
Mass Spectrometry-Based Proteomic Analysis of Potential Host Proteins Interacting with GP5 in PRRSV-Infected PAMs

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
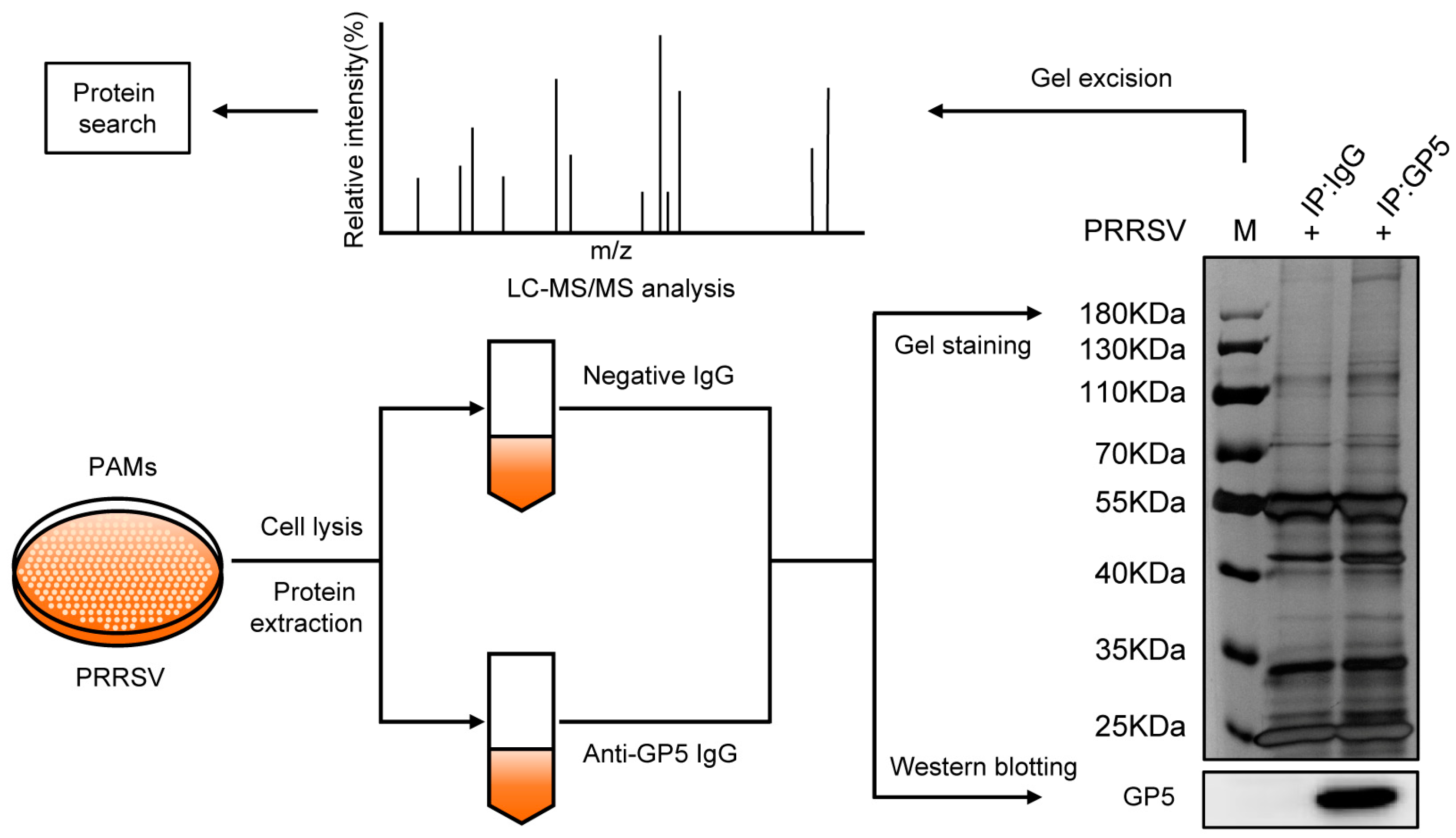
2.1. Identification of Potential GP5-Interacting Proteins in PAMs
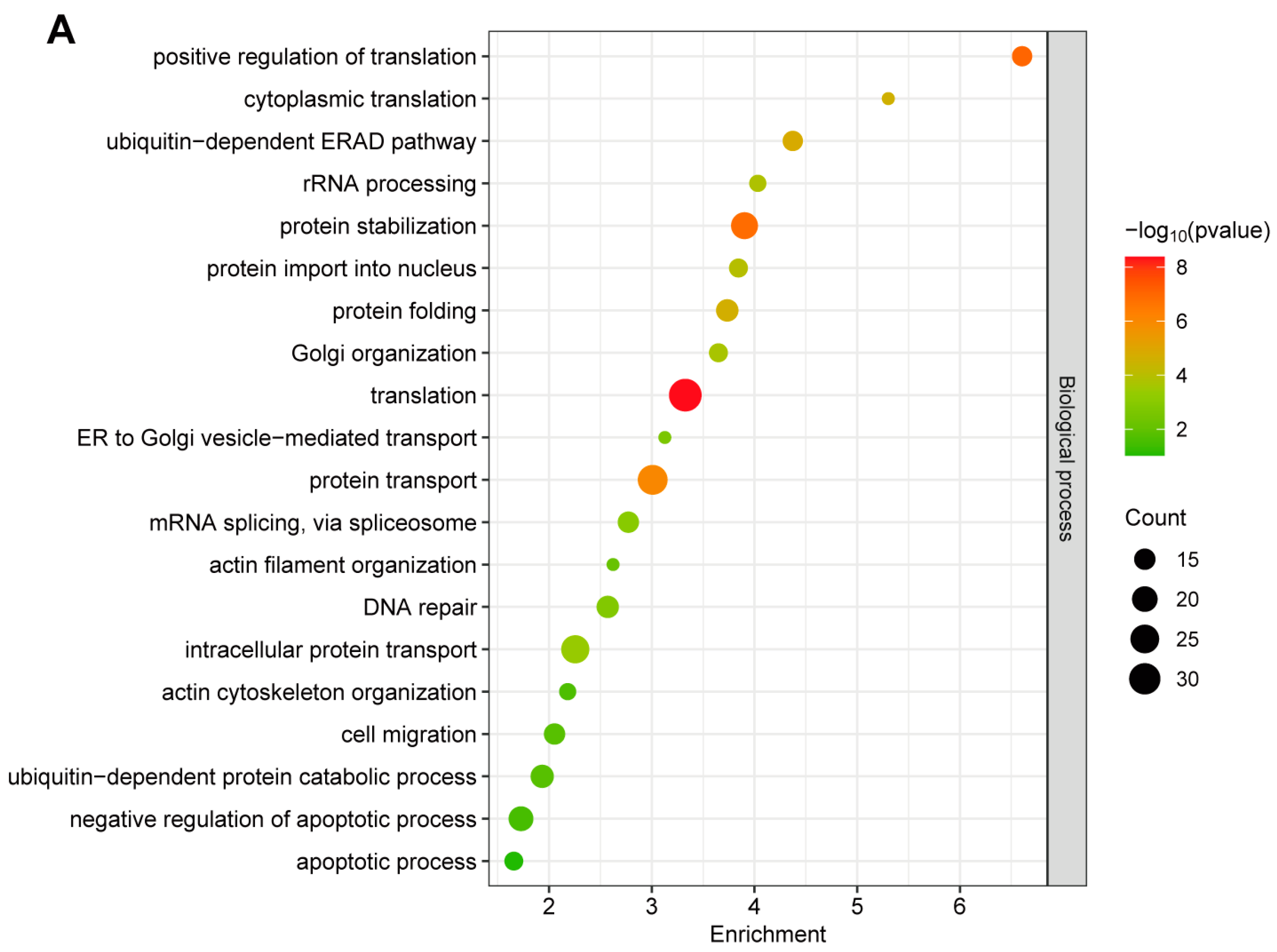
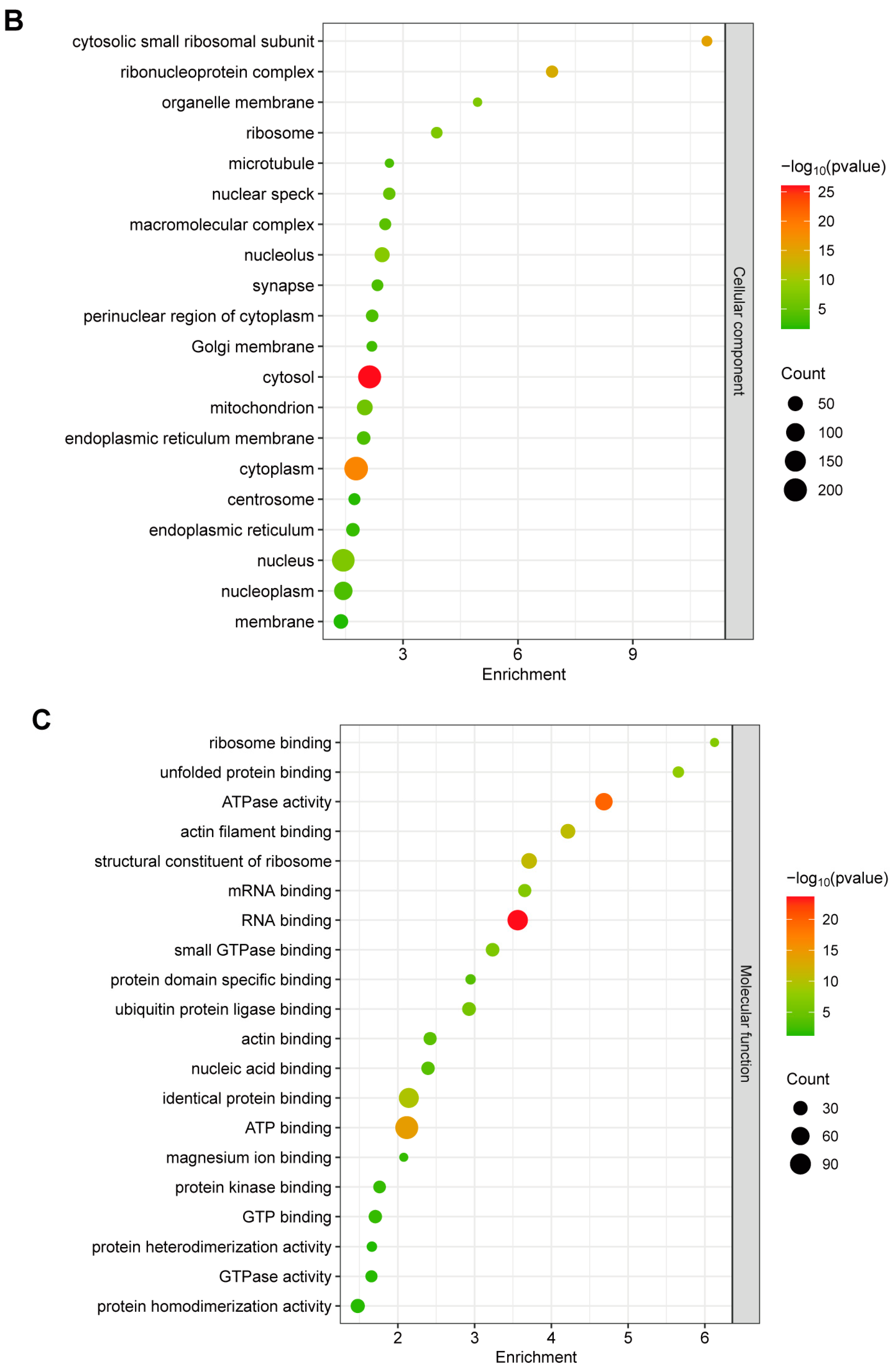
2.2. Gene Ontology (GO) Annotation of GP5-Interacting Proteins
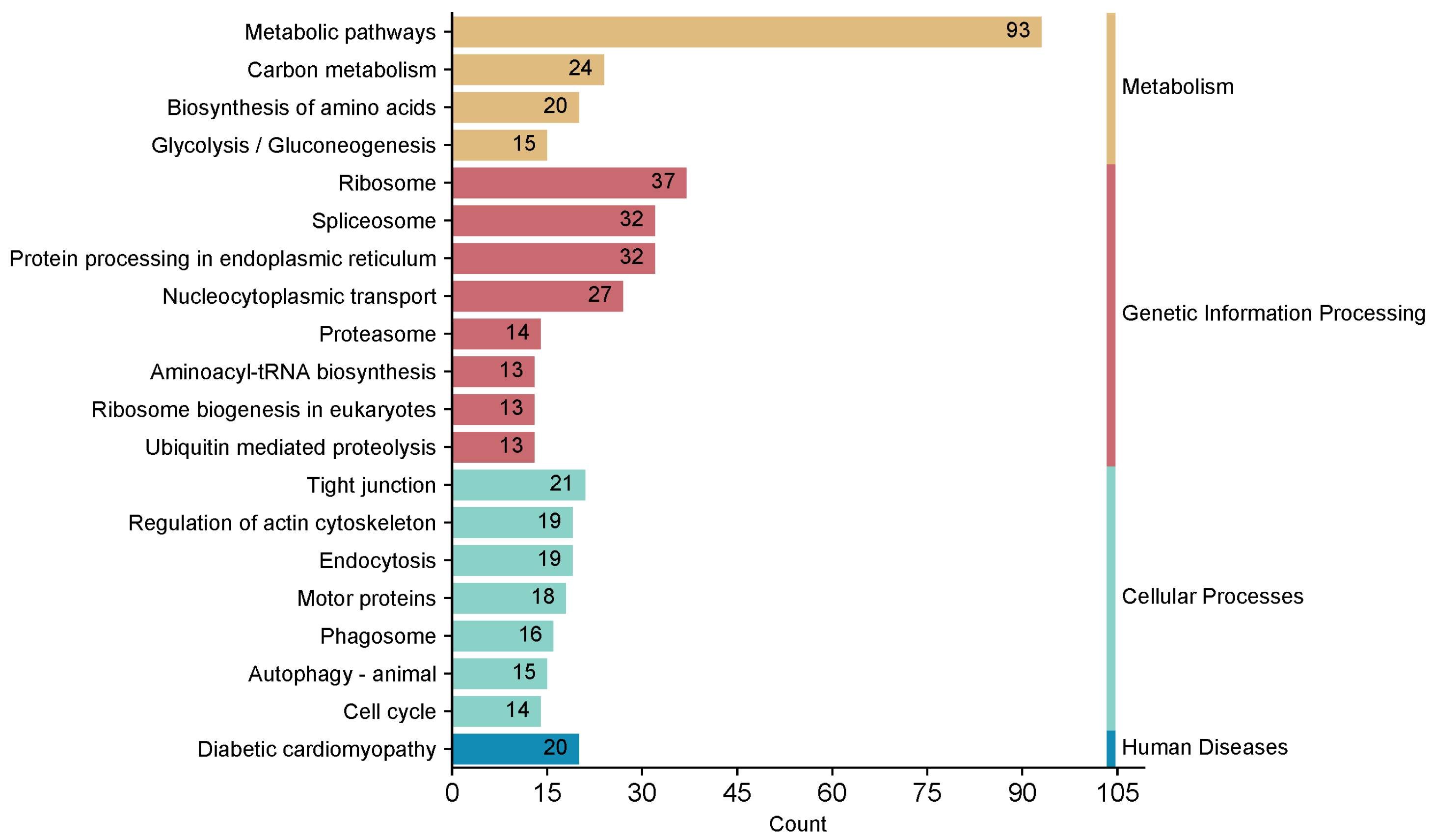
2.3. Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Annotation of GP5-Interacting Proteins
2.4. Protein–Protein Interaction (PPI) Network Analysis of GP5-Interacting Proteins
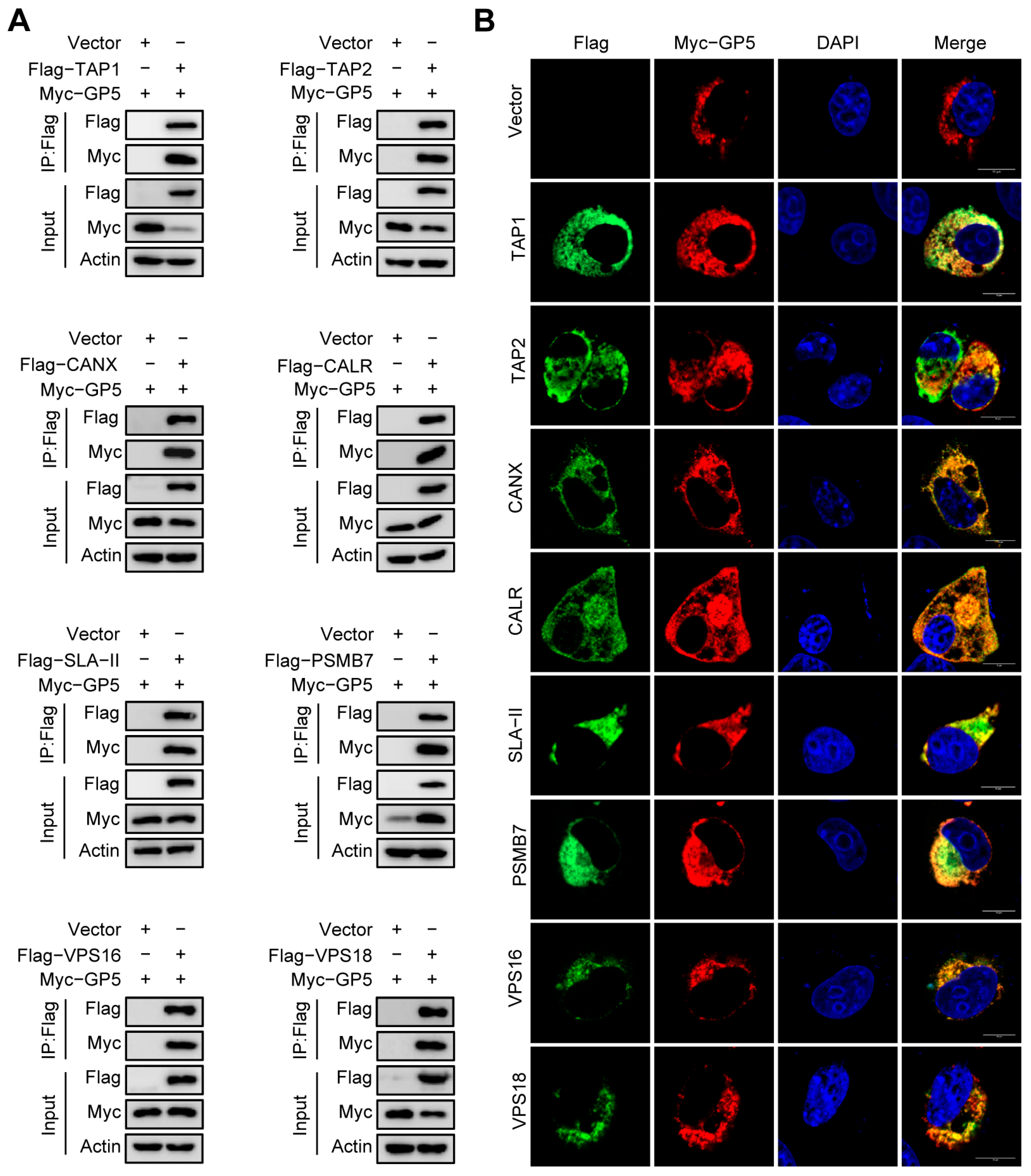
2.5. Validation of the Interaction between GP5 and Identified Host Proteins
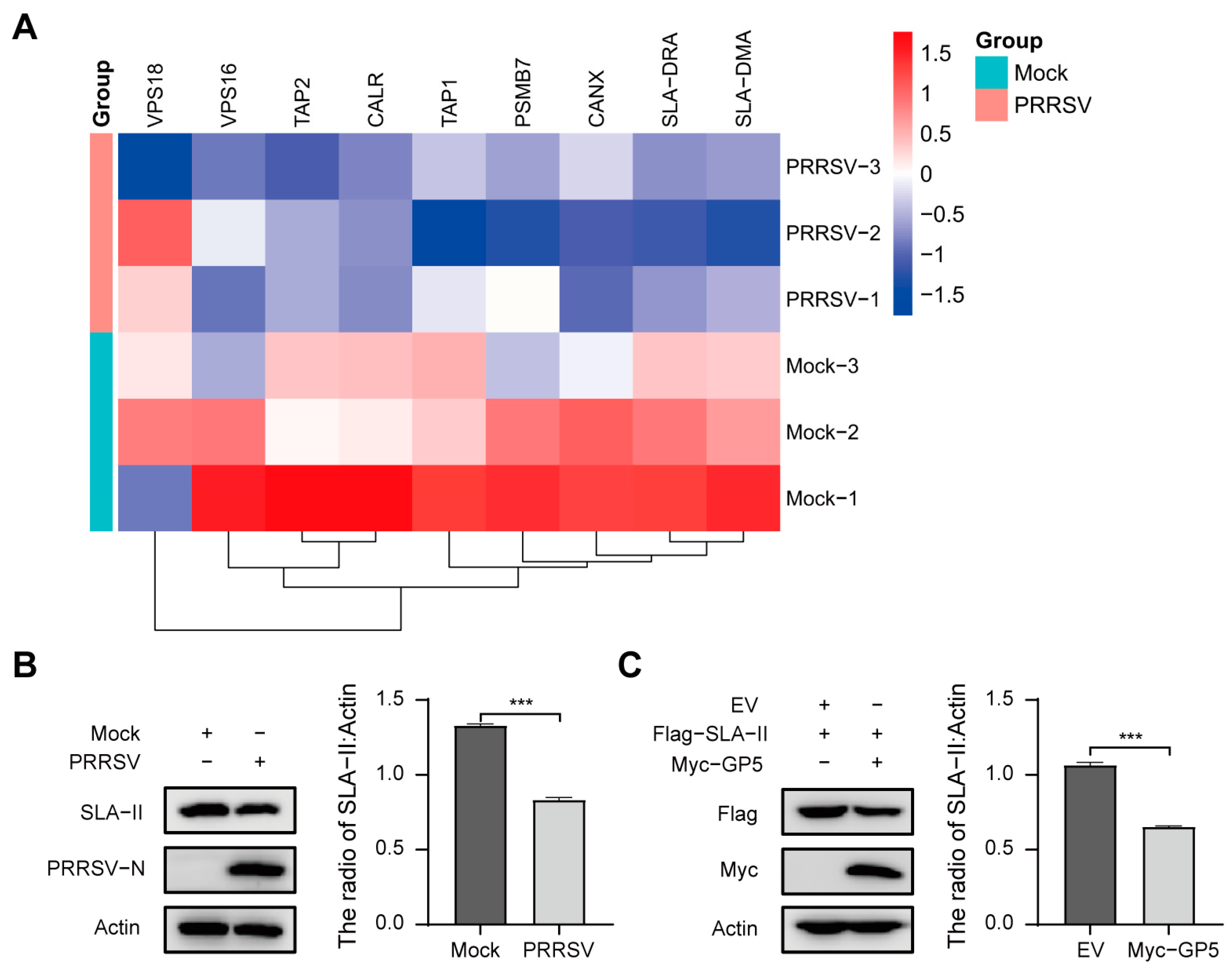
2.6. PRRSV May Inhibit the Progress of Antigen Processing and Presentation via GP5
3. Discussion
4. Materials and Methods
4.1. Cells, Virus, and Reagents
4.2. Plasmid Construction
4.3. Cell Transfection and Viral Infection
4.4. Co-IP Assay for LC-MS/MS Analysis
4.5. Sample Processing for TMT-Based Proteomics
4.6. LC-MS/MS Data Analysis
4.7. Bioinformatics Analysis
4.8. Co-IP Assay
4.9. Western Blotting
4.10. Immunofluorescence
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, L.J.; Yang, Y.; Xia, Q.Q.; Guan, Z.X.; Zhang, J.J.; Li, B.B.; Qiu, Y.F.; Liu, K.; Shao, D.H.; Ma, Z.Y.; et al. Genetic characterization of porcine reproductive and respiratory syndrome virus from Eastern China during 2017–2022. Front. Microbiol. 2022, 13, 971817. [Google Scholar] [CrossRef]
- Song, K.; Yu, J.Y.; Li, J.; Li, M.; Peng, L.Y.; Yi, P.F. Astragaloside IV Regulates cGAS-STING Signaling Pathway to Alleviate Immunosuppression Caused by PRRSV Infection. Viruses 2023, 15, 1586. [Google Scholar] [CrossRef]
- Su, N.Y.; Lin, Z.D.; Liu, X.; Sun, X.X.; Jin, X.X.; Feng, H.L.; Zhan, C.L.; Hu, X.Y.; Gu, C.Q.; Zhang, W.P.; et al. Pathological observation and transcriptomic analysis of thymus injury in PRRSV-infected piglets. Vet. Res. Commun. 2023, 47, 1949–1962. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.L.; Li, W.; Zhao, S.J.; Cui, Z.Y.; Chen, Y.; Zhang, Y.N.; Chen, J.; Xia, P.A. Proteomic Characterization of PAMs with PRRSV-ADE Infection. Viruses 2022, 15, 36. [Google Scholar] [CrossRef]
