


Radiotherapy Metastatic Prostate Cancer Cell Lines Treated with Gold Nanorods Modulate miRNA Signatures

 , ,
, ,  ,
,  , and
, and 
Abstract
1. Introduction
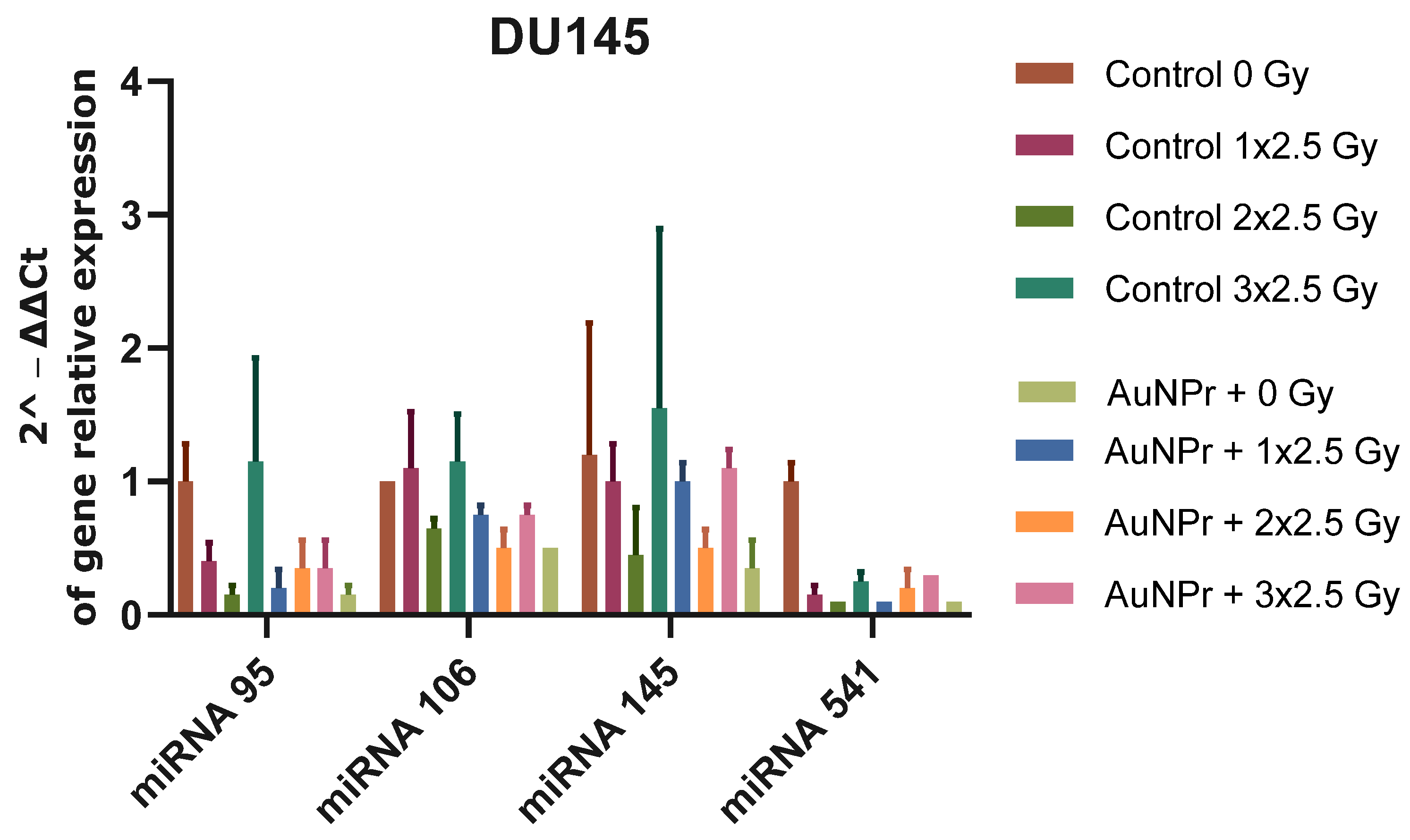
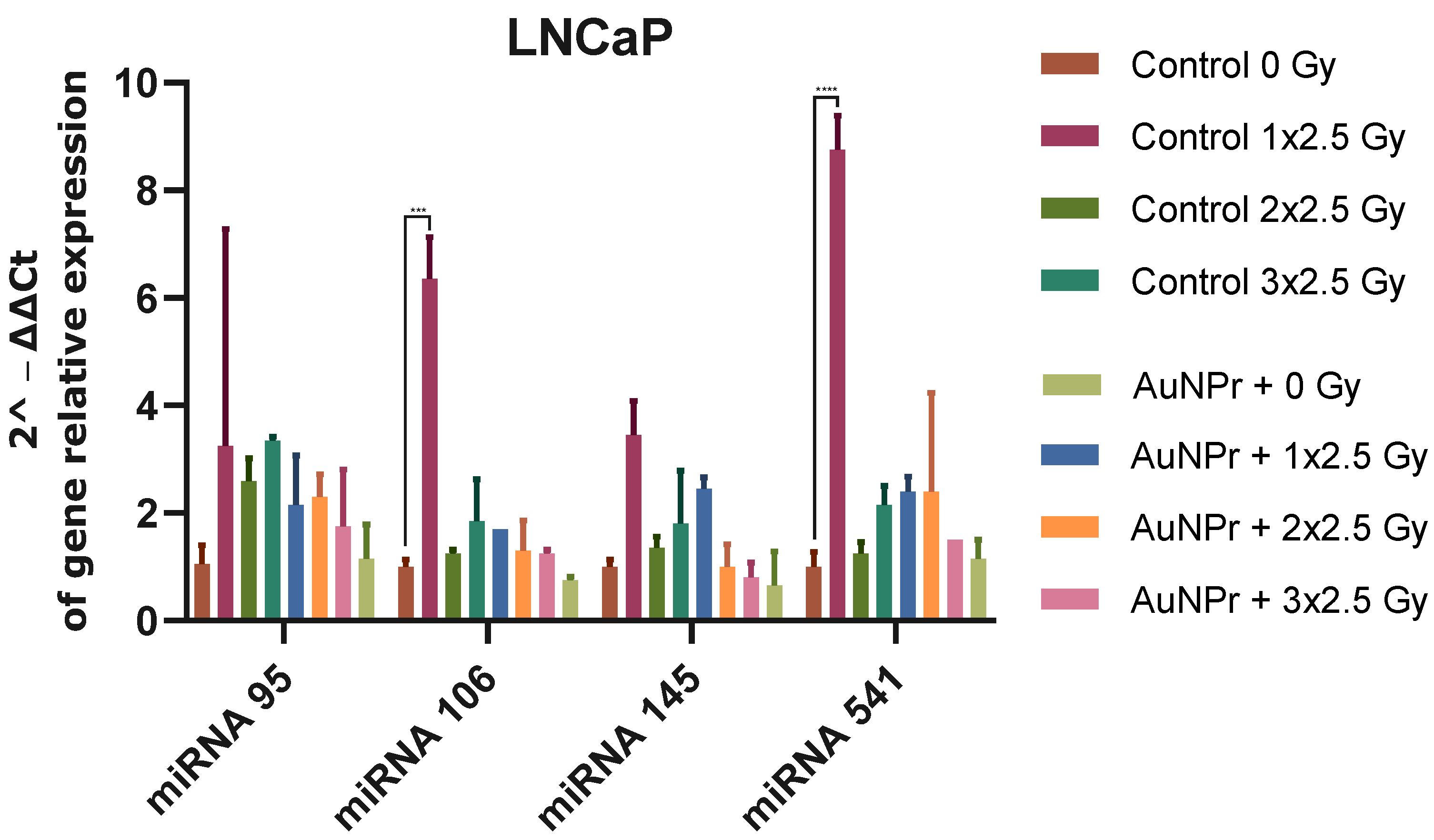
2. Results
3. Discussion
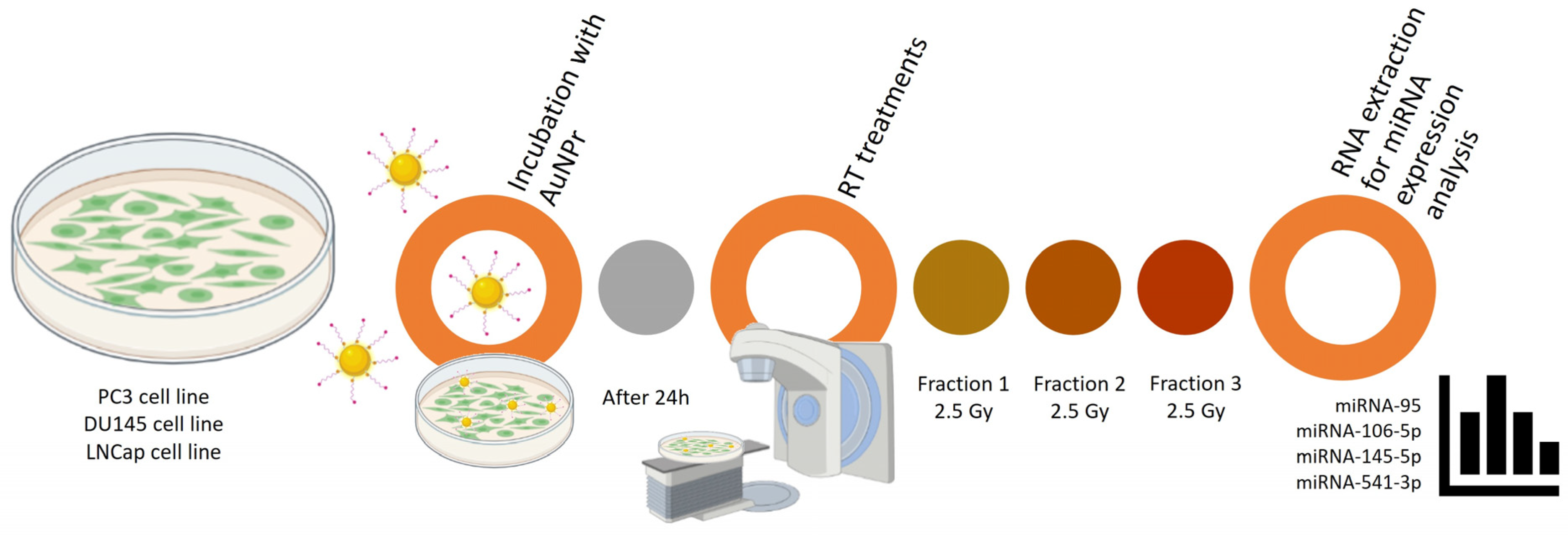
4. Materials and Methods
4.1. Reagents
4.2. Synthesis of Gold Nanorods
4.3. Cell Culture
4.4. Treatment Protocol
4.5. RNA Isolation and Gene Expression
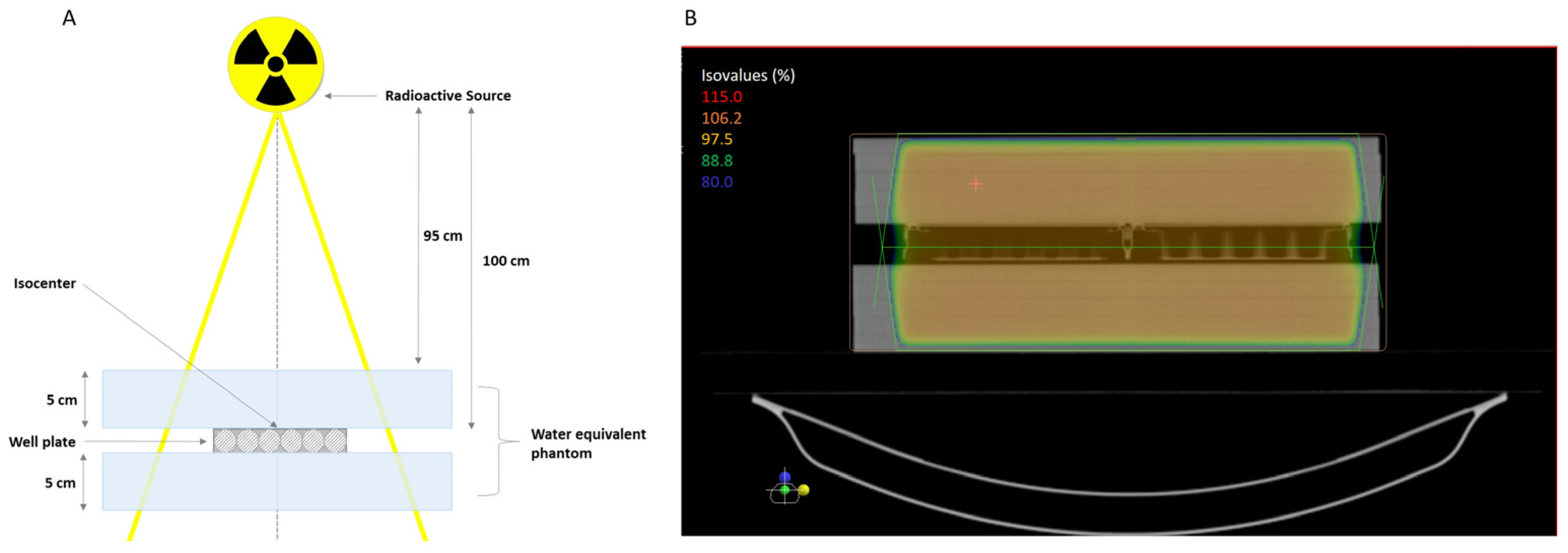
4.6. Cell Irradiation|Irradiation Setup
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peng, Y.; Croce, C.M. The role of micrornas in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed]
