
A Combination of β-Aescin and Newly Synthesized Alkylamidobetaines as Modern Components Eradicating the Biofilms of Multidrug-Resistant Clinical Strains of Candida glabrata

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Results
2.1. The Synthesis and Characterization of CnTMDAB Surfactants
2.2. Microbiological Investigation
- I.
- In the first stage, we aimed to select clinically drug-resistant strains of C. glabrata with the most vigorous intensity of biofilm production. Among the eight multidrug-resistant clinical strains of C. glabrata, the 2586 and 2853 strains were selected. Moreover, the reference strain C. glabrata ATCC 90030 was used.
- II.
- Further research evaluated the antimicrobial properties of β-aescin, newly synthesized AABs, and antifungal drugs (including the combination) (through their MIC90, MFC, and FICI values). Combinations of the tested compounds and their actions with antifungal medications were considered. An essential point of this research was determining the ability of biofilm eradication of compounds against C. glabrata strains. The anti-biofilm assays included qualitative and quantitative analyses of the impact of the combined compounds on fungal biofilms. These measurements were performed using fluorescence microscopy with computational analysis of the obtained pictures.
- III.
- The last stage was focused on the measurements of the potential toxicity of β-aescin and AABs. For this purpose, a hemolysis test and MTT assays on Balb/3T3 mouse embryonic fibroblasts were conducted to determine the usefulness of these compounds for medicine.
2.2.1. Selection of Candida glabrata Clinical Strains Based on Their Biofilm Production
2.2.2. Evaluation of the MIC and MFC of Antifungal Compounds against Selected Strains
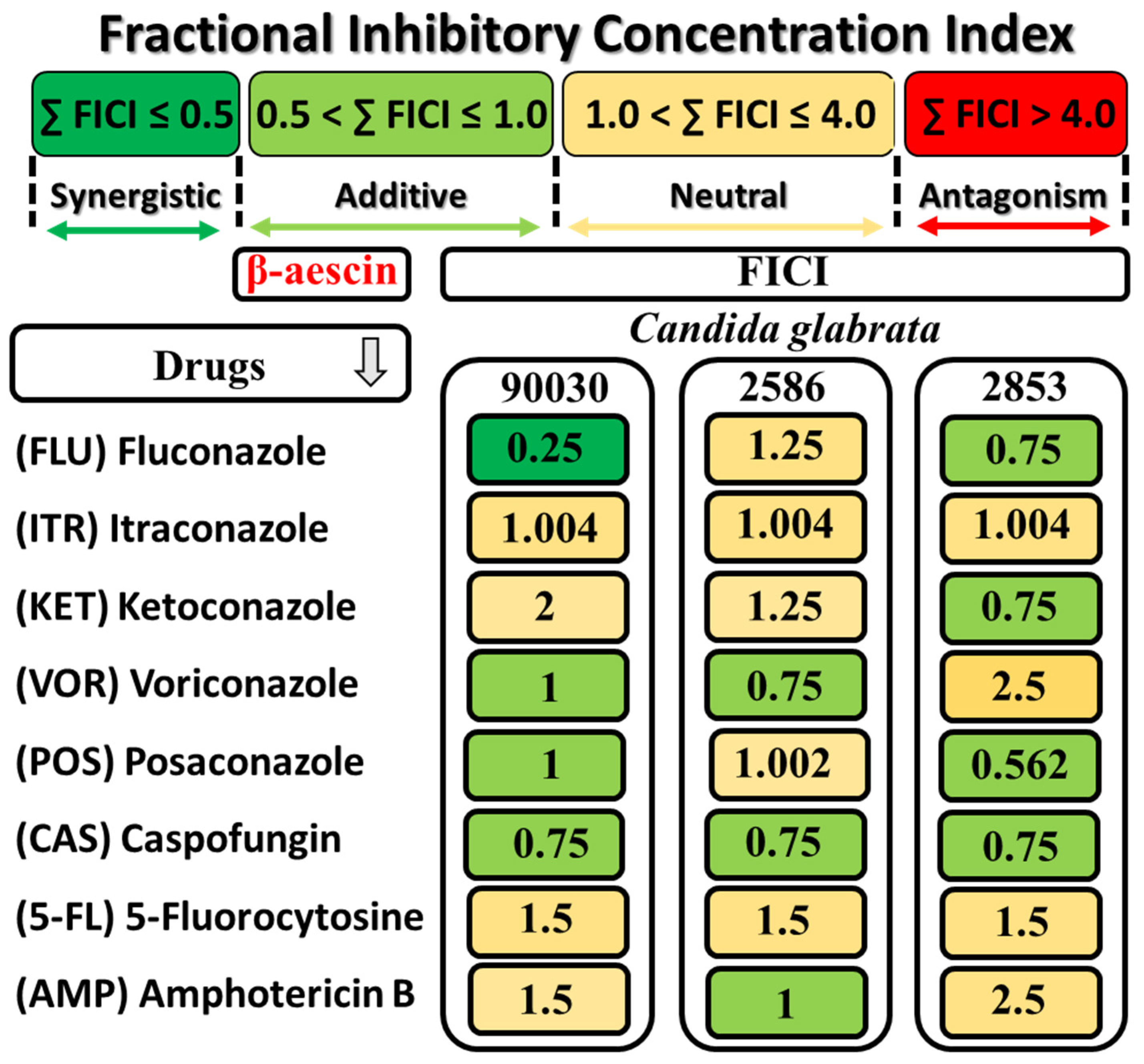
2.2.3. Evaluation of the Fractional Inhibitory Concentration Index (FICI) of β-AE and Antifungal Drugs
2.2.4. Interaction of AAB with β-AE and Selected Antifungal Agents
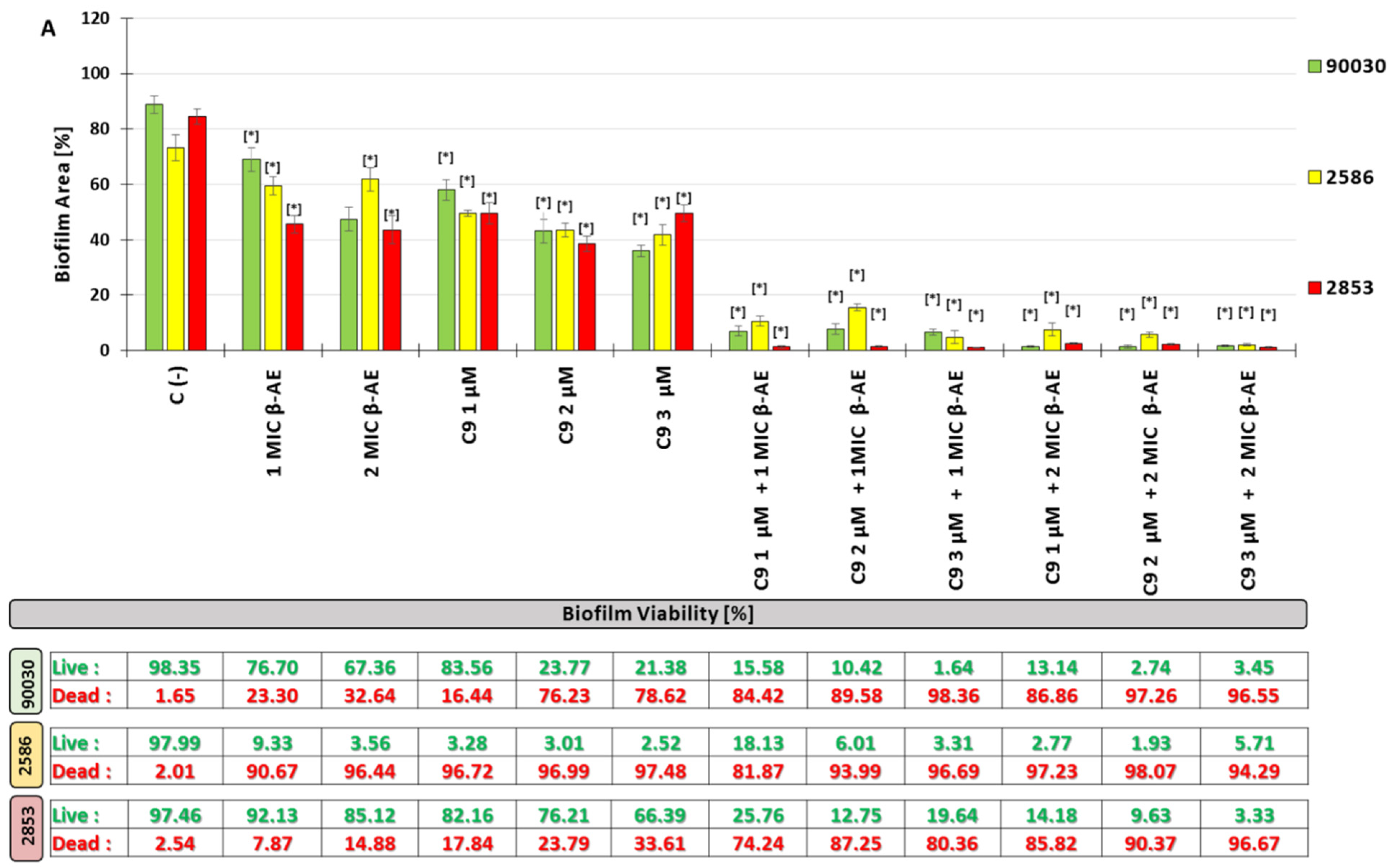
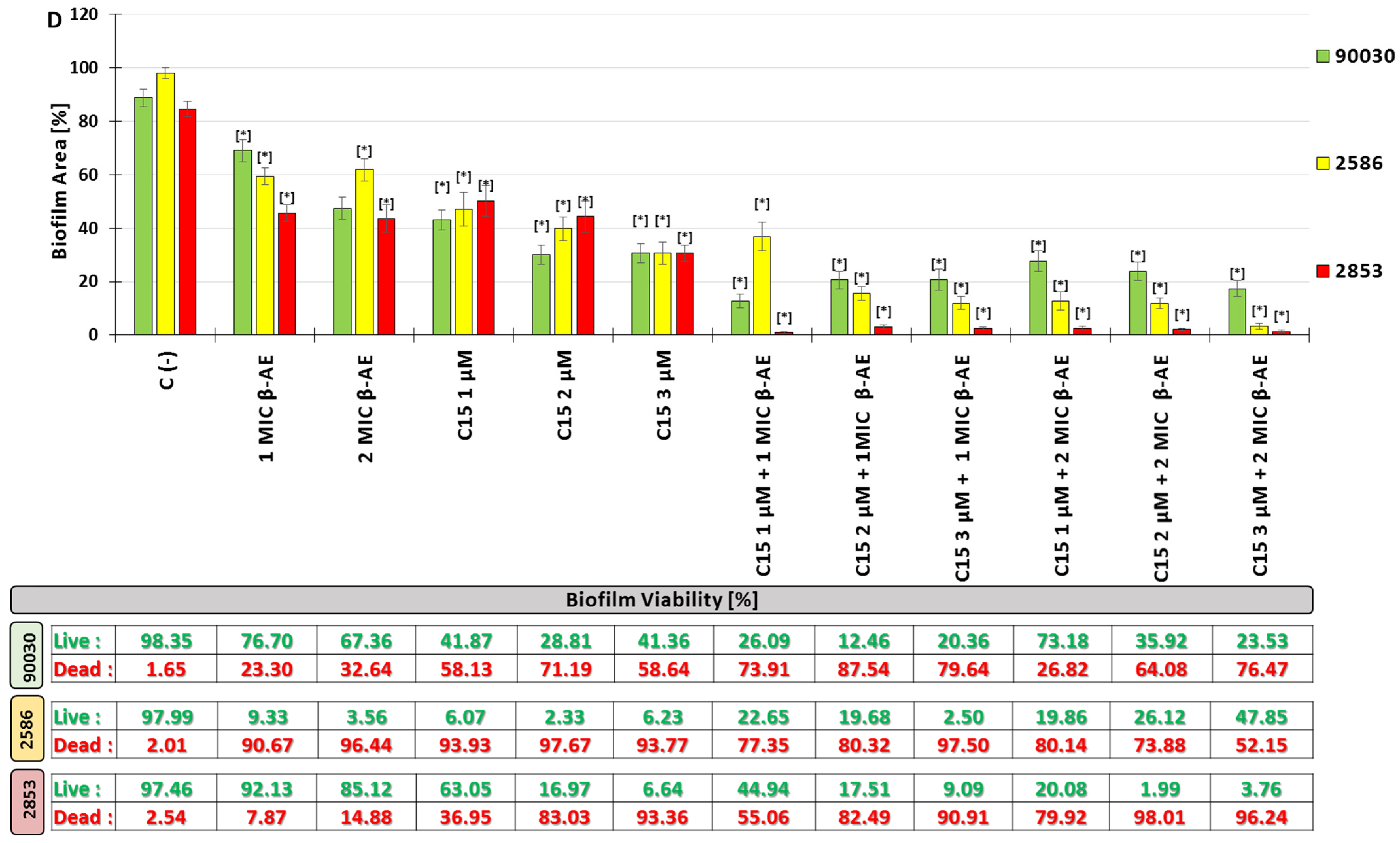
2.2.5. Influenced Combination of β-AE and Newly Synthesized AABs on Biofilm Eradication Multidrug-Resistant C. glabrata