- Zhang, L.J.; Feng, X.; Wang, H.D.; He, S.J.; Fan, H.J.; Liu, D.Y. Antibody-dependent enhancement of porcine reproductive and respiratory syndrome virus infection downregulates the levels of interferon-gamma/lambdas in porcine alveolar macrophages in vitro. Front. Vet. Sci. 2023, 10, 1150430. [Google Scholar] [CrossRef]
- Wang, H.M.; Liu, Y.G.; Tang, Y.D.; Liu, T.X.; Zheng, L.L.; Wang, T.Y.; Liu, S.G.; Wang, G.; Cai, X.H. A natural recombinant PRRSV between HP-PRRSV JXA1-like and NADC30-like strains. Transbound. Emerg. Dis. 2018, 65, 1078–1086. [Google Scholar] [CrossRef]
- Risser, J.; Ackerman, M.; Evelsizer, R.; Wu, S.; Kwon, B.; Hammer, J.M. Porcine reproductive and respiratory syndrome virus genetic variability a management and diagnostic dilemma. Virol. J. 2021, 18, 206. [Google Scholar] [CrossRef] [PubMed]
- Sinkora, M.; Toman, M.; Stepanova, K.; Stepanova, H.; Leva, L.; Sinkorova, J.; Moutelikova, R.; Salat, J.; Srutkova, D.; Schwarzer, M.; et al. The mechanism of immune dysregulation caused by porcine reproductive and respiratory syndrome virus (PRRSV). Microbes Infect. 2023, 25, 105146. [Google Scholar] [CrossRef]
- Zhou, L.; Ge, X.N.; Yang, H.C. Porcine Reproductive and Respiratory Syndrome Modified Live Virus Vaccine: A “Leaky” Vaccine with Debatable Efficacy and Safety. Vaccines 2021, 9, 362. [Google Scholar] [CrossRef] [PubMed]
- Chae, C. Commercial PRRS Modified-Live Virus Vaccines. Vaccines 2021, 9, 185. [Google Scholar] [CrossRef]
- Nan, Y.C.; Wu, C.Y.; Gu, G.Q.; Sun, W.Y.; Zhang, Y.J.; Zhou, E.M. Improved Vaccine against PRRSV: Current Progress and Future Perspective. Front. Microbiol. 2017, 8, 1635. [Google Scholar] [CrossRef]
- Li, W.; Sun, Y.Y.; Zhao, S.J.; Cui, Z.Y.; Chen, Y.; Xu, P.L.; Chen, J.; Zhang, Y.N.; Xia, P.A. Differences in Humoral Immune Response against the Type 2 Porcine Reproductive and Respiratory Syndrome Virus via Different Immune Pathways. Viruses 2022, 14, 1435. [Google Scholar] [CrossRef]
