
Fibroblasts and Endothelial Cells in Three-Dimensional Models: A New Tool for Addressing the Pathogenesis of Systemic Sclerosis as a Prototype of Fibrotic Vasculopathies
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Fine-Tuning of Optimal Cell Culture Conditions
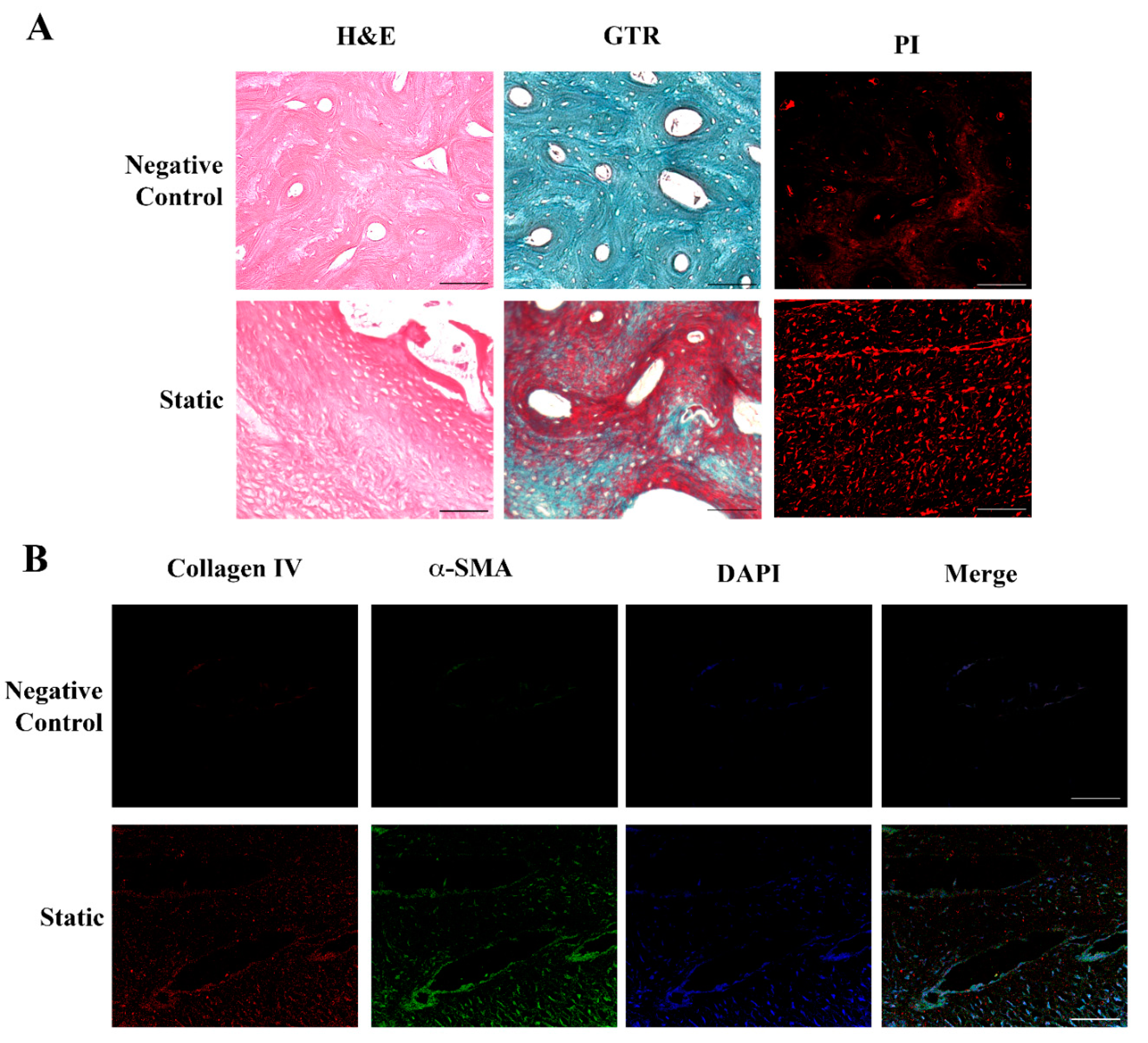
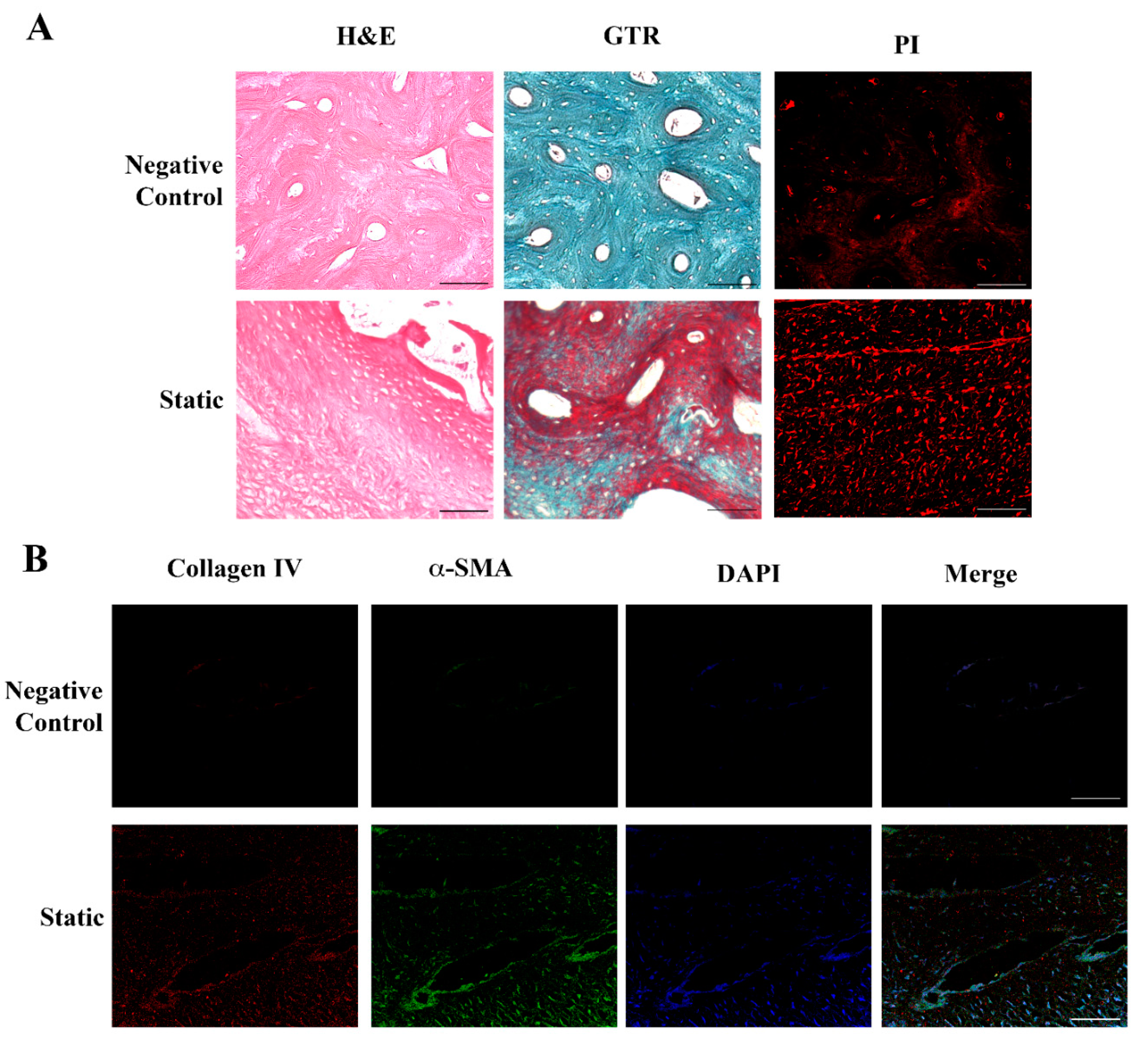
2.2. Human Dermal Fibroblasts Can Colonize and Reshape Bone Scaffold
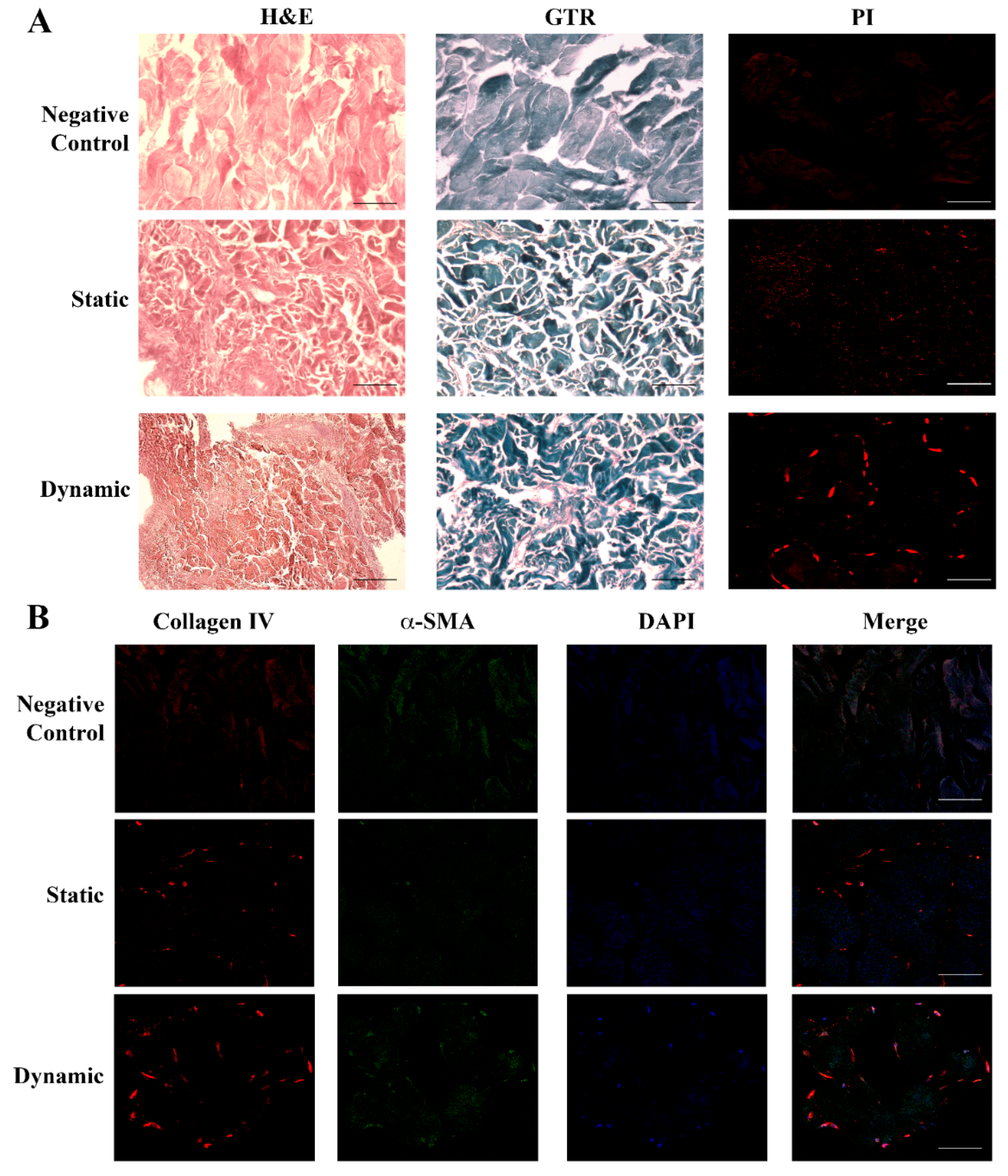
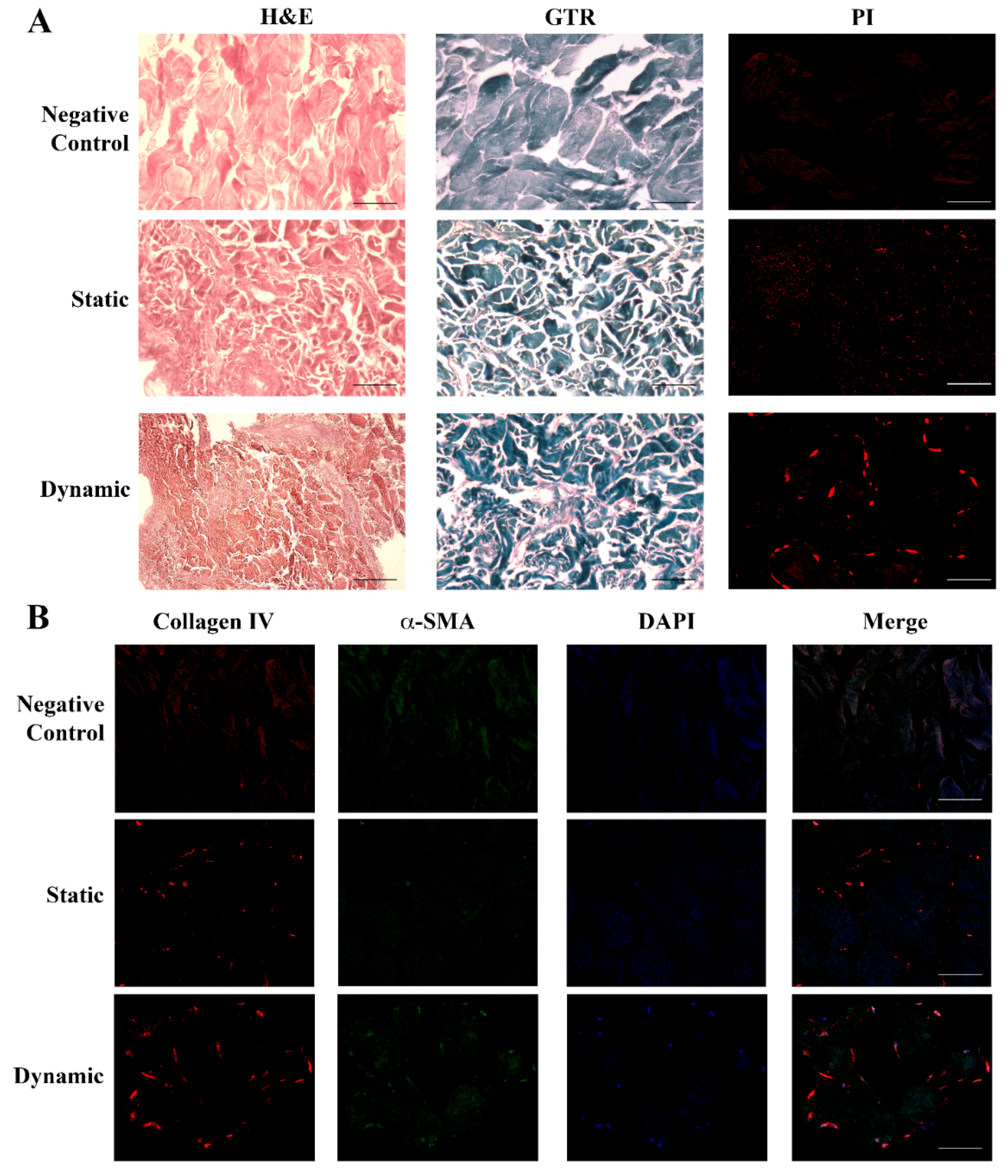
2.3. Human Dermal Fibroblasts Can Colonize Skin Scaffold and Develop a New Texture
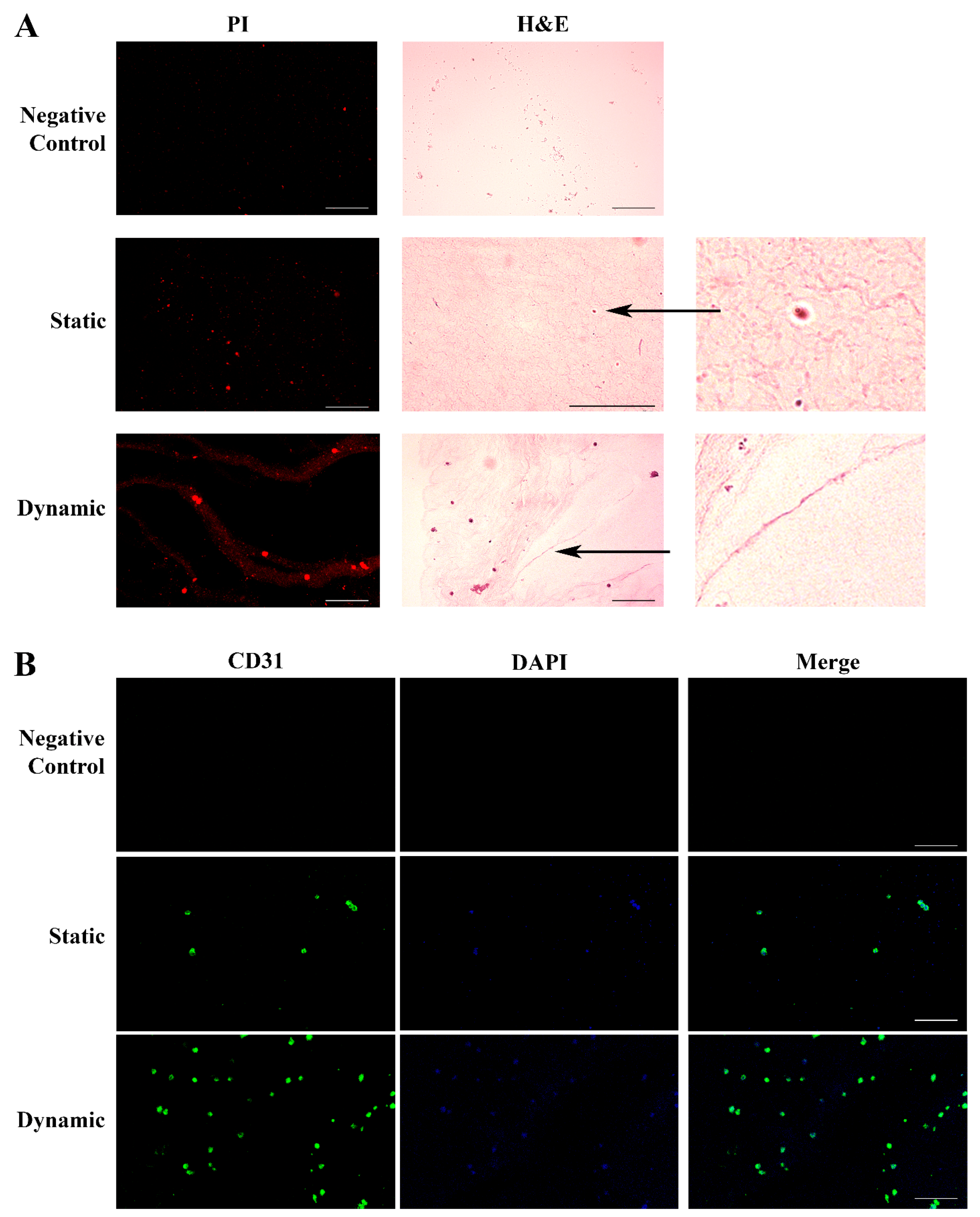
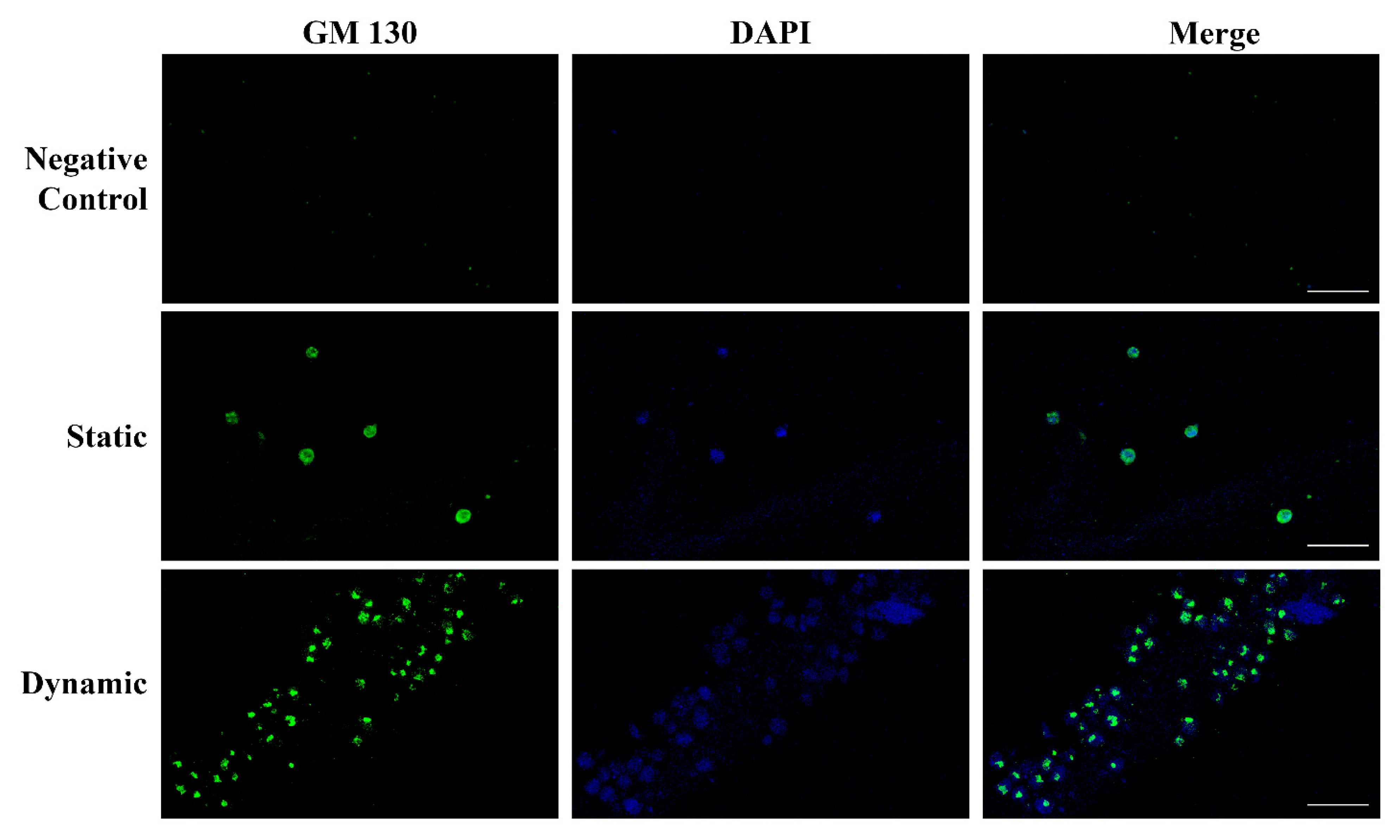
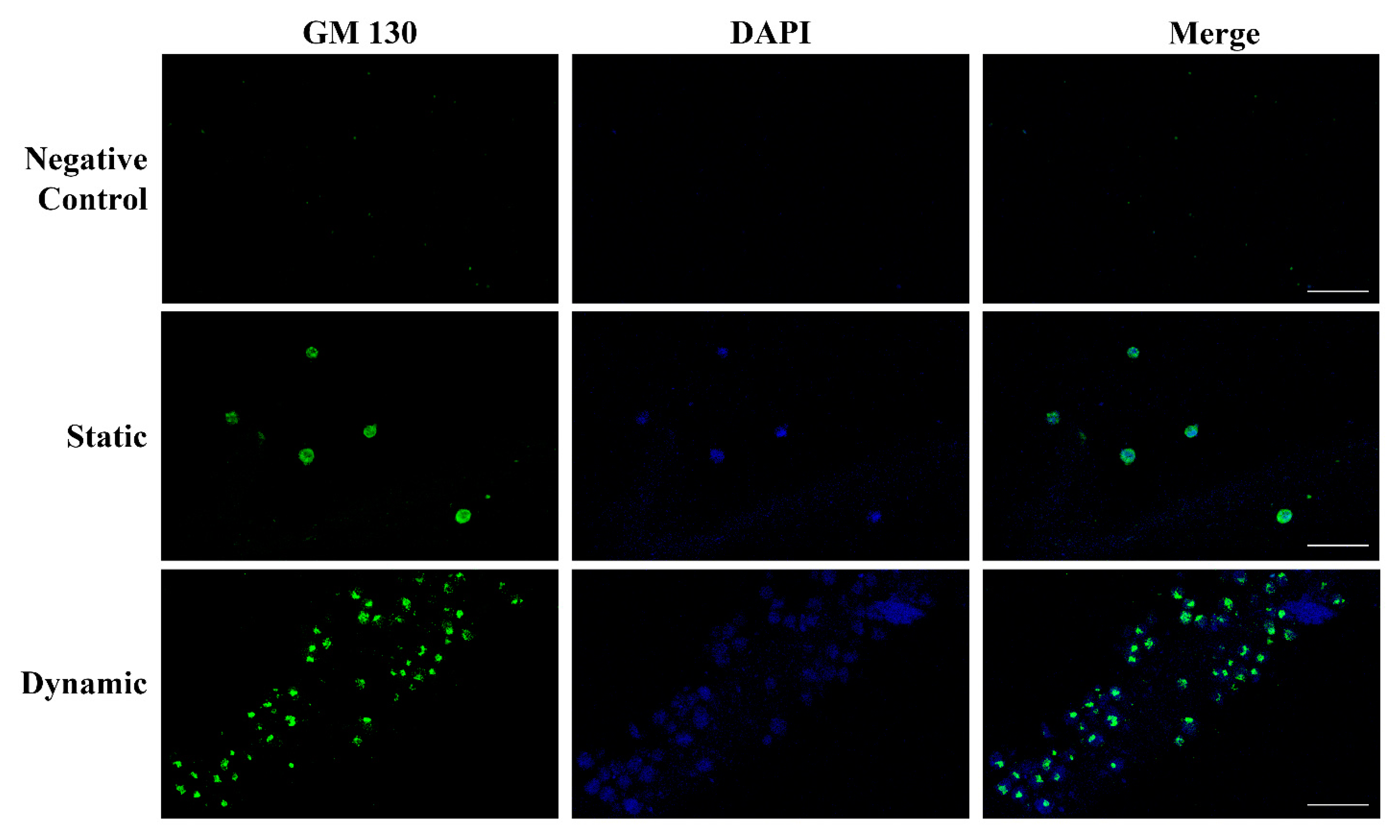
2.4. Endothelial Cells Can Develop Primitive Vascular Plexus in Lung ECM Hydrogel
3. Discussion
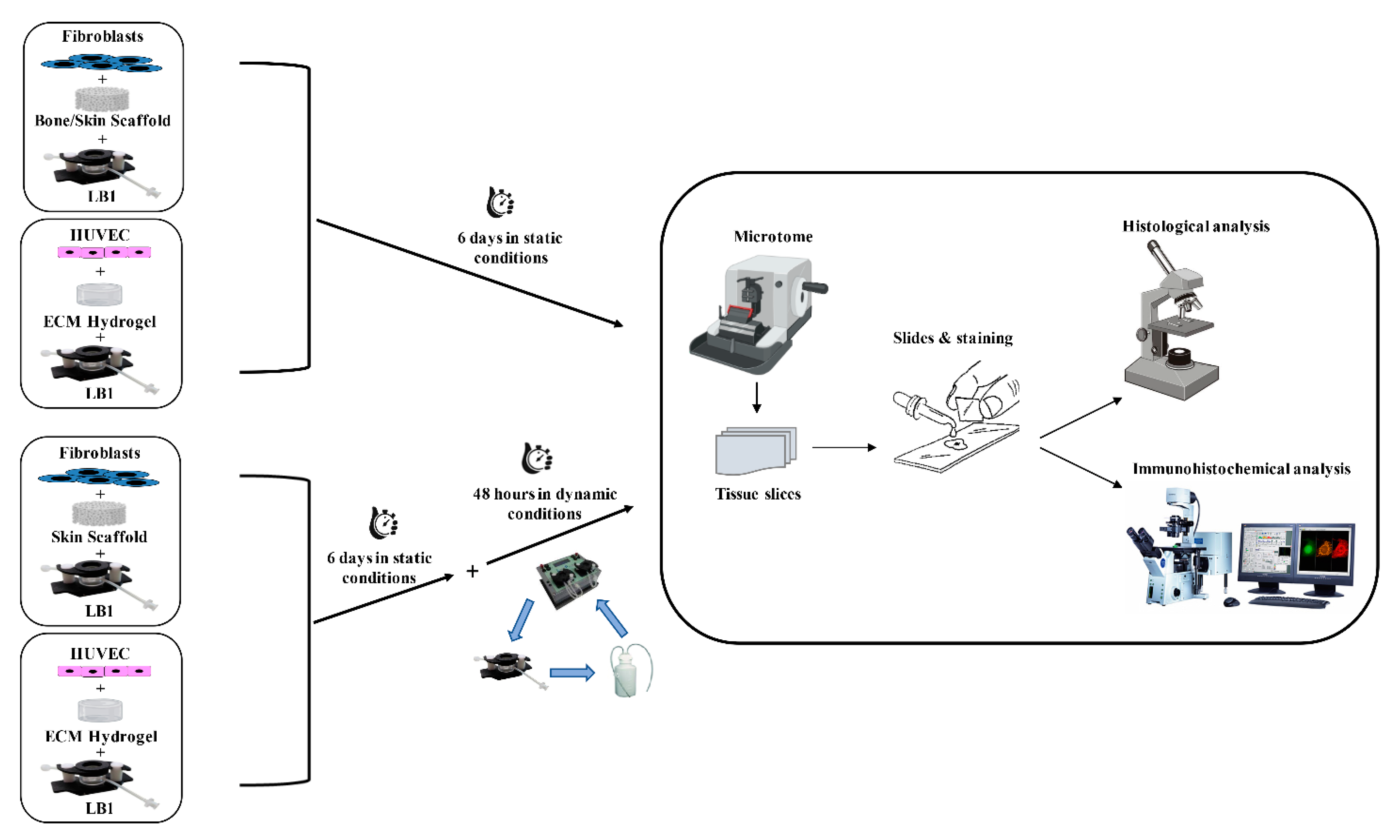
4. Materials and Methods
4.1. Modular Multi-Compartmental Cell Culture System
4.2. Primary Cells
4.2.1. Human Healthy Skin Fibroblasts
4.2.2. Human Macrovascular Endothelial Cells
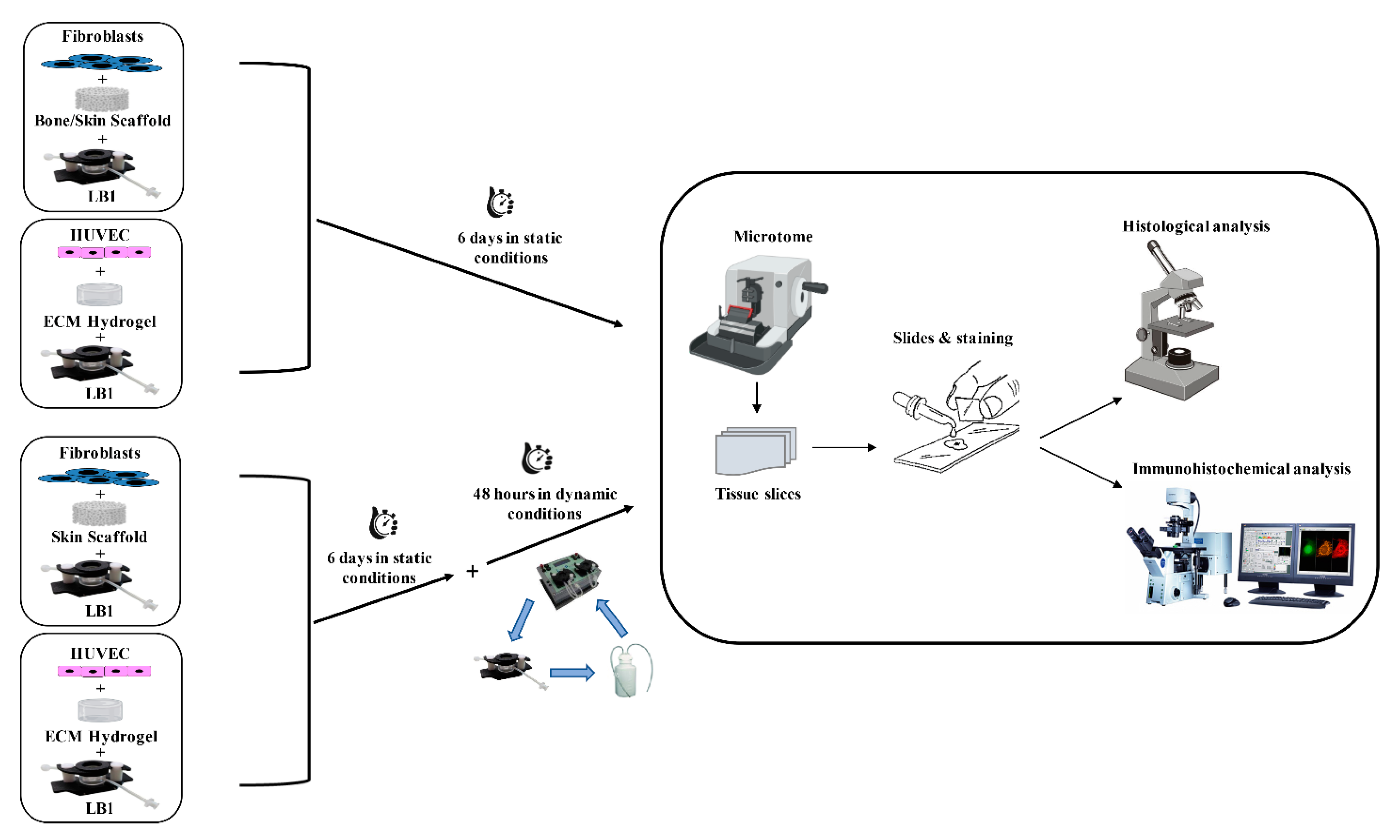
4.2.3. 3D Cell Cultures
4.3. Histological Analyses
4.4. Immunocytochemical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jensen, C.; Teng, Y. Is It Time to Start Transitioning From 2D to 3D Cell Culture? Front. Mol. Biosci. 2020, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Duval, K.; Grover, H.; Han, L.-H.; Mou, Y.; Pegoraro, A.F.; Fredberg, J.; Chen, Z. Modeling Physiological Events in 2D vs. 3D Cell Culture. Physiology 2017, 32, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Frangogiannis, N.G. Why animal model studies are lost in translation. J. Cardiovasc. Aging 2022, 2, 22. [Google Scholar] [CrossRef] [PubMed]
