

Age-Dependent Changes in the Occurrence and Segregation of GABA and Acetylcholine in the Rat Superior Cervical Ganglia

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
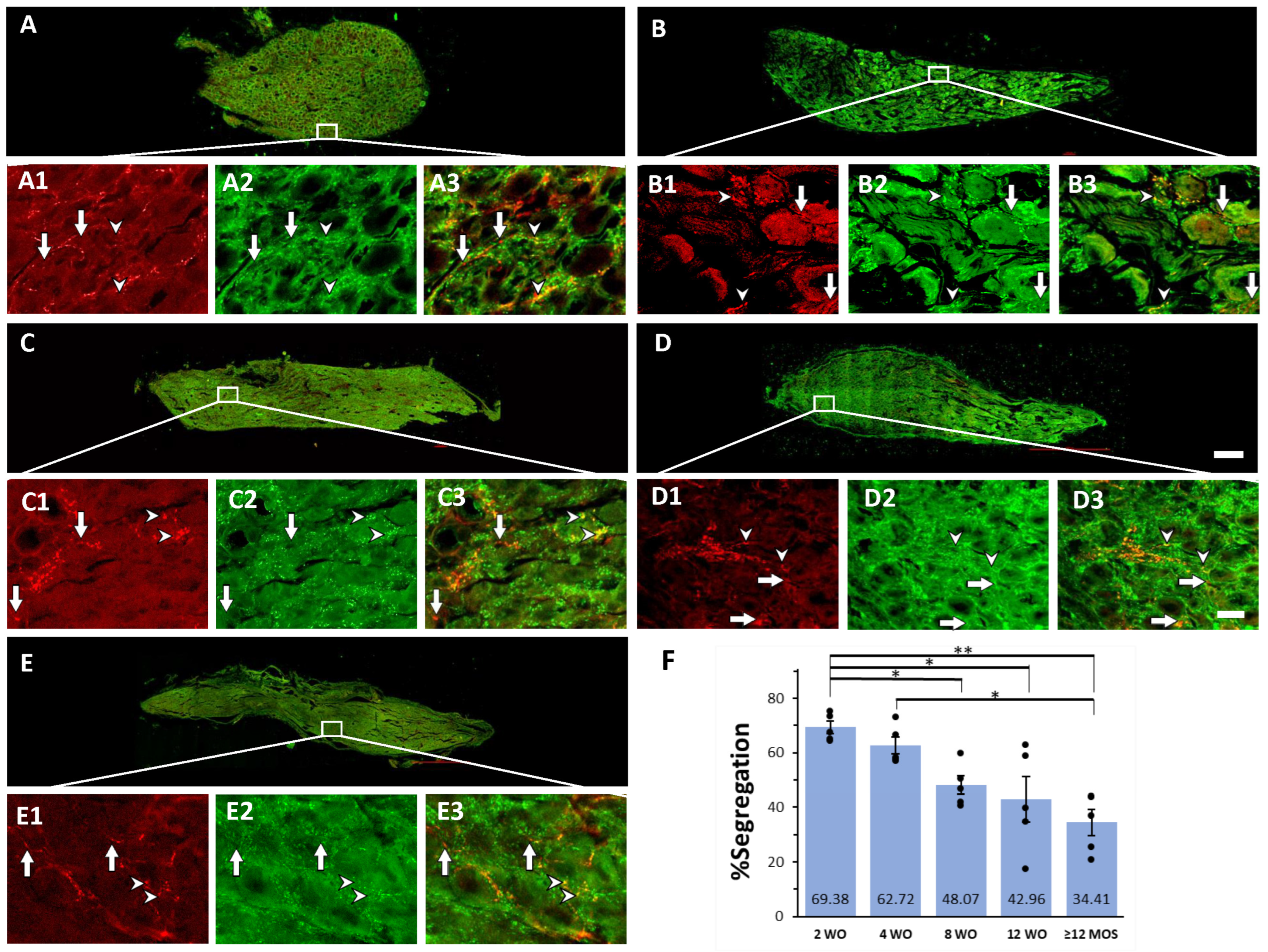
2.1. GAD67, VAChT, and Segregation Occurred in Preganglionic Varicosities
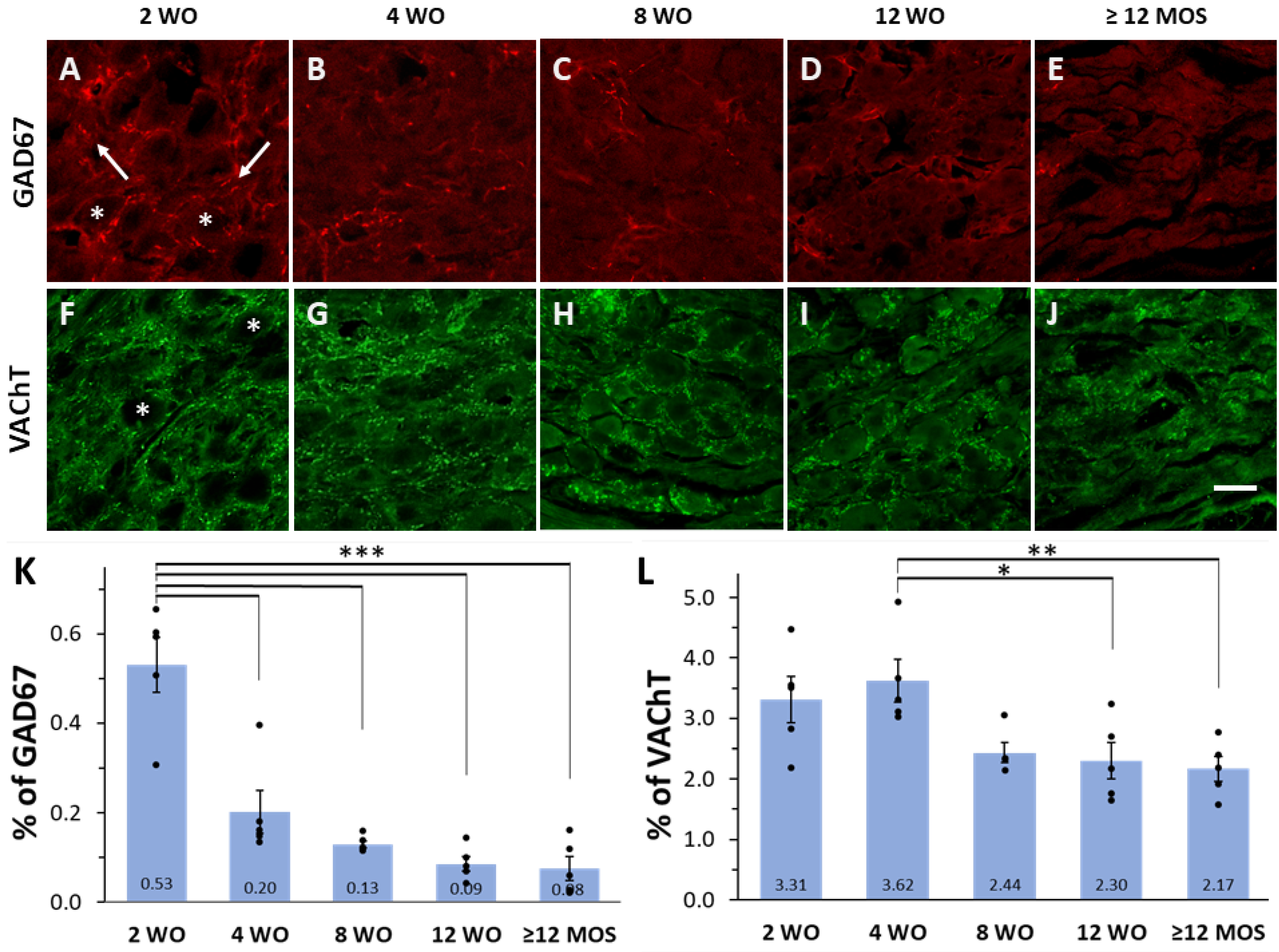
2.2. Expression of GAD67 and VAChT Depends on Age
2.3. Age-Dependent Variations in Segregation of GAD67 and VAChT
3. Discussion
4. Methods and Materials
4.1. Animals
4.2. Immunohistochemistry
4.3. Image Acquisition and Analysis
4.4. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adams, P.R.; Brown, D.A. Actions of gamma-aminobutyric acid on sympathetic ganglion cells. J. Physiol. 1975, 250, 85–120. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.L.; Olsen, R.W. GABA in the Nervous System: The View at Fifty Years; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2000; p. 52. [Google Scholar]
- Stein, V.; Nicoll, R.A. GABA generates excitement. Neuron 2003, 37, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Wolff, J.R.; Joó, F.; Kása, P.; Storm-Mathiesen, J.; Toldi, J.; Balcar, V.J. Presence of neurons with GABA-like immunoreactivity in the superior cervical ganglion of the rat. Neurosci. Lett. 1986, 71, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Kása, P.; Joó, F.; Dobó, E.; Wenthold, R.J.; Ottersen, O.P.; Storm-Mathisen, J.; Wolff, J.R. Heterogeneous distribution of GABA-immunoreactive nerve fibers and axon terminals in the superior cervical ganglion of adult rat. Neuroscience 1988, 26, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Wolff, J.R.; Kása, P.; Dobo, E.; Wenthold, R.J.; Joo, F. Quantitative analysis of the number and distribution of neurons richly innervated by GABA-immunoreactive axons in the rat superior cervical ganglion. J. Comp. Neurol. 1989, 282, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Hioki, H.; Nakamura, K.; Tanaka, Y.; Nakade, H.; Kaneko, T.; Iino, S.; Nojyo, Y. Gamma-aminobutyric acid-containing sympathetic preganglionic neurons in rat thoracic spinal cord send their axons to the superior cervical ganglion. J. Comp. Neurol. 2007, 502, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Elinos, D.; Rodríguez, R.; Martinez, L.A.; Zetina, M.E.; Cifuentes, F.; Morales, M.A. Segregation of acetylcholine and GABA in the rat superior cervical ganglia: Functional correlation. Front. Cell. Neurosci. 2016, 10, 91. [Google Scholar] [CrossRef]