- Kick, A.R.; Amaral, A.F.; Frias-De-Diego, A.; Cortes, L.M.; Fogle, J.E.; Crisci, E.; Almond, G.W.; Kaser, T. The Local and Systemic Humoral Immune Response Against Homologous and Heterologous Strains of the Type 2 Porcine Reproductive and Respiratory Syndrome Virus. Front. Immunol. 2021, 12, 637613. [Google Scholar] [CrossRef] [PubMed]
- Kappes, M.A.; Faaberg, K.S. PRRSV structure, replication and recombination: Origin of phenotype and genotype diversity. Virology 2015, 479–480, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.Z.; Lin, T.; Sun, L.C.; Li, Y.H.; Wang, X.M.; Gao, F.; Liu, R.X.; Chen, C.Y.; Tong, G.Z.; Yuan, S.S. N-linked glycosylation of GP5 of porcine reproductive and respiratory syndrome virus is critically important for virus replication in vivo. J. Virol. 2012, 86, 9941–9951. [Google Scholar] [CrossRef] [PubMed]
- Popescu, L.N.; Trible, B.R.; Chen, N.H.; Rowland, R.R.R. GP5 of porcine reproductive and respiratory syndrome virus (PRRSV) as a target for homologous and broadly neutralizing antibodies. Vet. Microbiol. 2017, 209, 90–96. [Google Scholar] [CrossRef]
- Du, L.P.; Yu, Z.Y.; Pang, F.J.; Xu, X.W.; Mao, A.H.; Yuan, W.Z.; He, K.W.; Li, B. Targeted Delivery of GP5 Antigen of PRRSV to M Cells Enhances the Antigen-Specific Systemic and Mucosal Immune Responses. Front. Cell. Infect. Microbiol. 2018, 8, 7. [Google Scholar] [CrossRef]
- Cui, J.R.; O’Connell, C.M.; Costa, A.; Pan, Y.; Smyth, J.A.; Verardi, P.H.; Burgess, D.J.; Van Kruiningen, H.J.; Garmendia, A.E. Correction: A PRRSV GP5-Mosaic vaccine: Protection of pigs from challenge and ex vivo detection of IFNgamma responses against several genotype 2 strains. PLoS ONE 2019, 14, e0213091. [Google Scholar] [CrossRef]
- Choi, J.C.; Kim, M.S.; Choi, H.Y.; Kang, Y.L.; Choi, I.Y.; Jung, S.W.; Jeong, J.Y.; Kim, M.C.; Cho, A.Y.; Lee, J.H.; et al. Porcine Reproductive and Respiratory Syndrome Virus Engineered by Serine Substitution on the 44th Amino Acid of GP5 Resulted in a Potential Vaccine Candidate with the Ability to Produce High Levels of Neutralizing Antibody. Vet. Sci. 2023, 10, 191. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Zhou, L.; He, B.C.; Zhang, X.H.; Ge, X.N.; Han, J.; Guo, X.; Yang, H.C. Nsp2 and GP5-M of Porcine Reproductive and Respiratory Syndrome Virus Contribute to Targets for Neutralizing Antibodies. Virol. Sin. 2019, 34, 631–640. [Google Scholar] [CrossRef]
