Abstract
The use of gene-editing tools, such as zinc finger nucleases, TALEN, and CRISPR/Cas, allows for the modification of physiological, morphological, and other characteristics in a wide range of crops to mitigate the negative effects of stress caused by anthropogenic climate change or biotic stresses. Importantly, these tools have the potential to improve crop resilience and increase yields in response to challenging environmental conditions. This review provides an overview of gene-editing techniques used in plants, focusing on the cultivated tomatoes. Several dozen genes that have been successfully edited with the CRISPR/Cas system were selected for inclusion to illustrate the possibilities of this technology in improving fruit yield and quality, tolerance to pathogens, or responses to drought and soil salinity, among other factors. Examples are also given of how the domestication of wild species can be accelerated using CRISPR/Cas to generate new crops that are better adapted to the new climatic situation or suited to use in indoor agriculture.
1. Introduction
Current global agricultural production faces significant losses due to increased environmental stress caused by climate change. This stress results from the fluctuations in temperature and precipitation, along with related factors such as soil salinization [1,2]. Consequently, the development of new tools to alleviate the negative effects of the current climate change scenario is crucial. In one avenue of development, to effectively respond to the various stresses caused by anthropogenic climate change, crops are currently undergoing genetic modification through new genomic techniques (NGTs) to enhance their resilience and adaptability [3]. Gene-editing tools, such as zinc finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), and clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated proteins (Cas), are the most commonly used NGTs for crop improvement.
Tomatoes (Solanum lycopersicum L.) are the most widely grown vegetable crop, with 186.1 million metric tons produced worldwide in 2022 [4]. However, drought, increasing global temperatures, the spread of insect pests, bacteria, and viruses pose a threat to their current production [5]. To overcome this, gene-editing tools have increased the tolerance of tomato plants to adverse factors, including heat, drought, salinity, bacteria, and viruses. Furthermore, tomato plants with increased levels of lycopene and carotenoids, along with other biochemical enhancements, have been effectively produced using gene-editing tools. Against this background, in this review, we present an overview of the gene-editing tools that are available and the strategies used to obtain edited plants, the results attained in tomatoes when using these tools, and a discussion of the weaknesses and strengths of the current approaches.
2. Gene-Editing Systems in Plants
2.1. Zinc Finger Nucleases (ZFNs)
Cys2-His2 (C2H2) zinc finger domains are present in many eukaryotic transcription factors. The classical C2H2 domain, with 28–30 amino acids, comprises two β-strands and an α-helix stabilized by a Zn2+ atom through conserved cysteine and histidine residues. Each C2H2 domain binds 3–4 nucleotides of DNA. Furthermore, the fusion of tandem C2H2 domains, separated by a conserved sequence of five amino acids, enables the binding to specific, longer DNA sequences. The zinc finger transcription factors were discovered in 1985 [6] and have since been utilized to regulate gene expression. ZFNs are chimeric proteins that comprise multiple C2H2 zinc finger domains, up to three, which are fused to the C-terminal cleavage domain of the endonuclease FokI, rendering ZFNs pioneering gene-editing tools to have been made accessible. The DNA sequence specificity of composite arrays of C2H2 zinc finger domains utilized to construct ZFNs determines the specificity of endonuclease cleavage. Dimerization of two different ZFN proteins results in a double strand break (DSB) in DNA with 5–7 nucleotide 5′ overhangs, as the Fok1 cleavage domain needs dimerization to cleave DNA (Figure 1a) [7,8]. The resulting DSBs in the DNA initiate endogenous mechanisms of DNA repair, predominantly through the non-homologous end-joining (NHEJ) pathway, which leads to the appearance of deletions and insertions at the repair site [9].
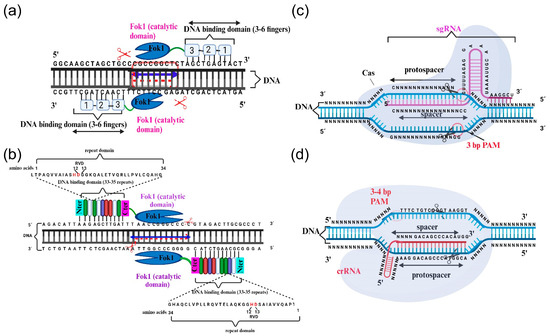
Figure 1.
Overview of the gene-editing systems analyzed in this review. (a) ZFNs, (b) TALENs, (c) CRISPR/Cas9, and (d) CRISPR/Cas12a. The main text provides a thorough description of all the elements depicted in the different panels. Created with BioRender.com (accessed on 12 January 2024).
The editing efficiency of the ZFN system is generally poor, with a success rate ranging from only 1% to 10% [9,10,11,12,13]. This limitation could be due to potential interactions between the neighboring C2H2 domains in the ZFNs, which affect their DNA binding specificity, although the underlying mechanism remains unclear [14]. Due to their low specificity, ZFNs can cause unintended mutations at off-target sites, as evidenced by earlier studies [8]. The first plant to be edited using the ZFN system was the model plant Arabidopsis thaliana [15], followed by maize [9], tobacco [11,16,17], and soybean [18]; however, the scope of crop editing using ZFNs has been limited due to the potential drawbacks of this approach [19,20,21].
2.2. Transcriptional Activator-like Effector Nucleases (TALENs)
TALENs are also chimeric proteins that can be engineered to cleave particular DNA sequences. Similar to ZFNs, TALENs comprises the C-terminal domain of FokI endonuclease and specific DNA binding domains, originating from the transcription activator-like effectors (TALE) from plant-pathogenic bacteria [22]. The TALE domain consists of numerous (between 12 and 27) consecutive repeats of 33–35 amino acids, each containing two α-helices and a short repeat variable diresidue known as RVD. RVD is essential for making sequence-specific DNA contacts. By using both bioinformatics tools and empirical data, the association between the RVD amino acid sequence and its binding to a specific nucleotide in the DNA was established [23,24]. Shortly afterwards, newly engineered TALE genes were utilized to selectively upregulate endogenous genes in various plant species [25]. The assembly of repeat modules for different RVD arrays allows for the generation of sequence-specific DNA binding domains to whatever length is desired. Like ZFNs, two monomeric TALENs are necessary to bind the target sites in the DNA and enable FokI to dimerize and cut the DNA (Figure 1b). Arabidopsis thaliana [26] and rice [27] were the first plant species to be edited using TALENs. To date, over 50 genes in 12 plant species, such as maize, wheat, barley, tobacco, soybean, potato, and tomato, among others, have undergone successful gene editing through the use of TALENs [28].
When considered in comparison to ZFNs, the interactions between DNA-binding domains of TALENs and their target nucleotides are simpler than those between ZFNs and their target trinucleotides. This means TALEN design is generally less complex than that of ZFNs. Additionally, TALENs offer other benefits over ZFNs, including higher editing efficiency (approximately 30%), and lower off-target mutations [28]. However, a clear disadvantage of TALENs is their significantly larger size when compared with ZFNs, which can pose challenges for their delivery and expression.
2.3. Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) and CRISPR-Associated Protein (Cas)
CRISPR and its associated locus encoding Cas proteins were initially discovered in the genomes of bacteria [29] and archaea [30]. The CRISPR/Cas system plays a crucial role in the adaptive immune responses against lytic bacteriophages and plasmids [31,32,33]. Upon initial infection by a phage, fragments of its DNA integrate into the CRISPR genomic locus. Upon new viral infection, the CRISPR locus is transcribed into the CRISPR RNA (crRNA), consisting of spacers with distinct sequences from various phages, indicative of past infections. These spacers are separated by repetitive endogenous sequences, known as direct repeats [34]. The Cas proteins possess nuclease and helicase domains that facilitate identifying and cutting exogenous DNA by following a crRNA-guided mechanism. Further additional processing requires trans-activating CRISPR RNA (tracrRNA) [35]. The tracrRNA–crRNA–Cas ribonuclease complex can recognize and cleave any exogenous DNA molecule that shares homology with the crRNA (Figure 1c). Another key element necessary for Cas cleavage activity is the so-called protospacer-adjacent motif (PAM) sequence, a conserved sequence of 2–6 base pairs flanking the target DNA sequence. Consequently, a sequence with perfect complementarity to the crRNA guide but lacking a PAM will be ignored by the Cas nuclease [36].
Soon after the molecular mechanism was uncovered, several research groups were able to edit genes in different organisms using the CRISPR/Cas system with great success [37,38,39,40,41]. A major step forward was the creation of a chimeric molecule called single guide RNA (sgRNA), which is a fusion of crRNA and tracrRNA and contains a short sequence complementary to the endogenous sequence of the gene to be edited. By varying the sgRNA sequence, it becomes possible to target different regions of DNA. Different CRISPR/Cas systems have been thoroughly investigated [42]. Two major classes, namely class 1 and class 2, comprise multi-subunit effector complexes or single-protein effector complexes, respectively, which are further divided into types (I, III, IV and II, V, VI, respectively) and subtypes [42,43]. Of the various Cas proteins identified, the most frequently used for gene editing are type II-A Cas9 from Streptococcus pyogenes [42] and type V-A Cas12a (Cpf1) from Acidaminococcus sp. And Lachnospiraceae bacterium [44].
2.3.1. Cas9
The most common method of gene editing utilizes the Cas9 protein of Streptococcus pyogenes. The Cas9 protein is a dual RNA-guided DNA endonuclease able to cleave the target DNA in both strands through recognizing the hairpin loop architecture of the crRNA–tracrRNA complex [45]. Cas9 guided by multiple sgRNAs can bind to endogenous genes with high specificity. In 2013, the first CRISPR/Cas9 edited plants of Arabidopsis thaliana, Nicotiana benthamiana, and Oryza sativa were obtained [46,47]. Different versions of Cas9 with high efficiency for gene editing have been identified and utilized in plants since then [48].
When compared to other gene-editing approaches, the CRISPR/Cas9 system exhibits high rates of editing efficiency, approximately 75% to 85% [49,50,51,52], surpassing those of ZFNs and TALENs [53]. However, the CRISPR/Cas9 system for gene editing has several drawbacks, including reduced editing efficiency due to PAM sequence mismatches, genetic mosaicism, and editing of non-targeted regions of the genome (i.e., off-target gene editing). To address these issues, a mutant version of Cas9 called Cas9 nickase (nCas9) has been engineered to generate single-strand DNA breaks and reduce off-target effects [54].
2.3.2. Cas12
Generally, Cas12a and their orthologs are smaller than Cas9 as they possess the nuclease domain but lack the helicase domain (Figure 1d). Certain Cas12a proteins serve as single effectors, which induce DSBs on the target DNA and are guided by smaller crRNAs (~42 nt) in contrast to those of Cas9 crRNAs (~100 nt) [44,55]. Cas12a has the ability to process pre-crRNA into mature crRNA independently of the tracrRNA. Additionally, there are other unique features of Cas12a proteins that distinguish them from Cas9. These include their preference for thymidine-rich PAM, whereas Cas9 prefers guanidine-rich sequences. Furthermore, after Cas12a cleaves the target DNA, it produces a 5′ overhang, increasing the efficiency of precise gene editing in AT-rich target regions, which Cas9 has difficulty accessing [44]. Beyond that, while Cas9 and Cas12a exhibit similar mismatches in their target DNA when assessed in vitro [56], it has been observed that Cas12a displays a lower off-target effect than Cas9 when conducting gene editing [57,58]. Initial successful outcomes following gene editing in plants using Cpf1, which is the Cas12a ortholog from Francisella novicida, were achieved in 2016 for Oryza sativa and Nicotiana benthamiana [59].
2.3.3. Other Cas Proteins
Cas13a is an RNA-guided RNA endonuclease from the bacterium Leptotrichia shahii that does not cleave DNA, but only single-stranded RNA [60]. Its use has been proposed to engineer resistance against plant RNA viruses and regulate gene expression post-transcriptionally [61]. Additionally, a few recent studies demonstrated that the hypercompact CRISPR/Cas gene-editing system of Cas12j encoded in some phage genomes can be used for efficient gene editing in plants with expanded target recognition capabilities compared to other Cas proteins [62,63,64].
2.4. Other Gene-Editing Tools
Several other gene-editing tools have been developed utilizing Cas9’s ability to bind to specific DNA sequences via their sgRNAs. These methods all employ non-functional variants of the Cas9 nuclease, also referred to as dead Cas9 (dCas9). dCas9 binds to the sgRNA target site on the DNA but does not cause DSBs, instead interfering with downstream transcription [65]. The fusion of specific deaminases with dCas9 enables the engineering of base editors, which direct the introduction of point mutations in specific sequences of a target DNA without initiating DSBs [66]. There are two classes of DNA base editors, namely cytosine base editors (CBEs) and adenine base editors (ABEs). ABEs convert the A=T base pair into G≡C, whereas CBEs convert G≡C into the A=T base pair [48,67]. Base editing has been successfully applied to a range of agronomic traits in crop plants, including wheat, rice, and tomatoes [68,69]. In prime editing, the dCas9 is fused with a reverse transcriptase (RT) enzyme, allowing for the introduction of base changes at the targeted site via the RT activity and using a prime editing guide RNA (pegRNA). This pegRNA not only specifies the target site but also encodes the desired edit [70]. Efficient cases of prime editing were previously reported for monocots, including rice, wheat, and maize, followed by mosses and dicots [71].
The different gene-editing methods described in this section allow for the precise modification of specific regions of the plant genome. CRISPR/Cas systems are the most widely used gene-editing tool due to their simple design, low cost, high efficiency, good repeatability, and short cycle time. Nevertheless, it is important to consider the advantages and disadvantages of ZFNs, TALENs, and CRISPR/Cas systems for gene editing to improve crop performance (Table 1).