2.2.6. Hemolysis Assays
2.2.7. Cell Proliferation Assays Balb/3T3 Mouse Embryonic Fibroblast (MTT Test)
3. Discussion
4. Materials and Methods
4.1. Compounds
- C9H19 molar mass: 328.49 (g/mol);
- C11H23 molar mass: 356.54 (g/mol);
- C13H27 molar mass: 384.60 (g/mol);
- C15H31 molar mass: 412.65 (g/mol).
- β-aescin (C55H86O24), CAS no.: 11072-93-8 (Sigma-Aldrich, Oakville, ON, Canada);
- Fluconazole (C10H8N6), CAS no.: 514222-44-7 (Glentham Life Sciences, Corsham, UK);
- Itraconazole (C35H38Cl2N8O4), CAS no.: 84625-61-6 (Glentham Life Sciences, Corsham, UK);
- Ketoconazole (C26H28Cl2N4O4), CAS no.: 65277-42-1 (Glentham Life Sciences, Corsham, UK);
- Voriconazole (C16H14F3N5O), CAS no.: 137234-62-9 (Glentham Life Sciences, Corsham, UK);
- Posaconazole (C37H42F2N8O4), CAS no.: 171228-49-2 (Glentham Life Sciences, Corsham, UK);
- Caspofungin (C52H88N10O15), CAS no.: 179463-17-3 (Glentham Life Sciences, Corsham, UK);
- 5-Fluorocytosine (C4H4FN3O), CAS no.: 2022-85-7 (Glentham Life Sciences, Corsham, UK);
- Amphotericin B (C47H73NO17), CAS no.: 1397-89-3 (Glentham Life Sciences, Corsham, UK).
