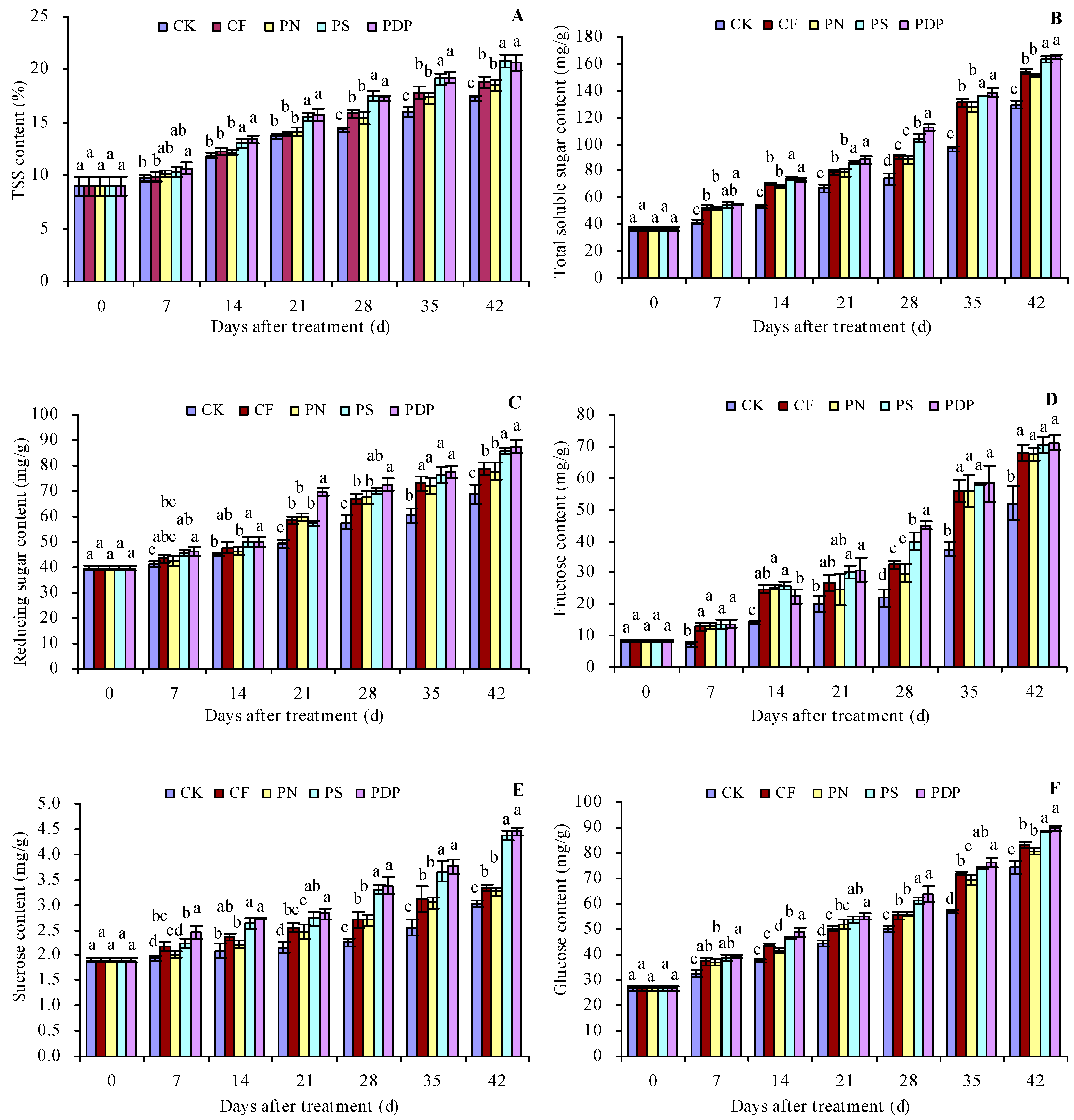
2.1. Sugar Content in the Grape Fruits
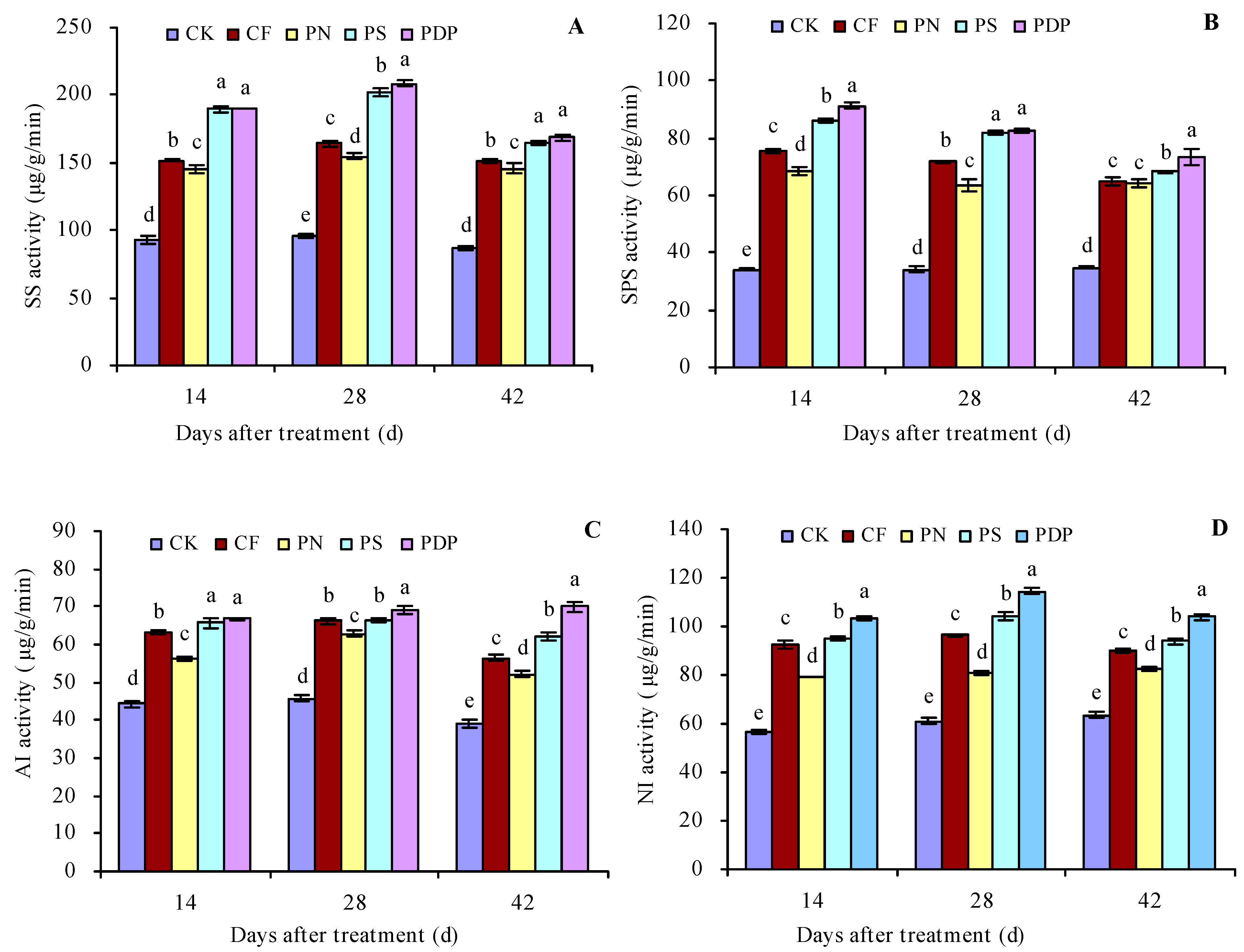
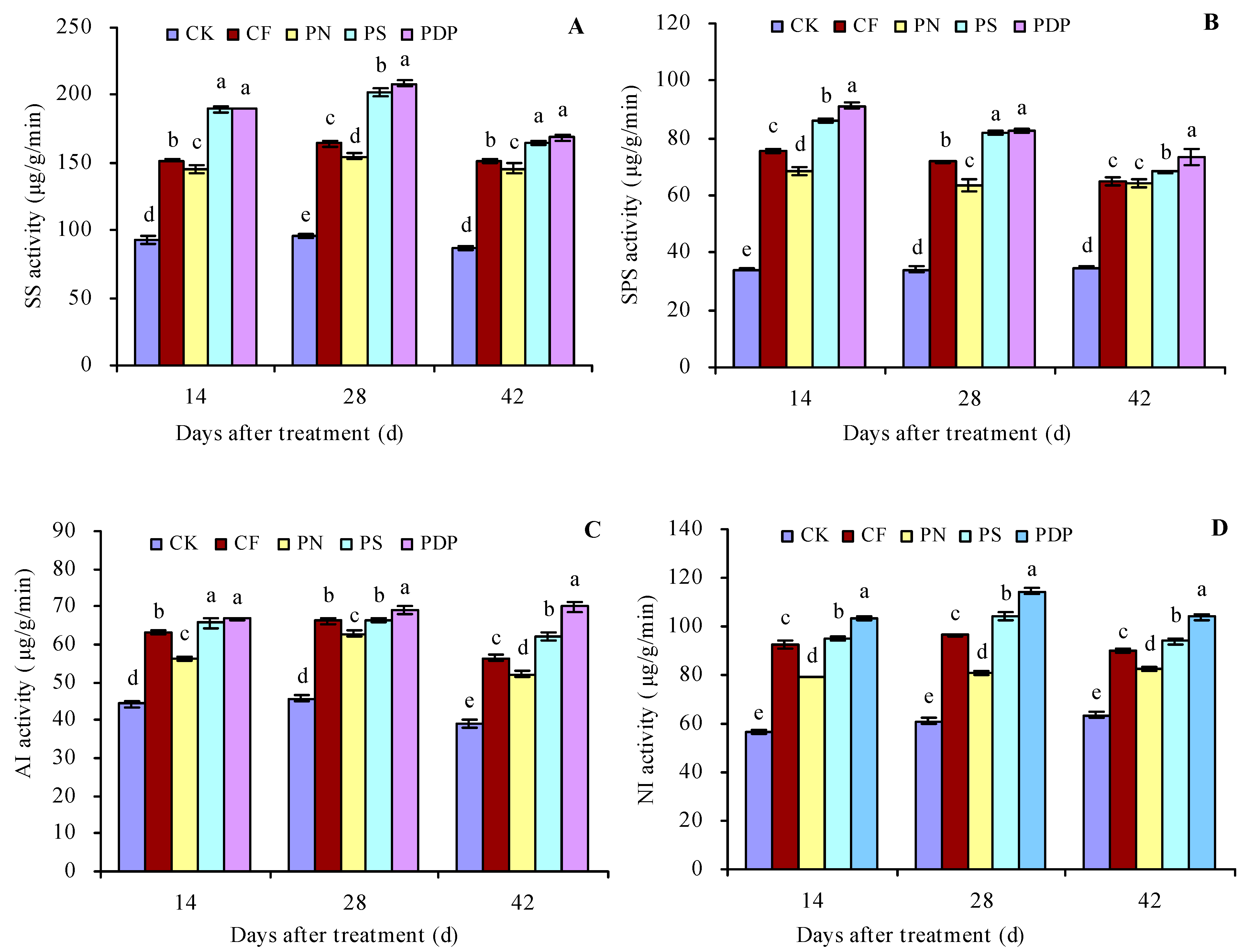
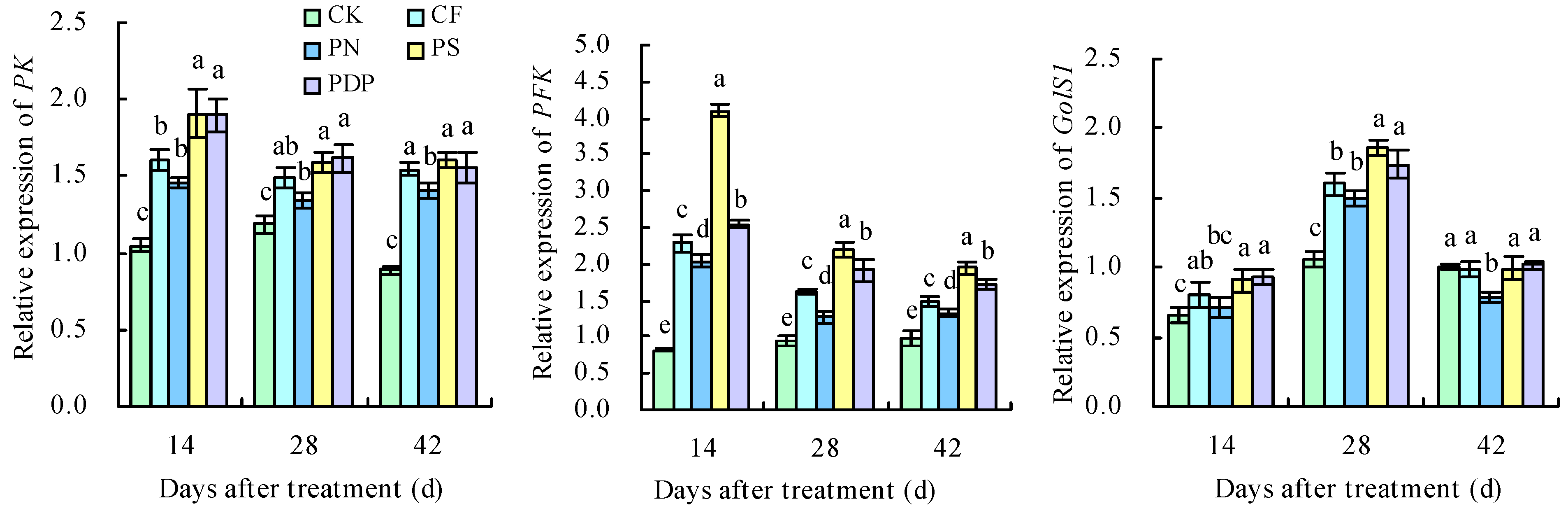
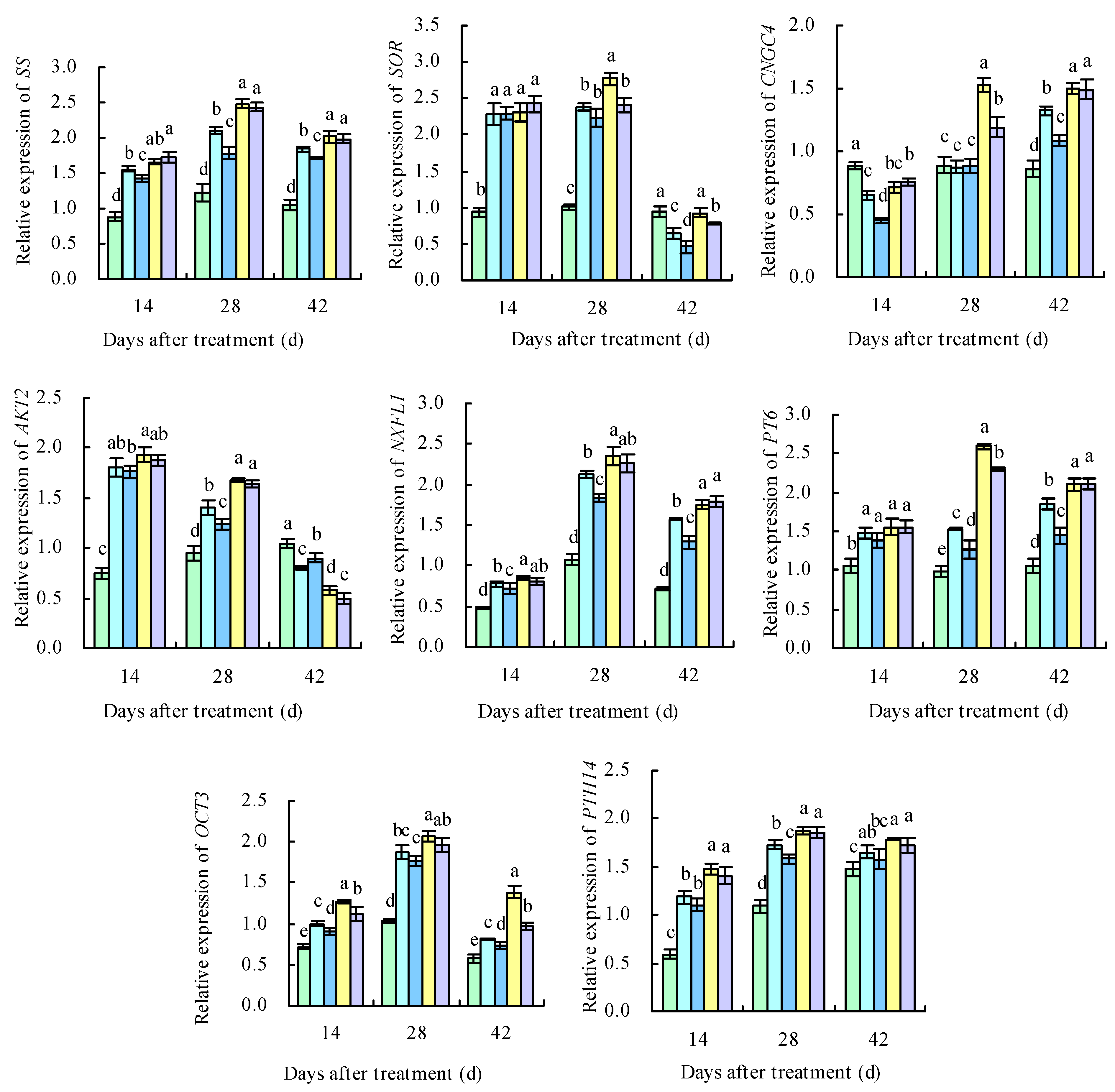
2.2. Sugar Metabolism-Related Enzyme Activity in Grape Fruits
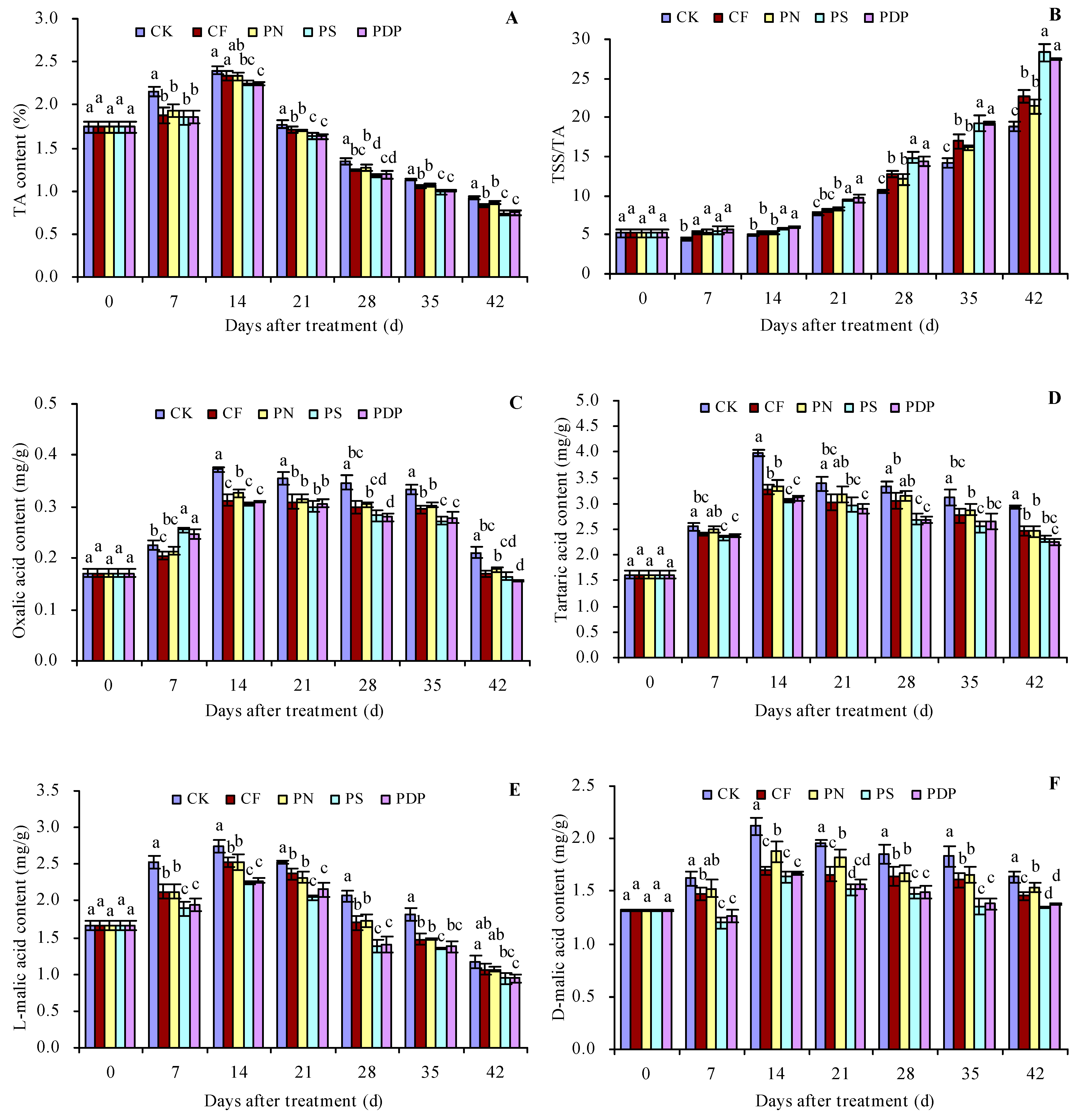
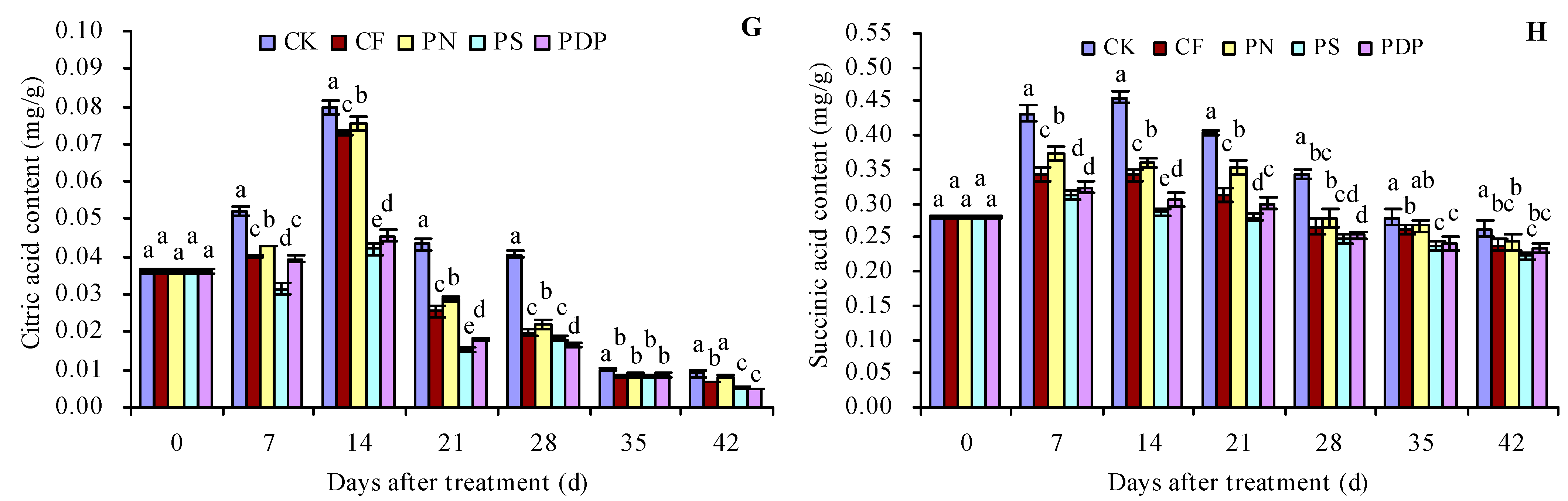
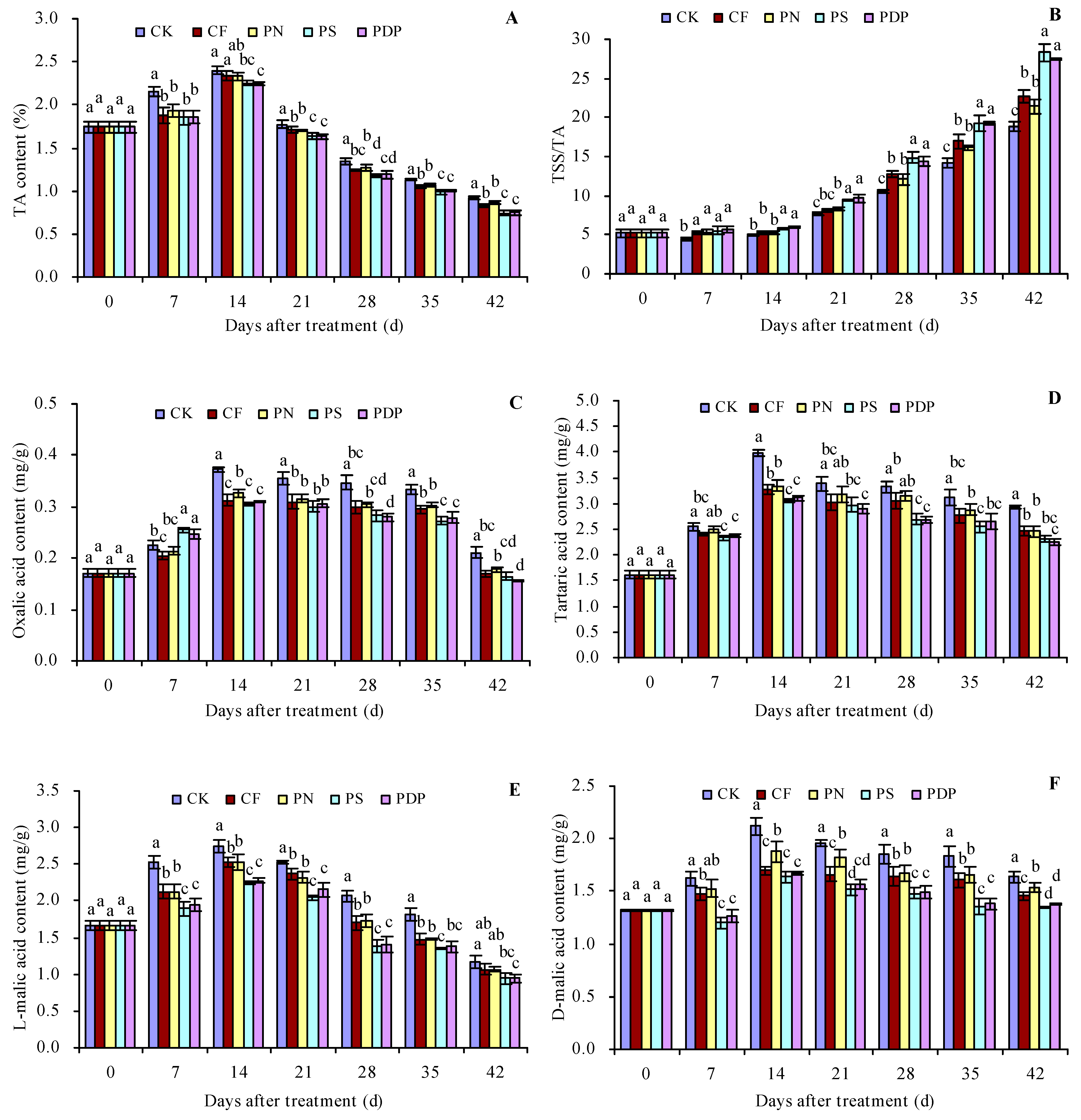
2.3. Organic Acid Content in Grape Fruits
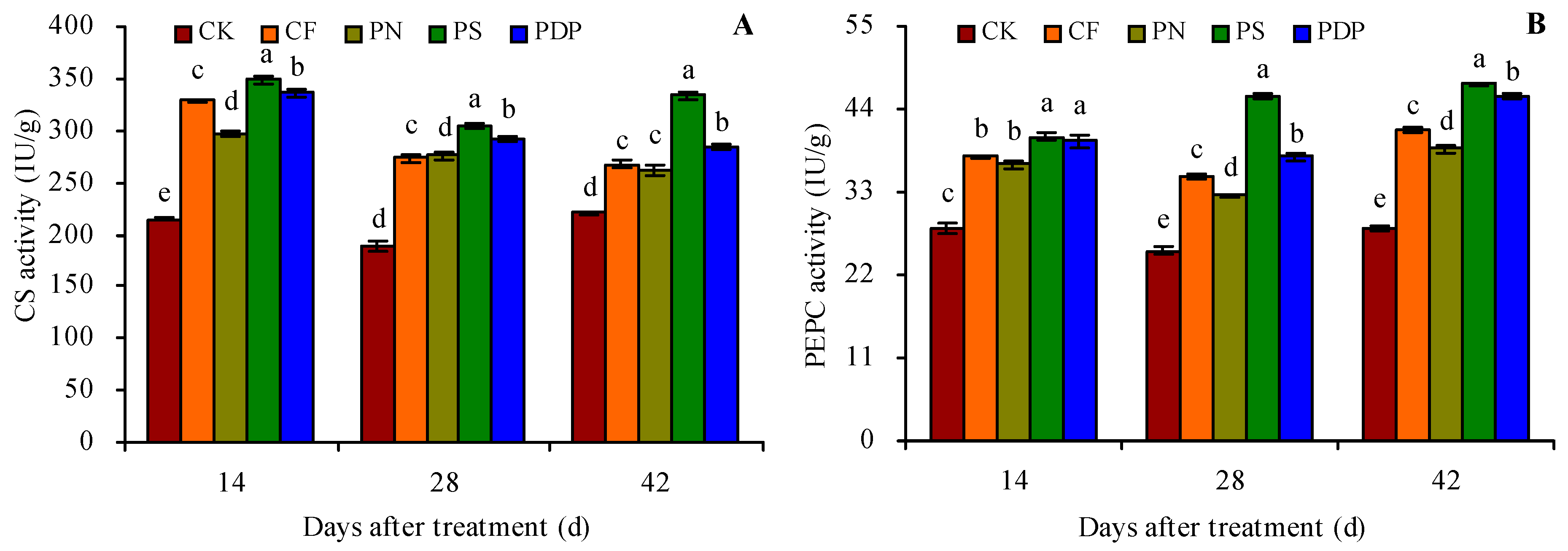
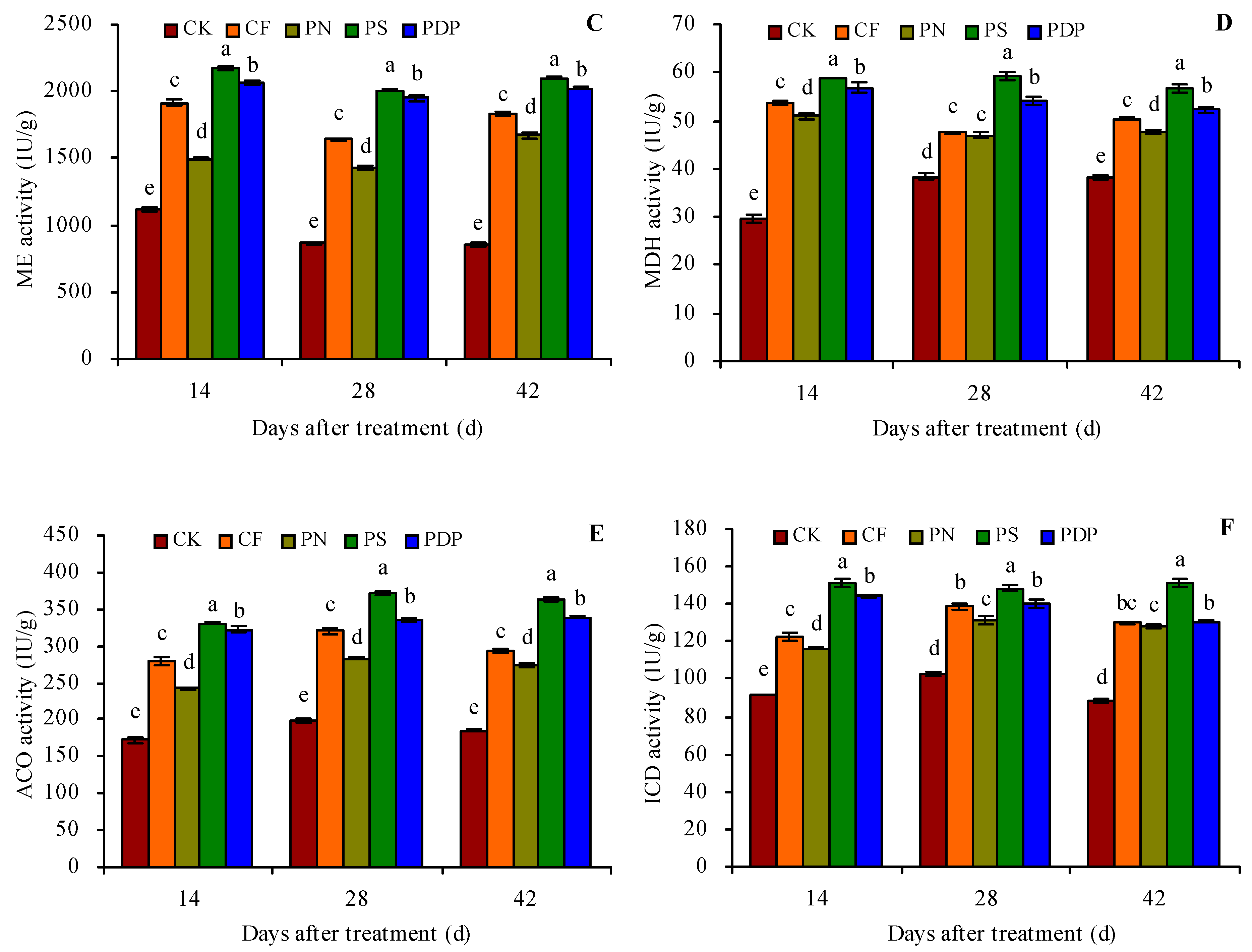
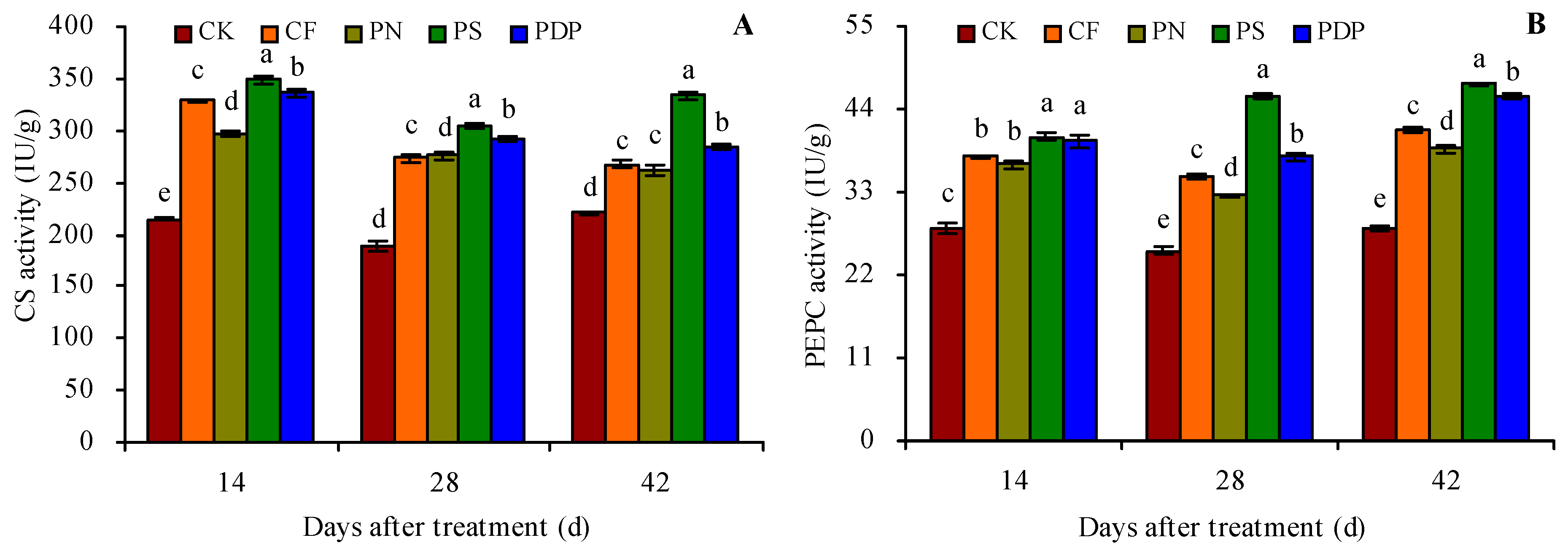
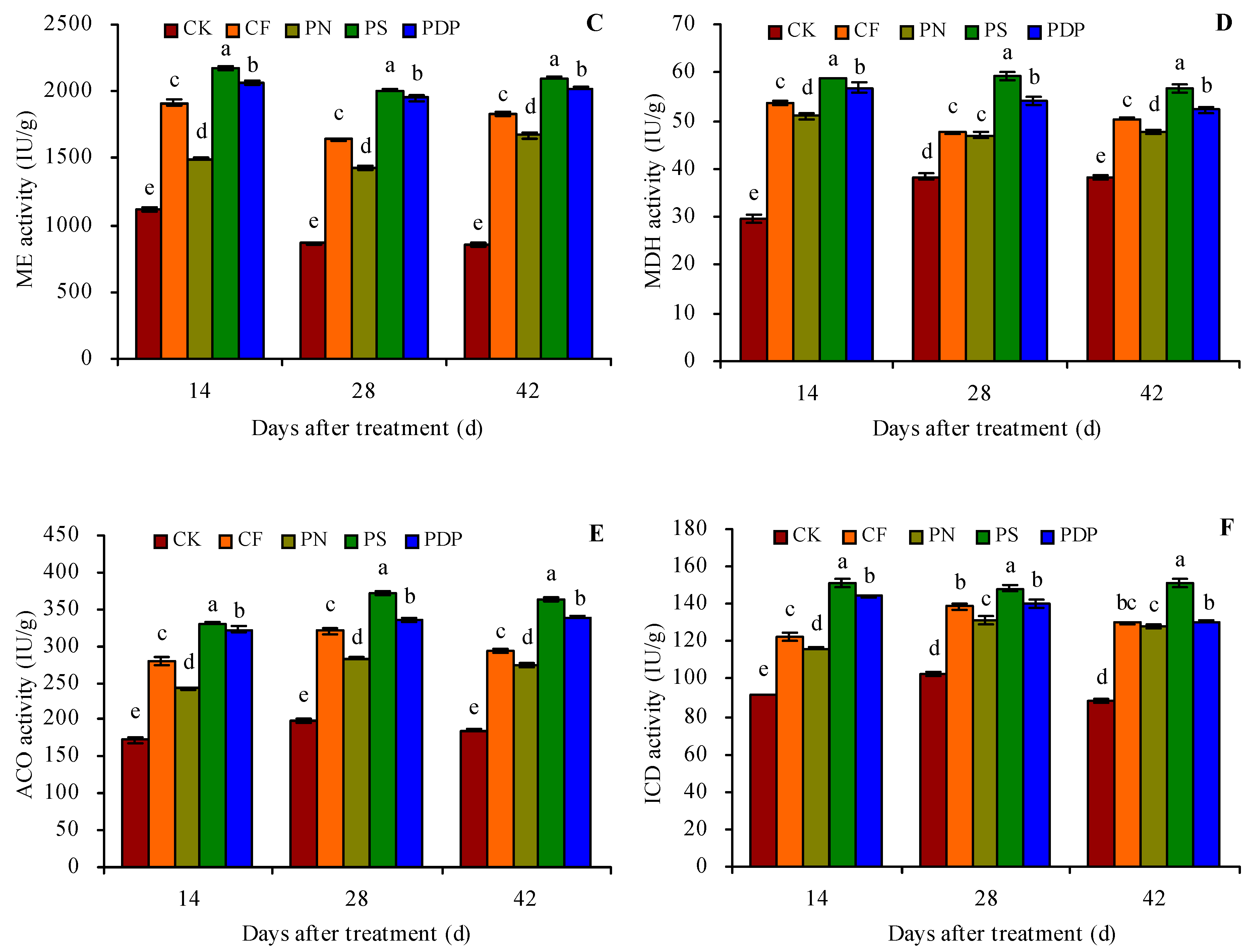
2.4. Organic Acid Metabolism-Related Enzyme Activity in Grape Fruits
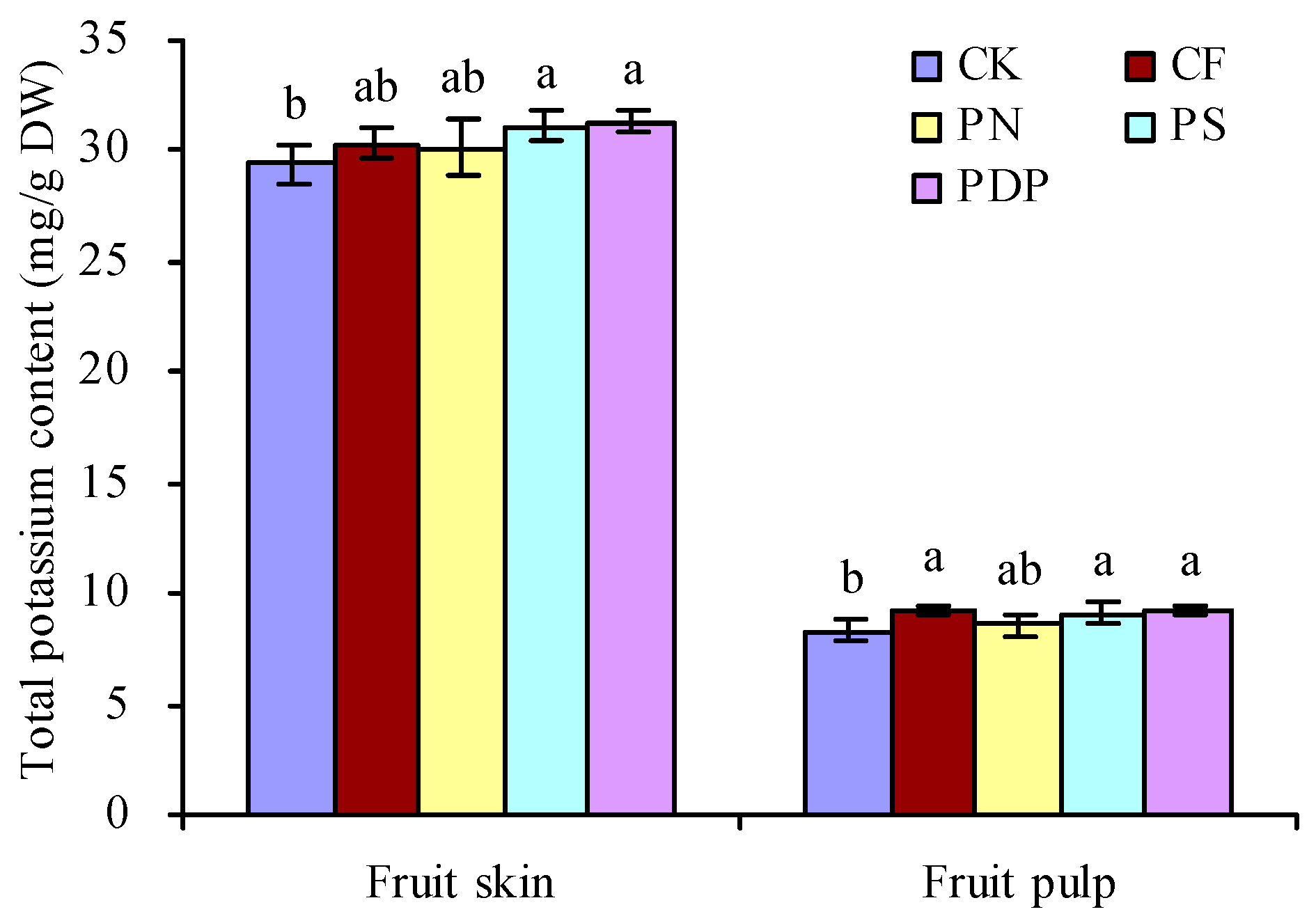
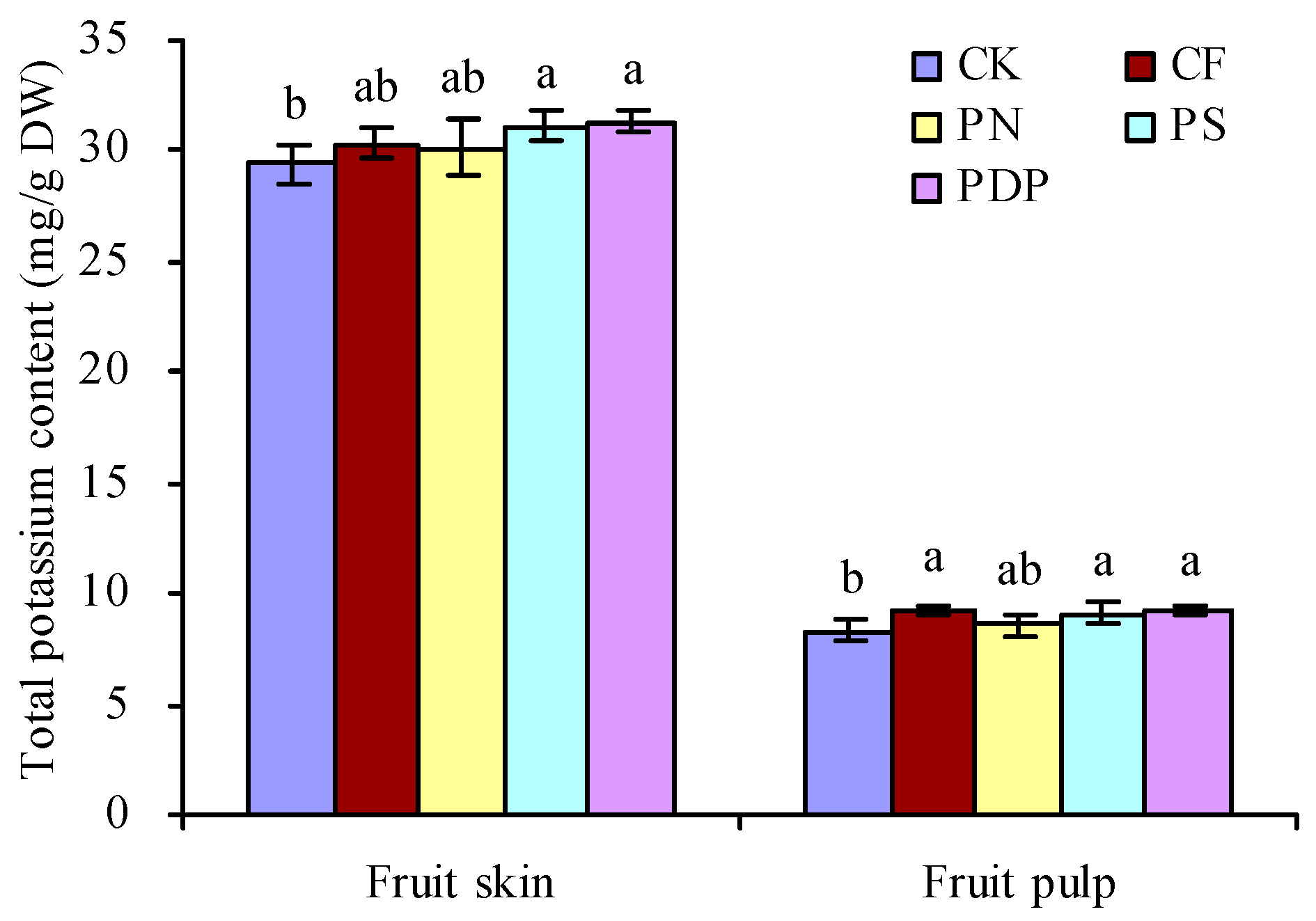
2.5. Total Potassium Content in Grape Fruits
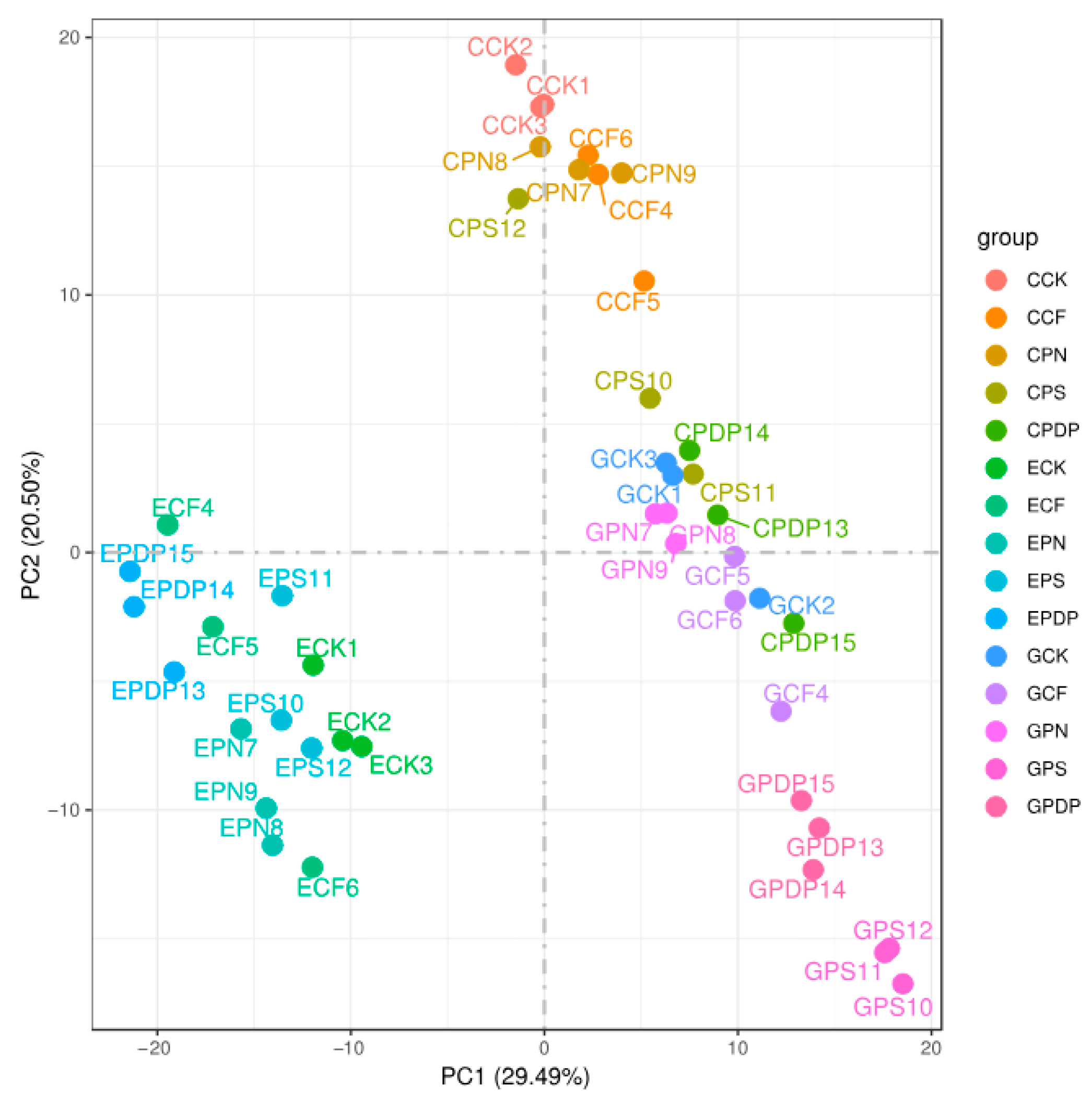
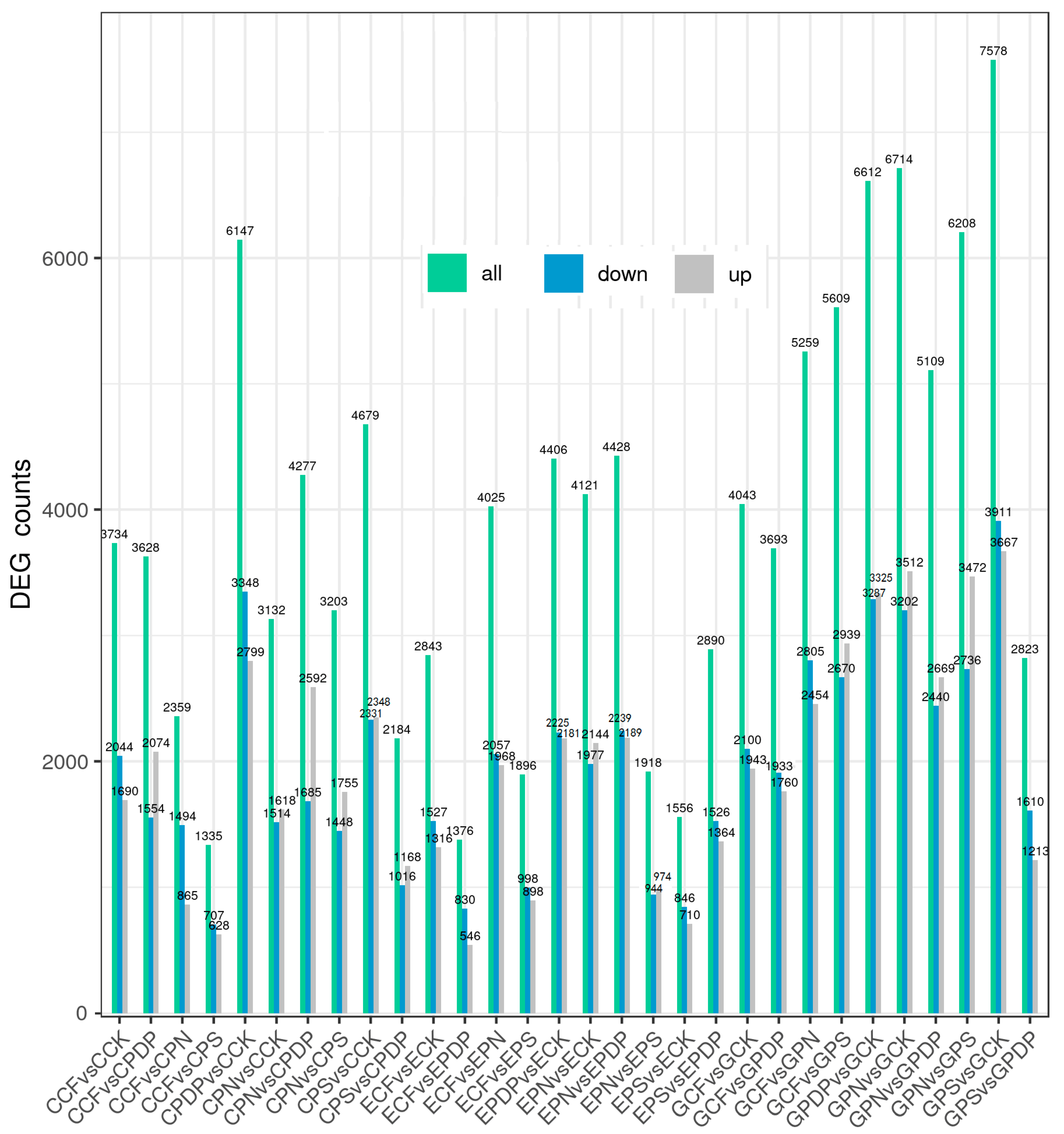
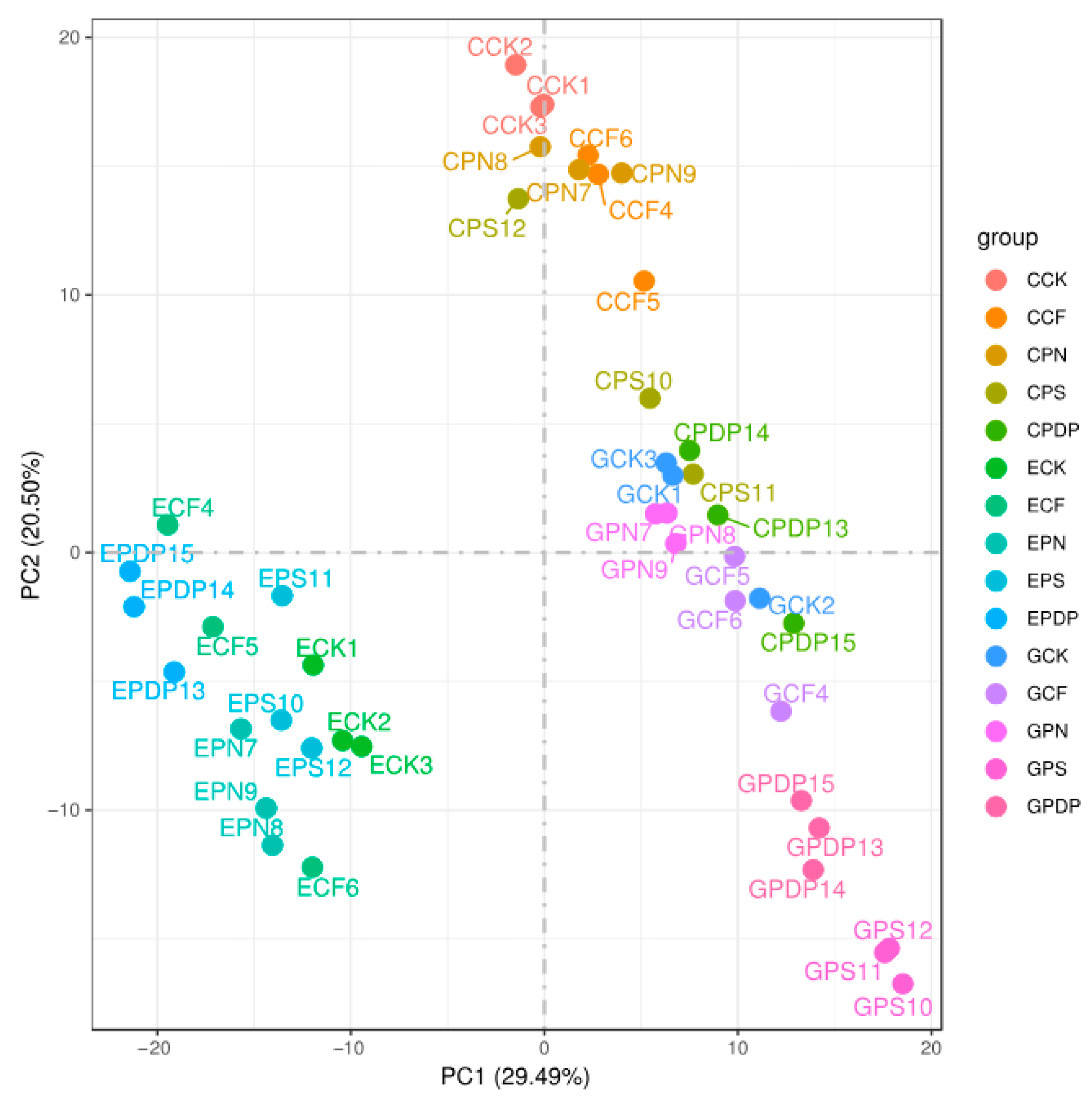
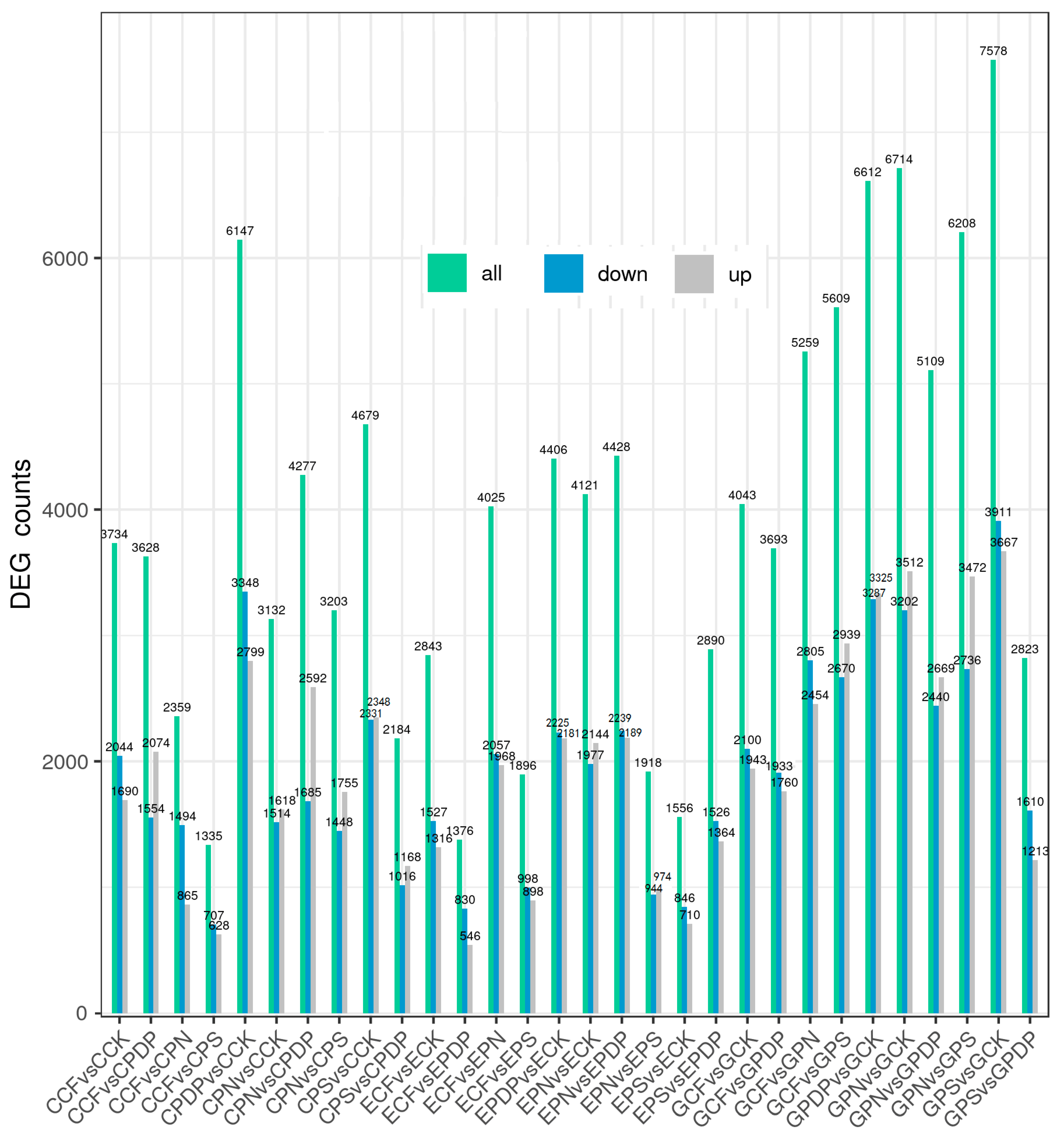
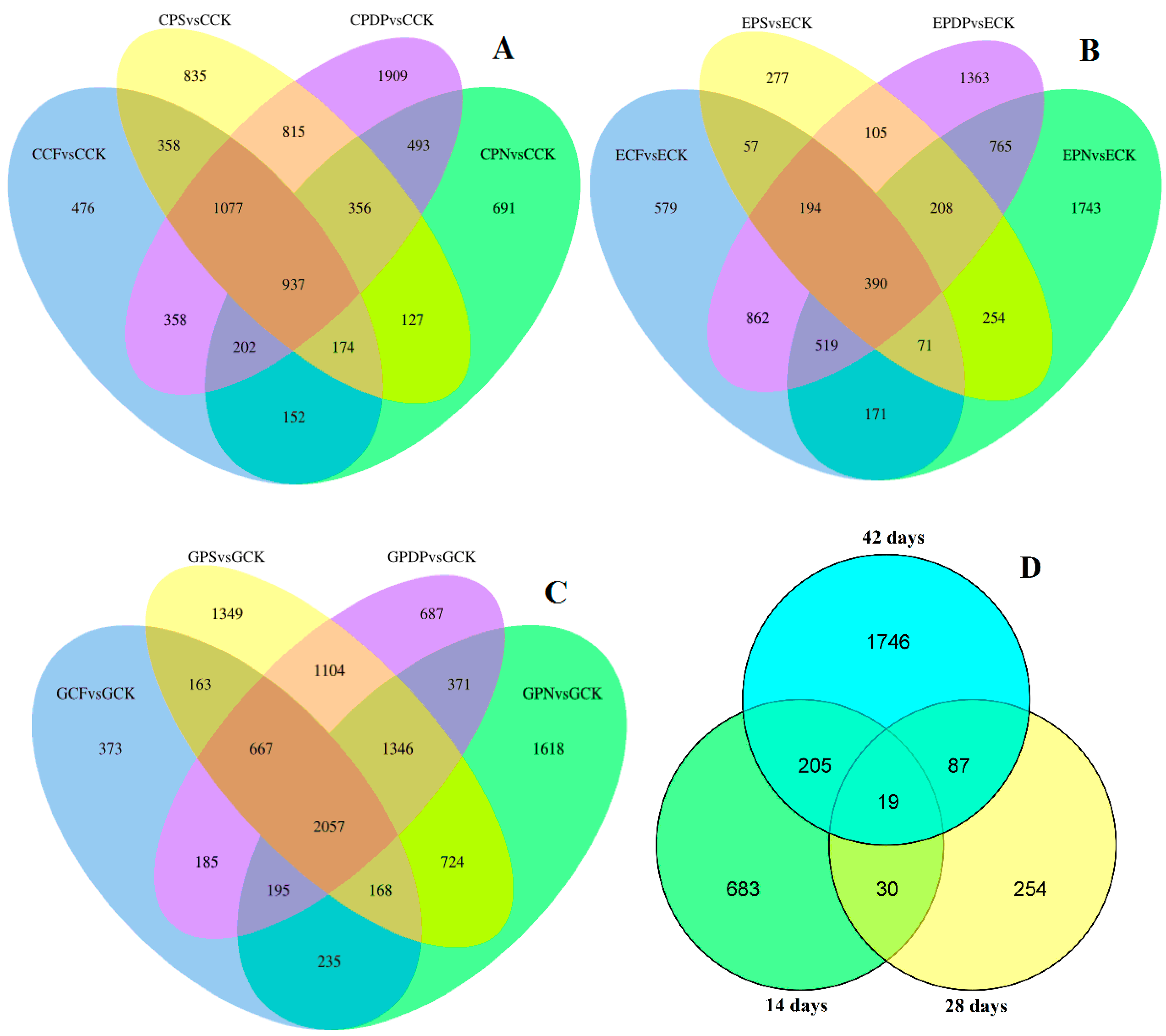
2.6. DEG Analysis following Potassium-Containing Fertilizer Treatment
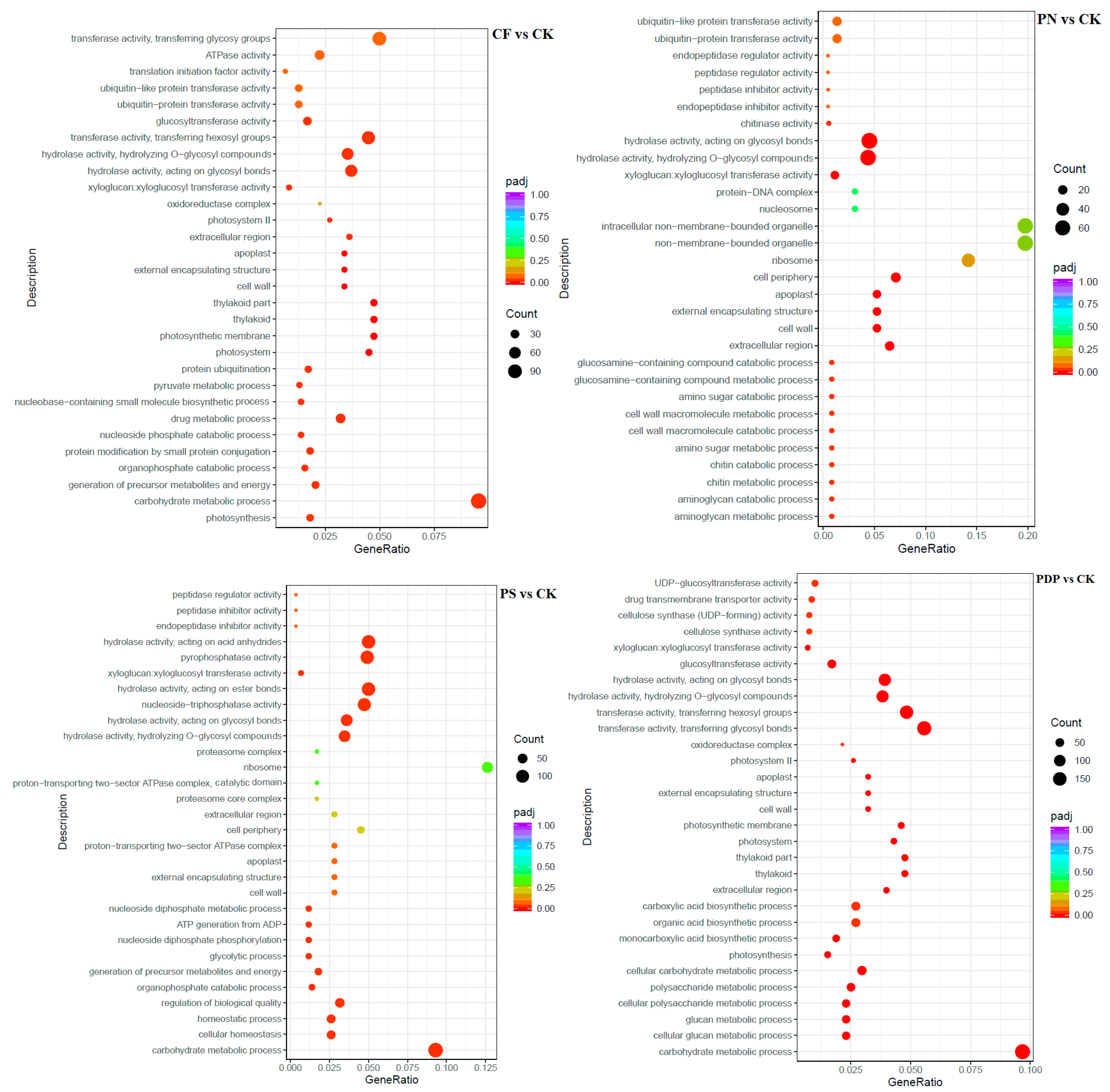
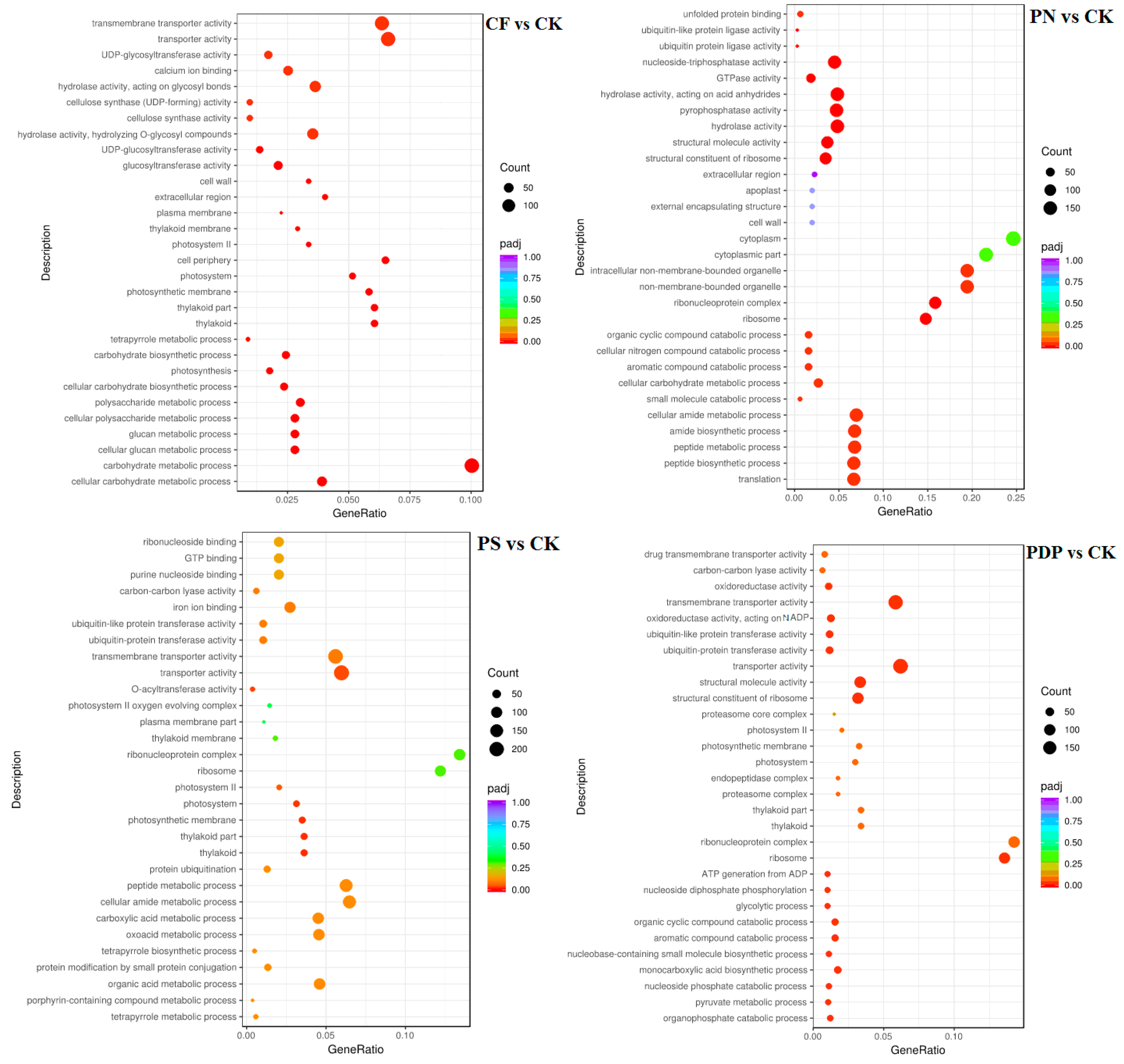
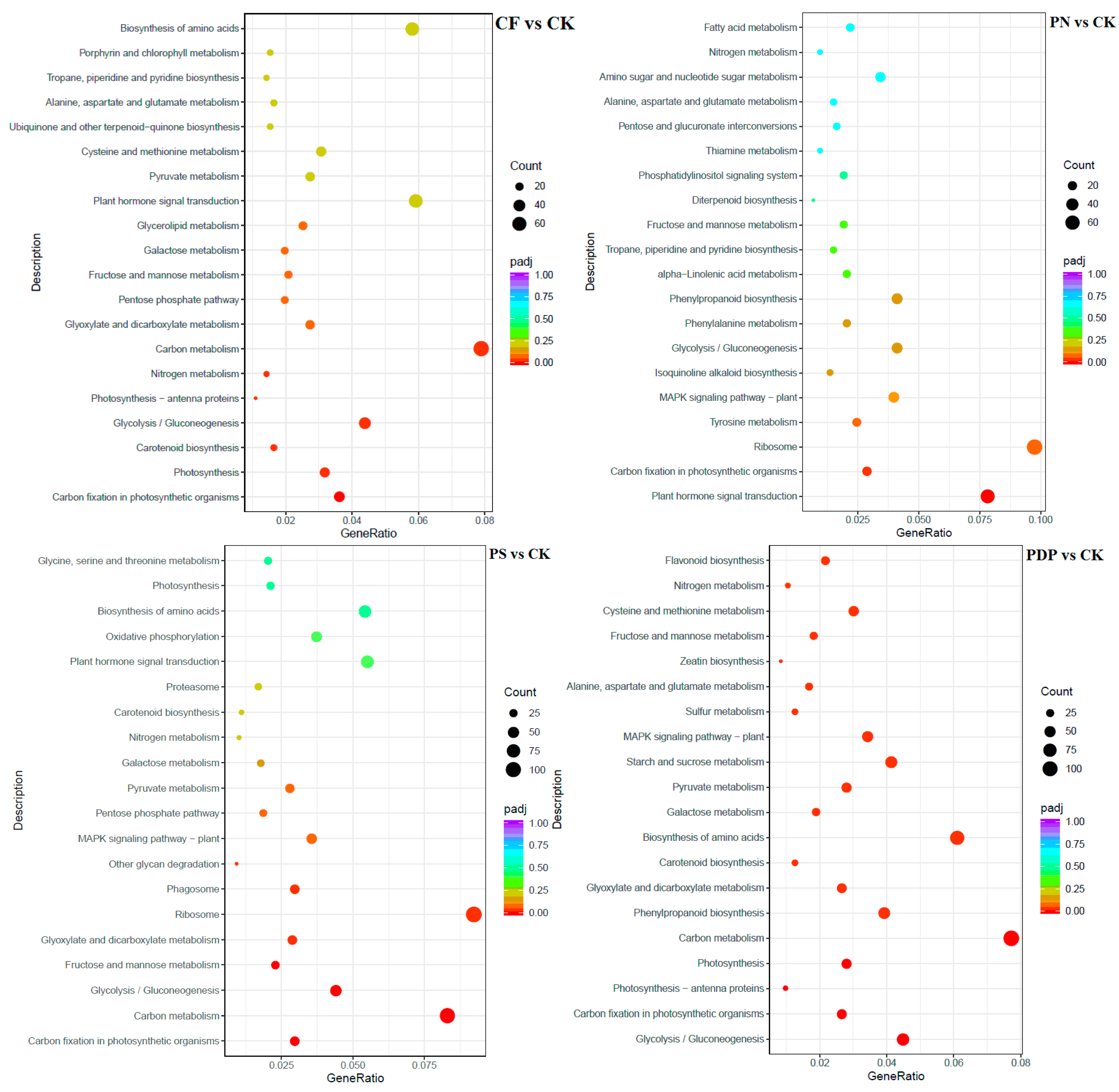
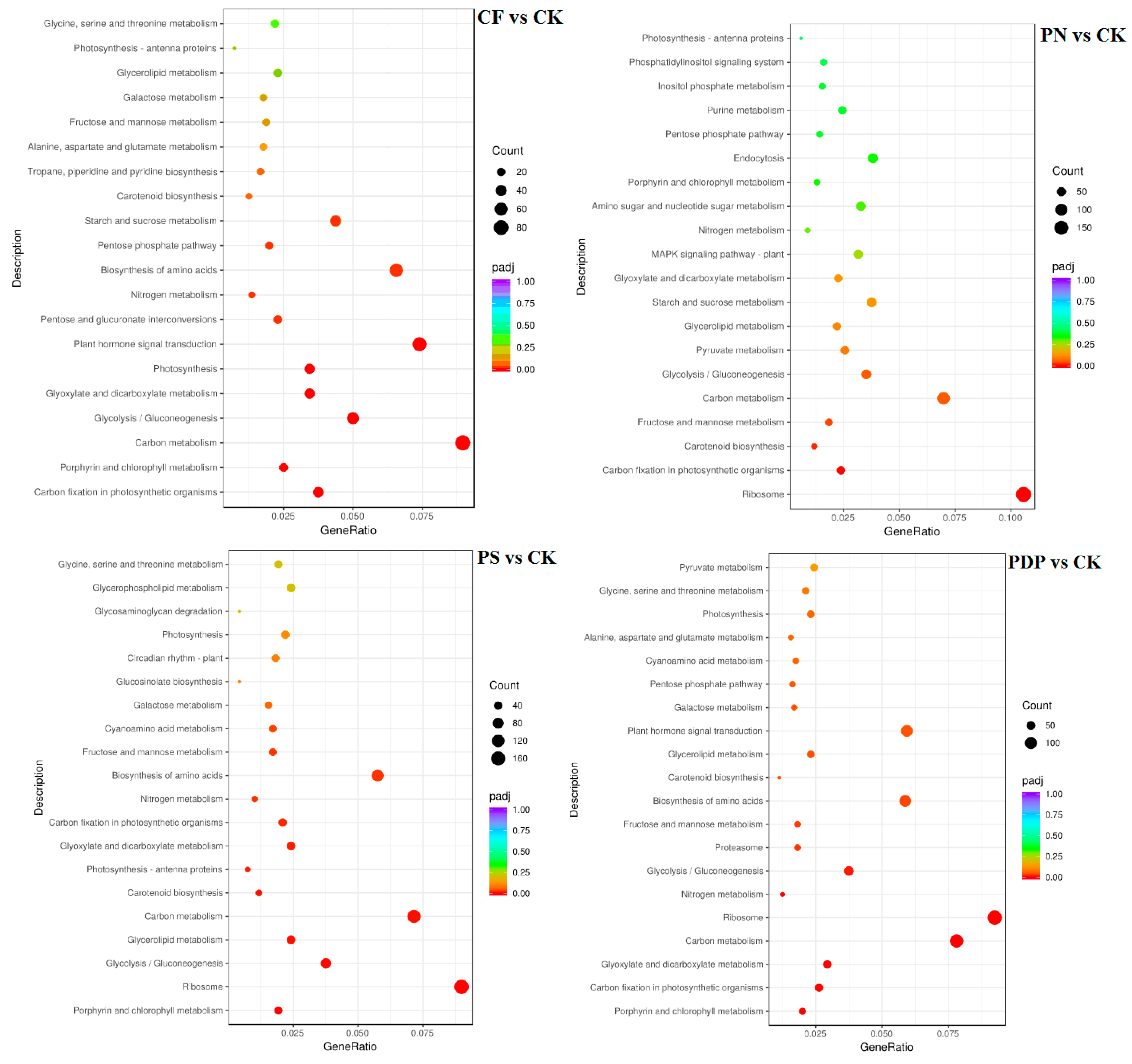
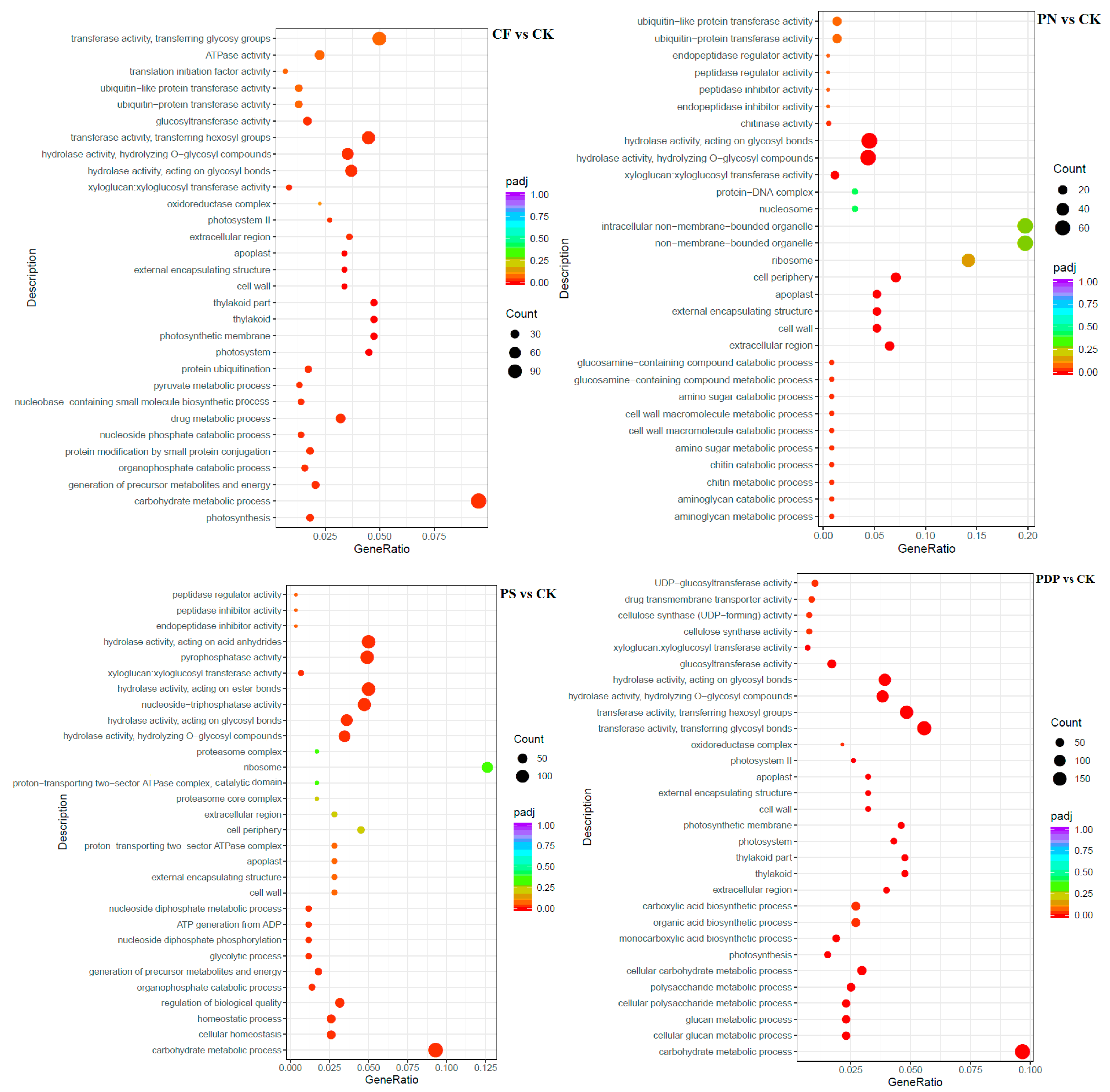
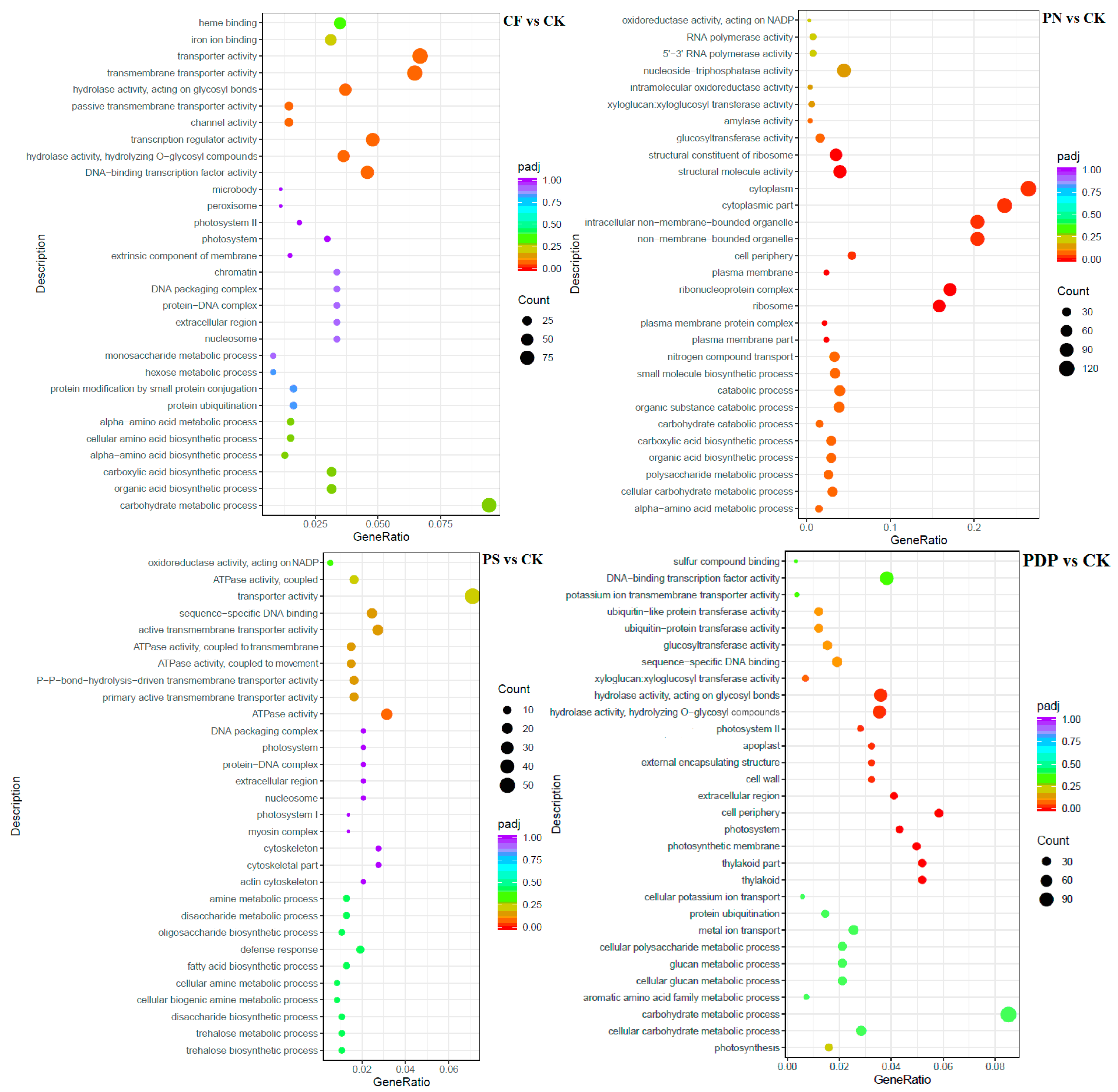
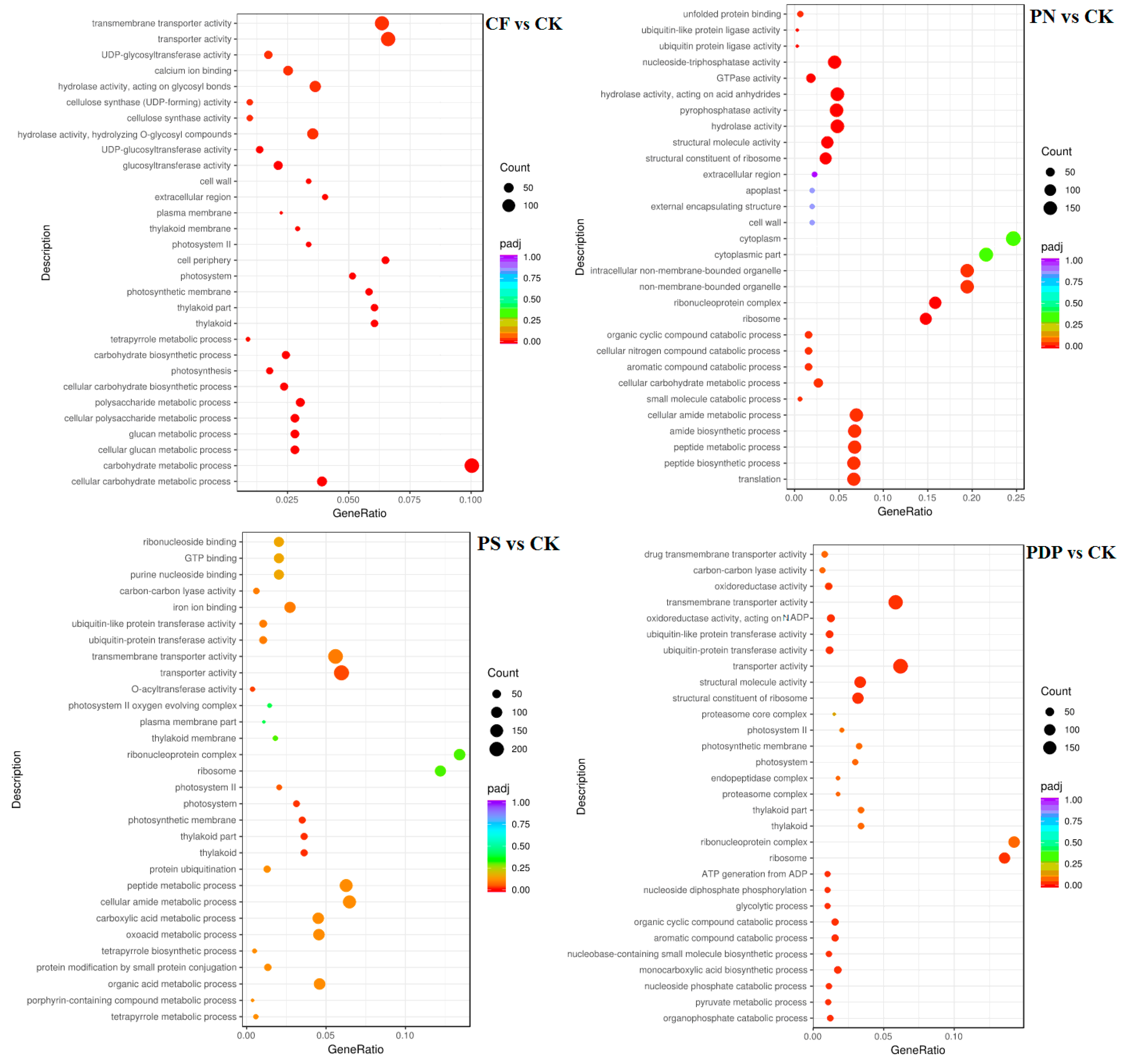
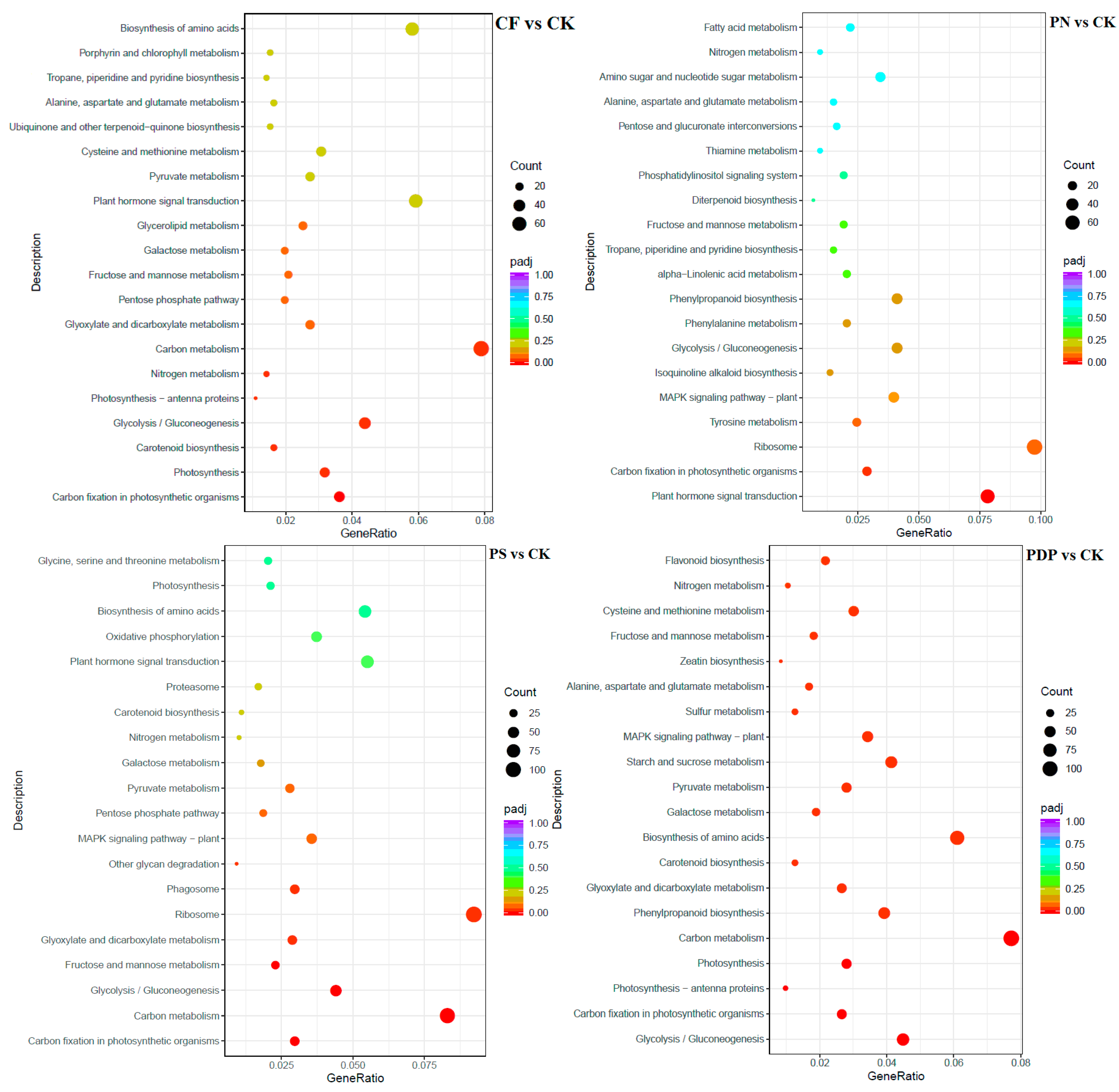
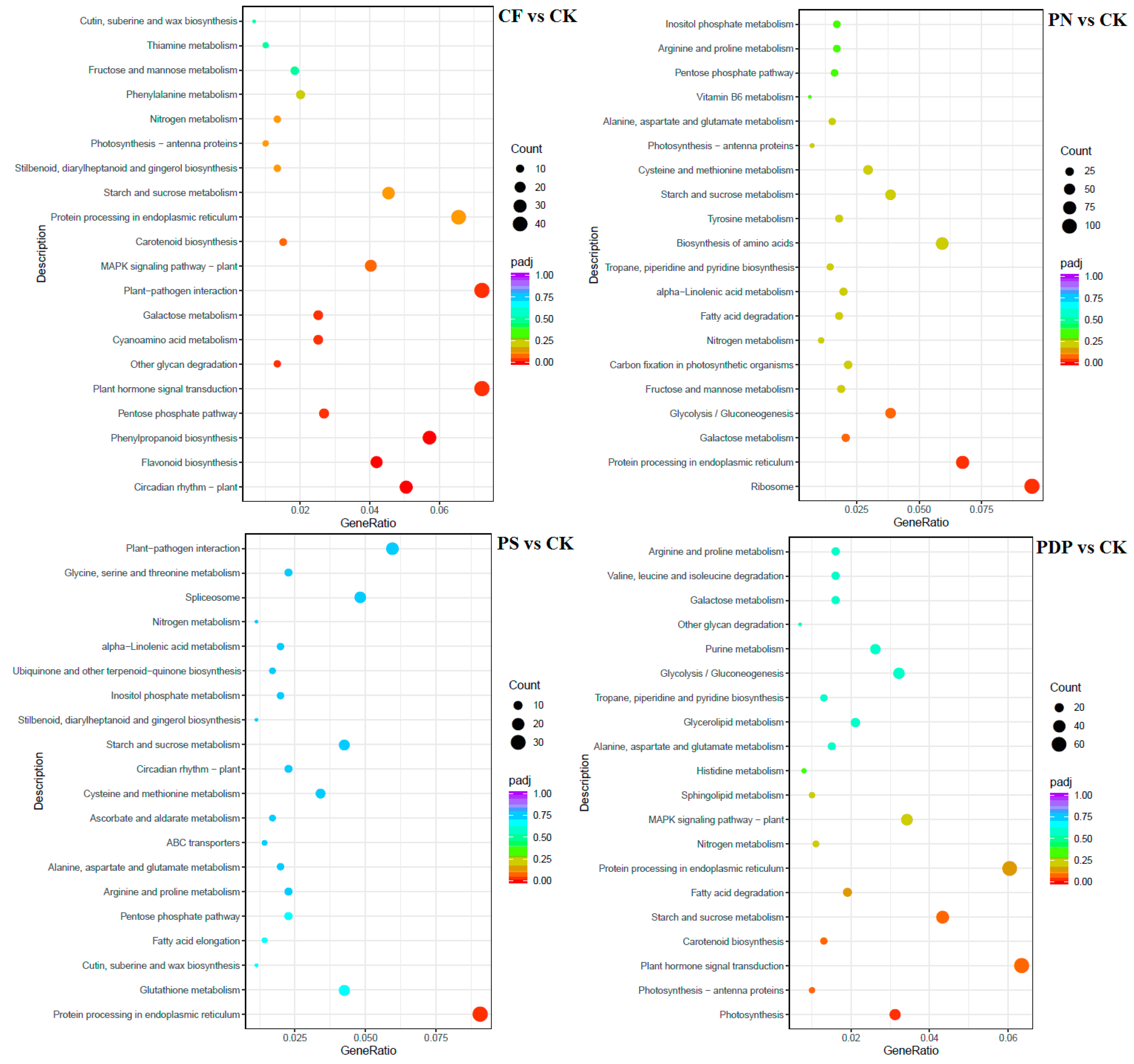
2.7. Functional Classification of DEGs
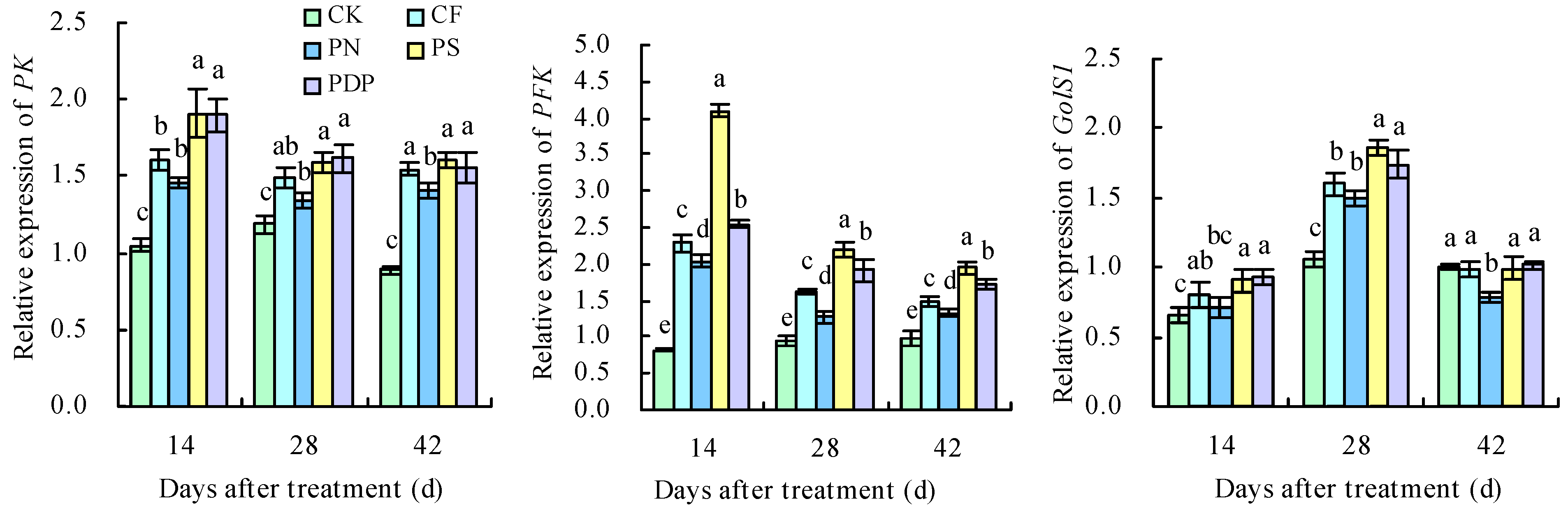
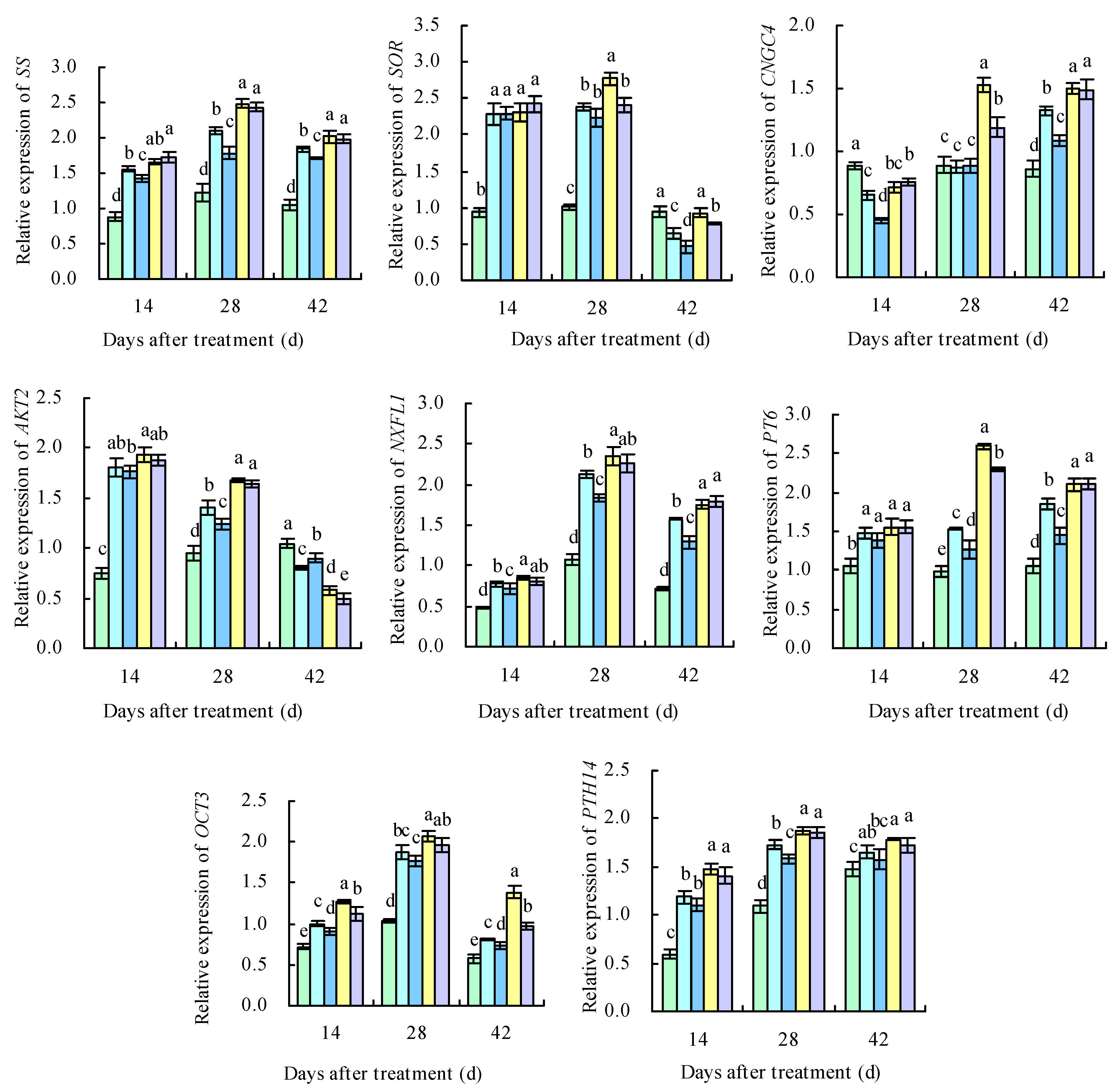
2.8. qRT-PCR Analysis of DEGs
3. Discussion
3.1. Effects of Potassium-Containing Fertilizers on the Sugar Content in Fruits
3.2. Effects of Potassium-Containing Fertilizers on the Activity of Sugar Metabolism-Related Enzymes in Fruits
3.3. Effects of Potassium-Containing Fertilizers on the Organic Acid Content in Fruits
3.4. Effects of Potassium-Containing Fertilizers on the Activity of Organic Acid Metabolism-Related Enzymes in Fruits
3.5. Effects of Potassium-Containing Fertilizers on the Sugar and Organic Acid Metabolism Pathways
4. Materials and Methods
4.1. Experimental Site and Materials
4.2. Experimental Design
4.3. Sampling
4.4. Sugar and Organic Acid Content Measurement
4.5. Determination of the Sugar and Organic Acid Metabolism-Related Enzyme Activity
4.6. Determination of the Total Potassium Content
4.7. Transcriptome Sequencing and Analysis of Differentially Expressed Genes (DEGs)
4.8. Quantitative RT-PCR (qRT-PCR) Analysis of DEGs
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anne-Francoise, A.; Jose-Miguel, M.; Chittaranjan, K. Genetics, Genomics, and Breeding of Grapes, 1st ed.; Science Publisher: Boca Raton, FL, USA, 2011; ISBN 9780429189418. [Google Scholar]