- Fernandez, A.; Suarez, P.; Castro, J.M.; Tabares, E.; Diaz-Guerra, M. Characterization of regions in the GP5 protein of porcine reproductive and respiratory syndrome virus required to induce apoptotic cell death. Virus Res. 2002, 83, 103–118. [Google Scholar] [CrossRef]
- Vu, H.L.; Kwon, B.; Yoon, K.J.; Laegreid, W.W.; Pattnaik, A.K.; Osorio, F.A. Immune evasion of porcine reproductive and respiratory syndrome virus through glycan shielding involves both glycoprotein 5 as well as glycoprotein 3. J. Virol. 2011, 85, 5555–5564. [Google Scholar] [CrossRef]
- Sun, L.C.; Li, Y.H.; Liu, R.X.; Wang, X.M.; Gao, F.; Lin, T.; Huang, T.; Yao, H.C.; Tong, G.Z.; Fan, H.J.; et al. Porcine reproductive and respiratory syndrome virus ORF5a protein is essential for virus viability. Virus Res. 2013, 171, 178–185. [Google Scholar] [CrossRef]
- Wei, Y.Q.; Dai, G.; Huang, M.; Wen, L.H.; Chen, R.A.; Liu, D.X. Construction of an infectious cloning system of porcine reproductive and respiratory syndrome virus and identification of glycoprotein 5 as a potential determinant of virulence and pathogenicity. Front. Microbiol. 2023, 14, 1227485. [Google Scholar] [CrossRef]
- Huang, J.; Jiang, P.; Li, Y.F.; Xu, J.R.; Jiang, W.M.; Wang, X.W. Inhibition of porcine reproductive and respiratory syndrome virus replication by short hairpin RNA in MARC-145 cells. Vet. Microbiol. 2006, 115, 302–310. [Google Scholar] [CrossRef]
- Young, J.E.; Dvorak, C.M.T.; Graham, S.P.; Murtaugh, M.P. Isolation of Porcine Reproductive and Respiratory Syndrome Virus GP5-Specific, Neutralizing Monoclonal Antibodies From Hyperimmune Sows. Front. Immunol. 2021, 12, 638493. [Google Scholar] [CrossRef]
- Wang, L.P.; Ding, X.; Xiao, J.J.; Jiménez-Gόngora, T.; Liu, R.Y.; Lozano-Duran, R. Inference of a Geminivirus-Host Protein-Protein Interaction Network through Affinity Purification and Mass Spectrometry Analysis. Viruses 2017, 9, 275. [Google Scholar] [CrossRef]
- LaPointe, A.T.; Gebhart, N.N.; Meller, M.E.; Hardy, R.W.; Sokoloski, K.J. Identification and Characterization of Sindbis Virus RNA-Host Protein Interactions. J. Virol. 2018, 92, e02171-17. [Google Scholar] [CrossRef]