- Saeidnia, S.; Manayi, A.; Abdollahi, M. From in vitro experiments to in vivo and clinical studies; pros and cons. Curr. Drug Discov. Technol. 2015, 12, 218–244. [Google Scholar] [CrossRef] [PubMed]
- Díaz, L.; Zambrano, E.; Flores, M.E.; Contreras, M.; Crispín, J.C.; Alemán, G.; Bravo, C.; Armenta, A.; Valdés, V.J.; Tovar, A.; et al. Ethical Considerations in Animal Research: The Principle of 3R’s. Rev. Investig. Clin. 2020, 73, 199–209. [Google Scholar] [CrossRef]
- Ghasemi, M.; Dehpour, A.R. Ethical Considerations in Animal Studies. J. Med. Ethics Hist. Med. 2009, 2, 12. [Google Scholar]
- Hartung, T. Food for Thought Look Back in Anger-What Clinical Studies Tell Us About Preclinical Work. ALTEX 2013, 30, 275–291. [Google Scholar] [CrossRef]
- Aske, K.C.; Waugh, C.A. Expanding the 3R Principles. EMBO Rep. 2017, 18, 1490–1492. [Google Scholar] [CrossRef]
- Habanjar, O.; Diab-Assaf, M.; Caldefie-Chezet, F.; Delort, L. 3D Cell Culture Systems: Tumor Application, Advantages, and Disadvantages. Int. J. Mol. Sci. 2021, 22, 12200. [Google Scholar] [CrossRef]
- Fang, Y.; Eglen, R.M. Three-Dimensional Cell Cultures in Drug Discovery and Development. SLAS Discov. 2017, 22, 456–472. [Google Scholar] [CrossRef]
- Mousavi, N. Characterization of in vitro 3D cultures. APMIS 2021, 129, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Haycock, J.W. 3D cell culture: A review of current approaches and techniques. Methods Mol. Biol. 2011, 695, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.C.; Moreira, A.F.; de Melo-Diogo, D.; Gaspar, V.M.; Carvalho, M.P.; Correia, I.J. 3D Tumor Spheroids: An Overview on the Tools and Techniques Used for Their Analysis. Biotechnol. Adv. 2016, 34, 1427–1441. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.P.; Gaspar, V.M.; Mano, J.F. Design of Spherically Structured 3D in Vitro Tumor Models—Advances and Prospects. Acta Biomater. 2018, 75, 11–34. [Google Scholar] [CrossRef] [PubMed]