Table 1.
Advantages and disadvantages of gene-editing systems in plants.
3. Methods for Obtaining Gene-Edited Plants
3.1. Methods for CRISPR/Cas Delivery
In order to induce DSBs and activate the endogenous DNA repair mechanism, which ultimately leads to gene editing, the Cas protein and sgRNA(s) components of the CRISPR/Cas system must reach the nucleus of the plant cell. This can be achieved through conventional delivery of genetically encoded CRISPR/Cas components or through preassembled gRNA/Cas ribonucleoprotein (RNP) complexes. The RNP complexes target and cleave the target sites in the DNA immediately after delivery and are rapidly degraded in those cells. Several recent systematic reviews have described the different methods used to deliver the CRISPR/Cas system to plants [72,73,74,75], and we will not provide an exhaustive description of them here. Below, you will find a brief overview of these methods, with an emphasis on recent developments for CRISPR/Cas delivery in tomatoes.
3.1.1. Particle Bombardment
The biolistic method delivers molecules into plant nuclei using accelerated nano-sized particles coated with nucleic acids or RNP complexes (Figure 2a) [76]. However, there are several disadvantages to this approach, including the random integration of the cargo DNA at multiple sites in the genome and the high cost of the equipment and reagents. Regenerating whole plantlets from transformed explants is a time-consuming process that is also dependent on the species and the genotype. RNP-mediated gene editing using particle bombardment has been successfully achieved in several species, but it has rarely been reported in tomatoes [77].
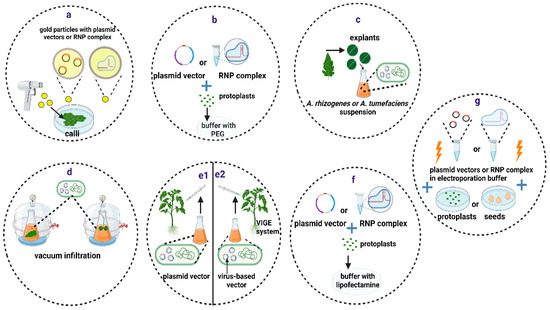
Figure 2.
Methods for delivery of CRISPR/Cas components into plant cells and tissues. (a) Particle bombardment. (b) PEG-mediated transfection. (c–e) Biological methods, including Agrobacterium-mediated stable transformation (c), vacuum infiltration (d), and direct injection of the Agrobacterium culture with an integrative plasmid vector (e1) or a virus-based plasmid vector (e2). (f,g) Other methods, including liposome-mediated transfection (f) and electroporation (g). Created with BioRender.com (accessed on 12 January 2024).
3.1.2. Polyethylene Glycol (PEG)-Mediated Transfection
In the presence of divalent cations, such as Ca2+ or Mg2+, and at high pH, PEG facilitates the incorporation of exogenous DNA or RNP complexes into plant protoplasts, but with low transformation efficiency (Figure 2b). This approach relies on the establishment of successful protoplast isolation and regeneration procedures. PEG-mediated transfection has been widely used to deliver vector DNA or RNPs into protoplasts of many plant species, including various tomato cultivars [78,79,80,81] and the wild tomato S. peruvianum [82].
3.1.3. Biological Methods
Biological methods for delivering DNA into plant nuclei rely on the natural ability of plant-pathogenic soil bacteria of the genus Rhizobium to integrate some of the genes present in their T-DNA into the genomic DNA of plant cells [83,84]. A. tumefaciens, also known as Rhizobium radiobacter, is the most commonly used species for delivering vectors encoding the CRISPR/Cas components. Conversely, A. rhizogenes, also known as R. rhizogenes, has received considerable attention in recent years during studies of root-specific processes [85]. A large number of laboratory strains of A. tumefaciens are available to transform a wide variety of plant species with varying success [86]. Recently, the A. tumefaciens and A. rhizogenes genomes were modified to improve their transformation efficiency and broaden their host range of plant species [87,88].
Agrobacterium spp. cell cultures containing the desired vectors in the exponential growth phase are used to transform plant tissues (Figure 2c). For plant species such as A. thaliana, the floral dip method [89] is preferred, while in most other cases, A. tumefaciens must be co-cultured with tissue explants, such as cotyledons, leaves, calli, or embryos. In such cases, a suitable protocol for regenerating whole plants from the few transformed cells with the T-DNA from A. tumefaciens is required. In the case of tomatoes, cotyledons can be easily transformed through co-culturing with various Agrobacterium spp. strains. Shoot induction can then be produced shortly afterwards with appropriate hormone combinations [90].
Transient expression of CRISPR/Cas components can be induced in plant tissues by delivering A. tumefaciens into plant cells through direct injection or vacuum infiltration (Figure 2d,e1) [91]. This method is appropriate for evaluating the gene-editing efficiency of different vectors in tomato leaves before conducting stable transformations [92,93]. Zhang and coauthors (2020) tested 195 sgRNAs for their ability to cause mutations in tomato leaves, and they found that 61.5% of them were suitable for gene editing [92].
Virus-induced gene editing (VIGE) relies on the use of viral-derived vector systems for the delivery of CRISPR/Cas components into plant cells [94]. The VIGE approach does not require genome integration of the CRISPR/Cas backbone, making it less time-consuming and more broadly applicable. However, the primary limitation of this approach is that most viruses are unable to infect meristematic or germline cells. As a result, regenerated edited plants are normally derived from other somatic cells [94]. Nonetheless, various approaches based on vectors derived from geminivirus [95] and potato virus X [96] have been successfully employed to produce stable edited tomato plants using direct injection (Figure 2e2). The results suggest that the VIGE approach is a reliable and adaptable technology that can be used for precise breeding of tomato traits.
3.1.4. Other Methods of CRISPR/Cas Delivery
Liposomes are lipid-based nanoparticles that have been shown to offer a useful tool for the delivery of genes and proteins (Figure 2f) into mammalian cells [97] and that have been used to deliver donor DNA and support subsequent successful gene editing in citrus protoplasts [98]. In addition, RNPs have been delivered in tobacco protoplasts through liposome nanoparticles, achieving editing efficiencies as high as 6.0% [99]. Moreover, recently, a similar protocol has been devised for the successful delivery of RNPs in maize protoplasts [100].
Electroporation, meanwhile, is a technique that increases the permeability of cell membranes in plant cells without cell walls (protoplasts) by exposing them to short and intense electric pulses (Figure 2g). This allows for the incorporation of exogenous nucleic acids or RNPs [101]. Recently, researchers have demonstrated the delivery of RNPs into cabbage and soybean protoplasts via electroporation, achieving editing efficiencies as high as 3.4% and 8.1%, respectively [102,103]. Electroporation has also been used in tomatoes for the delivery and transient expression of vectors encoding ZFNs [104], but no other reports with CRISPR/Cas vectors of RNPs can be found in this species so far.
3.2. Marker Genes Used with the CRISPR/Cas System
When using conventional CRISPR/Cas vectors, most gene-edited plants include the stable integration of the CRISPR/Cas system into the plant genome. Furthermore, the CRISPR/Cas expression module typically includes genes encoding selection markers for antibiotic or herbicide resistance. However, the presence of the transgene does not necessarily indicate that gene editing has taken place. Characterizing the mutations that have occurred in the target DNA often requires laborious procedures, including high-resolution melting PCR or Sanger sequencing. Additionally, to conform with biosafety regulations concerning genetically modified organisms (GMOs), the CRISPR/Cas expression module must be segregated in the progeny of gene-edited plants through self-pollination or crossing. Alternatively, the delivery of preassembled CRISPR/Cas components is considered a transgene-free method of gene editing. In this approach, the RNP complexes have high specificity for their target DNAs and are rapidly degraded within the cell after dissociating from the genomic DNA. The fusion of Cas proteins with fluorescent proteins, such as enhanced GFP (EGFP), enables the direct visualization of the RNP complexes in plant nuclei, which is highly convenient during the optimization of the delivery process.
For both approaches, it is beneficial to select a CRISPR/Cas-induced mutation that causes a visible phenotype. A frequently used endogenous target gene is the PHYTOENE DESATURASE (PDS) gene Solyc03g123760, which is involved in carotenoid biosynthesis. Mutations in this gene impair chlorophyll accumulation and produce an albino phenotype in many species, including tomatoes [105]. However, mutations in PDS affect the survival of the edited explants, which hinders the effective plant regeneration required for successive characterization. Recently, Rinne et al. (2021) proposed using a different endogenous marker gene to evaluate CRISPR/Cas gene-editing activity [106]. The MULTIPLE ANTIBIOTIC RESISTANCE 1 (MAR1) gene Solyc01g100610 encodes a transporter located in the mitochondria and chloroplasts, which plays a role in maintaining iron homeostasis and facilitates the transport of aminoglycoside antibiotics into these organelles. Mutations induced by the CRISPR/Cas system in MAR1 confer kanamycin resistance in tomatoes [106].
4. Gene Editing in Tomato Breeding
4.1. ZFNs
Only a limited number of studies are available on the application of ZFNs to edit tomato plants [107,108]. Shukla et al. (2016) conducted an experiment where they utilized ZFNs to target Solyc07g062650, which encodes a mitochondrial malate dehydrogenase (mMDH), in both the M82 and Moneymaker tomato cultivars [107]. PEG-mediated transfection was used to deliver four distinct ZFN constructs into protoplasts, and the regenerated plantlets were obtained via indirect organogenesis. The editing efficiency of mMDH was found to be low, ranging from 0.7% to 5.5%. The majority of mutants were found to have small deletions varying from 1 to 22 bp, while one mutant showed an insertion of 2 bp. Regenerated mutants displayed different phenotypes: heterozygous plants for any mutation that led to disruption of the mMDH open reading frame showed a decrease in growth and fruit yield, while homozygous plants for specific mutations exhibited an increase in fruit yield [107]. In another study, ZFNs were used to target the LEAFY-COTYLEDON1-LIKE4 (L1L4) gene Solyc05g005350, which encodes for the β-subunit of a nuclear factor Y, in the Heinz 1706 cultivar [108]. Electroporation of germinated seeds facilitated the transient expression of constructs encoding a pair of ZFNs designed to target exons 1 and 2 of the L1L4 gene. Based on the phenotypes observed in the edited plants from the T0 generation, over 65% efficiency was reported. Several mutations in the coding sequence of L1L4 have led to variations in plant traits, including seedling vigor, plant height, number of florets, and flowering and ripening times, thereby demonstrating that the affected L1L4 gene is a useful target for crop improvement [108].
4.2. TALENs
Three reports have been published to date on the use of TALENs for tomato gene editing. Lor et al. (2014) designed two pair of TALENs, under the control of an estrogen-inducible promoter, to target the PROCERA (PRO) gene in tomatoes, Solyc11g011260 [109]. This gene encodes a negative regulator of gibberellin signaling. Upon TALEN expression, 7 out of 40 regenerated plantlets carried deletions ranging from 1 to 88 bp, along with a 39-bp insertion and a 4-bp deletion, resulting in frameshifts that would lead to the production of truncated PRO proteins. Homozygous pro plants displayed phenotypes indicating an increased gibberellin response [109]. In another report, geminivirus replicons were used to express TALENs designed to target the ANTHOCYANIN 1 (ANT1) gene Solyc10g086260, encoding an MYB transcription factor whose overexpression results in purple plant tissue due to anthocyanin accumulation. The gene-editing efficiency of this system was about 14% [95]. In a recent report, a known mutation in the pepper gene that encodes EUKARYOTIC TRANSLATION INITIATION FACTOR 4E (eIF4E), Solyc03g005870, which is reported to be associated with potyvirus resistance, was generated through a gene knock-in strategy with a pair of TALENs in tomato [110]. The TALEN vector and the donor DNA template were introduced to cherry tomato leaves in a biolistic approach. One out of thirty-two regenerating shoots incorporated the donor template through NHEJ, and the edited plants with this allele showed the broadest potyvirus resistance spectrum achieved through genetic engineering in tomatoes so far [110].
4.3. CRISPR/Cas9
The CRISPR/Cas9 approach was first applied to tomato gene editing in 2014 [111,112]. As a proof of principle, Brooks et al. (2014) designed two sgRNAs to target the tomato homolog of the ARGONAUTE7 (AGO7) gene Solyc01g010970 [111], since loss-of-function mutations of AGO7 are known to produce filiform leaves [113]. The plasmid vector was delivered via Agrobacterium tumefaciens transformation of M82 cotyledons, and 14 out of 29 T0 plants displayed the wiry leaf phenotype characteristic of known ago7 alleles [111]. Ron et al. (2014), meanwhile, used A. rhizogenes-mediated transformation of cotyledon explants as a delivery method for the CRISPR/Cas9 vector, which was designed to target both a reporter gene (mGFP5) and the Solyc02g092370 gene encoding a SHORT-ROOT (SHR) protein in tomato hairy roots [112]. Consistent with the notion of a conserved role of SHR in both tomatoes and A. thaliana, transgenic hairy roots with no GFP expression and a reduced root meristem were identified. These roots contained small insertions and deletions in the coding region of the SHR gene [112].
In the past decade, the use of CRISPR/Cas9 for gene editing in tomatoes has been implemented in dozens of tomato cultivars [114,115], including wild species such as S. lycopersicum var. cerasiforme [110], S. pimpinellifolium [116,117,118,119,120,121], and S. peruvianum [82]. When searching the scientific literature available as of 31 December 2023, we identified 356 primary references that used CRISPR/Cas technology for gene editing in tomato and related species (Figure 3a–c and Supplementary Table S1). From this list, we selected a subset of references to be further discussed (Supplementary Table S2). A Sankey diagram constructed from the evidence presented in these selected references illustrates the relationship between tomato cultivars, their wild relatives, Cas enzyme types, A. tumefaciens strains, and the key genetic traits influenced by the genetic modifications induced by CRISPR/Cas9 (Figure 3d).