- Smolarz, B.; Durczyński, A.; Romanowicz, H.; Szyłło, K.; Hogendorf, P. miRNAs in Cancer (Review of Literature). Int. J. Mol. Sci. 2022, 23, 2805. [Google Scholar] [CrossRef] [PubMed]
- Otmani, K.; Lewalle, P. Tumor Suppressor miRNA in Cancer Cells and the Tumor Microenvironment: Mechanism of Deregulation and Clinical Implications. Front. Oncol. 2021, 11, 708765. [Google Scholar] [CrossRef] [PubMed]
- Forterre, A.; Komuro, H.; Aminova, S.; Harada, M. A Comprehensive Review of Cancer MicroRNA Therapeutic Delivery Strategies. Cancers 2020, 12, 1852. [Google Scholar] [CrossRef] [PubMed]
- Galvão-Lima, L.J.; Morais, A.H.F.; Valentim, R.A.M.; Barreto, E.J.S.S. miRNAs as biomarkers for early cancer detection and their application in the development of new diagnostic tools. Biomed. Eng. Online 2021, 20, 21. [Google Scholar] [CrossRef]
- Wang, L.; Lu, B.; He, M.; Wang, Y.; Wang, Z.; Du, L. Prostate Cancer Incidence and Mortality: Global Status and Temporal Trends in 89 Countries From 2000 to 2019. Front. Public Health 2022, 10, 811044. [Google Scholar] [CrossRef]
- Patel, A.R.; Klein, E.A. Risk factors for prostate cancer. Nat. Clin. Pract. Urol. 2009, 6, 87–95. [Google Scholar] [CrossRef]
- Rutz, J.; Benchellal, A.; Kassabra, W.; Maxeiner, S.; Bernd, A.; Kippenberger, S.; Zöller, N.; Chun, F.K.-H.; Juengel, E.; Blaheta, R.A. Growth, Proliferation and Metastasis of Prostate Cancer Cells Is Blocked by Low-Dose Curcumin in Combination with Light Irradiation. Int. J. Mol. Sci. 2021, 22, 9966. [Google Scholar] [CrossRef]
- Parker, C.; Castro, E.; Fizazi, K.; Heidenreich, A.; Ost, P.; Procopio, G.; Tombal, B.; Gillessen, S. Prostate cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2020, 31, 1119–1134. [Google Scholar] [CrossRef] [PubMed]
- Global Cancer Observatory: Cancer Today. Lyon, France: International Agency for Research on Cancer. 2020. Available online: http://gco.iarc.fr/today/home (accessed on 23 September 2023).