- Ackerman, E.E.; Kawakami, E.; Katoh, M.; Watanabe, T.; Watanabe, S.; Tomita, Y.; Lopes, T.J.; Matsuoka, Y.; Kitano, H.; Shoemaker, J.E.; et al. Network-Guided Discovery of Influenza Virus Replication Host Factors. MBio 2018, 9, e02002-18. [Google Scholar] [CrossRef]
- Shi, C.X.; Liu, Y.L.; Ding, Y.Z.; Zhang, Y.G.; Zhang, J. PRRSV receptors and their roles in virus infection. Arch. Microbiol. 2015, 197, 503–512. [Google Scholar] [CrossRef]
- Breedam, W.V.; Gorp, H.V.; Zhang, J.Q.; Crocker, P.R.; Delputte, P.L.; Nauwynck, H.J. The M/GP(5) glycoprotein complex of porcine reproductive and respiratory syndrome virus binds the sialoadhesin receptor in a sialic acid-dependent manner. PLoS Pathog. 2010, 6, e1000730. [Google Scholar] [CrossRef]
- Li, L.L.; Zhang, L.; Hu, Q.F.; Zhao, L.; Nan, Y.C.; Hou, G.P.; Chen, Y.Y.; Han, X.M.; Ren, X.L.; Zhao, Q.; et al. MYH9 Key Amino Acid Residues Identified by the Anti-Idiotypic Antibody to Porcine Reproductive and Respiratory Syndrome Virus Glycoprotein 5 Involve in the Virus Internalization by Porcine Alveolar Macrophages. Viruses 2019, 12, 40. [Google Scholar] [CrossRef]
- Zhang, M.; Zakhartchouk, A. Characterization of the interactome of the porcine reproductive and respiratory syndrome virus glycoprotein-5. Arch. Virol. 2018, 163, 1595–1605. [Google Scholar] [CrossRef]
- Liu, X.W.; Liu, X.; Bai, J.; Gao, Y.N.; Song, Z.B.; Nauwynck, H.; Wang, X.W.; Yang, Y.Q.; Jiang, P. Glyceraldehyde-3-Phosphate Dehydrogenase Restricted in Cytoplasmic Location by Viral GP5 Facilitates Porcine Reproductive and Respiratory Syndrome Virus Replication via Its Glycolytic Activity. J. Virol. 2021, 95, e0021021. [Google Scholar] [CrossRef]
- Hicks, J.A.; Yoo, D.; Liu, H.C. Interaction of porcine reproductive and respiratory syndrome virus major envelope proteins GP5 and M with the cellular protein Snapin. Virus Res. 2018, 249, 85–92. [Google Scholar] [CrossRef]
- Xue, B.Y.; Hou, G.P.; Zhang, G.X.; Huang, J.J.; Li, L.L.; Nan, Y.C.; Mu, Y.; Wang, L.Z.; Zhang, L.; Han, X.M.; et al. MYH9 Aggregation Induced by Direct Interaction With PRRSV GP5 Ectodomain Facilitates Viral Internalization by Permissive Cells. Front. Microbiol. 2019, 10, 2313. [Google Scholar] [CrossRef]