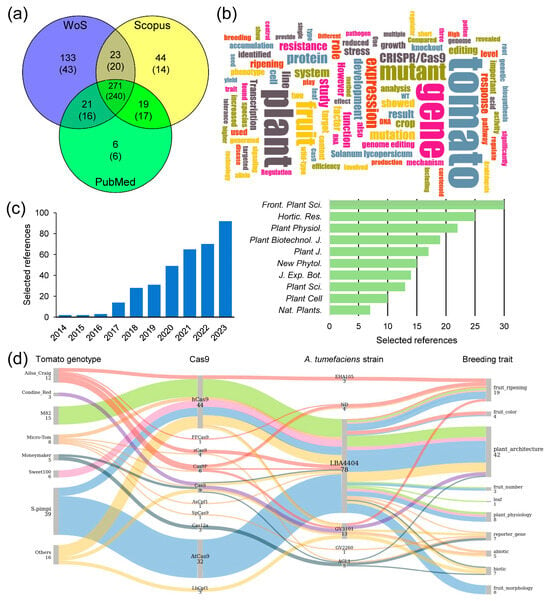
Figure 3.
Results of a reference search and text mining analysis. (a) Venn diagram of scientific articles downloaded from public databases using the search term: “(Cas9 tomato) NOT (review)” and restricted by date (from 1 January 2014 to 31 December 2023). Numbers in brackets indicate selected references after manual curation. (b) A word cloud generated with the 356 unique abstracts selected based on the criterion of including any mutant generated by gene editing in tomato and related species in the main text. The image was generated using the web application located at https://wordclouds.ethz.ch/ (accessed on 12 January 2024). (c) Selected papers about gene editing in tomato and related species, along with the top 10 plant science journals in which they were published. The raw data utilized for preparing panels a–c can be found in Supplementary Table S1. (d) The Sankey diagram illustrates the relationship between tomato cultivars, types of Cas enzyme, A. tumefaciens strains, and the key breeding traits it influences. The diagram was generated with SankeyMATIC and is based on data from Supplementary Table S2. The thickness of the lines and the colors represents the proportion of evidence connecting the different categories. Spimpi: S. pimpinellifolium genotypes; ND: not included.
Several studies have documented the domestication process from the most likely progenitor, S. pimpinellifolium, to modern tomato varieties [122]. Loss-of-function mutations in six loci that are important for key domestication traits have been identified from these and other studies: plant architecture (SELF PRUNING [SP] Solyc06g074350), fruit shape (OVATE [O] Solyc02g085500), fruit size (FASCIATED [FAS] Solyc11g071380, and FRUIT WEIGHT 2.2 [FW2.2] Solyc02g090730), fruit number (MULTIFLORA [MULT] Solyc02g077390), and nutritional quality (LYCOPENE BETA CYCLASE [CycB] Solyc04g040190). To reconstruct the domestication of tomatoes from S. pimpinellifolium, Zsögon et al. (2018) [121] designed a CRISPR/Cas9 vector containing six sgRNAs that targeted specific sequences in the coding regions of these six genes. Four of the six targeted loci were successfully edited in all 50 T1 lines tested, resulting in indel mutations in SP, O, FW2.2, and CycB [121]. In a subsequent round of gene editing, the researchers incorporated CLAVATA 3 (CLV3) Solyc11g071380, and developed a new CRISPR/Cas9 vector with eight sgRNAs targeting CLV3, FW2.2, MULT, and CycB genes. This resulted in 28 T1 lines with loss-of-function mutations in all four targeted loci [121]. The engineered lines exhibit a threefold increase in fruit size and a tenfold increase in fruit number when compared with the wild parent, S. pimpinellifolium. A significant finding was that the accumulation of fruit lycopene in these lines was enhanced fivefold in comparison to the cultivated tomato S. lycopersicum [121]. Similarly, Li et al. (2018) developed a multiplex CRISPR/Cas9 strategy to edit five genes in S. pimpinellifolium related to day-length sensitivity (SELF PRUNING 5G [SP5G] Solyc05g053850), shoot architecture (SP), flower and fruit production (CLV3 and WUSCHEL [WUS] Solyc02g083950), and vitamin content (GGP1 Solyc02g091510) [119]. Furthermore, Lemmon et al. (2018) used CRISPR/Cas9 to mutate orthologs of tomato domestication genes that control plant architecture (SP), flower production (SP5G), fruit size (CLAVATA1), and fruit abscission (JOINTLESS-2) in the orphan crop Physalis pruinosa [123]. This wild Solanaceae is distantly related to tomato and is grown in Central and South America for its subtly sweet berries. Overall, the results suggested that CRISPR/Cas9-mediated gene editing could be used to accelerate domestication in wild species and create crops that are better suited to changing climate scenarios [124,125].
4.3.1. CRISPR/Cas9-Edited Genes Related to Fruit Characteristics
Tomato plants are grown for their fleshy berry fruits, which come in varying sizes, shapes, and colors [126,127]. Tomatoes contain several health-promoting components, including vitamins, carotenoids, and phenolic compounds [128]. Tomato breeders have long utilized the limited genetic variation present in this species to enhance fruit quality traits. Some of the genes responsible for variations in fruit weight, shape, and biochemical composition have been identified and those might be used for precise gene editing using the CRISPR/Cas9 system (Supplementary Table S2), as will be exemplified below.
Tomato fruit ripening is a complex physiological and biochemical process that involves the degradation of chlorophyll, the accumulation of pigments (mainly carotenoids), the softening of the pulp, and the alteration of its organoleptic qualities. This process is controlled by ethylene, and requires three transcription factors: the MADS-box RIPENING INHIBITOR (RIN), the SBP-box COLORLESS NON-RIPENING (CNR), and the NAC transcription factor NON-RIPENING (NOR) [129]. One of the initial reports on the application of CRISPR/Cas9 technology in tomatoes targeted the RIN gene (Solyc05g012020) [130,131]. Fruits from RIN-knockout plants exhibited partial ripening and lower levels of lycopene compared to the wild type [131]. However, unexpectedly, these fruits showed excessive cell wall degradation. Furthermore, CRISPR/Cas9-induced mutations in CNR (Solyc02g077920) and NOR (Solyc10g006880) did not abolish ripening [132,133], indicating that the ripening transcriptional regulatory network is highly robust. These results are in stark contrast with those found in “classical” rin, Cnr, and nor mutants, which have obvious ripening-inhibited phenotypes. The difference in findings may be explained by the recent proposal that some of these mutants have gain-of-function mutations [134].
The color change that occurs during fruit ripening is due to the accumulation of β-carotene (orange) and lycopene (red), as well as a decrease in β-xanthophyll (yellow) and chlorophyll (green) levels [135]. Furthermore, the expression of SNAC4 (Solyc07g063420) and SNAC9 (Solyc04g005610) genes is induced by ethylene, and alterations in the pigmentation of mature fruits are observed when the expression levels of these genes are reduced by VIGS, indicating a positive role of both genes in this process [136]. In previous research, the SNAC9 gene was knocked out thoroughly using CRISPR/Cas9, resulting in an absence of expression of the protein. This led to a significant reduction in lycopene and total carotenoid contents in the mutants, possibly due to the direct regulation of key genes involved in carotenoid biosynthesis, such as PHYTOENE SYNTHASE 1 (PSY1) Solyc03g031860 [137]. A strategy using NGTs has recently been implemented to rapidly generate tomato lines with fruits of varying colors [135,138,139]. By genetically editing three genes related to fruit color, PSY1, MYB12 (Solyc01g079620), and STAY-GREEN 1 (Solyc08g080090), using a multiple CRISPR/Cas9 approach, it was possible to obtain transgene-free plants with fruits of different colors in less than a year. This strategy does not affect other important agronomic traits, such as yield and fruit quality [138].
Initial attempts to extend the shelf life of tomato fruits using CRISPR/Cas9 were focused on genes that encode enzymes responsible for degrading cell-wall pectins, such as pectate lyase (PL), polygalacturonase, and β-galactanase. However, only mutations in the PL gene Solyc03g111690, resulted in firmer fruits [140]. Simultaneous knockout of two genes, one encoding a fruit ripening-associated α-expansin (Solyc06g051800) and the other an endoglucanase (Solyc09g010210), using CRISPR/Cas9 improved fruit firmness [141]. Conversely, simultaneous overexpression of these genes promoted early fruit softening. These results support the conclusion that these two genes have a synergistic effect on fruit softening [141]. Furthermore, the bHLH transcription factor BRI1-EMS-SUPPRESSOR1 (BES1) encoded by Solyc04g079980 has been identified as an upstream regulator of fruit softening through the activation of cell-wall degradation during ripening [142]. CRISPR/Cas9-generated loss-of-function BES1 mutants resulted in firmer fruits and a longer shelf life during postharvest storage, without any negative impact on visual or nutritional quality.
4.3.2. CRISPR/Cas9-Edited Genes Related to Plant Architecture Traits
Modifying the plant architecture can significantly impact crop yield. This was demonstrated through mutations to the biosynthesis or signaling pathways of gibberellins, which increased crop yield by decreasing plant height in wheat and rice, an outcome that is of value during the “Green Revolution” [143]. Furthermore, in rice, the IDEAL PLANT ARCHITECTURE 1 (IPA1) gene has been determined to be a novel master regulator of the plant architecture, which can be used as a target during molecular design to improve grain yield. IPA1 encodes an SBP-box-like (SPL) transcription factor downstream of strigolactone signaling [144]. One of the IPA1 orthologs of tomatoes, SPL13 Solyc05g015840, has been shown to act downstream of strigolactones to suppress lateral bud growth by inhibiting cytokinin biosynthesis. The knockout lines of SPL13 generated with CRISPR/Cas9 displayed an increased shoot branching phenotype [133]. In other work, Kwon et al. (2020) developed a gene-editing staking approach to transform vine-like tomato plants into compact, early-yielding plants suitable for indoor agriculture [145,146]. Elsewhere, mutations in the classical flowering repressor gene SP, both natural and CRISPR/Cas9-induced, conferred a determinate growth habit in tomatoes [121]. Furthermore, mutating its paralog SP5G with CRISPR/Cas9 in the sp mutant background resulted in a shorter time to flowering and a more compact, determinate growth habit. This led to a quick burst of flower production and an early yield [147]. Meanwhile, cloning the short internodes dwarf mutant in the M82 cultivar was found to affect the well-known ERECTA (ER) gene [145]. In A. thaliana, this gene is known to regulate internode length [148]. The specific ER gene Solyc08g061560 was targeted using CRISPR/Cas9 in the previously generated sp sp5g double mutant [147]. The resulting plants exhibited a triple-determinate form, and when grown in a self-contained, climate-controlled LED hydroponic vertical farm system, they produced higher yields due to their bushy shoot architecture, rapid cycling, and highly compact fruit clusters [145].
Naturally occurring mutations in the MADS-box transcription factor genes JOINTLESS (J) and JOINTLESS2 (J2) in tomatoes have the potential to improve harvestability by modifying flower development, as they suppress the abscission zone in fruit peduncles [149]. The jointless pedicel trait has been successfully introgressed into small-fruited processed tomatoes and fresh-market tomatoes. However, the j2 loss-of-function mutation can lead to undesirable branching of inflorescences in genetic backgrounds that also carry a cryptic variant for the close homolog ENHANCER OF J2 (EJ2) Solyc03g114840, which was selected during domestication. This combination of j2 ej2 loss-of-function alleles results in excessive flower production and low fertility due to a poor fruit set [150]. Deletions induced by CRISPR/Cas9 in J2 Solyc12g038510, result in the jointless phenotype. When combined with CRISPR/Cas9-induced null mutations in EJ2, this leads to strongly branched inflorescence in tomato breeding lines carrying the suppressor of branching 3 (sb3) quantitative trait loci (QTL). These results suggest that genotypes carrying sb3 could be utilized to maintain a normal inflorescence architecture when generating jointless phenotypes via gene editing [150].
During the process of tomato domestication, natural genetic variants were selected based on their alterations in the expression of key genes involved in target agronomic traits, such as yield or plant architecture. In many cases, the molecular characterization of these genetic variants has shown that they do not affect the coding region of the gene, but rather its cis-regulatory regions, either in gene promoters or in regulatory introns. Structural variations, such as large indels, duplications, and chromosomal rearrangements, play a crucial role in plant evolution and agriculture. They impact traits such as shoot architecture, flowering time, fruit size, and stress resistance [151]. Gene editing may be used to study the effects of variations in regulatory regions by recreating specific mutations or mimicking the expression effects of natural cis-regulators, which could be achieved by using the CRISPR/Cas approach on defined tomato genotypes. Accordingly, Rodríguez-Leal et al. (2017) developed a genetic screen that uses heterozygous loss-of-function mutant backgrounds to efficiently assess the phenotypic effects of multiple CRISPR/Cas9-induced promoter variants for known genes that regulate three key productivity traits in tomato: fruit size (CLV3, WUS), inflorescence branching (MULT), and plant architecture (SP) [117]. The promoter alleles for CLV3 exhibited a full range of quantitative variation. The edited plants displayed phenotypes ranging from moderate to strong, capturing the full spectrum of allelic diversity and locule number variation previously identified [117]. This approach has been demonstrated to generate multiple regulatory mutations for the systematic assessment of the association between cis-regulatory regions and phenotypic variation. Furthermore, there is also potential for engineering gain-of-function alleles and thereby establishing a basis for dissecting the complex relationships between gene-regulatory alterations, which may facilitate quantitative trait control.