- OIV. Country Statistics of China in 2021. 2021. Available online: https://www.oiv.int (accessed on 27 April 2022).
- Gao, Y.M.; Tong, Y.A.; Ma, W.J. Study on grapery soil nutrients condition and balanced fertilization in Shaanxi Guanzhong area. J. Northwest A F Univ. (Nat. Sci. Ed.) 2006, 34, 41–44. [Google Scholar] [CrossRef]
- Shi, L. The Situations and Strategies of Application Offertilizer in Fruit and Vegetable Planting in Shaanxi Province. Master′s Thesis, Northwest A & F University, Yangling, China, 2005. [Google Scholar]
- Tang, J.L.; Yue, J. Sampling and analysis of soil nutrient status in Chengdu Plain vineyards. Guangxi Agric. Mech. 2019, 41, 80–81. [Google Scholar]
- He, H.H.; Li, W.J.; Li, Z.F.; Shang, H. Nutrient dynamics of gravel soil vineyards. Sino-Overseas Grapevine Wine 2021, 46, 31–35. [Google Scholar] [CrossRef]
- Guo, Y.; Sun, X.Z.; Song, X.L.; Wang, Q.C.; Li, Y.J.; Chen, S.Y. Study on the effects of potassium nutrition on growth and quality formation of cotton fiber. J. Shandong Agric. Univ. (Nat. Sci. Ed.) 2006, 37, 141–144. [Google Scholar]
- Lu, J.L.; Zhang, H.J.; Liu, Z.F. Effects of application of potassium on the yield and quality of grape in greenhouse. Chin. J. Soil Sci. 2015, 46, 694–697. [Google Scholar] [CrossRef]
- Li, M.; Zhang, J.X.; Guo, S.J.; Su, X.D.; Wu, B.; Zheng, Q.Q. Effect of combined application of fertilizer on fruit characters and yield of Crimson Seedless grape in gobi soil of dry region. Sino-Overseas Grapevine Wine 2010, 35, 20–23. [Google Scholar] [CrossRef]
- Wang, X.L.; Zhang, Z.W.; Shao, X.D.; Zhong, X.M.; Wang, F.C.; Shi, X.B.; Wang, Z.Q.; Wang, H.B. Correlation analysis between mineral element and quality index of ’Merlot’ grape fruit. Sino-Overseas Grapevine Wine 2022, 47, 10–20. [Google Scholar] [CrossRef]
- Yan, Y.S.; Lu, J.; Song, K.; Li, B.; Yang, J.H.; Zhang, C. Effects of potassium fertilization on the quality of ’Muscat Hamburg’ grapes. J. Tianjin Agric. Univ. 2022, 29, 14–18. [Google Scholar] [CrossRef]
- Lester, G.E.; Jifon, J.L.; Makus, D.J. Supplemental foliar potassium applications with or without a surfactant can enhance netted muskmelon quality. Hortscience 2006, 41, 741–744. [Google Scholar] [CrossRef]
- Husain, A.; Muhammad, S.; Rizwan, U.; Sikandar, H.; Muhammad, S. Dose optimization of potassium (K) for yield and quality increment of strawberry (Fragaria × ananassa Duch) chandler. Am. J. Exp. Agri. 2014, 4, 1526–1535. [Google Scholar]
- Manuel, N.; Fernando, A.; Vicente, M.; Francisco, R. K+ uptake in plant roots: The systems involved, their regulation and parallels in other organisms. J. Plant Physiol. 2014, 171, 688–695. [Google Scholar] [CrossRef]
