
Effect of Short Photoperiod on Behavior and Brain Plasticity in Mice Differing in Predisposition to Catalepsy: The Role of BDNF and Serotonin System


Abstract
1. Introduction
2. Results
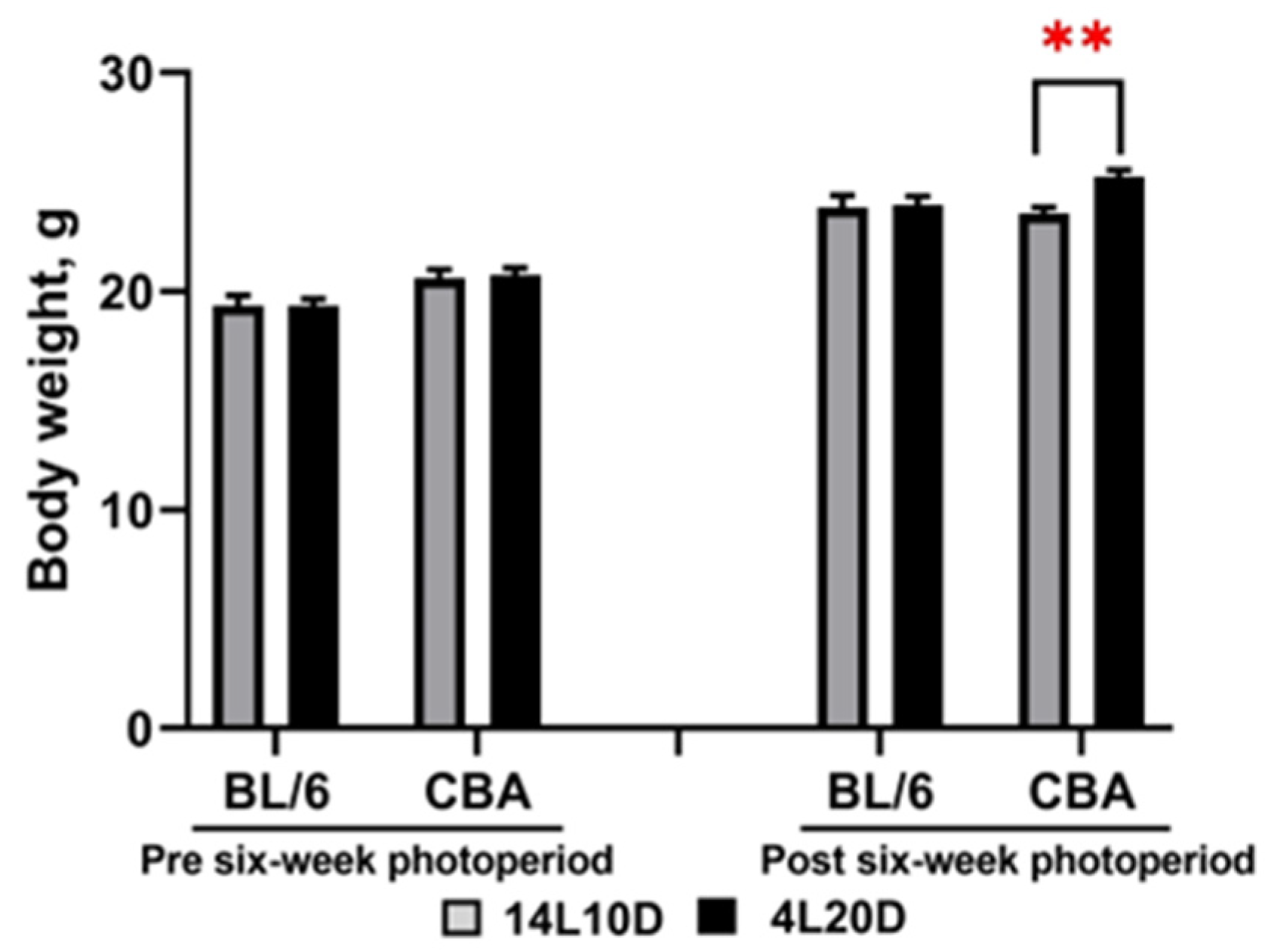
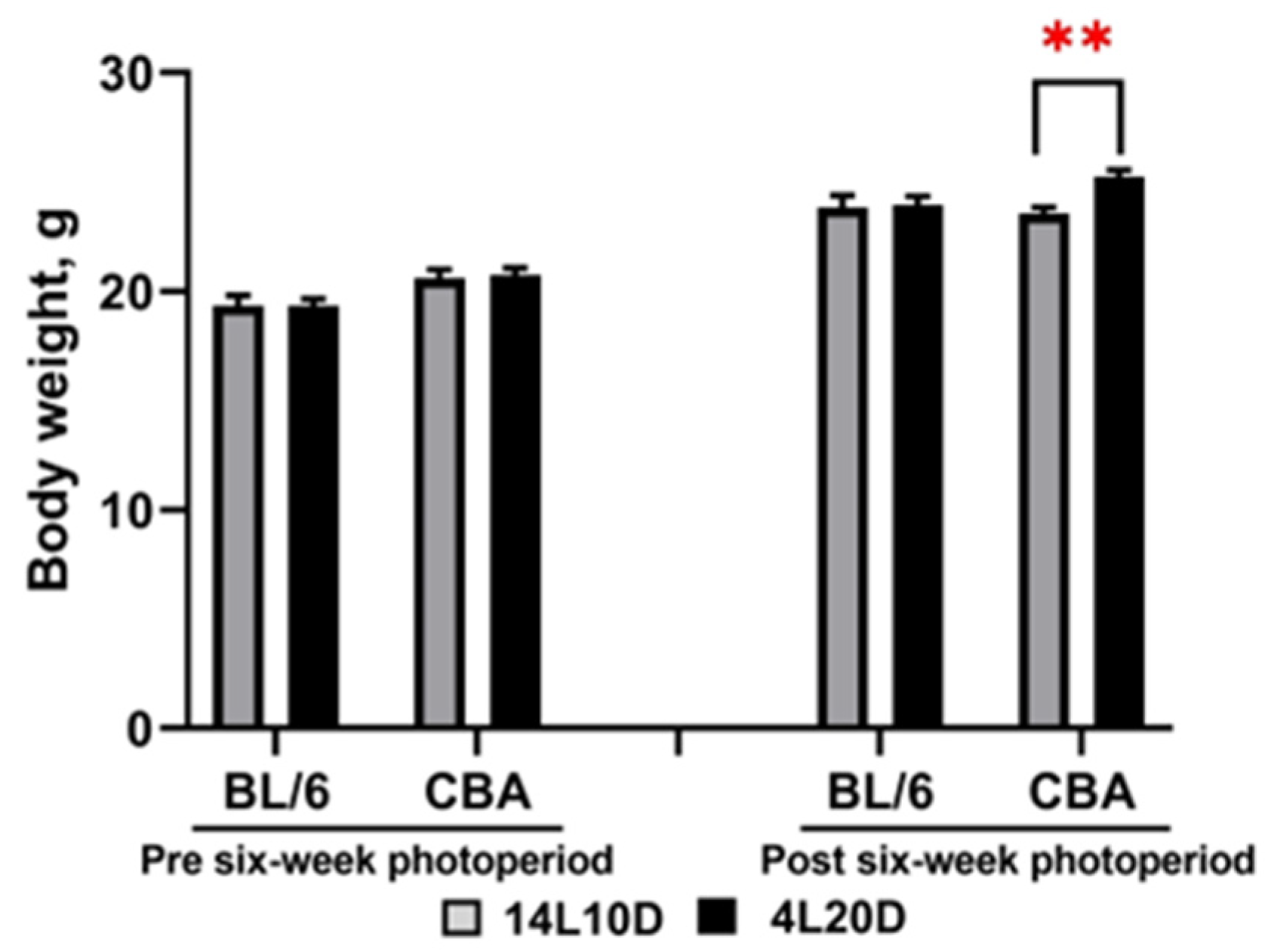
2.1. Effects of Short