- Shou, J.; Zhang, Q.; Wang, S.; Zhang, D. The prognosis of different distant metastases pattern in prostate cancer: A population based retrospective study. Prostate 2018, 78, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Li, Y.; Wang, J.; Gao, X.; Zhong, Q.; He, L.; Li, C.; Liu, M.; Liu, Y.; Ma, M.; et al. Guidelines for radiotherapy of prostate cancer (2020 edition). Precis. Radiat. Oncol. 2021, 5, 160–182. [Google Scholar] [CrossRef]
- Ni, J.; Bucci, J.; Chang, L.; Malouf, D.; Graham, P.; Li, Y. Targeting MicroRNAs in Prostate Cancer Radiotherapy. Theranostics 2017, 7, 3243–3259. [Google Scholar] [CrossRef]
- Tang, C.; Qi, J.; Wu, Y.; Luo, L.; Wang, Y.; Wu, Y.; Shi, X. Improving the prediction for the response to radiotherapy of clinical tumor samples by using combinatorial model of MicroRNA expression. Front. Genet. 2022, 13, 1069112. [Google Scholar] [CrossRef] [PubMed]
- Kachris, S.; Papadaki, C.; Rounis, K.; Tsitoura, E.; Kokkinaki, C.; Nikolaou, C.; Sourvinos, G.; Mavroudis, D. Circulating mirnas as potential biomarkers in prostate cancer patients undergoing radiotherapy. Cancer Manag. Res. 2021, 13, 8257–8271. [Google Scholar] [CrossRef] [PubMed]
- Ghamlouche, F.; Yehya, A.; Zeid, Y.; Fakhereddine, H.; Fawaz, J.; Liu, Y.-N.; Al-Sayegh, M.; Abou-Kheir, W. MicroRNAs as clinical tools for diagnosis, prognosis, and therapy in prostate cancer. Transl. Oncol. 2023, 28, 101613. [Google Scholar] [CrossRef] [PubMed]
- Soares, S.; Guerreiro, S.G.; Cruz-Martins, N.; Faria, I.; Baylina, P.; Sales, M.G.; Correa-Duarte, M.A.; Fernandes, R. The Influence of miRNAs on Radiotherapy Treatment in Prostate Cancer—A Systematic Review. Front. Oncol. 2021, 11, 704664. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Croce, C.M. microRNA involvement in human cancer. Carcinogenesis 2012, 33, 1126–1133. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, S.I. microRNA therapies in cancer. Mol. Cell. Ther. 2014, 2, 7. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, J.; Fu, S.; Wu, J. Gold Nanoparticles as Radiosensitizers in Cancer Radiotherapy. Int. J. Nanomed. 2020, 15, 9407–9430. [Google Scholar] [CrossRef]
- Piccolo, O.; Lincoln, J.D.; Melong, N.; Orr, B.C.; Fernandez, N.R.; Borsavage, J.; Berman, J.N.; Robar, J.; Ha, M.N. Radiation dose enhancement using gold nanoparticles with a diamond linear accelerator target: A multiple cell type analysis. Sci. Rep. 2022, 12, 1559. [Google Scholar] [CrossRef]
- Soares, S.; Faria, I.; Aires, F.; Monteiro, A.; Pinto, G.; Sales, M.G.; Correa-Duarte, M.A.; Guerreiro, S.G.; Fernandes, R. Application of Gold Nanoparticles as Radiosensitizer for Metastatic Prostate Cancer Cell Lines. Int. J. Mol. Sci. 2023, 24, 4122. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zheng, X.; Chen, L.; Gong, X.; Yang, H.; Duan, X.; Zhu, Y. Multifunctional Gold Nanoparticles in Cancer Diagnosis and Treatment. Int. J. Nanomed. 2022, 17, 2041–2067. [Google Scholar] [CrossRef]
- MacFarlane, L.-A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.C.; Doudna, J.A. Molecular Mechanisms of RNA Interference. Annu. Rev. Biophys. 2013, 42, 217–239. [Google Scholar] [CrossRef] [PubMed]