- Merino-Jiménez, C.; Miguel, F.; Feria-Pliego, J.A.; Zetina-Rosales, M.E.; Cifuentes, F.; Morales, M.A. Sympathetic hyperactivity and age affect segregation and expression of neurotransmitters. Front. Cell. Neurosci. 2018, 12, 411. [Google Scholar] [CrossRef]
- de Groat, W.C. The actions of gamma-aminobutyric acid and related amino acids on mammalian autonomic ganglia. J. Pharmacol. Exp. Ther. 1970, 172, 384–396. [Google Scholar]
- Bowery, N.G.; Brown, D.A. γ-Aminobutyrylcholine: Actions on GABA and acetylcholine receptors. J. Pharm. Pharmacol. 1972, 24, 663–666. [Google Scholar] [CrossRef]
- González-Burgos, G.R.; Biali, F.C.; Nicola Siri, L.C.; Cardinali, D.P. Effect of gamma-aminobutyric acid on synaptic transmission and long-term potentiation in rat superior cervical ganglion. Brain Res. 1994, 658, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Martínez, L.A.; Cifuentes, F.; Morales, M.A. Ganglionic Long-Term Potentiation in Prehypertensive and Hypertensive Stages of Spontaneously Hypertensive Rats Depends on GABA Modulation. Neural Plast. 2019, 2019, 7437894. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.D.; Kriegstein, A.R. Defining the role of GABA in cortical development. J. Physiol. 2009, 587, 1873–1879. [Google Scholar] [CrossRef] [PubMed]
- Wolff, J.R.; Joo, F.; Kása, P. Modulation by GABA of neuroplasticity in the central and peripheral nervous system. Neurochem. Res. 1993, 18, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Sossin, W.S.; Sweet-Cordero, A.; Scheller, R.H. Dale’s hypothesis revised: Differential neuropeptides derived from a common precursor are targeted to different processes. Proc. Natl. Acad. Sci. USA 1990, 87, 4845–4848. [Google Scholar] [CrossRef] [PubMed]
- Sulzer, D.; Rayport, S. Dale´s princip0le and glutamate corelease from ventral midbrain dopamine neurons. Amino Acids 2000, 19, 45–52. [Google Scholar] [CrossRef]
- Zhang, S.; Qi, J.; Li, X.; Wang, H.L.; Britt, J.P.; Hoffman, A.F.; Bonci, A.; Lupica, C.R.; Morales, M. Dopaminergic and glutamatergic microdomains in a subset of rodent mesoaccumbens axons. Nat. Neurosci. 2015, 18, 386–392. [Google Scholar] [CrossRef]