4.2. The Preparation and Chemical Characterization of AAB
4.2.1. Materials
4.2.2. The synthesis of N-[3-(dimethylamine)propyl]-N-methylalkylamides
4.2.3. The Synthesis of CnTMDAB
4.2.4. The Characterization of Betaine-Type Surfactants
4.3. Strains and Growth Condition
4.4. Assessment of Biofilm Production Ability
4.5. Minimal Inhibitory and Fungicidal Concentrations
4.6. Fractional Inhibitory Concentration Index (FICI) of β-Aescin and Antifungal Compounds
4.7. Interaction of the AABs with the Selected Antifungal Agents
4.8. Biofilm Eradication of Multidrug-Resistant C. glabrata Strains on Polystyrene Surfaces by β-Aescin and AAB Combination
4.9. Hemolysis Assay
4.10. Fibroblast Cell Culture
4.11. Cell Proliferation Assays (MTT Test)
4.12. Fluorescence Microscopy (FM) and Computational Analysis of Pictures
4.13. Statistical Analysis
5. Conclusions
- AAB compounds have the potential to be used as adjuvant compounds due to their demonstrated synergy and additive effects with selected antifungal drugs.
- The combination of β-AE with AABs is a highly effective mixture for eradicating C. glabrata biofilms.
- Due to its low cytotoxicity and hemolytic potential, the AAB C9 may be used in practice after further clinical trials.
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lohse, M.B.; Gulati, M.; Johnson, A.D.; Nobile, C.J. Development and regulation of single and multi-species Candida albicans biofilms. Nat. Rev. Microbiol. 2018, 16, 19–31. [Google Scholar] [CrossRef]
- Malinovska, Z.; Conkova, E.; Vaczi, P. Biofilm formation in Medically important Candida species. J. Fungi 2023, 9, 955. [Google Scholar] [CrossRef]
- Maccallum, D.M. Hosting infections: Experimental models to assay Candida virulence. Int. J. Microbiol. 2012, 2012, 363764. [Google Scholar] [CrossRef] [PubMed]
- Pohl, C.H. Recent advances and opportunities in the study of Candida albicans polymicrobial biofilms. Front. Cell. Infect. Microbiol. 2022, 12, 836379. [Google Scholar] [CrossRef] [PubMed]
- Ponde, N.O.; Lortal, L.; Ramage, G.; Naglik, J.R.; Richardson, J.P. Candida albicans biofilms and polymicrobial interactions. Crit. Rev. Microbiol. 2021, 47, 91–111. [Google Scholar] [CrossRef] [PubMed]
- Marak, M.B.; Dhanashree, B. Antifungal susceptibility and biofilm production of Candida spp. isolated from clinical samples. Int. J. Microbiol. 2018, 2018, 7495218. [Google Scholar] [CrossRef] [PubMed]
- Nami, S.; Aghebati-Maleki, A.; Morovati, H.; Aghebati-Maleki, L. Current antifungal drugs and immunotherapeutic approaches as promising strategies to treatment of fungal diseases. Biomed. Pharmacother. 2019, 110, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Franiczek, R.; Gleńsk, M.; Krzyżanowska, B.; Włodarczyk, M. β-Aescin as subinhibitory concentration (sub-MIC) enhances susceptibility of Candida glabrata clinical isolates to nystatin. Med. Mycol. 2015, 53, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Ciurea, C.N.; Kosovski, L.B.; Mare, A.D.; Toma, F.; Pintea-Simon, I.A.; Man, A. Candida and candidiasis-opportunism versus pathogenicity: A review of the virulence traits. Microorganisms 2020, 8, 857. [Google Scholar] [CrossRef] [PubMed]
- Hachem, R.; Hanna, H.; Kontoyiannis, D.; Jiang, Y.; Raad, I. The changing epidemiology of invasive candidiases: Candida glabrata and Candida krusei as the leading causes of candidemia in hematologic malignancy. Cancer 2008, 112, 2493–2499. [Google Scholar] [CrossRef]
- Oberoi, J.K.; Wattal, C.; Goel, N.; Raveendran, R.; Datta, S.; Prasad, K. Non-albicans Candida species in bloodstream infections in a tertiary care hospital in New Delhi, India. Indian. J. Med. Res. 2012, 136, 997–1003. [Google Scholar]
- Maione, A.; Imparato, M.; Galdiero, M.; Alteris, E.D.; Feola, A.; Galdiero, E.; Guida, M. Effect of escin alone or in combination with antifungal agents on resistant Candida glabrata biofilms: Mechanisms of action. Antibiotics 2023, 12, 1210. [Google Scholar] [CrossRef] [PubMed]
- Asmundsdottir, L.R.; Erlendsdottir, H.; Gottfredsson, M. National study of candidemia, antifungal use, and antifungal drug resistance in Iceland, 2000 to 2011. J. Clin. Microbiol. 2013, 51, 841–848. [Google Scholar] [CrossRef]
- Bodey, G.P.; Mardani, M.; Hanna, H.A.; Boktour, M.; Abbas, J.; Girgawy, E.; Hachem, R.Y.; Kontoyiannis, D.P.; Raad, I.I. The epidemiology of Candida albicans and Candida glabrata fungemia in immunocompromised patients with cancer. Am. J. Med. 2002, 112, 380–385. [Google Scholar] [CrossRef]