- Cooper, B.T.; Sanfilippo, N.J. Concurrent chemoradiation for high-risk prostate cancer. World J. Clin. Oncol. 2015, 6, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Aminabad, N.S.; Farshbaf, M.; Akbarzadeh, A. Recent Advances of Gold Nanoparticles in Biomedical Applications: State of the Art. Cell Biochem. Biophys. 2019, 77, 123–137. [Google Scholar] [CrossRef]
- Huang, X.; Taeb, S.; Jahangiri, S.; Emmenegger, U.; Tran, E.; Bruce, J.; Mesci, A.; Korpela, E.; Vesprini, D.; Wong, C.S.; et al. miRNA-95 Mediates Radioresistance in Tumors by Targeting the Sphingolipid Phosphatase SGPP1. Cancer Res. 2013, 73, 6972–6986. [Google Scholar] [CrossRef]
- Razdan, A.; de Souza, P.; Roberts, T.L. Role of MicroRNAs in Treatment Response in Prostate Cancer. Curr. Cancer Drug Targets 2018, 18, 929–944. [Google Scholar] [CrossRef]
- Ambs, S.; Prueitt, R.L.; Yi, M.; Hudson, R.S.; Howe, T.M.; Petrocca, F.; Wallace, T.A.; Liu, C.-G.; Volinia, S.; Calin, G.A.; et al. Genomic Profiling of MicroRNA and Messenger RNA Reveals Deregulated MicroRNA Expression in Prostate Cancer. Cancer Res. 2008, 68, 6162–6170. [Google Scholar] [CrossRef]
- Li, B.; Shi, X.; Nori, D.; Chao, C.K.; Chen, A.M.; Valicenti, R.; White, R.d.V. Down-regulation of microRNA 106b is involved in p21-mediated cell cycle arrest in response to radiation in prostate cancer cells. Prostate 2011, 71, 567–574. [Google Scholar] [CrossRef]
- Xue, G.; Ren, Z.; Chen, Y.; Zhu, J.; Du, Y.; Pan, D.; Li, X.; Hu, B. A feedback regulation between miR-145 and DNA methyltransferase 3b in prostate cancer cell and their responses to irradiation. Cancer Lett. 2015, 361, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Josson, S.; Sung, S.; Lao, K.; Chung, L.W.; Johnstone, P.A. Radiation modulation of MicroRNA in prostate cancer cell lines. Prostate 2008, 68, 1599–1606. [Google Scholar] [CrossRef]
- Gong, P.; Zhang, T.; He, D.; Hsieh, J.-T. MicroRNA-145 Modulates Tumor Sensitivity to Radiation in Prostate Cancer. Radiat. Res. 2015, 184, 630–638. [Google Scholar] [CrossRef]
- Hatano, K.; Kumar, B.; Zhang, Y.; Coulter, J.B.; Hedayati, M.; Mears, B.; Ni, X.; Kudrolli, T.A.; Chowdhury, W.H.; Rodriguez, R.; et al. A functional screen identifies miRNAs that inhibit DNA repair and sensitize prostate cancer cells to ionizing radiation. Nucleic Acids Res. 2015, 43, 4075–4086. [Google Scholar] [CrossRef] [PubMed]
- El Bezawy, R.; Tripari, M.; Percio, S.; Cicchetti, A.; Tortoreto, M.; Stucchi, C.; Tinelli, S.; Zuco, V.; Doldi, V.; Gandellini, P.; et al. SPOP Deregulation Improves the Radiation Response of Prostate Cancer Models by Impairing DNA Damage Repair. Cancers 2020, 12, 1462. [Google Scholar] [CrossRef]
- He, Z.; Shen, F.; Qi, P.; Zhai, Z.; Wang, Z. miR-541-3p enhances the radiosensitivity of prostate cancer cells by inhibiting HSP27 expression and downregulating β-catenin. Cell Death Discov. 2021, 7, 18. [Google Scholar] [CrossRef]