- Guo, A.; Lin, X.J.; Gao, H.H.; Zheng, X.L.; Chen, M.; Zheng, G.H. Effects of potassium fertilization on sugar metabolism and related enzymatic activities in Ficus carica. Fujian J. Agric. Sci. 2019, 34, 1388–1396. [Google Scholar] [CrossRef]
- Wu, C.Y. Effect of Potassium on Photosynthesis, Fruit Quality and Sugar Metabolism of Juiube. Ph.D. Thesis, China Agricultural University, Beijing, China, 2016. [Google Scholar]
- Zhou, M. Research on Nutrients Change Rule of Vitis davidii foěx and Influence of Potassium to Fruit Sugar Accumulation. Ph.D. Thesis, Hunan Agricultural University, Changsha, China, 2017. [Google Scholar]
- Huang, H.; Zhao, X.Y.; Xiao, Q.; Hu, W.J.; Wang, P.; Luo, Y.Y.; Xia, H.; Lin, L.J.; Lv, X.L.; Liang, D.; et al. Identification of key genes induced by different potassium levels provides insight into the formation of fruit quality in grapes. Int. J. Mol. Sci. 2023, 24, 1218. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; Andrianteranagna, M.; Cuellar, T.; Cherel, I.; Gibrat, R.; Boeglin, M.; Moreau, B.; Paris, N.; Verdeil, J.L.; Zimmermann, S.D.; et al. Characterization of the grapevine Shaker K+ channel VvK3.1 supports its function in massive potassium fluxes necessary for berry potassium loading and pulvinus-actuated leaf movements. New Phytol. 2019, 222, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Fontes, N.; Gerós, H.; Delrot, S. Grape berry vacuole: A complex and heterogeneous membrane system specialized in the accumulation of solutes. Am. J. Enol. Vitic. 2011, 62, 270–278. [Google Scholar] [CrossRef]
- Zhang, W. Effects of Potassium on Fruit Quality and its Association with Metabolic Pathway of Trehalose 6-phosphate in Apple. Ph.D. Thesis, Northwest A & F University, Yangling, China, 2017. [Google Scholar]
- Guo, Z.G.; Li, W.F.; Mao, J.; Zuo, C.W.; Chen, B.H. Effects of potassium fertilizer on endogenous hormone content and acid metabolism in fruit of apple cv. ‘Red Delicious’. Trans. Chin. Soc. Agric. Eng. 2019, 35, 281–290. [Google Scholar]
- Ramos, M.C.; Romero, M.P. Potassium uptake and redistribution in Cabernet Sauvignon and Syrah grape tissues and its relationships with grape quality parameters. J. Sci. Food Agric. 2017, 97, 3269–3277. [Google Scholar] [CrossRef] [PubMed]
- Sustr, M.; Soukup, A.; Tylova, E. Potassium in root growth and development. Plants 2019, 8, 435. [Google Scholar] [CrossRef] [PubMed]
- Gu, N.; Zhao, L.P.; Zhao, X.M. A review and perspective on slow and controlled release fertilizer in China. Appl. Mech. Mater. 2014, 535, 222–225. [Google Scholar] [CrossRef]
- Pushpavathi, Y.; Satisha, J.; Satisha, G.C.; Reddy, M.L. Influence of potassium fertilization on yield, petiole and soil nutrient status of table grape cv. Sharad seedless. J. Plant Nutr. 2021, 44, 2218–2227. [Google Scholar] [CrossRef]