- Frías-De-León, M.G.; Hernández-Castro, R.; Conde-Cuevas, E.; García-Coronel, I.H.; Vázquez-Aceituno, V.A.; Soriano-Ursúa, M.A.; Farfán-García, E.D.; Ocharán-Hernández, E.; Rodríguez-Cerdeira, C.; Arenas, R.; et al. Candida glabrata antifungal resistance and virulence factors, a perfect pathogenic combination. Pharmaceutics 2021, 13, 1529. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.F.; Silva, S.; Henriques, M. Candida glabrata: A review of its features and resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 673–688. [Google Scholar] [CrossRef]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.W.; Azeredo, J. Candida glabrata, Candida parapsilosis, and Candida tropicalis: Biology, epidemiology, pathogenicity and antifungal resistance. FEMS Microb. Rev. 2012, 36, 288–305. [Google Scholar] [CrossRef]
- Biernasiuk, A.; Malm, A. Synergistic interactions between linalool and some antimycotic agents against Candida spp. as a basis for developing new antifungal preparations. Appl. Sci. 2023, 13, 5686. [Google Scholar] [CrossRef]
- Mroczyńska, M.; Brillowska-Dabrowska, A. Virulence of clinical Candida isolates. Pathogens 2021, 10, 466. [Google Scholar] [CrossRef]
- Kaur, J.; Nobile, C.J. Antifungal drug-resistance mechanisms in Candida biofilms. Curr. Opin. Microbiol. 2023, 71, 102237. [Google Scholar] [CrossRef]
- Römling, U.; Balsalobre, C. Biofilm infections, their resilience to therapy and innovative treatment strategies. J. Intern. Med. 2012, 272, 541–561. [Google Scholar] [CrossRef] [PubMed]
- Junqueira, J.C.; Mylonakis, E. Candida biofilms. Front. Microbiol. 2023, 13, 1128600. [Google Scholar] [CrossRef] [PubMed]
- Arendrup, M.C.; Patterson, T.F. Multidrug-resistant Candida: Epidemiology, molecular mechanisms, and treatment. J. Infect. Dis. 2017, 216, S445–S451. [Google Scholar] [CrossRef]
- Lu, L.; Hu, W.; Tian, Z.; Yuan, D.; Yi, G.; Zhou, Y.; Cheng, Q.; Zhu, J.; Li, M. Developing natural products as potential anti-biofilm agents. Chin. Med. 2019, 14, 11. [Google Scholar] [CrossRef]
- Sirtori, C.R. Aescin: Pharmacology, pharmacokinetics and therapeutic profile. Pharmacol. Res. 2001, 44, 183–193. [Google Scholar] [CrossRef]
- Hostettmann, K.; Marston, A. Saponins; Cambridge University Press: Cambridge, UK; Cambridge University Press: New York, NY, USA, 1995. [Google Scholar]
- Sparg, S.G.; Light, M.E.; van Staden, J. Biological activities and distribution of plant saponins. J. Ethnopharmacol. 2007, 94, 219–243. [Google Scholar] [CrossRef]
- Netala, V.R.; Ghosh, S.B.; Bobbu, P.; Anitha, D.; Tartte, V. Triterpenoid saponins: A review. Food Res. Int. 2014, 59, 16–40. [Google Scholar]
- Geisler, R.; Dargel, C.; Hellweg, T. The biosurfactant β-aescin: A review on the physicochemical properties and its interaction with lipid model membranes and Langmuir monolayers. Molecules 2020, 25, 117. [Google Scholar] [CrossRef]
- Clendennen, S.K.; Boaz, N.W. Chapter 14—Betaine Amphoteric Surfactants—Synthesis, properties and applications. In Biobased Surfactants, 2nd ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 447–469. [Google Scholar]
- Alzobaidi, S.; Da, C.; Tran, V.; Prodanovic, M.; Johnson, K.P. High-temperature ultralow water content carbon dioxide-in-water foam stabilized with viscoelastic zwitterionic surfactants. J. Colloid. Interface Sci. 2017, 488, 79–91. [Google Scholar] [CrossRef]
- Dey, J.; Ghosh, R.; Mahapatra, R.D. Self-Assembly of Unconventional Low-Molecular-Mass Amphiphiles Containing a PEG Chain. Langmuir 2019, 35, 848–861. [Google Scholar] [CrossRef] [PubMed]
- Lamch, Ł.; Witek, K.; Jarek, E.; Obłąk, E.; Warszyński, P.; Wilk, K.A. New mild amphoteric sulfohydroxybetaine-type surfactants containing different labile spacers: Synthesis, surface properties, and performance. J. Colloid. Interface Sci. 2020, 558, 220–229. [Google Scholar] [CrossRef]
- Domingo, X. Amphoteric Surfactants; Lomax, E.G., Ed.; Marcel Dekker: New York, NY, USA, 1996; p. 567. [Google Scholar]
- Liu, H.; Du, Y.; Yang, J.; Zhu, H. Structural characterization and antimicrobial activity of chitosan/betaine derivative complex. Carbohydr. Polym. 2003, 55, 291–297. [Google Scholar] [CrossRef]
- Robbins, N.; Caplan, T.; Cowen, L.E. Molecular evolution of antifungal drug resistance. Annu. Rev. Microbiol. 2017, 71, 735–775. [Google Scholar] [CrossRef] [PubMed]
- Nikoomanesh, F.; Falahatinejad, M.; Cernakova, L.; Dos Santos, A.L.S.; Mohammadi, S.R.; Rafiee, M.; Rodrigues, C.F.; Roudbary, M. Combination of farnesol with common antifungal drugs: Inhibitory effect against Candida species isolated from women with RVVC. Medicina 2023, 59, 743. [Google Scholar] [CrossRef]