- Leung, C.-M.; Li, S.-C.; Chen, T.-W.; Ho, M.-R.; Hu, L.-Y.; Liu, W.-S.; Wu, T.T.; Hsu, P.-C.; Chang, H.-T.; Tsai, K.-W. Comprehensive microRNA profiling of prostate cancer cells after ionizing radiation treatment. Oncol. Rep. 2014, 31, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Sousa, D.P.; Conde, J. Gold Nanoconjugates for miRNA Modulation in Cancer Therapy: From miRNA Silencing to miRNA Mimics. ACS Mater. Au 2022, 2, 626–640. [Google Scholar] [CrossRef]
- Zhu, L.; Wang, M.; Chen, N.; Zhang, Y.; Xu, T.; Zhuang, W.; Xiao, S.; Dai, L. Mechanisms of microRNA action in rectal cancer radiotherapy. Chin. Med. J. 2022, 135, 2017–2025. [Google Scholar] [CrossRef]
- Ma, W.; Ma, C.-N.; Zhou, N.-N.; Li, X.-D.; Zhang, Y.-J. Up-regulation of miR-328-3p sensitizes non-small cell lung cancer to radiotherapy. Sci. Rep. 2016, 6, 31651. [Google Scholar] [CrossRef]
- Ma, W.; Ma, C.-N.; Li, X.-D.; Zhang, Y.-J. Examining the effect of gene reduction in miR-95 and enhanced radiosensitivity in non-small cell lung cancer. Cancer Gene Ther. 2016, 23, 66–71. [Google Scholar] [CrossRef]
- Zheng, L.; Zhang, Y.; Liu, Y.; Zhou, M.; Lu, Y.; Yuan, L.; Zhang, C.; Hong, M.; Wang, S.; Li, X. MiR-106b induces cell radioresistance via the PTEN/PI3K/AKT pathways and p21 in colorectal cancer. J. Transl. Med. 2015, 13, 252. [Google Scholar] [CrossRef]
- Ivanovska, I.; Ball, A.S.; Diaz, R.L.; Magnus, J.F.; Kibukawa, M.; Schelter, J.M.; Kobayashi, S.V.; Lim, L.; Burchard, J.; Jackson, A.L.; et al. Micrornas in the mir-106b family regulate p21/cdkn1a and promote cell cycle progression. Mol. Cell. Biol. 2008, 28, 2167–2174. [Google Scholar] [CrossRef]
- Hudson, R.S.; Yi, M.; Esposito, D.; A Glynn, S.; Starks, A.M.; Yang, Y.; Schetter, A.J.; Watkins, S.K.; A Hurwitz, A.; Dorsey, T.H.; et al. MicroRNA-106b-25 cluster expression is associated with early disease recurrence and targets caspase-7 and focal adhesion in human prostate cancer. Oncogene 2013, 32, 4139–4147. [Google Scholar] [CrossRef]
- Petrocca, F.; Visone, R.; Onelli, M.R.; Shah, M.H.; Nicoloso, M.S.; de Martino, I.; Iliopoulos, D.; Pilozzi, E.; Liu, C.G.; Negrini, M.; et al. E2f1-regulated micrornas impair tgfbeta-dependent cell-cycle arrest and apoptosis in gastric cancer. Cancer Cell 2008, 13, 272–286. [Google Scholar] [CrossRef] [PubMed]
- Larne, O.; Hagman, Z.; Lilja, H.; Bjartell, A.; Edsjö, A.; Ceder, Y. miR-145 suppress the androgen receptor in prostate cancer cells and correlates to prostate cancer prognosis. Carcinogenesis 2015, 36, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Malla, B.; Zaugg, K.; Vassella, E.; Aebersold, D.M.; Pra, A.D. Exosomes and Exosomal MicroRNAs in Prostate Cancer Radiation Therapy. Int. J. Radiat. Oncol. Biol. Phys. 2017, 98, 982–995. [Google Scholar] [CrossRef] [PubMed]
- Malla, B.; Aebersold, D.M.; Pra, A.D. Protocol for serum exosomal miRNAs analysis in prostate cancer patients treated with radiotherapy. J. Transl. Med. 2018, 16, 223. [Google Scholar] [CrossRef] [PubMed]
- Borges, H.L.; Linden, R.; Wang, J.Y. DNA damage-induced cell death: Lessons from the central nervous system. Cell Res. 2008, 18, 17–26. [Google Scholar] [CrossRef] [PubMed]