- Xu, Z.Y.; Yan, J.; Cai, Z.X.; Sun, M.; Su, Z.W.; Shen, Z.J.; Ma, R.J.; Yu, M.L. Correlation between soluble sugar, organic acid and phenolic substances with tasted flavor in peach fruit. Jiangsu J. Agric. Sci. 2022, 38, 190–199. [Google Scholar]
- Zhong, H.X. Sugar Accumulation Mechanism and Key Gene Mining in Grape Fruit. Ph.D. Thesis, Xinjiang Agricultural University, Urumqi, China, 2021. [Google Scholar]
- Liu, M.B. Effects of Potassium Fertilizer on Photosynthetic Characteristics of Grape Leaves and Fruit Quality. Master’s Thesis, Northwest A&F University, Yangling, China, 2022. [Google Scholar]
- Wu, K.J.; Hu, C.X.; Wang, J.; Guo, J.D.; Sun, X.C.; Tan, Q.L.; Zhao, X.H.; Wu, S.W. Comparative effects of different potassium sources on soluble sugars and organic acids in tomato. Sci. Hortic-Amst. 2023, 308, 111601. [Google Scholar] [CrossRef]
- Shi, X.B.; Wang, X.D.; Wang, B.L.; Wang, Z.Q.; Ji, X.H.; Wang, X.L.; Liu, F.Z.; Wang, H.B. Requirement characteristics of mineral elements in different developmental phases of kyoho grapevine. Sci. Agric. Sin. 2019, 52, 2686–2694. [Google Scholar]
- Wind, J.; Smeekens, S.; Hanson, J. Sucrose: Metabolite and signaling molecule. Phytochemistry 2010, 71, 1610–1614. [Google Scholar] [CrossRef] [PubMed]
- Su, J.Y.; Yao, Y.; Liu, Y.H.; Han, Q.Y.; Zhang, W.L. Function, structure and catalytic mechanism of sucrose phosphate synthase: A review. Chin. J. Biotechnol. 2021, 37, 1858–1868. [Google Scholar] [CrossRef]
- Liu, H.J.; Shi, C.Y.; Zhang, L.M.; Zhang, H.F.; Wang, Z.Z.; Cha, S.S. Effect of potassium on related enzyme activities in sugar metabolism of edible sweet potato. J. Plant Nutr. Fertil. 2012, 18, 724–732. [Google Scholar]
- Wu, S.W.; Zhang, C.M.; Li, M.; Tan, Q.L.; Sun, X.C.; Pan, Z.Y.; Deng, X.X.; Hu, C.X. Effects of potassium on fruit soluble sugar and citrate accumulations in ’Cara Cara‘ navel orange (Citrus sinensis L. Osbeck). Sci. Hortic-Amst. 2021, 283, 110057. [Google Scholar] [CrossRef]
- Shen, C.W.; Wang, J.; Jin, X.; Liu, N.; Fan, X.S.; Dong, C.X.; Shen, Q.R.; Xu, Y.C. Potassium enhances the sugar assimilation in leaves and fruit by regulating the expression of key genes involved in sugar metabolism of Asian pears. Plant Growth Regul. 2017, 83, 287–300. [Google Scholar] [CrossRef]
- Wu, C.Y.; Jiang, H.; Li, T.H.; Yu, J. Effect of soil potassium fertilization on leaf photosynthesis and fruit quality of jujube. Soil Fertil. Sci. China 2018, 276, 99–106. [Google Scholar]
- Zhang, W.; Zhang, X.; Wang, Y.F.; Zhang, N.S.; Guo, Y.P.; Ren, X.L.; Zhao, Z.Y. Potassium fertilization arrests malate accumulation and alters soluble sugar metabolism in apple fruit. Biol. Open 2018, 7, bio024745. [Google Scholar] [CrossRef]
- Kodur, S. Effects of juice pH and potassium on juice and wine quality, and regulation of potassium in grapevines through rootstocks (Vitis): A short review. Vitis 2011, 50, 1–6. [Google Scholar]