- Vallieres, C.; Singh, N.; Alexander, C.; Avery, S.V. Repurposing nonantifungal approved drugs for synergistic targeting of fungal pathogens. ACS Infect. Dis. 2020, 6, 2950–2958. [Google Scholar] [CrossRef] [PubMed]
- Paluch, E.; Okińczyc, P.; Zwyrzykowska-Wodzińska, A.; Szperlik, J.; Żarowska, B.; Duda-Madej, A.; Bąbelewski, P.; Włodarczyk, M.; Wojtasik, W.; Kupczyński, R.; et al. Composition and antimicrobial activity of Ilex leaves water extracts. Molecules 2021, 26, 7442. [Google Scholar] [CrossRef]
- Okińczyc, P.; Paluch, E.; Franiczek, R.; Widelski, J.; Wojtanowski, K.K.; Mroczek, T.; Krzyżanowska, B.; Skalicka-Woźniak, K.; Sroka, Z. Antimicrobial Activity of Apis mellifera L. and Trigona sp. Propolis from Nepal and Its Phytochemical Analysis. Biomed. Pharmacother. 2020, 129, 110435. [Google Scholar] [CrossRef] [PubMed]
- Potter, D.A.; Kimmerer, T.W. Inhibition of herbivory on young holly leaves: Evidence for the defensive role of saponins. Oecologia 1989, 78, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Zhu, N.; Ze-Ren-Wang-Mu, N.; Shen, Y. Two new antifungal saponins from the Tibetian herbal medicine Clematis tangutica. Planta Med. 2003, 69, 547–551. [Google Scholar] [PubMed]
- Zhang, J.D.; Cao, Y.B.; Xu, Z.; Sun, H.H.; An, M.M.; Yan, L.; Chen, H.S.; Gao, P.H.; Wang, Y.; Jia, X.M.; et al. In vitro and in vivo antifungal activities of eight steroid saponins from Tribulus terrestris L. with potent activity against fluconazole-resistant fungal pathogens. Biol. Pharm. Bull. 2005, 28, 2211–2215. [Google Scholar] [CrossRef]
- Ngo-Mback, M.; Babii, C.; Dongmo, P.J.; Toghueo, M.K.; Stefan, M.; Boyom, F.F. Anticandidal and synergistic effect of essential oil fraction from three aromatic plants used in Cameroon. J. Mycol. Med. 2020, 30, 100940. [Google Scholar] [CrossRef] [PubMed]
- Rosato, A.; Altini, E.; Sblano, S.; Salvagno, L.; Maggi, F.; de Michele, G.; Carocci, A.; Clodoveo, M.L.; Corbo, F.; Fracchiolla, G. Synergistic activity of new diclofenac and essential oils combinations against different Candida spp. Antibiotics 2021, 10, 688. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, H. Membrane interactions of phytochemicals as their molecular mechanism applicable to the discovery of drug leads from plants. Molecules 2015, 20, 18923–18966. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, Y.; Liu, X.; Wang, J.; Wei, L.; Feng, Y. Effect of a hydrophilic head group on Krafft temperature, surface activities and rheological behaviors of erucyl amidobetaines. J. Surface. Detergent. 2014, 17, 295–301. [Google Scholar] [CrossRef]
- Linfield, W.M. Soap and lime soap dispersants. J. Am. Oil Chem. Soc. 1978, 55, 87–91. [Google Scholar] [CrossRef]
- Sheehan, D.J.; Hitchcock, C.A.; Sibley, C.M. Current and emerging azole antifungal agents. Clin. Microbiol. Rev. 1999, 12, 40–79. [Google Scholar] [CrossRef]
- Allen, D.; Wilson, D.; Drew, R.; Perfect, J. Azole antifungals: 35 years of invasive fungal infection management. Expert. Rev. Anti Infect. Ther. 2015, 13, 787–798. [Google Scholar] [CrossRef]
- Paluch, E.; Piecuch, A.; Obłąk, E.; Lamch; Wilk, K.A. Antifungal Activity of Newly Synthesized Chemodegradable Dicephalic-Type Cationic Surfactants. Colloids Surf. B Biointerfaces 2018, 164, 34–41. [Google Scholar] [CrossRef]
- Piętka-Ottlik, M.; Frąckowiak, R.; Maliszewska, I.; Kołwzan, B.; Wilk, K.A. Ecotoxicity and biodegradability dicephalic-type cationic surfactants. Colloid. Surf. B Biointerfaces 2018, 89, 1103–1111. [Google Scholar]
- Piecuch, A.; Lamch, Ł.; Paluch, E.; Obłąk, E.; Wilk, K.A. Biofilm prevention by dicephalic cationic surfactants and their interaction with DNA. J. Appl. Microbiol. 2016, 121, 682–692. [Google Scholar] [CrossRef]
- Bujdáková, H.; Didiášová, M.; Drahovská, H.; Černáková, L. Role of cell surface hydrophobicity in Candida albicans biofilm. Cent. Eur. J. Biol. 2013, 8, 259–262. [Google Scholar] [CrossRef]
- Obłąk, E.; Piecuch, A.; Krasowska, A.; Łuczyński, J. Antifungal gemini quaternary ammonium salts. Microbiol. Res. 2013, 168, 630–638. [Google Scholar] [CrossRef]
- Almeida, J.A.S.; Faneca, H.; Carvalho, R.A.; Marques, E.F.; Pais, A.A.C.C. Dicationic Alkylammonium Bromide Gemini Surfactants. Membrane Perturbation and Skin Irritation. PLoS ONE 2011, 6, e26965. [Google Scholar] [CrossRef] [PubMed]
- The European Committee on Antimicrobial Susceptibility Testing—EUCAST. Available online: www.eucast.org (accessed on 1 November 2023).