- Fortin, G.M.; Ducrot, C.; Giguère, N.; Kouwenhoven, W.M.; Bourque, M.J.; Pacelli, C.; Varaschin, R.K.; Brill, M.; Singh, S.; Wiseman, P.W.; et al. Segregation of dopamine and glutamate release sites in dopamine neuron axons: Regulation by striatal target cells. FASEB J. 2019, 33, 400–417. [Google Scholar] [CrossRef]
- Agnati, L.F.; Guidolin, D.; Cervetto, C.; Maura, G.; Marcoli, M. Brain Structure and Function: Insights from Chemical Neuroanatomy. Life 2023, 13, 940. [Google Scholar] [CrossRef]
- Vega, A.; Luther, J.; Birren, S.; Morales, M. Segregation of the classical transmitters norepinephrine and acetylcholine and the neuropeptide Y in sympathetic neurons: Modulation by ciliary neurotrophic factor or prolonged growth in culture. Dev. Neurobiol. 2010, 70, 913–928. [Google Scholar] [CrossRef]
- Vega, A.; Cancino-Rodezno, A.; Valle-Leija, P.; Sánchez-Tafolla, B.; Elinos, D.; Cifuentes, F.; Morales, M.A. Neurotrophin-dependent plasticity of neurotransmitter segregation in the rat superior cervical ganglion in vivo. Dev. Neurobiol. 2016, 76, 832–846. [Google Scholar] [CrossRef] [PubMed]
- Gascoigne, D.A.; Serdyukova, N.A.; Aksenov, D.P. Early Development of the GABAergic System and the Associated Risks of Neonatal Anesthesia. Int. J. Mol. Sci. 2021, 22, 12951. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, R.; Romo-Parra, H.; Maqueda, J.; Vivar, C.; Ramírez, M.; Morales, M.A.; Lamas, M. Plasticity of the GABAergic phenotype of the “glutamatergic” granule cells of the rat dentate gyrus. J. Neurosci. 2003, 23, 5594–5598. [Google Scholar] [CrossRef] [PubMed]
- Naderipoor, P.; Amani, M.; Abedi, A.; Sakhaie, N.; Sadegzadeh, F.; Saadati, H. Alterations in the behavior, cognitive function, and BDNF level in adult male rats following neonatal blockade of GABA-A receptors. Brain Res. Bull. 2021, 169, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhan, L.; Zeng, W.; Li, K.; Sun, W.; Xu, Z.C.; Xu, E. Downregulation of hippocampal GABA after hypoxia-induced seizures in neonatal rats. Neurochem. Res. 2011, 36, 2409–2416. [Google Scholar] [CrossRef] [PubMed]
- Kilb, W. Development of the GABAergic system from birth to adolescence. Neuroscientist 2012, 18, 613–630. [Google Scholar] [CrossRef] [PubMed]