- Zhang, X. Effects of Different Potassium Level Onsugar and Acid Metabolism in ‘Gala’ Apple Fruit. Master’s Thesis, Northwest A&F University, Yangling, China, 2016. [Google Scholar]
- Zhao, Y.; Zhu, H.J.; Yang, D.D.; Gong, C.S.; Liu, W.G. Research progress of citric acid metabolism in the fruit. Acta Hortic. Sin. 2022, 49, 2579–2596. [Google Scholar] [CrossRef]
- Shen, Z.J.; Ma, R.J.; Yu, M.L.; Cai, Z.X.; Song, H.F.; Li, X. Regularity analysis of main sugar and acid in fruit development of peach. Acta Agric. Boreali-Sin. 2007, 22, 130–134. [Google Scholar]
- Jia, H.F.; Wang, Y.H.; Sun, M.Z.; Li, B.B.; Han, Y.; Zhao, Y.X.; Li, X.L.; Ding, N.; Li, C.; Ji, W.L.; et al. Sucrose functions as a signal involved in the regulation of strawberry fruit development and ripening. New Phytol. 2013, 198, 453–465. [Google Scholar] [CrossRef]
- Armengaud, P.; Sulpice, R.; Miller, A.J.; Stitt, M.; Amtmann, A.; Gibon, Y. Multilevel analysis of primary metabolism provides new insights into the role of potassium nutrition for glycolysis and nitrogen assimilation in Arabidopsis roots. Plant Physiol. 2009, 150, 772–785. [Google Scholar] [CrossRef]
- Guo, Y.; Sun, X.Z.; Song, X.L.; Wang, Q.C.; Li, Y.J.; Chen, S.Y. Effects of potassium nutrition on growth and leaf physiological characteristics at seedling stage of cotton. J. Plant Nutr. Fertil. 2006, 12, 363–368. [Google Scholar]
- Zhnag, C.X.; Yang, T.Y.; Luo, J.; Wang, X.L.; Jiang, A.L.; Ye, Z.W. Effects of different fertilizers and application methods on photosynthetic characteristics andquality of kyoho grape. Southwest China J. Agric. Sci. 2010, 23, 440–443. [Google Scholar] [CrossRef]
- Chen, J.W.; Zhang, S.L.; Zhang, L.C. Sugar transport, metabolism, accumulation and their regulation in fruits. Physiol. Mol. Biol. Plants 2004, 30, 1–10. [Google Scholar]
- Weber, A.; Servaites, J.C.; Geiger, D.R.; Kofler, H.; Hille, D.; Gröner, F.; Hebbeker, U.; Flügge, U.I. Identification, purification, and molecular cloning of a putative plastidic glucose translocator. Plant Cell 2000, 12, 787. [Google Scholar] [CrossRef] [PubMed]
- Martinoia, E.; Massonneau, A.; Frangne, N. Transport processes of solutes across the vacuolar membrane of higher plants. Plant Cell Physiol. 2000, 41, 1175–1185. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Weerasinghe, R.R.; Perdue, T.D.; Cakmakci, N.G.; Taylor, J.P.; Marzluff, W.F.; Jones, A.M. A Golgi-localized hexose transporter is involved in heterotrimeric G protein-mediated early development in Arabidopsis. Mol. Biol. Cell 2006, 17, 4257–4269. [Google Scholar] [CrossRef]
- Poschet, G.; Hannich, B.; Raab, S.; Jungkunz, I.; Klemens, P.A.W.; Krueger, S.; Wic, S.; Neuhaus, H.E.; Büttner, M. A novel Arabidopsis vacuolar glucose exporter is involved in cellular sugar homeostasis and affects the composition of seed storage compounds. Plant Physiol. 2011, 157, 1664–1676. [Google Scholar] [CrossRef] [PubMed]