Photoperiod on Body Weight
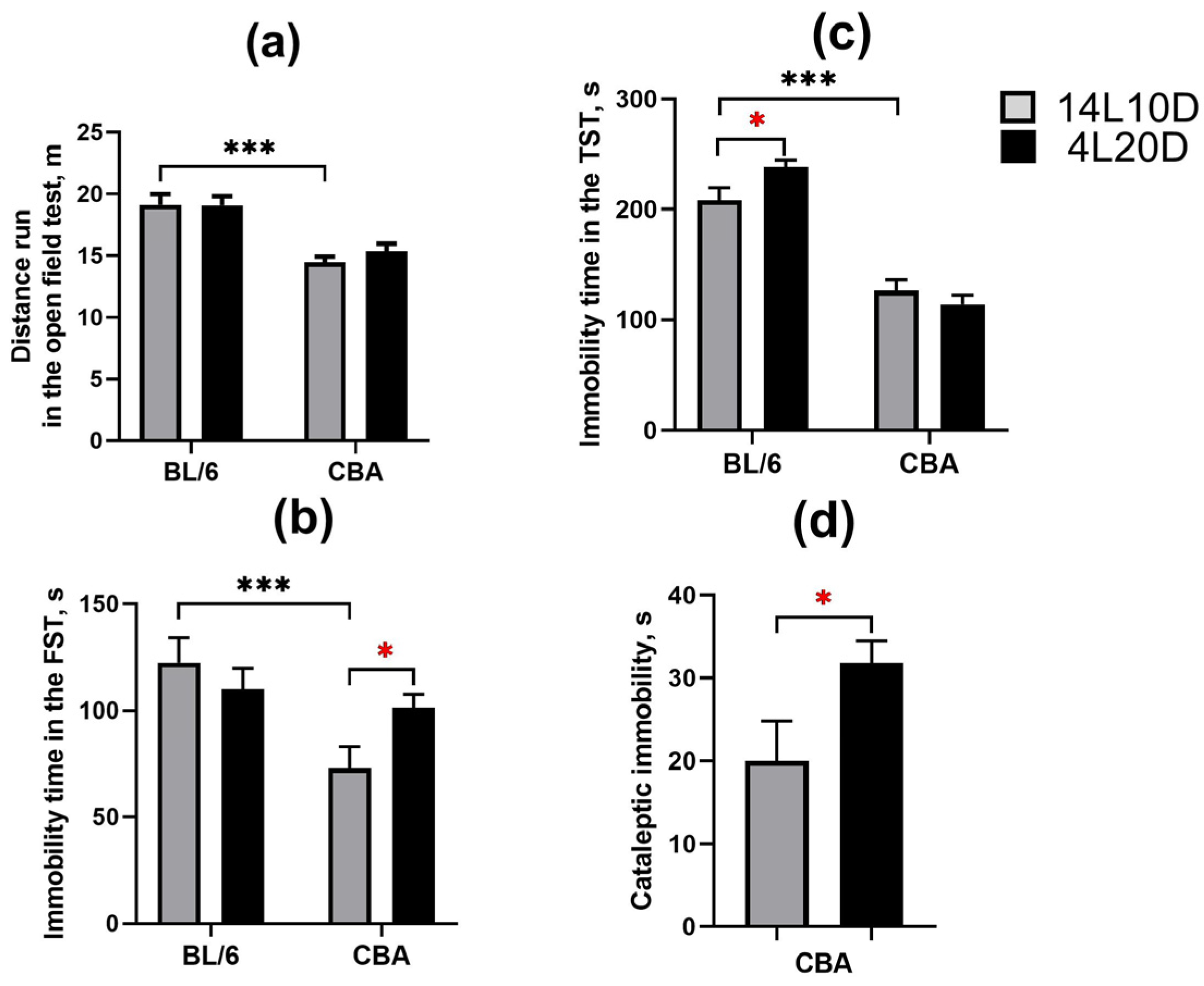
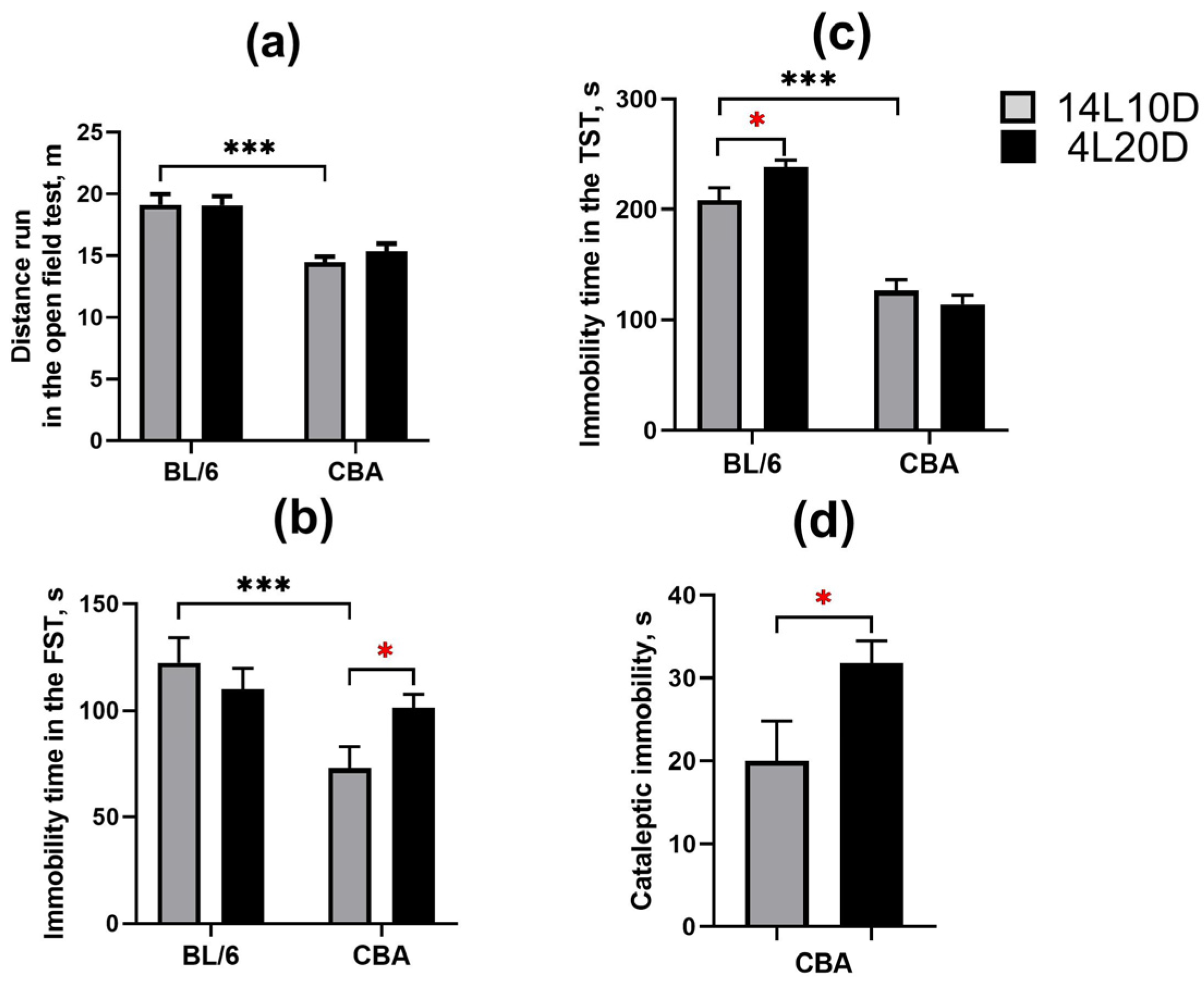