4.3.3. CRISPR/Cas9-Edited Genes Related to Adaptive Stress Responses
Systematic reviews have compiled extant studies on gene editing with CRISPR/Cas9 in agricultural crops in the scientific literature [152,153,154]. Various biotic stresses, such as diseases and pests, as well as abiotic stresses such as cold, heat, drought, and salinity, affect tomato crop production, and Supplementary Table S2 shows that CRISPR/Cas9 gene editing has been used to address many of these traits. Those applications will be summarized in this section.
The MILDEW RESISTANCE LOCUS O (MLO) encodes an integral transmembrane protein that is highly conserved in monocots and dicots. It is a key factor in susceptibility to powdery mildew caused by the pathogenic fungus Oidium neolycopersici. In cultivated tomato and the wild species S. peruvianum, CRISPR/Cas9-induced knock-out lines of MLO1 orthologs Solyc04g049090 and pSolyc04g049090 conferred resistance to O. neolycopersici without generating any other undesired phenotypic effects [82,155,156]. Furthermore, the DOWNY MILDEW RESISTANT 6 (DMR6) gene in A. thaliana encodes a putative 2-oxoglutarate Fe(II)-dependent dioxygenase that has been identified as a factor in susceptibility to bacterial and oomycete pathogens. The tomato genome contains two orthologs of DMR6, DMR6-1 (Solyc03g080190) and DMR6-2 (Solyc06g073080). Inactivation of DMR6-1 using CRISPR/Cas9 resulted in enhanced disease resistance against different tomato pathogens, such as bacteria, oomycetes, and fungi, without an obvious growth penalty [157]. Notably, the enhanced pathogen resistance observed in the dmr6-1 tomato mutants was correlated with increased levels of salicylic acid. The biochemical characterization of DMR6 enzymes suggests that they play a role in converting salicylic acid to its inactive form [157]. In other work, the CRISPR/Cas9 system has been used to mutate the susceptibility gene POWDERY MILDEW RESISTANCE 4 (PMR4) Solyc07g053980, with the product functioning as a callose synthase conferring resistance to O. neolycopersici and Phytophtora infestans in tomatoes [158,159]. These results demonstrate that the CRISPR/Cas9 system is suitable for facilitating broad resistance to bacterial and fungal pathogens by precisely targeting susceptibility genes and negative regulators involved in the plant defense mechanism.
Tomato yellow leaf curl virus (TYLCV) is a highly destructive viral pathogen that affects tomato crops globally. TYLCV is transmitted by the whitefly Bemisia tabaci when it feeds on the phloem sap of plants. Previous strategies to confer resistance to TYLCV in tomato plants have focused on the viral coat protein and DNA replicase protein, CP and RepA, respectively. Efficient viral interference was achieved by targeting the TYLCV genome with CRISPR/Cas9 for these sequences, resulting in reduced TYLCV accumulation in transgenic tomato plants [160]. Several QTL related to TYLCV resistance in tomatoes, including Ty-1 to Ty-6, have been identified [161]. ty-5 confers broad-spectrum resistance and encodes a missense allele of the tomato homolog of the PELOTA messenger RNA surveillance factor, which is involved in ribosome recycling during protein translation. Gene editing of the PELOTA gene Solyc04g009810, using a CRISPR/Cas9 approach resulted in resistance to TYLCV as a result of restricting the proliferation of viral DNA, likely by slowing or inhibiting ribosome recycling and decreasing viral protein synthesis in infected cells [156]. Targeting susceptibility factors encoded by the host plant genome, rather than the viral genome, offers a promising approach to achieving resistance to pathogens without the need for stable inheritance of CRISPR/Cas9 components.
In the context of global warming, drought stress is becoming a critical challenge in tomato production. As previously discussed [162], the development of new CRISPR/Cas9-based approaches to improve drought tolerance in tomatoes is essential to reduce yield loss. Several key factors have been considered (Supplementary Table S2). Among those, in a scenario where the tomato LATERAL ORGAN BOUNDARIES DOMAIN 40 (LBD40) Solyc02g085910 was highly expressed in the roots and fruits and its expression was significantly induced by PEG and salt treatment, LBD40 knockout mutants generated by CRISPR/Cas9 gene editing improved the water-holding ability of tomatoes under drought conditions, suggesting that LBD40 was a negative regulator of drought tolerance in this species [163]. Furthermore, two studies have highlighted the role of AUXIN RESPONSE FACTOR 4 (ARF4) Solyc11g069190 in mediating the tolerance to salinity and osmotic stress in tomatoes [164,165]. The expression of the ARF4 gene was downregulated in tomato seedlings under ABA and water deficit conditions. Downregulating the expression of ARF4 with CRISPR/Cas9 was observed to enhance salt and osmotic stress tolerance recovery. This resulted in an obvious leaf curling phenotype, which reduced transpiration, and a significant increase in root length and density compared to wild type plants under stress conditions [164,165]. More recently, the expression of SP2G (Solyc02g079290) and SP3C (Solyc03g026050) was found to be significantly increased in the leaves of the drought-resistant wild species S. pennellii and in domesticated tomatoes after irrigation was stopped [166]. Furthermore, three independent CRISPR/Cas9-mediated SP3C homozygous mutants exhibited increased root length and reduced lateral root branching compared to the wild type. These traits are associated with greater drought tolerance in tomatoes and could be fine-tuned for agronomic gain [166]. Further research into the mechanisms of action of key transcription factors that regulate the abiotic stress response in tomatoes may expand the resources and tools at our disposal for developing multi-stress tolerant tomato varieties.
4.4. CRISPR/Cas12a
Two different Cas12a variants were used to stably transform tomato plants so they targeted Solyc01g079620, which encodes the MYB12 transcription factor required for flavonoid biosynthesis. A mutation in this gene leads to the production of pink-colored fruits [167]. The gene-editing efficiency of this system ranged from 7.7% to 48.8%, with a tendency to induce a broader range of deletions but no insertions, in contrast to Cas9 [168]. Furthermore, as demonstrated by Vu et al. (2020), the CRISPR/Cas12a system has the advantage of an increased efficiency in genome editing via the homology-directed repair (HDR) pathway when compared to Cas9. The authors reported an HDR efficiency of 4.5% of the ANT1 visible marker when using a geminiviral replicon system [169]. In another study, to confirm the applicability of the Cas12a-mediated HDR approach for tomato genome editing of a potential agronomic trait, a known mutation in the HIGH-AFFINITY K+ TRANSPORTER 1;2 (HKT1;2) gene Solyc07g014680, which determines salinity tolerance [170], was engineered. However, the efficiency of HDR was low, at only 0.7%. The edited plants were salt-tolerant in both homozygous and heterozygous states [169]. In other work, a higher HDR efficiency of 4.3% was reported when using a new Cas12a variant that is capable of performing the cleavage of the target DNA at low temperatures [171]. Together, these results confirm that Cas12a-mediated HDR induces efficient gene targeting, which may be used to obtain allele-specific traits and marker-free tomato plants. More recently, Cas12 has been fused to a CBE with the aim of conferring resistance to the herbicide chlorsulfuron via precise editing of the ACETOLACTATE SYNTHASE (ALS) gene Solyc03g044330 [172]. More than 20 chlorsulfuron-resistant lines were obtained in this way and the mutations were confirmed to be highly specific. In addition, the authors edited the Solyc08g061560 gene, which encodes the ER kinase receptor. Mutations in this gene in A. thaliana resulted in a compact inflorescence [148]. The phenotype of the homozygous edited tomato plants in this gene included a compact architecture, short petioles, and densely clustered inflorescence [172].
5. Conclusions
Over the last 15 years, significant progress has been made in crop gene editing using the NGTs of ZFNs, TALENs, and CRISPR/Cas. Among these, the CRISPR/Cas-based editing system has emerged as the preferred option due to its many benefits. The high efficiency of gene editing induced by the Cas proteins, together with the flexibility in the design of gene-specific RNA guides, allows the chosen experimental design to be refined while considering the genotype of the tomato variety, thus supporting the ultimate objective pursued with the genetic modification. A wealth of evidence supports the utility of the CRISPR/Cas system for the genetic editing of multiple genes in different varieties of cultivated tomatoes and related wild species. In this review, we conducted a systematic search of the references and identified 356 scientific articles that reported primary results on the modification of a gene using NGTs in cultivated tomatoes or related wild species. From these, we selected 47 genes that affect key genetic traits that we determine to warrant further discussion. Some of these genes were found to impact the plant architecture, resulting in increased shoot and flower branching, a more compact growth habit, and a shorter flowering time. Engineering such variations can thus facilitate the development of new tomato varieties that are better suited to indoor farming and have higher crop yields than those at present. Furthermore, we have discussed tomato plants that have been gene edited to increase their tolerance to various pathogens and viruses, including Oidium neolycopersici, Phytophtora infestans, and TYLCV. Additionally, gene-edited tomatoes are being developed to exhibit greater tolerance to drought or salt/osmotic stress.
The widespread application of CRISPR/Cas methodologies for precise genetic modification will facilitate the development of tomato plants that are more tolerant to the multiple stresses associated with the effects of human-driven climate change. These new varieties will not only be suitable for controlled greenhouse conditions but also for outdoor cultivation in order to maintain or even enhance the yield under adverse environmental conditions. On the other hand, the use of CRISPR/Cas could enable the design and production of new tomato varieties with better-quality fruits enriched in vitamins and other essential or bio-healthy compounds. However, in many countries, the regulatory management of crops generated using NGTs does not differ from that of crops produced using conventional genetic improvement techniques, such as mutagenesis, while in other countries, the regulation of NGT crops parallels that of GMOs. This has resulted in regulatory oversight, making it difficult to generalize scientific advances aimed at improving food security in developing countries.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms25052606/s1.
Author Contributions
Conceptualization, E.L. and J.M.P.-P.; methodology, O.Y.; formal analysis, E.L. and J.M.P.-P.; investigation, O.Y.; resources, J.M.P.-P.; data curation, O.Y. and E.L.; writing—original draft preparation, O.Y. and E.L.; writing—review and editing, J.M.P.-P.; visualization, J.M.P.-P.; supervision, E.L. and J.M.P.-P.; project administration, J.M.P.-P.; funding acquisition, J.M.P.-P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by MCIN/AEI/10.13039/501100011033, grant TED2021-132256B-C22, the Conselleria d’Innovació, Universitats, Ciència i Societat Digital, grant AGROALNEXT/2022/036, by the “ERDF A way of making Europe”, and by the “European Union NextGenerationEU/PRTR”. O.Y. is recipient of a PhD fellowship of the “Acoge CV-UCRANIA personal investigador” program.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data used for this review have been included in the Supplementary tables.
Acknowledgments
We are grateful to Taras Pasternak and other members of our laboratory for constructive discussions and critical suggestions.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Anwar, M.R.; Liu, D.L.; Macadam, I.; Kelly, G. Adapting Agriculture to Climate Change: A Review. Theor. Appl. Climatol. 2013, 113, 225–245. [Google Scholar] [CrossRef]
- Kumar, V.; Srivastava, A.K.; Sytar, O. Editorial: Plants for Future Climate: Responses and Adaptations to Combined, Multifactorial, and Sequential Stresses. Front. Plant Sci. 2023, 14, 1290649. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Study on the Status of New Genomic Techniques under Union Law and in Light of the Court of Justice Ruling in Case C-528/16. Available online: https://food.ec.europa.eu/document/download/5135278b-3098-4011-a286-a316209c01cd_en?filename=gmo_mod-bio_ngt_eu-study.pdf (accessed on 12 January 2024).
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 12 January 2024).