- Giusti, S.; Sbrana, T.; La Marca, M.; Di Patria, V.; Martinucci, V.; Tirella, A.; Domenici, C.; Ahluwalia, A. A Novel Dual-Flow Bioreactor Simulates Increased Fluorescein Permeability in Epithelial Tissue Barriers. Biotechnol. J. 2014, 9, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Paolillo, M.; Colombo, R.; Serra, M.; Belvisi, L.; Papetti, A.; Ciusani, E.; Comincini, S.; Schinelli, S. Stem-Like Cancer Cells in a Dynamic 3D Culture System: A Model to Study Metastatic Cell Adhesion and Anti-Cancer Drugs. Cells 2019, 8, 1434. [Google Scholar] [CrossRef] [PubMed]
- Vozzi, F.; Mazzei, D.; Vinci, B.; Vozzi, G.; Sbrana, T.; Ricotti, L.; Forgione, N.; Ahluwalia, A. A Flexible Bioreactor System for Constructing in Vitro Tissue and Organ Models. Biotechnol. Bioeng. 2011, 108, 2129–2140. [Google Scholar] [CrossRef]
- Varga, J.; Abraham, D. Systemic sclerosis: A prototypic multisystem fibrotic disorder. J. Clin. Investig. 2007, 117, 557–567. [Google Scholar] [CrossRef]
- Rosendahl, A.H.; Schönborn, K.; Krieg, T. Pathophysiology of systemic sclerosis (scleroderma). Kaohsiung J. Med. Sci. 2022, 38, 187–195. [Google Scholar] [CrossRef]
- Patnaik, E.; Lyons, M.; Tran, K.; Pattanaik, D. Endothelial Dysfunction in Systemic Sclerosis. Int. J. Mol. Sci. 2023, 24, 14385. [Google Scholar] [CrossRef]
- Raschi, E.; Chighizola, C.B.; Cesana, L.; Privitera, D.; Ingegnoli, F.; Mastaglio, C.; Meroni, P.L.; Borghi, M.O. Immune Complexes Containing Scleroderma-Specific Autoantibodies Induce a Profibrotic and Proinflammatory Phenotype in Skin Fibroblasts. Arthritis Res. Ther. 2018, 20, 187. [Google Scholar] [CrossRef]