- Qi, P.F.; Liu, K.; Wei, J.C.; Li, Y.M.; Li, B.B.; Shao, D.H.; Wu, Z.C.; Shi, Y.Y.; Tong, G.Z.; Qiu, Y.F.; et al. Nonstructural Protein 4 of Porcine Reproductive and Respiratory Syndrome Virus Modulates Cell Surface Swine Leukocyte Antigen Class I Expression by Downregulating β2-Microglobulin Transcription. J. Virol. 2017, 91, e01755-16. [Google Scholar] [CrossRef]
- Wang, J.J.; Zhang, M.S.; Cui, X.C.; Gao, X.; Sun, W.F.; Ge, X.N.; Zhang, Y.N.; Guo, X.; Han, J.; Zhou, L.; et al. Attenuated Porcine Reproductive and Respiratory Syndrome Virus Regains Its Fatal Virulence by Serial Passaging in Pigs or Porcine Alveolar Macrophages To Increase Its Adaptation to Target Cells. Microbiol. Spectr. 2022, 10, e0308422. [Google Scholar] [CrossRef]
- Murata, S.; Yashiroda, H.; Tanaka, K. Molecular mechanisms of proteasome assembly. Nat. Rev. Mol. Cell Biol. 2009, 10, 104–115. [Google Scholar] [CrossRef]
- Oh, J.; Lee, C. Proteomic characterization of a novel structural protein ORF5a of porcine reproductive and respiratory syndrome virus. Virus Res. 2012, 169, 255–263. [Google Scholar] [CrossRef]
- Zhang, X.X.; Chen, Y.J.; Li, S.B.; Wang, J.L.; He, Z.; Yan, J.C.; Liu, X.H.; Guo, C.H. MARCO Inhibits Porcine Reproductive and Respiratory Syndrome Virus Infection through Intensifying Viral GP5-Induced Apoptosis. Microbiol. Spectr. 2023, 11, e0475322. [Google Scholar] [CrossRef]
- Xiong, Z.X.; Niu, X.Y.; Song, Y.B.; Danping, S.; Wang, F.; Chen, R.A.; He, D.S. Evolution of porcine reproductive and respiratory syndrome virus GP5 and GP3 genes under swIFN-beta immune pressure and interferon regulatory factor-3 activation suppressed by GP5. Res. Vet. Sci. 2015, 101, 175–179. [Google Scholar] [CrossRef]
- Smith, S.M. Strategies for the Purification of Membrane Proteins. Methods Mol. Biol. 2017, 1485, 389–400. [Google Scholar] [CrossRef]
- Feroz, H.; Kwon, H.; Peng, J.; Oh, H.; Ferlez, B.; Baker, C.S.; Golbeck, J.H.; Bazan, G.C.; Zydney, A.L.; Kumar, M. Improving extraction and post-purification concentration of membrane proteins. Analyst 2018, 143, 1378–1386. [Google Scholar] [CrossRef]
- Lubec, G.; Afjehi-Sadat, L. Limitations and pitfalls in protein identification by mass spectrometry. Chem. Rev. 2007, 107, 3568–3584. [Google Scholar] [CrossRef]