- Kumar, L.; Chhogyel, N.; Gopalakrishnan, T. Climate Change and Future of Agri-Food Production. In Future Foods; Elsevier: Amsterdam, The Netherlands, 2022; pp. 49–79. [Google Scholar]
- Miller, J.; McLachlan, A.D.; Klug, A. Repetitive Zinc-Binding Domains in the Protein Transcription Factor IIIA from Xenopus Oocytes. EMBO J. 1985, 4, 1609–1614. [Google Scholar] [CrossRef]
- Pattanayak, V.; Guilinger, J.P.; Liu, D.R. Determining the Specificities of TALENs, Cas9, and Other Genome-Editing Enzymes. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2014; pp. 47–78. [Google Scholar]
- Petolino, J.F. Genome Editing in Plants via Designed Zinc Finger Nucleases. Vitr. Cell. Dev. Biol.-Plant 2015, 51, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shukla, V.K.; Doyon, Y.; Miller, J.C. Precise Genome Modification in the Crop Species Zea Mays Using Zinc-Finger Nucleases. Nature 2009, 459, 437–441. [Google Scholar] [CrossRef] [PubMed]
- De Pater, S.; Neuteboom, L.W.; Pinas, J.E.; Hooykaas, P.J.J.; Van Der Zaal, B.J. ZFN-induced Mutagenesis and Gene-targeting in Arabidopsis through Agrobacterium-mediated Floral Dip Transformation. Plant Biotechnol. J. 2009, 7, 821–835. [Google Scholar] [CrossRef]
- Cai, C.Q.; Doyon, Y.; Ainley, W.M.; Miller, J.C.; DeKelver, R.C.; Moehle, E.A.; Rock, J.M.; Lee, Y.-L.; Garrison, R.; Schulenberg, L.; et al. Targeted Transgene Integration in Plant Cells Using Designed Zinc Finger Nucleases. Plant Mol. Biol. 2009, 69, 699–709. [Google Scholar] [CrossRef]
- Townsend, J.A.; Wright, D.A.; Winfrey, R.J.; Fu, F. High-Frequency Modification of Plant Genes Using Engineered Zinc-Finger Nucleases. Nature 2009, 459, 442–445. [Google Scholar] [CrossRef]
- Osakabe, K.; Osakabe, Y.; Toki, S. Site-Directed Mutagenesis in Arabidopsis Using Custom-Designed Zinc Finger Nucleases. Proc. Natl. Acad. Sci. USA 2010, 107, 12034–12039. [Google Scholar] [CrossRef]
- Mueller, A.L.; Corbi-Verge, C.; Giganti, D.O.; Ichikawa, D.M.; Spencer, J.M.; MacRae, M.; Garton, M.; Kim, P.M.; Noyes, M.B. The Geometric Influence on the Cys2His2 Zinc Finger Domain and Functional Plasticity. Nucleic Acids Res. 2020, 48, 6382–6402. [Google Scholar] [CrossRef]
- Lloyd, A.; Plaisier, C.L.; Carroll, D. Targeted Mutagenesis Using Zinc-Finger Nucleases in Arabidopsis. Proc. Natl. Acad. Sci. USA 2005, 102, 2232–2237. [Google Scholar] [CrossRef]
- Tovkach, A.; Zeevi, V.; Tzfira, T. A Toolbox and Procedural Notes for Characterizing Novel Zinc Finger Nucleases for Genome Editing in Plant Cells. Plant J. 2009, 57, 747–757. [Google Scholar] [CrossRef]
- Petolino, J.F.; Worden, A.; Curlee, K. Zinc Finger Nuclease-Mediated Transgene Deletion. Plant Mol. Biol. 2010, 73, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Curtin, S.J.; Zhang, F.; Sander, J.D.; Haun, W.J.; Starker, C.; Baltes, N.J.; Reyon, D.; Dahlborg, E.J.; Goodwin, M.J.; Coffman, A.P.; et al. Targeted Mutagenesis of Duplicated Genes in Soybean with Zinc-Finger Nucleases. Plant Physiol. 2011, 156, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.M.; Musunuru, K. Expanding the Genetic Editing Tool Kit: ZFNs, TALENs, and CRISPR-Cas9. J. Clin. Investig. 2014, 124, 4154–4161. [Google Scholar] [CrossRef]
- Mushtaq, M.; Bhat, J.A.; Mir, Z.A.; Sakina, A.; Ali, S.; Singh, A.K.; Tyagi, A.; Salgotra, R.K.; Dar, A.A.; Bhat, R. CRISPR/Cas Approach: A New Way of Looking at Plant-Abiotic Interactions. J. Plant Physiol. 2018, 224–225, 156–162. [Google Scholar] [CrossRef] [PubMed]
- González Castro, N.G.; Bjelic, J.; Malhotra, G.; Huang, C.; Alsaffar, S.H. Comparison of the Feasibility, Efficiency, and Safety of Genome Editing Technologies. Int. J. Mol. Sci. 2021, 22, 10355. [Google Scholar] [CrossRef] [PubMed]
- Schornack, S.; Moscou, M.J.; Ward, E.R.; Horvath, D.M. Engineering Plant Disease Resistance Based on TAL Effectors. Annu. Rev. Phytopathol. 2013, 51, 383–406. [Google Scholar] [CrossRef] [PubMed]
- Boch, J.; Scholze, H.; Schornack, S.; Landgraf, A.; Hahn, S.; Kay, S.; Lahaye, T.; Nickstadt, A.; Bonas, U. Breaking the Code of DNA Binding Specificity of TAL-Type III Effectors. Science 2009, 326, 1509–1512. [Google Scholar] [CrossRef] [PubMed]
- Moscou, M.J.; Bogdanove, A.J. A Simple Cipher Governs DNA Recognition by TAL Effectors. Science 2009, 326, 1501. [Google Scholar] [CrossRef]
- Morbitzer, R.; Römer, P.; Boch, J. Regulation of Selected Genome Loci Using de Novo-Engineered Transcription Activator-like Effector (TALE)-Type Transcription Factors. Proc. Natl. Acad. Sci. USA 2010, 107, 21617–21622. [Google Scholar] [CrossRef]
- Cermak, T.; Doyle, E.L.; Christian, M.; Wang, L.; Zhang, Y.; Schmidt, C.; Baller, J.A.; Somia, N.V.; Bogdanove, A.J.; Voytas, D.F. Efficient Design and Assembly of Custom TALEN and Other TAL Effector-Based Constructs for DNA Targeting. Nucleic Acids Res. 2011, 39, e82. [Google Scholar] [CrossRef]
- Li, T.; Liu, B.; Spalding, M.H. High-Efficiency TALEN-Based Gene Editing Produces Disease-Resistant Rice. Nat. Biotechnol. 2012, 30, 390–392. [Google Scholar] [CrossRef]
- Malzahn, A.; Lowder, L.; Qi, Y. Plant Genome Editing with TALEN and CRISPR. Cell Biosci. 2017, 7, 21. [Google Scholar] [CrossRef]
- Groenen, P.M.A.; Bunschoten, A.E.; van Soolingen, D.; Errtbden, J.D. Nature of DNA Polymorphism in the Direct Repeat Cluster of Mycobacterium Tuberculosis; Application for Strain Differentiation by a Novel Typing Method. Mol. Microbiol. 1993, 10, 1057–1065. [Google Scholar] [CrossRef]
- Mojica, F.J.M.; Juez, G.; Rodriguez-Valera, F. Transcription at Different Salinities of Haloferax Mediterranei Sequences Adjacent to Partially Modified Pst I Sites. Mol. Microbiol. 1993, 9, 613–621. [Google Scholar] [CrossRef]
- Mojica, F.J.M.; Diez-Villasenor, C.; Garcia-Martinez, J. Intervening Sequences of Regularly Spaced Prokaryotic Repeats Derive from Foreign Genetic Elements. J. Mol. Evol. 2005, 60, 174–182. [Google Scholar] [CrossRef]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR Provides Acquired Resistance against Viruses in Prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Marraffini, L.A.; Sontheimer, E.J. CRISPR Interference Limits Horizontal Gene Transfer in Staphylococci by Targeting DNA. Science 2008, 322, 1843–1845. [Google Scholar] [CrossRef] [PubMed]
- Brouns, S.J.J.; Jore, M.M.; Lundgren, M.; Westra, E.R.; Slijkhuis, R.J.H.; Snijders, A.P.L.; Dickman, M.J.; Makarova, K.S.; Koonin, E.V.; van der Oost, J. Small CRISPR RNAs Guide Antiviral Defense in Prokaryotes. Science 2008, 321, 960–964. [Google Scholar] [CrossRef] [PubMed]
- Deltcheva, E.; Chylinski, K.; Sharma, C.M.; Gonzales, K.; Chao, Y.; Pirzada, Z.A.; Eckert, M.R.; Vogel, J.; Charpentier, E. CRISPR RNA Maturation by Trans-Encoded Small RNA and Host Factor RNase III. Nature 2011, 471, 602–607. [Google Scholar] [CrossRef]
- Collias, D.; Beisel, C.L. CRISPR Technologies and the Search for the PAM-Free Nuclease. Nat. Commun. 2021, 12, 555. [Google Scholar] [CrossRef]
- Jinek, M.; East, A.; Cheng, A.; Lin, S.; Ma, E.; Doudna, J. RNA-Programmed Genome Editing in Human Cells. eLife 2013, 2, e00471. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Hwang, W.Y.; Fu, Y.; Reyon, D.; Maeder, M.L.; Tsai, S.Q.; Sander, J.D.; Peterson, R.T.; Yeh, J.-R.J.; Joung, J.K. Efficient Genome Editing in Zebrafish Using a CRISPR-Cas System. Nat. Biotechnol. 2013, 31, 227–229. [Google Scholar] [CrossRef] [PubMed]
- Mali, P.; Yang, L.; Esvelt, K.M. RNA-Guided Human Genome Engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Wolf, Y.I.; Iranzo, J. Evolutionary Classification of CRISPR–Cas Systems: A Burst of Class 2 and Derived Variants. Nat. Rev. Microbiol. 2020, 18, 67–83. [Google Scholar] [CrossRef]
- Makarova, K.S.; Koonin, E.V. Annotation and Classification of CRISPR-Cas Systems. In CRISPR: Methods and Protocols; Elsevier: Amsterdam, The Netherlands, 2015; pp. 47–75. [Google Scholar]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O. Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Nishimasu, H.; Ran, F.A.; Hsu, P.D. Crystal Structure of Cas9 in Complex with Guide RNA and Target DNA. Cell 2014, 156, 935–949. [Google Scholar] [CrossRef]
- Li, J.-F.; Norville, J.E.; Aach, J. Multiplex and Homologous Recombination–Mediated Genome Editing in Arabidopsis and Nicotiana Benthamiana Using Guide RNA and Cas9. Nat. Biotechnol. 2013, 31, 688–691. [Google Scholar] [CrossRef]
- Shan, Q.; Wang, Y.; Li, J. Targeted Genome Modification of Crop Plants Using a CRISPR-Cas System. Nat. Biotechnol. 2013, 31, 686–688. [Google Scholar] [CrossRef]
- Wada, N.; Ueta, R.; Osakabe, Y.; Osakabe, K. Precision Genome Editing in Plants: State-of-the-Art in CRISPR/Cas9-Based Genome Engineering. BMC Plant Biol. 2020, 20, 234. [Google Scholar] [CrossRef]
- Pan, C.; Ye, L.; Qin, L. CRISPR/Cas9-Mediated Efficient and Heritable Targeted Mutagenesis in Tomato Plants in the First and Later Generations. Sci. Rep. 2016, 6, 24765. [Google Scholar] [CrossRef]
- Ueta, R.; Abe, C.; Watanabe, T. Rapid Breeding of Parthenocarpic Tomato Plants Using CRISPR/Cas9. Sci. Rep. 2017, 7, 507. [Google Scholar] [CrossRef]