- Raschi, E.; Privitera, D.; Bodio, C.; Lonati, P.A.; Borghi, M.O.; Ingegnoli, F.; Meroni, P.L.; Chighizola, C.B. Scleroderma-Specific Autoantibodies Embedded in Immune Complexes Mediate Endothelial Damage: An Early Event in the Pathogenesis of Systemic Sclerosis. Arthritis Res. Ther. 2020, 22, 265. [Google Scholar] [CrossRef]
- Kutsuna, Y.J.; Iwamoto, N.; Ichinose, K.; Aibara, N.; Nakashima, K.; Nakamura, H.; Koike, Y.; Murota, H.; Ueki, Y.; Miyamoto, H.; et al. Immune Complexome Analysis of a Rich Variety of Serum Immune Complexes Identifies Disease-Characteristic Immune Complex Antigens in Systemic Sclerosis. J. Autoimmun. 2023, 134, 102954. [Google Scholar] [CrossRef]
- Sieminski, A.L.; Semino, C.E.; Gong, H.; Kamm, R.D. Primary Sequence of Ionic Self-assembling Peptide Gels Affects Endothelial Cell Adhesion and Capillary Morphogenesis. J. Biomed. Mater. Res. A 2008, 87A, 494–504. [Google Scholar] [CrossRef]
- Caliari, S.R.; Burdick, J.A. A Practical Guide to Hydrogels for Cell Culture. Nat. Methods 2016, 13, 405–414. [Google Scholar] [CrossRef]
- Ravichandran, Y.; Goud, B.; Manneville, J.-B. The Golgi Apparatus and Cell Polarity: Roles of the Cytoskeleton, the Golgi Matrix, and Golgi Membranes. Curr. Opin. Cell Biol. 2020, 62, 104–113. [Google Scholar] [CrossRef]
- Truchetet, M.E.; Brembilla, N.C.; Chizzolini, C. Current Concepts on the Pathogenesis of Systemic Sclerosis. Clin. Rev. Allergy Immunol. 2023, 64, 262–283. [Google Scholar] [CrossRef]
- Volkmann, E.R.; Andréasson, K.; Smith, V. Systemic sclerosis. Lancet 2023, 401, 304–318. [Google Scholar] [CrossRef] [PubMed]
- Cavazzana, I.; Vojinovic, T.; Airo’, P.; Fredi, M.; Ceribelli, A.; Pedretti, E.; Lazzaroni, M.G.; Garrafa, E.; Franceschini, F. Systemic Sclerosis-Specific Antibodies: Novel and Classical Biomarkers. Clin. Rev. Allergy Immunol. 2023, 64, 412–430. [Google Scholar] [CrossRef] [PubMed]