- Paluch, E.; Szperlik, J.; Lamch, Ł.; Wilk, K.A.; Obłąk, E. Biofilm Eradication and Antifungal Mechanism of Action against Candida albicans of Cationic Dicephalic Surfactants with a Labile Linker. Sci. Rep. 2021, 11, 8896. [Google Scholar] [CrossRef] [PubMed]
- Sæbø, I.P.; Bjørås, M.; Franzyk, H.; Helgesen, E.; Booth, J.A. Optimization of the Hemolysis Assay for the Assessment of Cytotoxicity. Int.J. Mol. Sci. 2023, 24, 2914. [Google Scholar] [CrossRef] [PubMed]
- Paluch, E.; Sobierajska, P.; Okinczyc, P.; Widelski, J.; Duda-Madej, A.; Krzyzanowska, B.; Krzyzek, P.; Ogorek, R.; Szperlik, J.; Chmielowiec, J.; et al. Nanoapatites Doped and Co-Doped with Noble Metal Ions as Modern Antibiofilm Materials for Biomedical Applications against Drug-Resistant Clinical Strains of Enterococcus faecalis VRE and Staphylococcus aureus MRSA. Int. J. Mol. Sci. 2022, 23, 1533. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical structure | R | Molecular Weight (g/mol) | 1H NMR, CDCl3 | ESI-MS (MNa+) | Elemental Analyses (Calculated) | Melting Point (°C) | TK (°C) | CMC (mol/dm3) | Abbreviation | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| δ (ppm) | %C | %H | %N | ||||||||
 | C9H19 | 328.49 | 0.84–0.91 [t, 3H, -COCH2CH2(CH2)kCH3]; 1.22–1.34 [m, 2kH, -COCH2CH2(CH2)kCH3]; 1.57–1.60 [m, 2H, -COCH2CH2(CH2)kCH3]; 1.99 [m, 2H, -N+CH2CH2CH2N-]; 2.28–2.31 [t, 2H, -COCH2CH2(CH2)kCH3]; 3.02–3.05 [s, 3H, -N(CH3)-]; 3.34–3.45 [s, 6H; -N+(CH3)2-]; 3.63–3.67 [m, 4H, -N+CH2CH2CH2N-]; 3.99–4.03 [s, 2H, -N+CH2COO−]. | 351.5 | 65.79 (65.81) | 11.10 (11.07) | 8.52 (8.53) | - | <0 | 3.0 × 10−2 | C9 |
| C11H23 | 356.54 | 379.5 | 67.31 (67.37) | 11.30 (11.33) | 7.91 (7.86) | 45–47 | ~5 | 8.7 × 10−3 | C11 | ||
| C13H27 | 384.60 | 407.6 | 68.65 (68.70) | 11.52 (11.55) | 7.26 (7.29) | 58–59 | 20.5 | 2.5 × 10−3 | C13 | ||
| C15H31 | 412.65 | 435.6 | 69.84 (69.85) | 11.77 (11.75) | 6.78 (6.79) | 70–71 | 36.4 | 7.4 × 10−3 | C15 | ||
| R | Mw (g/mol) | Abbreviation | ATCC 90030 | 2586 | 2853 |
|---|---|---|---|---|---|
| MIC90 | MIC90 | MIC90 | |||
| C9H19 | 328.49 | C9 | 3 | 3 | 1 |
| C11H23 | 356.54 | C11 | >3 | >3 | >3 |
| C13H27 | 384.60 | C13 | >3 | >3 | >3 |
| C15H31 | 412.65 | C15 | >3 | >3 | >3 |
| Abb. | Compound | ATCC 90030 | 2586 | 2853 | |||
|---|---|---|---|---|---|---|---|
| MIC90 | MFC | MIC90 | MFC | MIC90 | MFC | ||
| β-AE | β-aescin | 128 | 256 | 256 | 256 | 128 | 128 |
| FLU | Fluconazole | 2048 | 2048 | 2048 | 2048 | 32 | 2048 |
| ITR | Itraconazole | 4096 | 4096 | 4096 | 4096 | 4096 | 4096 |
| KET | Ketoconazole | 256 | 1024 | 512 | 1024 | 4 | 1024 |
| VOR | Voriconazole | 512 | 1024 | 512 | 1024 | 2 | 512 |
| POS | Posaconazole | 256 | 1024 | 512 | 1024 | 1 | 256 |
| CAS | Caspofungin | 0.06 | 0.06 | 0.03 | 0.06 | 0.03 | 0.06 |
| 5-FL | 5-Fluorocytosine | 512 | 1024 | 512 | 1024 | 256 | 512 |
| AMP | Amphotericin B | 0.25 | 0.5 | 0.25 | 1 | 0.25 | 0.5 |
| Abb. | Compound | ATCC 90030 | 2586 | 2853 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C9 | C11 | C13 | C15 | C9 | C11 | C13 | C15 | C9 | C11 | C13 | C15 | ||