- D’Ambrosio, C.; Stigliani, A.L.; Giorio, G. CRISPR/Cas9 Editing of Carotenoid Genes in Tomato. Transgenic Res. 2018, 27, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Wang, H.; Sun, C.; Li, Q.; Jiang, H.; Du, M.; Li, C.-B.; Li, C. Efficient Generation of Pink-Fruited Tomatoes Using CRISPR/Cas9 System. J. Genet. Genom. 2018, 45, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wolt, J.D. Risk Associated with Off-Target Plant Genome Editing and Methods for Its Limitation. Emerg. Top. Life Sci. 2017, 1, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Ma, X.; Gao, F.; Guo, Y. Off-Target Effects in CRISPR/Cas9 Gene Editing. Front. Bioeng. Biotechnol. 2023, 11, 1143157. [Google Scholar] [CrossRef] [PubMed]
- Nadakuduti, S.S.; Enciso-Rodríguez, F. Advances in Genome Editing With CRISPR Systems and Transformation Technologies for Plant DNA Manipulation. Front. Plant Sci. 2021, 11, 637159. [Google Scholar] [CrossRef]
- Jones, S.K.; Hawkins, J.A.; Johnson, N.V.; Jung, C.; Hu, K.; Rybarski, J.R.; Chen, J.S.; Doudna, J.A.; Press, W.H.; Finkelstein, I.J. Massively Parallel Kinetic Profiling of Natural and Engineered CRISPR Nucleases. Nat. Biotechnol. 2021, 39, 84–93. [Google Scholar] [CrossRef]
- Paul, B.; Montoya, G. CRISPR-Cas12a: Functional Overview and Applications. Biomed. J. 2020, 43, 8–17. [Google Scholar] [CrossRef]
- Kulcsár, P.I.; Tálas, A.; Ligeti, Z. A Cleavage Rule for Selection of Increased-Fidelity SpCas9 Variants with High Efficiency and No Detectable off-Targets. Nat. Commun. 2023, 14, 5746. [Google Scholar] [CrossRef]
- Endo, A.; Masafumi, M.; Kaya, H.; Toki, S. Efficient Targeted Mutagenesis of Rice and Tobacco Genomes Using Cpf1 from Francisella Novicida. Sci. Rep. 2016, 6, 38169. [Google Scholar] [CrossRef]
- Abudayyeh, O.O.; Gootenberg, J.S.; Konermann, S.; Joung, J.; Slaymaker, I.M.; Cox, D.B.T.; Shmakov, S.; Makarova, K.S.; Semenova, E.; Minakhin, L.; et al. C2c2 Is a Single-Component Programmable RNA-Guided RNA-Targeting CRISPR Effector. Science 2016, 353, aaf5573. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.Z.; Amin, I.; Hameed, A.; Mansoor, S. CRISPR–Cas13a: Prospects for Plant Virus Resistance. Trends Biotechnol. 2018, 36, 1207–1210. [Google Scholar] [CrossRef] [PubMed]
- Pausch, P.; Al-Shayeb, B.; Bisom-Rapp, E. CRISPR-CasΦ from Huge Phages Is a Hypercompact Genome Editor. Science 2020, 369, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Sretenovic, S.; Fan, T.; Cheng, Y.; Li, G.; Qi, A.; Tang, X. Hypercompact CRISPR–Cas12j2 (CasΦ) Enables Genome Editing, Gene Activation, and Epigenome Editing in Plants. Plant Commun. 2022, 3, 100453. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhong, Z.; Wu, Z. Genome Editing in Plants Using the Compact Editor CasΦ. Proc. Natl. Acad. Sci. USA 2023, 120, e2216822120. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.S.; Larson, M.H.; Gilbert, L.A. Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression. Cell 2013, 152, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Hua, K.; Tao, X.; Yuan, F.; Wang, D.; Zhu, J.-K. Precise A·T to G·C Base Editing in the Rice Genome. Mol. Plant 2018, 11, 627–630. [Google Scholar] [CrossRef]
- Nidhi, S.; Anand, U.; Oleksak, P. Novel CRISPR–Cas Systems: An Updated Review of the Current Achievements, Applications, and Future Research Perspectives. Int. J. Mol. Sci. 2021, 22, 3327. [Google Scholar] [CrossRef]
- Li, J.; Zhang, C.; He, Y. Plant Base Editing and Prime Editing: The Current Status and Future Perspectives. J. Integr. Plant Biol. 2023, 65, 444–467. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Joshi, R.K.; Zhao, K. Base Editing in Crops: Current Advances, Limitations and Future Implications. Plant Biotechnol. J. 2020, 18, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-Replace Genome Editing without Double-Strand Breaks or Donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.V.; Nguyen, N.T.; Kim, J. Prime Editing: Mechanism Insight and Recent Applications in Plants. Plant Biotechnol. J. 2023, 22, 19–36. [Google Scholar] [CrossRef] [PubMed]
- Alghuthaymi, M.A.; Ahmad, A.; Khan, Z.; Khan, S.H.; Ahmed, F.K.; Faiz, S.; Nepovimova, E.; Kuča, K.; Abd-Elsalam, K.A. Exosome/Liposome-like Nanoparticles: New Carriers for CRISPR Genome Editing in Plants. Int. J. Mol. Sci. 2021, 22, 7456. [Google Scholar] [CrossRef] [PubMed]
- Laforest, L.C.; Nadakuduti, S.S. Advances in Delivery Mechanisms of CRISPR Gene-Editing Reagents in Plants. Front. Genome Ed. 2022, 4, 830178. [Google Scholar] [CrossRef] [PubMed]
- Rustgi, S.; Naveed, S.; Windham, J.; Zhang, H.; Demirer, G.S. Plant Biomacromolecule Delivery Methods in the 21st Century. Front. Genome Ed. 2022, 4, 1011934. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Xu, C.; Li, T.; Ma, H.; Gong, J.; Li, X.; Sun, X.; Hu, X. Application of Nanotechnology in Plant Genetic Engineering. Int. J. Mol. Sci. 2023, 24, 14836. [Google Scholar] [CrossRef]
- Cunningham, F.J.; Demirer, G.S.; Goh, N.S.; Zhang, H.; Landry, M.P. Nanobiolistics: An Emerging Genetic Transformation Approach. Methods Mol. Biol. 2020, 2124, 141–159. [Google Scholar] [CrossRef]
- Zhang, Y.; Iaffaldano, B.; Qi, Y. CRISPR Ribonucleoprotein-Mediated Genetic Engineering in Plants. Plant Commun. 2021, 2, 100168. [Google Scholar] [CrossRef]
- Nicolia, A.; Andersson, M.; Hofvander, P. Tomato Protoplasts as Cell Target for Ribonucleoprotein (RNP)-Mediated Multiplexed Genome Editing. Plant Cell Tissue Organ Cult. 2021, 144, 463–467. [Google Scholar] [CrossRef]
- Liu, Y.; Andersson, M.; Granell, A. Establishment of a DNA-Free Genome Editing and Protoplast Regeneration Method in Cultivated Tomato (Solanum lycopersicum). Plant Cell Rep. 2022, 41, 1843–1852. [Google Scholar] [CrossRef] [PubMed]
- Kang, G.H.; Kang, B.-C.; Han, J.-S. DNA-Free Genome Editing in Tomato Protoplasts Using CRISPR/Cas9 Ribonucleoprotein Delivery. Hortic. Environ. Biotechnol. 2023, 65, 1–12. [Google Scholar] [CrossRef]
- Slaman, E.; Lammers, M.; Angenent, G.C.; de Maagd, R.A. High-Throughput SgRNA Testing Reveals Rules for Cas9 Specificity and DNA Repair in Tomato Cells. Front. Genome Ed. 2023, 5, 1196763. [Google Scholar] [CrossRef]
- Lin, C.-S.; Hsu, C.-T.; Yuan, Y.-H. DNA-Free CRISPR-Cas9 Gene Editing of Wild Tetraploid Tomato Solanum Peruvianum Using Protoplast Regeneration. Plant Physiol. 2022, 188, 1917–1930. [Google Scholar] [CrossRef]
- Pǎcurar, D.I.; Thordal-Christensen, H.; Pǎcurar, M.L.; Pamfil, D.; Botez, M.L.; Bellini, C. Agrobacterium Tumefaciens: From Crown Gall Tumors to Genetic Transformation. Physiol. Mol. Plant Pathol. 2011, 76, 76–81. [Google Scholar] [CrossRef]
- Hwang, H.H.; Yu, M.; Lai, E.M. Agrobacterium-Mediated Plant Transformation: Biology and Applications. Arab. Book 2017, 15, e0186. [Google Scholar] [CrossRef]
- Kiryushkin, A.S.; Ilina, E.L.; Guseva, E.D.; Pawlowski, K.; Demchenko, K.N. Hairy CRISPR: Genome Editing in Plants Using Hairy Root Transformation. Plants 2022, 11, 51. [Google Scholar] [CrossRef] [PubMed]
- De Saeger, J.; Park, J.; Chung, H.S.; Hernalsteens, J.P.; Van Lijsebettens, M.; Inzé, D.; Van Montagu, M.; Depuydt, S. Agrobacterium Strains and Strain Improvement: Present and Outlook. Biotechnol. Adv. 2021, 53, 107677. [Google Scholar] [CrossRef]
- Rodrigues, S.D.; Karimi, M.; Impens, L.; van Lerberge, E.; Coussens, G.; Aesaert, S.; Rombaut, D.; Holtappels, D.; Ibrahim, H.M.M.; van Montagu, M.; et al. Efficient CRISPR-Mediated Base Editing in Agrobacterium spp. Proc. Natl. Acad. Sci. USA 2021, 118, e2013338118. [Google Scholar] [CrossRef]
- Raman, V.; Rojas, C.M.; Vasudevan, B.; Dunning, K.; Kolape, J.; Oh, S.; Yun, J.; Yang, L.; Li, G.; Pant, B.D.; et al. Agrobacterium Expressing a Type III Secretion System Delivers Pseudomonas Effectors into Plant Cells to Enhance Transformation. Nat. Commun. 2022, 13, 2581. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral Dip: A Simplified Method for Agrobacterium-Mediated Transformation of Arabidopsis Thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Yaroshko, O.; Pasternak, T.; Larriba, E.; Pérez-Pérez, J.M. Optimization of Callus Induction and Shoot Regeneration from Tomato Cotyledon Explants. Plants 2023, 12, 2942. [Google Scholar] [CrossRef] [PubMed]
- Chincinska, I.A. Leaf Infiltration in Plant Science: Old Method, New Possibilities. Plant Methods 2021, 17, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Roberts, H.M.; Van Eck, J. Generation and Molecular Characterization of CRISPR/Cas9-Induced Mutations in 63 Immunity-Associated Genes in Tomato Reveals Specificity and a Range of Gene Modifications. Front. Plant Sci. 2020, 11, 10. [Google Scholar] [CrossRef]
- Hong, Y.; Meng, J.; He, X.; Zhang, Y.; Liu, Y.; Zhang, C.; Qi, H.; Luan, Y. Editing MiR482b and MiR482c Simultaneously by CRISPR/Cas9 Enhanced Tomato Resistance to Phytophthora Infestans. Phytopathology® 2021, 111, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Rössner, C.; Lotz, D.; Becker, A. VIGS Goes Viral: How VIGS Transforms Our Understanding of Plant Science. Annu. Rev. Plant Biol. 2022, 73, 703–728. [Google Scholar] [CrossRef] [PubMed]
- Čermák, T.; Baltes, N.J.; Čegan, R.; Zhang, Y.; Voytas, D.F. High-Frequency, Precise Modification of the Tomato Genome. Genome Biol. 2015, 16, 232. [Google Scholar] [CrossRef] [PubMed]
- Uranga, M.; Aragones, V.; Garcia, A. RNA Virus-Mediated Gene Editing for Tomato Trait Breeding. bioRxiv 2023. [Google Scholar] [CrossRef]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering Precision Nanoparticles for Drug Delivery. Nat. Rev. Drug Discov. 2020, 20, 101–124. [Google Scholar] [CrossRef]