- El Bezawy, R.; Tinelli, S.; Tortoreto, M.; Doldi, V.; Zuco, V.; Folini, M.; Stucchi, C.; Rancati, T.; Valdagni, R.; Gandellini, P.; et al. Mir-205 enhances radiation sensitivity of prostate cancer cells by impairing dna damage repair through pkcepsilon and zeb1 inhibition. J. Exp. Clin. Cancer Res. 2019, 38, 51. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Tao, W.; Ni, S.; Chen, Q.; Zhao, Z.; Ma, L.; Fu, Y.; Jiao, Z. Tumor-suppressive microrna-145 induces growth arrest by targeting senp1 in human prostate cancer cells. Cancer Sci. 2015, 106, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Scarabelli, L.; Sánchez-Iglesias, A.; Pérez-Juste, J.; Liz-Marzán, L.M. A “tips and tricks” practical guide to the synthesis of gold nanorods. J. Phys. Chem. Lett. 2015, 6, 4270–4279. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Cell Line Used in the Literature | Function | Target | miRNA Expression after Irradiation | Functional Role | Therapeutic Strategy | References | |
|---|---|---|---|---|---|---|---|---|
| In the Literature (2–8 Gy) | Our Study (3 × 2.5 Gy) | |||||||
| hsa-miRNA-95 | PC3 | - | SGPPI | ↑ (6 Gy) | ↑ PC3, LNCaP, HPrEPiC↓ DU145 | RR | Antagomirs | [28,29] |
| hsa-miRNA-106b | LNCaP | OM | P21 P53 Caspase-7 | ↓ (6 Gy) | ↑ PC3, LNCaP, HPrEPiC ↓ DU145 | RR | Antagomirs | [30,31] |
| hsa-miRNA-145 | LNCaP, PC3 | TS | DNMT3b SPOP ZEB1 | ↑ (2 Gy) | ↑ PC3, DU145, HPrEPiC ↓ LNCAP | RR | Mimicking | [32,33,34,35,36] |
| hsa-miRNA-541-3p | LNCaP, DU145, PC3, and PrEC | TS | HSP27 | ↑ (2–8 Gy) | ↑ PC3, LNCaP, HPrEPiC ↓ DU145 | RR | Mimicking | [37] |
| MiRNA | Primer Forward |
|---|---|
| miR-95 | UUCAACGGGUAUUUAUUGAGCA |
| miR-106-5p | UAAAGUGCUGACAGUGCAGAU |
| miR-145-5p | GUCCAGUUUUCCCAGGAAUCCCU |
| miR-541-3p | UGGUGGGCACAGAAUCUGGACU |
| Universal | Universal PCR Primer |
| RNU6 | GUGCUCGCUUCGGCAGCACAUAUACUAAAAUUGGAACGAUACAGAGAAGAUUAGCAUGGCCCCUGCGCAAGGAU-GACACGCAAAUUCGUGAAGCGUUCCAUAUUUU |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares, S.; Aires, F.; Monteiro, A.; Pinto, G.; Faria, I.; Sales, G.; Correa-Duarte, M.A.; Guerreiro, S.; Fernandes, R. Radiotherapy Metastatic Prostate Cancer Cell Lines Treated with Gold Nanorods Modulate miRNA Signatures. Int. J. Mol. Sci. 2024, 25, 2754. https://doi.org/10.3390/ijms25052754
Soares S, Aires F, Monteiro A, Pinto G, Faria I, Sales G, Correa-Duarte MA, Guerreiro S, Fernandes R. Radiotherapy Metastatic Prostate Cancer Cell Lines Treated with Gold Nanorods Modulate miRNA Signatures. International Journal of Molecular Sciences. 2024; 25(5):2754. https://doi.org/10.3390/ijms25052754
Chicago/Turabian StyleSoares, Sílvia, Fátima Aires, Armanda Monteiro, Gabriela Pinto, Isabel Faria, Goreti Sales, Miguel A. Correa-Duarte, Susana Guerreiro, and Rúben Fernandes. 2024. "Radiotherapy Metastatic Prostate Cancer Cell Lines Treated with Gold Nanorods Modulate miRNA Signatures" International Journal of Molecular Sciences 25, no. 5: 2754. https://doi.org/10.3390/ijms25052754
APA StyleSoares, S., Aires, F., Monteiro, A., Pinto, G., Faria, I., Sales, G., Correa-Duarte, M. A., Guerreiro, S., & Fernandes, R. (2024). Radiotherapy Metastatic Prostate Cancer Cell Lines Treated with Gold Nanorods Modulate miRNA Signatures. International Journal of Molecular Sciences, 25(5), 2754. https://doi.org/10.3390/ijms25052754