| β-AE | β-aescin | 128 (1) | 128 (1) | 128 (1) | 128 (1) | 128 (2) | 128 (2) | 128 (2) | 128 (2) | 128 (1) | 128 (1) | 128 (1) | 128 (1) |
| FLU | Fluconazole | 128 (16) | 32 (64) | 32 (64) | 32 (64) | 256 (8) | 32 (64) | 32 (64) | 32 (64) | 32 (1) | 16 (2) | 16 (2) | 2 (16) |
| ITR | Itraconazole | 4096 (1) | 4096 (1) | 4096 (1) | 4096 (1) | 4096 (1) | 4096 (1) | 4096 (1) | 4096 (1) | 4096 (1) | 4096 (1) | 4096 (1) | 4096 (1) |
| KET | Ketoconazole | 16 (16) | 16 (16) | 16 (16) | 4 (64) | 64 (8) | 32 (16) | 16 (32) | 8 (64) | 4 (1) | 4 (1) | 2 (2) | 1 (4) |
| VOR | Voriconazole | 8 (64) | 8 (64) | 8 (64) | 8 (64) | 64 (8) | 8 (64) | 8 (64) | 8 (64) | 2 (1) | 1 (2) | 1 (2) | 256 (128) |
| POS | Posaconazole | 512 (2) | 32 (8) | 32 (8) | 512 (2) | 512 (1) | 256 (2) | 256 (2) | 512 (1) | 8 (8) | 4 (4) | 4 (4) | 256 (256) |
| CAS | Caspofungin | 0.06 (1) | 0.06 (1) | 0.06 (1) | 0.03 (1) | 0.03 (1) | 0.03 (1) | 0.03 (1) | 0.03 (1) | 0.03 (1) | 0.03 (1) | 0.03 (1) | 0.03 (1) |
| 5-FL | 5-Fluorocytosine | 512 (1) | 512 (1) | 512 (1) | 512 (1) | 512 (1) | 512 (1) | 512 (1) | 512 (1) | 256 (1) | 256 (1) | 256 (1) | 256 (1) |
| AMP | Amphotericin B | 0.25 (1) | 0.12 (2) | 0.12 (2) | 0.06 (4) | 0.12 (2) | 0.12 (2) | 0.12 (2) | 0.06 (4) | 0.25 (1) | 0.12 (2) | 0.12 (2) | 0.06 (4) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paluch, E.; Bortkiewicz, O.; Widelski, J.; Duda-Madej, A.; Gleńsk, M.; Nawrot, U.; Lamch, Ł.; Długowska, D.; Sobieszczańska, B.; Wilk, K.A. A Combination of β-Aescin and Newly Synthesized Alkylamidobetaines as Modern Components Eradicating the Biofilms of Multidrug-Resistant Clinical Strains of Candida glabrata. Int. J. Mol. Sci. 2024, 25, 2541. https://doi.org/10.3390/ijms25052541
Paluch E, Bortkiewicz O, Widelski J, Duda-Madej A, Gleńsk M, Nawrot U, Lamch Ł, Długowska D, Sobieszczańska B, Wilk KA. A Combination of β-Aescin and Newly Synthesized Alkylamidobetaines as Modern Components Eradicating the Biofilms of Multidrug-Resistant Clinical Strains of Candida glabrata. International Journal of Molecular Sciences. 2024; 25(5):2541. https://doi.org/10.3390/ijms25052541
Chicago/Turabian StylePaluch, Emil, Olga Bortkiewicz, Jarosław Widelski, Anna Duda-Madej, Michał Gleńsk, Urszula Nawrot, Łukasz Lamch, Daria Długowska, Beata Sobieszczańska, and Kazimiera A. Wilk. 2024. "A Combination of β-Aescin and Newly Synthesized Alkylamidobetaines as Modern Components Eradicating the Biofilms of Multidrug-Resistant Clinical Strains of Candida glabrata" International Journal of Molecular Sciences 25, no. 5: 2541. https://doi.org/10.3390/ijms25052541
APA StylePaluch, E., Bortkiewicz, O., Widelski, J., Duda-Madej, A., Gleńsk, M., Nawrot, U., Lamch, Ł., Długowska, D., Sobieszczańska, B., & Wilk, K. A. (2024). A Combination of β-Aescin and Newly Synthesized Alkylamidobetaines as Modern Components Eradicating the Biofilms of Multidrug-Resistant Clinical Strains of Candida glabrata. International Journal of Molecular Sciences, 25(5), 2541. https://doi.org/10.3390/ijms25052541