- van den Hoogen, F.; Khanna, D.; Fransen, J.; Johnson, S.R.; Baron, M.; Tyndall, A.; Matucci-Cerinic, M.; Naden, R.P.; Medsger, T.A., Jr.; Carreira, P.E.; et al. 2013 classification criteria for systemic sclerosis: An American college of rheumatology/European league against rheumatism collaborative initiative. Ann. Rheum. Dis. 2013, 72, 1747–1755. [Google Scholar] [CrossRef] [PubMed]
- Valentini, G.; Marcoccia, A.; Cuomo, G.; Vettori, S.; Ludici, M.; Bondanini, F.; Santoriello, C.; Ciani, A.; Cozzolino, D.; De Matteis, G.M.; et al. Early Systemic Sclerosis: Analysis of the Disease Course in Patients with Marker Autoantibody and/or Capillaroscopic Positivity. Arthritis Care Res. 2014, 66, 1520–1527. [Google Scholar] [CrossRef] [PubMed]
- Marangoni, R.G.; Varga, J.; Tourtellotte, W.G. Animal Models of Scleroderma: Recent Progress. Curr. Opin. Rheumatol. 2016, 28, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Yue, X.; Yu, X.; Petersen, F.; Riemekasten, G. Recent Advances in Mouse Models for Systemic Sclerosis. Autoimmun. Rev. 2018, 17, 1225–1234. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; Yue, X.; Wax, J.; Kasper, B.; Yin, J.; Wang, X.; Zhang, L.; Ahmadi, M.; Heidecke, H.; Müller, A.; et al. Both T and B Cells Are Indispensable for the Development of a PBMC Transfer-Induced Humanized Mouse Model for SSc. Arthritis Res. Ther. 2022, 24, 209. [Google Scholar] [CrossRef]
- Vazquez-Armendariz, A.I.; Barroso, M.M.; El Agha, E.; Herold, S. 3D In Vitro Models: Novel Insights into Idiopathic Pulmonary Fibrosis Pathophysiology and Drug Screening. Cells 2022, 11, 1526. [Google Scholar] [CrossRef]
- Paradiso, F.; Quintela, M.; Lenna, S.; Serpelloni, S.; James, D.; Caserta, S.; Conlan, S.; Francis, L.; Taraballi, F. Studying Activated Fibroblast Phenotypes and Fibrosis-Linked Mechanosensing Using 3D Biomimetic Models. Macromol. Biosci. 2022, 22, e2100450. [Google Scholar] [CrossRef]