- Mahmoud, L.M.; Kaur, P.; Stanton, D.; Grosser, J.W.; Dutt, M. A Cationic Lipid Mediated CRISPR/Cas9 Technique for the Production of Stable Genome Edited Citrus Plants. Plant Methods 2022, 18, 33. [Google Scholar] [CrossRef]
- Liu, W.; Rudis, M.R.; Cheplick, M.H.; Millwood, R.J.; Yang, J.P.; Ondzighi-Assoume, C.A.; Montgomery, G.A.; Burris, K.P.; Mazarei, M.; Chesnut, J.D.; et al. Lipofection-Mediated Genome Editing Using DNA-Free Delivery of the Cas9/GRNA Ribonucleoprotein into Plant Cells. Plant Cell Rep. 2020, 39, 245–257. [Google Scholar] [CrossRef]
- Nagy, B.; Öktem, A.; Ferenc, G.; Ungor, D.; Kalac, A.; Kelemen-Valkony, I.; Fodor, E.; Nagy, I.; Dudits, D.; Ayaydin, F. CRISPR/Cas9 Mutagenesis through Introducing a Nanoparticle Complex Made of a Cationic Polymer and Nucleic Acids into Maize Protoplasts. Int. J. Mol. Sci. 2023, 24, 16137. [Google Scholar] [CrossRef]
- Ozyigit, I.I. Gene Transfer to Plants by Electroporation: Methods and Applications. Mol. Biol. Rep. 2020, 47, 3195–3210. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Lee, J.; Choi, S.A.; Kim, Y.S.; Koo, O.; Choi, S.H.; Ahn, W.S.; Jie, E.Y.; Kim, S.W. Efficient Genome Editing Using CRISPR–Cas9 RNP Delivery into Cabbage Protoplasts via Electro-Transfection. Plant Biotechnol. Rep. 2020, 14, 695–702. [Google Scholar] [CrossRef]
- Subburaj, S.; Agapito-Tenfen, S.Z. Establishment of Targeted Mutagenesis in Soybean Protoplasts Using CRISPR/Cas9 RNP Delivery via Electro−transfection. Front. Plant Sci. 2023, 14, 1255819. [Google Scholar] [CrossRef]
- Hilioti, Z. Non-Transgenic Approach to Deliver ZFNs in Seeds for Targeted Genome Engineering. Methods Mol. Biol. 2018, 1867, 187–199. [Google Scholar] [CrossRef]
- Komatsu, H.; Abdellatif, I.M.Y.; Yuan, S.; Ono, M.; Nonaka, S.; Ezura, H.; Ariizumi, T.; Miura, K. Genome Editing in PDS Genes of Tomatoes by Non-Selection Method and of Nicotiana Benthamiana by One Single Guide RNA to Edit Two Orthologs. Plant Biotechnol. 2020, 37, 213. [Google Scholar] [CrossRef]
- Rinne, J.; Witte, C.P.; Herde, M. Loss of MAR1 Function Is a Marker for Co-Selection of CRISPR-Induced Mutations in Plants. Front. Genome Ed. 2021, 3, 723384. [Google Scholar] [CrossRef]
- Shukla, V.; Gupta, M.; Urnov, F. Targeted Modification of Malate Dehydrogenase. Patent nº US9523098B2, 20 December 2016. [Google Scholar]
- Hilioti, Z.; Ganopoulos, I.; Ajith, S.; Bossis, I.; Tsaftaris, A. A Novel Arrangement of Zinc Finger Nuclease System for in Vivo Targeted Genome Engineering: The Tomato LEC1-LIKE4 Gene Case. Plant Cell Rep. 2016, 35, 2241–2255. [Google Scholar] [CrossRef] [PubMed]
- Lor, V.S.; Starker, C.G.; Voytas, D.F. Targeted Mutagenesis of the Tomato PROCERA Gene Using Transcription Activator-Like Effector Nucleases. Plant Physiol. 2014, 166, 1288–1291. [Google Scholar] [CrossRef] [PubMed]
- Kuroiwa, K.; Danilo, B.; Perrot, L. An Iterative Gene-editing Strategy Broadens EIF4E1genetic Diversity in Solanum lycopersicum and Generates Resistance to Multiple Potyvirus Isolates. Plant Biotechnol. J. 2023, 21, 918–930. [Google Scholar] [CrossRef] [PubMed]
- Brooks, C.; Nekrasov, V.; Lippman, Z.B.; Van Eck, J. Efficient Gene Editing in Tomato in the First Generation Using the Clustered Regularly Interspaced Short Palindromic Repeats/CRISPR-Associated9 System. Plant Physiol. 2014, 166, 1292–1297. [Google Scholar] [CrossRef] [PubMed]
- Ron, M.; Kajala, K.; Pauluzzi, G. Hairy Root Transformation Using Agrobacterium Rhizogenes as a Tool for Exploring Cell Type-Specific Gene Expression and Function Using Tomato as a Model. Plant Physiol. 2014, 166, 455–469. [Google Scholar] [CrossRef] [PubMed]
- Yifhar, T.; Pekker, I.; Peled, D.; Friedlander, G.; Pistunov, A.; Sabban, M.; Wachsman, G.; Alvarez, J.P.; Amsellem, Z.; Eshed, Y. Failure of the Tomato Trans-Acting Short Interfering RNA Program to Regulate AUXIN RESPONSE FACTOR3 and ARF4 Underlies the Wiry Leaf Syndrome. Plant Cell 2012, 24, 3575–3589. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, M.; Boopathi, T.; Paramasivan, M. A Status-Quo Review on CRISPR-Cas9 Gene Editing Applications in Tomato. Int. J. Biol. Macromol. 2021, 190, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, J.K.; Singh, A.K.; Behera, T.K. CRISPR/Cas Genome Editing in Tomato Improvement: Advances and Applications. Front. Plant Sci. 2023, 14, 1121209. [Google Scholar] [CrossRef]
- Hayut, S.F.; Bessudo, C.M.; Levy, A.A. Targeted Recombination between Homologous Chromosomes for Precise Breeding in Tomato. Nat. Commun. 2017, 8, 15605. [Google Scholar] [CrossRef]
- Rodríguez-Leal, D.; Lemmon, Z.H.; Man, J. Engineering Quantitative Trait Variation for Crop Improvement by Genome Editing. Cell 2017, 171, 470–480.e8. [Google Scholar] [CrossRef]
- Ye, J.; Wang, X.; Hu, T. An InDel in the Promoter of Al-ACTIVATED MALATE TRANSPORTER9 Selected during Tomato Domestication Determines Fruit Malate Contents and Aluminum Tolerance. Plant Cell 2017, 29, 2249–2268. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yang, X.; Yu, Y. Domestication of Wild Tomato Is Accelerated by Genome Editing. Nat. Biotechnol. 2018, 36, 1160–1163. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhang, B.; Keyhaninejad, N. A Common Genetic Mechanism Underlies Morphological Diversity in Fruits and Other Plant Organs. Nat. Commun. 2018, 9, 4734. [Google Scholar] [CrossRef] [PubMed]
- Zsögön, A.; Čermák, T.; Naves, E.R.; Notini, M.M.; Edel, K.H.; Weinl, S.; Freschi, L.; Voytas, D.F.; Kudla, J.; Peres, L.E.P. De Novo Domestication of Wild Tomato Using Genome Editing. Nat. Biotechnol. 2018, 36, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
- Blanca, J.; Montero-Pau, J.; Sauvage, C.; Bauchet, G.; Illa, E.; Díez, M.J.; Francis, D.; Causse, M.; van der Knaap, E.; Cañizares, J. Genomic Variation in Tomato, from Wild Ancestors to Contemporary Breeding Accessions. BMC Genom. 2015, 16, 257. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, Z.H.; Reem, N.T.; Dalrymple, J.; Soyk, S.; Swartwood, K.E.; Rodriguez-Leal, D.; Van Eck, J.; Lippman, Z.B. Rapid Improvement of Domestication Traits in an Orphan Crop by Genome Editing. Nat. Plants 2018, 4, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.Z.; Zaidi, S.S.-A.; Amin, I.; Mansoor, S. A CRISPR Way for Fast-Forward Crop Domestication. Trends Plant Sci. 2019, 24, 293–296. [Google Scholar] [CrossRef]
- Vu, T.V.; Das, S.; Tran, M.T. Precision Genome Engineering for the Breeding of Tomatoes: Recent Progress and Future Perspectives. Front. Genome Ed. 2020, 2, 612137. [Google Scholar] [CrossRef]
- Tanksley, S.D. The Genetic, Developmental, and Molecular Bases of Fruit Size and Shape Variation in Tomato. Plant Cell Online 2004, 16, S181–S189. [Google Scholar] [CrossRef]
- Chattopadhyay, T.; Hazra, P.; Akhtar, S.; Maurya, D.; Mukherjee, A.; Roy, S. Skin Colour, Carotenogenesis and Chlorophyll Degradation Mutant Alleles: Genetic Orchestration behind the Fruit Colour Variation in Tomato. Plant Cell Rep. 2021, 40, 767–782. [Google Scholar] [CrossRef]
- Quinet, M.; Angosto, T.; Yuste-Lisbona, F.J.; Blanchard-Gros, R.; Bigot, S.; Martinez, J.-P.; Lutts, S. Tomato Fruit Development and Metabolism. Front. Plant Sci. 2019, 10, 1554. [Google Scholar] [CrossRef]
- Karlova, R.; Chapman, N.; David, K.; Angenent, G.C.; Seymour, G.B.; de Maagd, R.A. Transcriptional Control of Fleshy Fruit Development and Ripening. J. Exp. Bot. 2014, 65, 4527–4541. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Nishizawa-Yokoi, A.; Endo, M.; Mikami, M.; Toki, S. CRISPR/Cas9-Mediated Mutagenesis of the RIN Locus That Regulates Tomato Fruit Ripening. Biochem. Biophys. Res. Commun. 2015, 467, 76–82. [Google Scholar] [CrossRef]
- Ito, Y.; Sekiyama, Y.; Nakayama, H.; Nishizawa-Yokoi, A.; Endo, M.; Shima, Y.; Nakamura, N.; Kotake-Nara, E.; Kawasaki, S.; Hirose, S.; et al. Allelic Mutations in the Ripening-Inhibitor Locus Generate Extensive Variation in Tomato Ripening. Plant Physiol. 2020, 183, 80–95. [Google Scholar] [CrossRef]
- Gao, Y.; Zhu, N.; Zhu, X.; Wu, M.; Jiang, C.-Z.; Grierson, D.; Luo, Y.; Shen, W.; Zhong, S.; Fu, D.-Q.; et al. Diversity and Redundancy of the Ripening Regulatory Networks Revealed by the FruitENCODE and the New CRISPR/Cas9 CNR and NOR Mutants. Hortic. Res. 2019, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Song, X.; Zheng, Q.; Liu, Y.; Yu, J.; Zhou, Y.; Xia, X. The Transcription Factor SPL13 Mediates Strigolactone Suppression of Shoot Branching by Inhibiting Cytokinin Synthesis in Solanum lycopersicum. J. Exp. Bot. 2023, 74, 5722–5735. [Google Scholar] [CrossRef]
- Wang, R.; Angenent, G.C.; Seymour, G.; de Maagd, R.A. Revisiting the Role of Master Regulators in Tomato Ripening. Trends Plant Sci. 2020, 25, 291–301. [Google Scholar] [CrossRef]
- Naeem, M.; Zhao, W.; Ahmad, N.; Zhao, L. Beyond Green and Red: Unlocking the Genetic Orchestration of Tomato Fruit Color and Pigmentation. Funct. Integr. Genom. 2023, 23, 243. [Google Scholar] [CrossRef] [PubMed]
- Kou, X.; Zhao, Y.; Wu, C.; Jiang, B.; Zhang, Z.; Rathbun, J.R.; He, Y.; Xue, Z. SNAC4 and SNAC9 Transcription Factors Show Contrasting Effects on Tomato Carotenoids Biosynthesis and Softening. Postharvest Biol. Technol. 2018, 144, 9–19. [Google Scholar] [CrossRef]