- Kotsias, F.; Cebrian, I.; Alloatti, A. Antigen processing and presentation. Int. Rev. Cell Mol. Biol. 2019, 348, 69–121. [Google Scholar] [CrossRef]
- Muntjewerff, E.M.; Meesters, L.D.; van den Bogaart, G. Antigen Cross-Presentation by Macrophages. Front. Immunol. 2020, 11, 1276. [Google Scholar] [CrossRef]
- Mantel, I.; Sadiq, B.A.; Blander, J.M. Spotlight on TAP and its vital role in antigen presentation and cross-presentation. Mol. Immunol. 2022, 142, 105–119. [Google Scholar] [CrossRef]
- Pishesha, N.; Harmand, T.J.; Ploegh, H.L. A guide to antigen processing and presentation. Nat. Rev. Immunol. 2022, 22, 751–764. [Google Scholar] [CrossRef]
- Kasahara, M.; Hayashi, M.; Tanaka, K.; Inoko, H.; Sugaya, K.; Ikemura, T.; Ishibashi, T. Chromosomal localization of the proteasome Z subunit gene reveals an ancient chromosomal duplication involving the major histocompatibility complex. Proc. Natl. Acad. Sci. USA 1996, 93, 9096–9101. [Google Scholar] [CrossRef]
- Wartosch, L.; Günesdogan, U.; Graham, S.C.; Luzio, J.P. Recruitment of VPS33A to HOPS by VPS16 Is Required for Lysosome Fusion with Endosomes and Autophagosomes. Traffic 2015, 16, 727–742. [Google Scholar] [CrossRef]
- Valečka, J.; Almeida, C.R.; Su, B.; Pierre, P.; Gatti, E. Autophagy and MHC-restricted antigen presentation. Mol. Immunol. 2018, 99, 163–170. [Google Scholar] [CrossRef]
- An, T.Q.; Li, J.N.; Su, C.M.; Yoo, D. Molecular and Cellular Mechanisms for PRRSV Pathogenesis and Host Response to Infection. Virus Res. 2020, 286, 197980. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, W.; Hu, D.; Su, R.; Ji, M.; Huang, Y.; Shereen, M.A.; Xu, X.; Luo, Z.; Zhang, Q.; et al. HIV-1 Nef Interacts with LMP7 To Attenuate Immunoproteasome Formation and Major Histocompatibility Complex Class I Antigen Presentation. MBio 2020, 11, e02221-19. [Google Scholar] [CrossRef]
- Zhang, F.; Zang, T.M.; Stevenson, E.M.; Lei, X.; Copertino, D.C.; Mota, T.M.; Boucau, J.; Garcia-Beltran, W.F.; Jones, R.B.; Bieniasz, P.D. Inhibition of major histocompatibility complex-I antigen presentation by sarbecovirus ORF7a proteins. Proc. Natl. Acad. Sci. USA 2022, 119, e2209042119. [Google Scholar] [CrossRef]