- Sfriso, R.; Zhang, S.; Bichsel, C.A.; Steck, O.; Despont, A.; Guenat, O.T.; Rieben, R. 3D Artificial Round Section Micro-Vessels to Investigate Endothelial Cells under Physiological Flow Conditions. Sci. Rep. 2018, 8, 5898. [Google Scholar] [CrossRef]
- Sfriso, R.; Rieben, R. 3D Cell-Culture Models for the Assessment of Anticoagulant and Anti-Inflammatory Properties of Endothelial Cells. Methods Mol. Biol. 2020, 2110, 83–97. [Google Scholar] [CrossRef]
- Kalender, M.; Bulbul, M.; Kolbasi, B.; Keskin, I. In 2D and 3D Cell Culture Models, Effects of Endothelial Cells on E-Cadherin/β-Catenin Expression Levels and Spheroid Sizes in Ishikawa Cells. Asian Pac. J. Cancer Prev. 2022, 23, 39–51. [Google Scholar] [CrossRef]
- Herbert, S.P.; Stainier, D.Y.R. Molecular Control of Endothelial Cell Behaviour during Blood Vessel Morphogenesis. Nat. Rev. Mol. Cell Biol. 2011, 12, 551–564. [Google Scholar] [CrossRef]
- Kučera, T.; Lammert, E. Ancestral Vascular Tube Formation and its Adoption by Tumors. Biol. Chem. 2009, 390, 985–994. [Google Scholar] [CrossRef]
- Schmidt, A.; Brixius, K.; Bloch, W. Endothelial Precursor Cell Migration during Vasculogenesis. Circ. Res. 2007, 101, 125–136. [Google Scholar] [CrossRef]
- DeLisser, H.M.; Christofidou-Solomidou, M.; Strieter, R.M.; Burdick, M.D.; Robinson, C.S.; Wexler, R.S.; Kerr, J.S.; Garlanda, C.; Merwin, J.R.; Madri, J.A.; et al. Involvement of Endothelial PECAM-1/CD31 in Angiogenesis. Am. J. Pathol. 1997, 151, 671–677. [Google Scholar]
- Cao, G.; O’Brien, C.D.; Zhou, Z.; Sanders, S.M.; Greenbaum, J.N.; Makrigiannakis, A.; DeLisser, H.M. Involvement of Human PECAM-1 in Angiogenesis and in Vitro Endothelial Cell Migration. Am. J. Physiol.-Cell Physiol. 2002, 282, C1181–C1190. [Google Scholar] [CrossRef] [PubMed]