- Feng, Y.; Kou, X.; Yuan, S.; Wu, C.; Zhao, X.; Xue, Z.; Li, Q.; Huang, Z.; Sun, Y. CRISPR/Cas9-Mediated SNAC9 Mutants Reveal the Positive Regulation of Tomato Ripening by SNAC9 and the Mechanism of Carotenoid Metabolism Regulation. Hortic. Res. 2023, 10, uhad019. [Google Scholar] [CrossRef]
- Ilahy, R.; Tlili, I.; Siddiqui, M.W.; Hdider, C.; Lenucci, M.S. Inside and Beyond Color: Comparative Overview of Functional Quality of Tomato and Watermelon Fruits. Front. Plant. Sci. 2019, 10, 769. [Google Scholar] [CrossRef]
- Yang, T.; Ali, M.; Lin, L.; Li, P.; He, H.; Zhu, Q.; Sun, C.; Wu, N.; Zhang, X.; Huang, T.; et al. Recoloring Tomato Fruit by CRISPR/Cas9-Mediated Multiplex Gene Editing. Hortic. Res. 2023, 10, uhac214. [Google Scholar] [CrossRef]
- Wang, D.; Samsulrizal, N.H.; Yan, C.; Allcock, N.S.; Craigon, J.; Blanco-Ulate, B.; Ortega-Salazar, I.; Marcus, S.E.; Bagheri, H.M.; Perez-Fons, L.; et al. Characterization of CRISPR Mutants Targeting Genes Modulating Pectin Degradation in Ripening Tomato. Plant Physiol. 2019, 179, 544. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; Lin, Y.; Wang, C.; Lu, J.; Liu, Z.; He, Z.; Shu, X.; Chen, W.; Wu, R.; Li, B.; et al. Expansin SlExp1 and Endoglucanase SlCel2 Synergistically Promote Fruit Softening and Cell Wall Disassembly in Tomato. Plant Cell 2023, koad291. [Google Scholar] [CrossRef]
- Liu, H.; Liu, L.; Liang, D.; Zhang, M.; Jia, C.; Qi, M.; Liu, Y.; Shao, Z.; Meng, F.; Hu, S.; et al. SlBES1 Promotes Tomato Fruit Softening through Transcriptional Inhibition of PMEU1. iScience 2021, 24, 102926. [Google Scholar] [CrossRef]
- Peng, J.; Richards, D.E.; Hartley, N.M.; Murphy, G.P.; Devos, K.M.; Flintham, J.E.; Beales, J.; Fish, L.J.; Worland, A.J.; Pelica, F.; et al. ‘Green Revolution’ Genes Encode Mutant Gibberellin Response Modulators. Nature 1999, 400, 256–261. [Google Scholar] [CrossRef]
- Song, X.; Meng, X.; Guo, H.; Cheng, Q.; Jing, Y.; Chen, M.; Liu, G.; Wang, B.; Wang, Y.; Li, J.; et al. Targeting a Gene Regulatory Element Enhances Rice Grain Yield by Decoupling Panicle Number and Size. Nat. Biotechnol. 2022, 40, 1403–1411. [Google Scholar] [CrossRef] [PubMed]
- Kwon, C.-T.; Heo, J.; Lemmon, Z.H.; Capua, Y.; Hutton, S.F.; Van Eck, J.; Park, S.J.; Lippman, Z.B. Rapid Customization of Solanaceae Fruit Crops for Urban Agriculture. Nat. Biotechnol. 2020, 38, 182–188. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, C.A.; McIntyre, C.L.; Dry, I.B.; Hani, S.M.; Hochman, Z.; Bonnett, G.D. Vertical Farms Bear Fruit. Nat. Biotechnol. 2020, 38, 160–162. [Google Scholar] [CrossRef]
- Soyk, S.; Müller, N.A.; Park, S.J. Variation in the Flowering Gene SELF PRUNING 5G Promotes Day-Neutrality and Early Yield in Tomato. Nat. Genet. 2017, 49, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Chen, Y.; Liu, Y.; Shang, J.; Sun, X.; Du, J. Multifaceted Roles of the ERECTA Family in Plant Organ Morphogenesis. J. Exp. Bot. 2022, 73, 7208–7218. [Google Scholar] [CrossRef] [PubMed]
- Roldan, M.V.G.; Périlleux, C.; Morin, H.; Huerga-Fernandez, S.; Latrasse, D.; Benhamed, M.; Bendahmane, A. Natural and Induced Loss of Function Mutations in SlMBP21 MADS-Box Gene Led to Jointless-2 Phenotype in Tomato. Sci. Rep. 2017, 7, 4402. [Google Scholar] [CrossRef] [PubMed]
- Soyk, S.; Lemmon, Z.H.; Sedlazeck, F.J.; Jiménez-Gómez, J.M.; Alonge, M.; Hutton, S.F.; Van Eck, J.; Schatz, M.C.; Lippman, Z.B. Duplication of a Domestication Locus Neutralized a Cryptic Variant That Caused a Breeding Barrier in Tomato. Nat. Plants 2019, 5, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Alonge, M.; Wang, X.; Benoit, M.; Soyk, S.; Pereira, L.; Zhang, L.; Suresh, H.; Ramakrishnan, S.; Maumus, F.; Ciren, D.; et al. Major Impacts of Widespread Structural Variation on Gene Expression and Crop Improvement in Tomato. Cell 2020, 182, 145–161. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, X.; Zhang, Y.; Zhang, Q. CRISPR/Cas Genome Editing Improves Abiotic and Biotic Stress Tolerance of Crops. Front. Genome Ed. 2022, 4, 987817. [Google Scholar] [CrossRef]
- Nascimento, F.D.; Rocha, A.D.; Soares, J.M.; Mascarenhas, M.S.; Ferreira, M.D.; Morais Lino, L.S.; Ramos, A.P.; Diniz, L.E.; Mendes, T.A.; Ferreira, C.F.; et al. Gene Editing for Plant Resistance to Abiotic Factors: A Systematic Review. Plants 2023, 12, 305. [Google Scholar] [CrossRef]
- Sardar, A. Genetic Amelioration of Fruit and Vegetable Crops to Increase Biotic and Abiotic Stress Resistance through CRISPR Genome Editing. Front. Plant Sci. 2023, 14, 1260102. [Google Scholar] [CrossRef]
- Nekrasov, V.; Wang, C.; Win, J. Rapid Generation of a Transgene-Free Powdery Mildew Resistant Tomato by Genome Deletion. Sci. Rep. 2017, 7, 482. [Google Scholar] [CrossRef]
- Pramanik, D.; Shelake, R.M.; Park, J. CRISPR/Cas9-Mediated Generation of Pathogen-Resistant Tomato against Tomato Yellow Leaf Curl Virus and Powdery Mildew. Int. J. Mol. Sci. 2021, 22, 1878. [Google Scholar] [CrossRef]
- Thomazella, D.P.T.; Seong, K.; Mackelprang, R. Loss of Function of a DMR6 Ortholog in Tomato Confers Broad-Spectrum Disease Resistance. Proc. Natl. Acad. Sci. USA 2021, 118, e2026152118. [Google Scholar] [CrossRef]
- Santillán Martínez, M.I.; Bracuto, V.; Koseoglou, E.; Appiano, M.; Jacobsen, E.; Visser, R.G.F.; Wolters, A.-M.A.; Bai, Y. CRISPR/Cas9-Targeted Mutagenesis of the Tomato Susceptibility Gene PMR4 for Resistance against Powdery Mildew. BMC Plant Biol. 2020, 20, 284. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Maioli, A.; Yan, Z.; Bai, Y.; Valentino, D.; Milani, A.M.; Pompili, V.; Comino, C.; Lanteri, S.; Moglia, A.; et al. CRISPR/Cas9-Based Knock-Out of the PMR4 Gene Reduces Susceptibility to Late Blight in Two Tomato Cultivars. Int. J. Mol. Sci. 2022, 23, 14542. [Google Scholar] [CrossRef] [PubMed]
- Tashkandi, M.; Ali, Z.; Aljedaani, F. Engineering Resistance against Tomato Yellow Leaf Curl Virus via the CRISPR/Cas9 System in Tomato. Plant Signal. Behav. 2018, 13, e1525996. [Google Scholar] [CrossRef]
- El-Sappah, A.H.; Qi, S.; Soaud, S.A.; Huang, Q.; Saleh, A.M.; Abourehab, M.A.S.; Wan, L.; Cheng, G.; Liu, J.; Ihtisham, M.; et al. Natural Resistance of Tomato Plants to Tomato Yellow Leaf Curl Virus. Front. Plant Sci. 2022, 13, 1081549. [Google Scholar] [CrossRef]
- Taheri, S.; Gantait, S.; Azizi, P.; Mazumdar, P. Drought Tolerance Improvement in Solanum lycopersicum: An Insight into “OMICS” Approaches and Genome Editing. 3 Biotech 2022, 12, 63. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, J.; Xu, J. CRISPR/Cas9 Targeted Mutagenesis of SlLBD40, a Lateral Organ Boundaries Domain Transcription Factor, Enhances Drought Tolerance in Tomato. Plant Sci. 2020, 301, 110683. [Google Scholar] [CrossRef]
- Bouzroud, S.; Gasparini, K.; Hu, G.; Barbosa, M.A.M.; Rosa, B.L.; Fahr, M.; Bendaou, N.; Bouzayen, M.; Zsögön, A.; Smouni, A.; et al. Down Regulation and Loss of Auxin Response Factor 4 Function Using CRISPR/Cas9 Alters Plant Growth, Stomatal Function and Improves Tomato Tolerance to Salinity and Osmotic Stress. Genes 2020, 11, 272. [Google Scholar] [CrossRef]
- Chen, M.; Zhu, X.; Liu, X.; Wu, C.; Yu, C.; Hu, G.; Chen, L.; Chen, R.; Bouzayen, M.; Zouine, M.; et al. Knockout of Auxin Response Factor SlARF4 Improves Tomato Resistance to Water Deficit. Int. J. Mol. Sci. 2021, 22, 3347. [Google Scholar] [CrossRef] [PubMed]
- Moreira, J.D.; Quiñones, A.; Lira, B.S.; Robledo, J.M.; Curtin, S.J.; Vicente, M.H.; Ribeiro, D.M.; Ryngajllo, M.; Jiménez-Gómez, J.M.; Peres, L.E.P.; et al. SELF PRUNING 3C Is a Flowering Repressor That Modulates Seed Germination, Root Architecture, and Drought Responses. J. Exp. Bot. 2022, 73, 6226–6240. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Moreno, J.-P.; Tzfadia, O.; Forment, J.; Presa, S.; Rogachev, I.; Meir, S.; Orzaez, D.; Aharoni, A.; Granell, A. Characterization of a New Pink-Fruited Tomato Mutant Results in the Identification of a Null Allele of the SlMYB12 Transcription Factor. Plant Physiol. 2016, 171, 1821–1836. [Google Scholar] [CrossRef]
- Bernabé-Orts, J.M.; Casas-Rodrigo, I.; Minguet, E.G.; Landolfi, V.; Garcia-Carpintero, V.; Gianoglio, S.; Vázquez-Vilar, M.; Granell, A.; Orzaez, D. Assessment of Cas12a-mediated Gene Editing Efficiency in Plants. Plant Biotechnol. J. 2019, 17, 1971–1984. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.V.; Sivankalyani, V.; Kim, E.; Doan, D.T.H.; Tran, M.T.; Kim, J.; Sung, Y.W.; Park, M.; Kang, Y.J.; Kim, J. Highly Efficient Homology-directed Repair Using CRISPR/Cpf1-geminiviral Replicon in Tomato. Plant Biotechnol. J. 2020, 18, 2133–2143. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Raddatz, N.; Aman, R.; Kim, S.; Park, H.C.; Jan, M.; Baek, D.; Khan, I.U.; Oh, D.-H.; Lee, S.Y.; et al. A Single Amino-Acid Substitution in the Sodium Transporter HKT1 Associated with Plant Salt Tolerance. Plant Physiol. 2016, 171, 2112–2126. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.V.; Doan, D.T.H.; Tran, M.T. Improvement of the LbCas12a-CrRNA System for Efficient Gene Targeting in Tomato. Front. Plant Sci. 2021, 12, 722552. [Google Scholar] [CrossRef]
- Huang, X.; Jia, H.; Jin, X. Efficient Transgene-Free Genome Editing in Plants in the T0 Generation Based on a Co-Editing Strategy. bioRxiv 2023. [Google Scholar] [CrossRef]

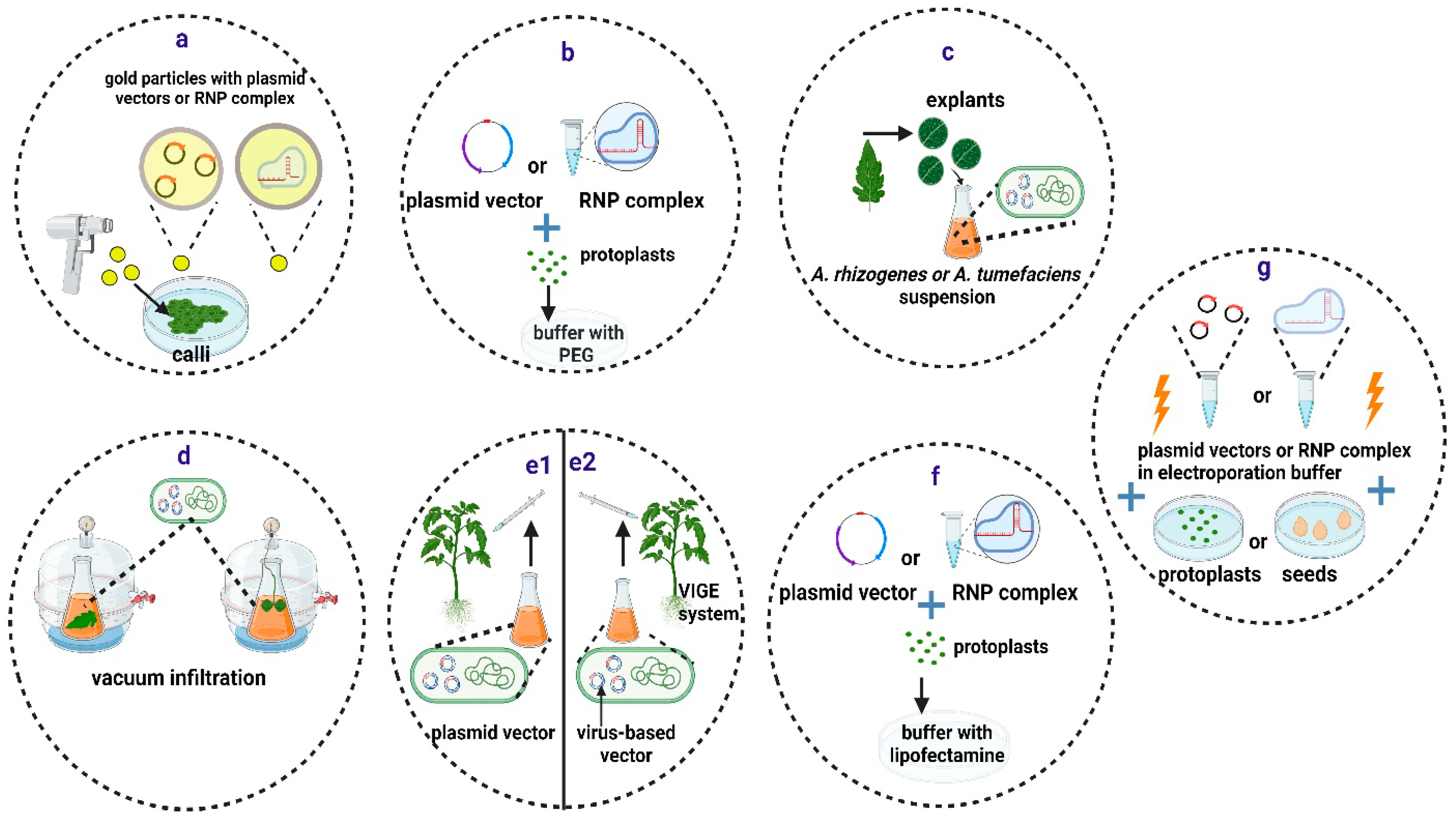
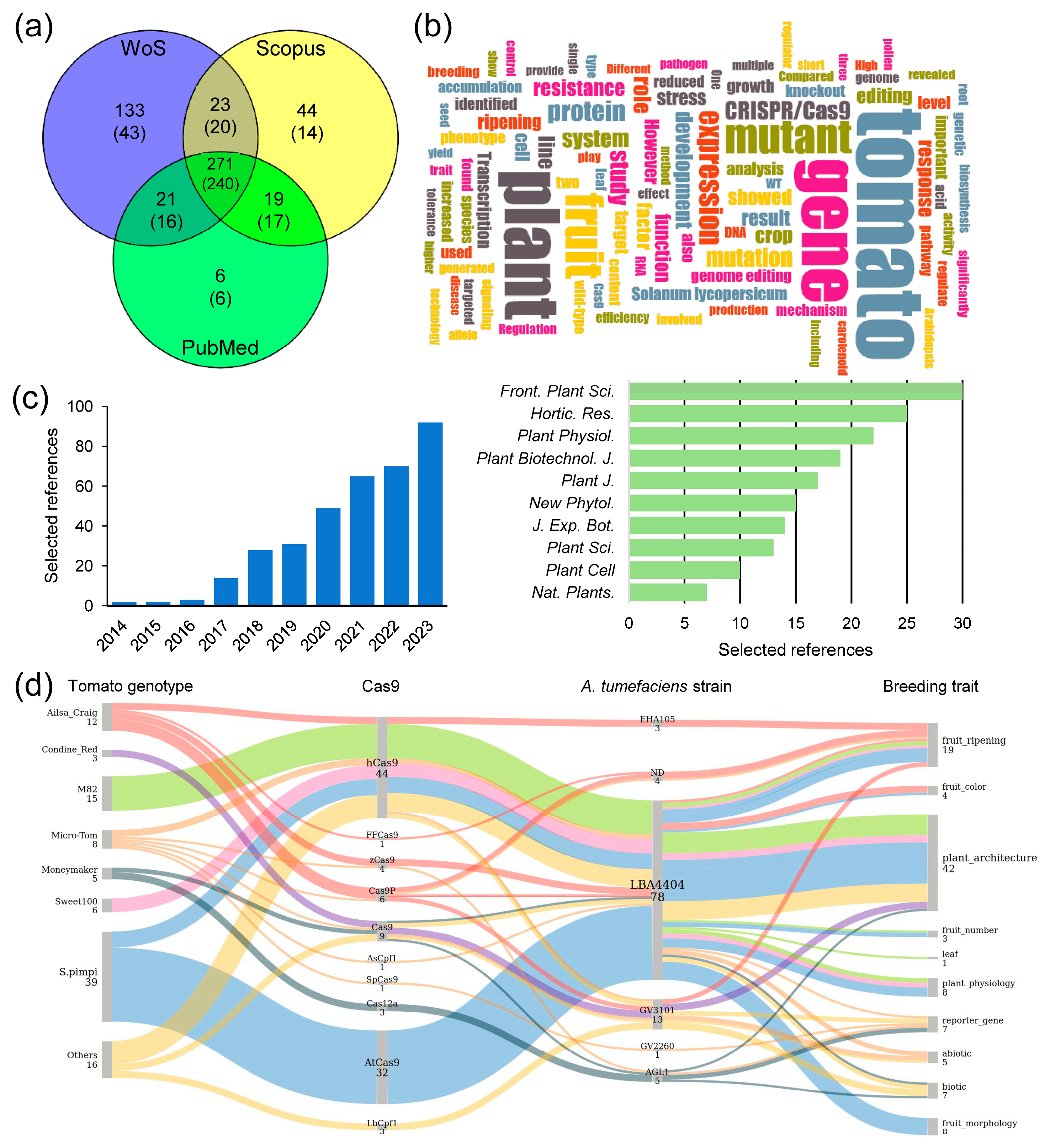
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).