- Stumptner-Cuvelette, P.; Morchoisne, S.; Dugast, M.; Le Gall, S.; Raposo, G.; Schwartz, O.; Benaroch, P. HIV-1 Nef impairs MHC class II antigen presentation and surface expression. Proc. Natl. Acad. Sci. USA 2001, 98, 12144–12149. [Google Scholar] [CrossRef]
- Li, P.; Wang, N.; Zhou, D.; Yee, C.S.; Chang, C.H.; Brutkiewicz, R.R.; Blum, J.S. Disruption of MHC class II-restricted antigen presentation by vaccinia virus. J. Immunol. 2005, 175, 6481–6488. [Google Scholar] [CrossRef]
- Forsyth, K.S.; Eisenlohr, L.C. Giving CD4+ T cells the slip: Viral interference with MHC class II-restricted antigen processing and presentation. Curr. Opin. Immunol. 2016, 40, 123–129. [Google Scholar] [CrossRef]
- Cui, Z.; Zhou, L.; Zhao, S.; Li, W.; Li, J.; Chen, J.; Zhang, Y.; Xia, P. The Host E3-Ubiquitin Ligase TRIM28 Impedes Viral Protein GP4 Ubiquitination and Promotes PRRSV Replication. Int. J. Mol. Sci. 2023, 24, 10965. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Wang, Y.; Zhang, M.; Zhao, S.; Wang, M.; Zhao, R.; Chen, J.; Zhang, Y.; Xia, P. Mass Spectrometry-Based Proteomic Analysis of Potential Host Proteins Interacting with GP5 in PRRSV-Infected PAMs. Int. J. Mol. Sci. 2024, 25, 2778. https://doi.org/10.3390/ijms25052778
Li W, Wang Y, Zhang M, Zhao S, Wang M, Zhao R, Chen J, Zhang Y, Xia P. Mass Spectrometry-Based Proteomic Analysis of Potential Host Proteins Interacting with GP5 in PRRSV-Infected PAMs. International Journal of Molecular Sciences. 2024; 25(5):2778. https://doi.org/10.3390/ijms25052778
Chicago/Turabian StyleLi, Wen, Yueshuai Wang, Mengting Zhang, Shijie Zhao, Mengxiang Wang, Ruijie Zhao, Jing Chen, Yina Zhang, and Pingan Xia. 2024. "Mass Spectrometry-Based Proteomic Analysis of Potential Host Proteins Interacting with GP5 in PRRSV-Infected PAMs" International Journal of Molecular Sciences 25, no. 5: 2778. https://doi.org/10.3390/ijms25052778
APA StyleLi, W., Wang, Y., Zhang, M., Zhao, S., Wang, M., Zhao, R., Chen, J., Zhang, Y., & Xia, P. (2024). Mass Spectrometry-Based Proteomic Analysis of Potential Host Proteins Interacting with GP5 in PRRSV-Infected PAMs. International Journal of Molecular Sciences, 25(5), 2778. https://doi.org/10.3390/ijms25052778