- Lamalice, L.; Le Boeuf, F.; Huot, J. Endothelial Cell Migration during Angiogenesis. Circ. Res. 2007, 100, 782–794. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Huang, N.F.; Hsu, S. Mechanotransduction in Endothelial Cell Migration. J. Cell Biochem. 2005, 96, 1110–1126. [Google Scholar] [CrossRef] [PubMed]
- Sonmez, U.M.; Cheng, Y.-W.; Watkins, S.C.; Roman, B.L.; Davidson, L.A. Endothelial Cell Polarization and Orientation to Flow in a Novel Microfluidic Multimodal Shear Stress Generator. Lab. Chip 2020, 20, 4373–4390. [Google Scholar] [CrossRef] [PubMed]
- Hikita, T.; Mirzapourshafiyi, F.; Barbacena, P.; Riddell, M.; Pasha, A.; Li, M.; Kawamura, T.; Brandes, R.P.; Hirose, T.; Ohno, S.; et al. PAR-3 Controls Endothelial Planar Polarity and Vascular Inflammation under Laminar Flow. EMBO Rep. 2018, 19, e45253. [Google Scholar] [CrossRef] [PubMed]
- De Pieri, A.; Rochev, Y.; Zeugolis, D.I. Scaffold-Free Cell-Based Tissue Engineering Therapies: Advances, Shortfalls and Forecast. NPJ Regen. Med. 2021, 6, 18. [Google Scholar] [CrossRef]
- De Pieri, A.; Korman, B.D.; Jüngel, A.; Wuertz-Kozak, K. Engineering Advanced In Vitro Models of Systemic Sclerosis for Drug Discovery and Development. Adv. Biol. 2021, 5, e2000168. [Google Scholar] [CrossRef]
- Kramer, B.; Corallo, C.; van den Heuvel, A.; Crawford, J.; Olivier, T.; Elstak, E.; Giordano, N.; Vulto, P.; Lanz, H.L.; Janssen, R.A.J.; et al. High-Throughput 3D Microvessel-on-a-Chip Model to Study Defective Angiogenesis in Systemic Sclerosis. Sci. Rep. 2022, 12, 16930. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Suhaeri, M.; Ha, S.S.; Oh, S.J.; Kim, S.