- Obenland, D.; Feliziani, E.; Zhu, S.; Zhao, X.; Margosan, D.A.; Gabler, F.M.; Van Zyl, S.; Romanazzi, G.; Smilanick, J.L.; Beno-Moualem, D.; et al. Potassium application to table grape clusters after veraison increases soluble solids by enhancing berry water loss. Sci. Hortic-Amst. 2015, 187, 58–64. [Google Scholar] [CrossRef]
- Zhang, C.; Jia, H.F.; Wang, J.; Jiu, S.T.; Wang, M.Q. Effectiveness and concentration of foliar-application of potassium fertilizers ongrapevine evaluated by expression of potassium uptake related genes. J. Plant Nutr. Fertil. 2016, 22, 1091–1101. [Google Scholar] [CrossRef]
- Xue, B.G. Effects of Potassium on Fruit Ouality and EjUGDI and EjTP3 Gene Cloning and Expression of Loquat. Master’s Thesis, South Western University of Finance and Economics, Chongqing, China, 2020. [Google Scholar]
- Hua, B.; Mercier, R.W.; Leng, Q.; Berkowitz, G.A. Plants do it differently: A new basis for potassium/sodium selectivity in the pore of an ion channel. Plant Physiol. 2003, 132, 1353–1361. [Google Scholar] [CrossRef] [PubMed]
- Harada, H.; Leigh, R.A. Genetic mapping of natural variation in potassium concentrations in shoots of Arabidopsis thaliana. J. Exp. Bot. 2006, 57, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Lacombe, B.; Pilot, G.; Michard, E.; Gaymard, F.; Sentenac, H.; Thibaud, J.B. A shaker-like K+ channel with weak rectification is expressed in both source and sink phloem tissues of Arabidopsis. Plant Cell 2000, 12, 837–851. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agrochemical Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000; ISBN 9787109066441. [Google Scholar]
- Hu, R.P. The Effects of Shed and Potassium Fertilizer on the Germination and High-Quality Berry Formation of Grape under Rain-Sheltered Cultivation. Ph.D. Thesis, Sichuan Agricultural University, Chengdu, China, 2017. [Google Scholar]
- Bai, B.Z. Plant Physiology and Biochemistry; China Agricultural Science and Technology Press: Beijing, China, 2003; ISBN 7801197909. [Google Scholar]
- Chen, M.; Jiang, Q.; Yin, X.R.; Lin, Q.; Chen, J.Y.; Allan, A.C.; Xu, C.J.; Chen, K.S. Effect of hot air treatment on organic acid-and sugar-metabolism in Ponkan (Citrus reticulata) fruit. Sci. Hortic-Amst. 2012, 147, 118–125. [Google Scholar] [CrossRef]
- Hu, Z.Q.; Wang, H.C.; Hu, G.B. Measurement of sugars, organic acids and vitamin C in litchi fruit by highperformance liquid chromatography. J. Fruit Sci. 2005, 22, 582–585. [Google Scholar]