- Takla, M.; Saadeh, K.; Tse, G.; Huang, C.L.H.; Jeevaratnam, K. Ageing and the Autonomic Nervous System. In Biochemistry and Cell Biology of Ageing: Part IV, Clinical Science, Subcellular Biochemistry; Harris, J.R., Korolchuk, V.I., Eds.; Springer: Cham, Switzerland, 2023; Volume 103, pp. 201–252. [Google Scholar]
- Wolff, J.R.; Joó, F.; Dames, W.; Fehér, O. Induction and maintenance of free postsynaptic membrane thickenings in the adult superior cervical ganglion. J. Neurocytol. 1979, 8, 549–563. [Google Scholar] [CrossRef]
- Phillips, R.J.; Powley, T.L. Innervation of the gastrointestinal tract: Patterns of aging. Auton. Neurosci. 2007, 136, 1–19. [Google Scholar] [CrossRef]
- Gibson, G.E.; Peterson, C. Aging Decreases Oxidative Metabolism and the Release and Synthesis of Acetylcholine. J. Neurochem. 1981, 37, 978–984. [Google Scholar] [CrossRef]
- Smith, D.O. Reduced capabilities of synaptic transmission in aged rats. Exp. Neurol. 1979, 66, 650–666. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández, A.; González-Sierra, C.; Zetina, M.E.; Cifuentes, F.; Morales, M.A. Age-Dependent Changes in the Occurrence and Segregation of GABA and Acetylcholine in the Rat Superior Cervical Ganglia. Int. J. Mol. Sci. 2024, 25, 2588. https://doi.org/10.3390/ijms25052588
Hernández A, González-Sierra C, Zetina ME, Cifuentes F, Morales MA. Age-Dependent Changes in the Occurrence and Segregation of GABA and Acetylcholine in the Rat Superior Cervical Ganglia. International Journal of Molecular Sciences. 2024; 25(5):2588. https://doi.org/10.3390/ijms25052588
Chicago/Turabian StyleHernández, Alfredo, Constanza González-Sierra, María Elena Zetina, Fredy Cifuentes, and Miguel Angel Morales. 2024. "Age-Dependent Changes in the Occurrence and Segregation of GABA and Acetylcholine in the Rat Superior Cervical Ganglia" International Journal of Molecular Sciences 25, no. 5: 2588. https://doi.org/10.3390/ijms25052588
APA StyleHernández, A., González-Sierra, C., Zetina, M. E., Cifuentes, F., & Morales, M. A. (2024). Age-Dependent Changes in the Occurrence and Segregation of GABA and Acetylcholine in the Rat Superior Cervical Ganglia. International Journal of Molecular Sciences, 25(5), 2588. https://doi.org/10.3390/ijms25052588