-H.; Park, K. Human Lung Fibroblast-Derived Matrix Facilitates Vascular Morphogenesis in 3D Environment and Enhances Skin Wound Healing. Acta Biomater. 2017, 54, 333–344. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bodio, C.; Milesi, A.; Lonati, P.A.; Chighizola, C.B.; Mauro, A.; Pradotto, L.G.; Meroni, P.L.; Borghi, M.O.; Raschi, E. Fibroblasts and Endothelial Cells in Three-Dimensional Models: A New Tool for Addressing the Pathogenesis of Systemic Sclerosis as a Prototype of Fibrotic Vasculopathies. Int. J. Mol. Sci. 2024, 25, 2780. https://doi.org/10.3390/ijms25052780
Bodio C, Milesi A, Lonati PA, Chighizola CB, Mauro A, Pradotto LG, Meroni PL, Borghi MO, Raschi E. Fibroblasts and Endothelial Cells in Three-Dimensional Models: A New Tool for Addressing the Pathogenesis of Systemic Sclerosis as a Prototype of Fibrotic Vasculopathies. International Journal of Molecular Sciences. 2024; 25(5):2780. https://doi.org/10.3390/ijms25052780
Chicago/Turabian StyleBodio, Caterina, Alessandra Milesi, Paola Adele Lonati, Cecilia Beatrice Chighizola, Alessandro Mauro, Luca Guglielmo Pradotto, Pier Luigi Meroni, Maria Orietta Borghi, and Elena Raschi. 2024. "Fibroblasts and Endothelial Cells in Three-Dimensional Models: A New Tool for Addressing the Pathogenesis of Systemic Sclerosis as a Prototype of Fibrotic Vasculopathies" International Journal of Molecular Sciences 25, no. 5: 2780. https://doi.org/10.3390/ijms25052780
APA StyleBodio, C., Milesi, A., Lonati, P. A., Chighizola, C. B., Mauro, A., Pradotto, L. G., Meroni, P. L., Borghi, M. O., & Raschi, E. (2024). Fibroblasts and Endothelial Cells in Three-Dimensional Models: A New Tool for Addressing the Pathogenesis of Systemic Sclerosis as a Prototype of Fibrotic Vasculopathies. International Journal of Molecular Sciences, 25(5), 2780. https://doi.org/10.3390/ijms25052780