- Cecilia, D.S.; Gianpiero, Z.; Alberto, F.; Andrea, M.; Dal, M.A.; Luca, V.; Genny, B.; Paola, T.; Carla, A.; Elisa, Z.; et al. The high polyphenol content of grapevine cultivar Tannat berries is conferred primarily by genes that are not shared with the reference genome. Plant Cell 2013, 25, 4777–4788. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Clean Reads | Clean Bases | GC Content | Q20 Percentage | Q30 Percentage |
|---|---|---|---|---|---|---|
| CCK | 47,431,499 | 46,848,865 | 7.03 G | 46.59% | 97.28% | 92.61% |
| CCF | 46,678,397 | 45,942,010 | 6.89 G | 46.55% | 97.44% | 93.06% |
| CPN | 47,272,605 | 46,232,352 | 6.93 G | 46.53% | 97.43% | 93.02% |
| CPS | 45,467,270 | 44,788,052 | 6.72 G | 46.59% | 97.56% | 93.26% |
| CPDP | 50,756,362 | 50,014,186 | 7.5 G | 46.57% | 97.57% | 93.34% |
| ECK | 47,031,424 | 46,243,460 | 6.94 G | 46.42% | 97.43% | 93.06% |
| ECF | 45,249,647 | 44,366,584 | 6.66 G | 46.43% | 97.71% | 93.66% |
| EPN | 48,886,265 | 48,140,646 | 7.22 G | 46.74% | 97.72% | 93.62% |
| EPS | 47,141,936 | 46,461,278 | 6.97 G | 46.52% | 97.84% | 93.87% |
| EPDP | 48,923,110 | 48,030,740 | 7.20 G | 46.38% | 97.52% | 93.13% |
| GCK | 44,575,317 | 43,594,313 | 6.54 G | 46.43% | 96.92% | 91.86% |
| GCF | 45,679,846 | 44,867,686 | 6.73 G | 46.43% | 97.38% | 92.74% |
| GPN | 47,601,313 | 46,808,660 | 7.02 G | 46.66% | 97.86% | 93.90% |
| GPS | 46,993,310 | 46,268,872 | 6.94 G | 46.45% | 97.74% | 93.74% |
| GPDP | 45,907,787 | 44,653,306 | 6.70 G | 47.14% | 97.70% | 93.73% |
| KEGG ID | Metabolic Pathways | 14 Days after Treatment | 28 Days after Treatment | 42 Days after Treatment | Total Number of DEGs for Pathway Annotations | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CF vs. CK | PN vs. CK | PS vs. CK | PDP vs. CK | CF vs. CK | PN vs. CK | PS vs. CK | PDP vs. CK | CF vs. CK | PN vs. CK | PS vs. CK | PDP vs. CK | |||
| vvi03010 | Ribosome | 9 | 71 | 109 | 0 | 0 | 106 | 0 | 0 | 0 | 177 | 167 | 148 | 787 |
| vvi01200 | Carbon metabolism | 72 | 0 | 98 | 110 | 0 | 0 | 0 | 0 | 86 | 0 | 133 | 125 | 624 |
| vvi00860 | Porphyrin and chlorophyll metabolism | 17 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 24 | 0 | 36 | 32 | 109 |
| vvi04075 | Plant hormone signal transduction | 0 | 57 | 0 | 0 | 0 | 0 | 0 | 0 | 71 | 0 | 0 | 95 | 223 |
| vvi00051 | Fructose and mannose metabolism | 0 | 0 | 27 | 26 | 0 | 0 | 0 | 0 | 0 | 31 | 32 | 29 | 145 |
| vvi00010 | Glycolysis/Gluconeogenesis | 40 | 0 | 52 | 64 | 0 | 0 | 0 | 0 | 48 | 0 | 70 | 80 | 354 |
| vvi00710 | Carbon fixation in photosynthetic organisms | 33 | 21 | 35 | 38 | 0 | 0 | 0 | 0 | 36 | 40 | 39 | 42 | 284 |
| vvi01230 | Biosynthesis of amino acids | 0 | 0 | 0 | 87 | 0 | 0 | 0 | 0 | 63 | 0 | 107 | 94 | 351 |
| vvi00906 | Carotenoid biosynthesis | 15 | 0 | 0 | 18 | 0 | 0 | 0 | 0 | 0 | 20 | 22 | 18 | 93 |
| No. | Gene ID in NCBI | Gene Description | p Value |
|---|---|---|---|
| 1 | 100255934 | Pyruvate kinase (PK) | 0.012 |
| 2 | 100249662 | Phosphofructokinase (PFK) | 1.2 × 10−3 |
| 3 | 100260266 | Galactinol synthase-1 (GolS1) | 3.0 × 10−11 |
| 4 | 100249279 | Sucrose synthase (SS) | 9.3 × 10−15 |
| 5 | 100233111 | Shaker-like potassium channel (SOR) | 1.0 × 10−5 |
| 6 | 100265816 | Cyclic nucleotide-gated ion channel 4 (CNGC4) | 1.9 × 10−3 |
| 7 | 100261806 | Potassium channel AKT2/3 (AKT2) | 0.013 |
| 8 | 100268010 | NF-X1-type zinc finger protein NFXL1 (NFXL1) | 6.7 × 10−6 |
| 9 | 100251966 | Potassium transporter 6 (PT6) | 0.013 |
| 10 | 100263786 | Organic cation/carnitine transporter 3 (OCT3) | 3.2 × 10−4 |
| 11 | 100257925 | Low-affinity inorganic phosphate transporter 1 (PHT14) | 1.4 × 10−12 |
| Gene Name | Gene ID in NCBI | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) |
|---|---|---|---|
| PK | 100255934 | AGGATCAGGTCAACTCCTCT | ATCACAACGATGAGCTTTGC |
| PFK | 100249662 | TAAGCCGATCCTTTCCTCAC | CTAATGGGTGGATGCTCAGT |
| GolS1 | 100260266 | GCAGATGTGTTTACAGCCG | TTCACATAGTCCCCGTTTCC |
| SS | 100249279 | TTCAGCCCCGTTGAGAATAA | CAACAAGTCCCGTCAAGTTC |
| SOR | 100233111 | GCAACCTAACAGCTCCTTTG | CGCAGAAGCCTACACAATTC |
| CNGC4 | 100265816 | AGAGTCAAGAGAAGTGCGAG | TCTTGGCCGGATAAATGACA |
| AKT2 | 100261806 | GAATGGCGCAGATGTTATCC | AATTCGGTGTCCCATCTCTC |
| NFXL1 | 100268010 | TCACGTTTTCTGTCCAATGC | AATAAAACGCTTTGGCTCCC |
| PT6 | 100251966 | CCCGAGAGTCCAAAGATTGA | TTCTTCACCATGCTAGACCC |
| OCT3 | 100263786 | GAGACCGGCTATAAGAGAGC | GAGGAGAGGGGTTGAATCTG |
| PHT14 | 100257925 | GCGTTAATCGACAAGATGGG | AGAGTACATCACCACGAACC |
| ACTIN | 100232866 | CTTGCATCCCTCAGCACCTT | TCCTGTGGACAATGGATGGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Lu, Y.; Zhang, X.; Hu, W.; Lin, L.; Deng, Q.; Xia, H.; Liang, D.; Lv, X. Effects of Potassium-Containing Fertilizers on Sugar and Organic Acid Metabolism in Grape Fruits. Int. J. Mol. Sci. 2024, 25, 2828. https://doi.org/10.3390/ijms25052828
Wang J, Lu Y, Zhang X, Hu W, Lin L, Deng Q, Xia H, Liang D, Lv X. Effects of Potassium-Containing Fertilizers on Sugar and Organic Acid Metabolism in Grape Fruits. International Journal of Molecular Sciences. 2024; 25(5):2828. https://doi.org/10.3390/ijms25052828
Chicago/Turabian StyleWang, Jin, Yuhang Lu, Xuemei Zhang, Wenjie Hu, Lijin Lin, Qunxian Deng, Hui Xia, Dong Liang, and Xiulan Lv. 2024. "Effects of Potassium-Containing Fertilizers on Sugar and Organic Acid Metabolism in Grape Fruits" International Journal of Molecular Sciences 25, no. 5: 2828. https://doi.org/10.3390/ijms25052828
APA StyleWang, J., Lu, Y., Zhang, X., Hu, W., Lin, L., Deng, Q., Xia, H., Liang, D., & Lv, X. (2024). Effects of Potassium-Containing Fertilizers on Sugar and Organic Acid Metabolism in Grape Fruits. International Journal of Molecular Sciences, 25(5), 2828. https://doi.org/10.3390/ijms25052828