2.2. Effects of Short Photoperiod on Behavior
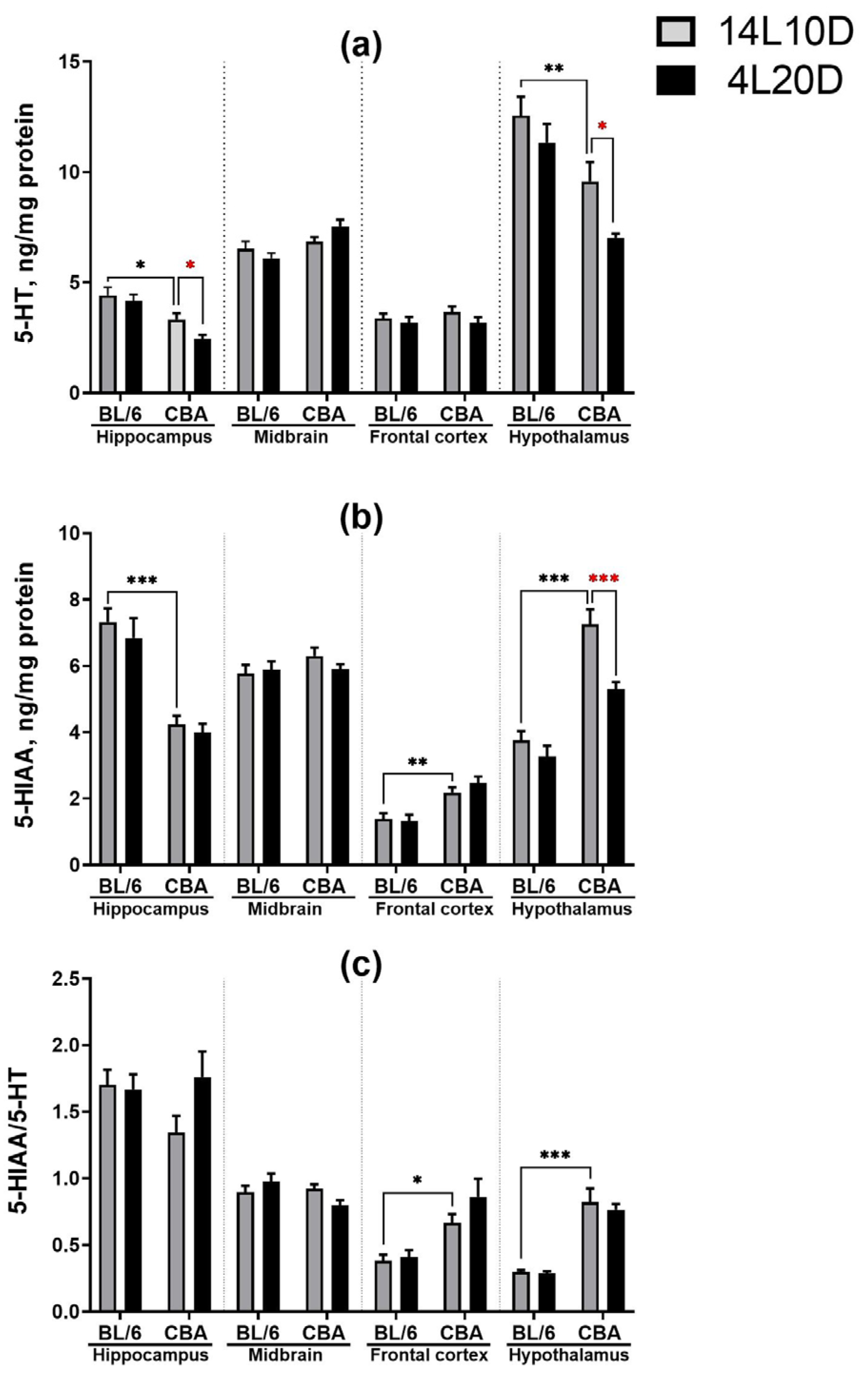
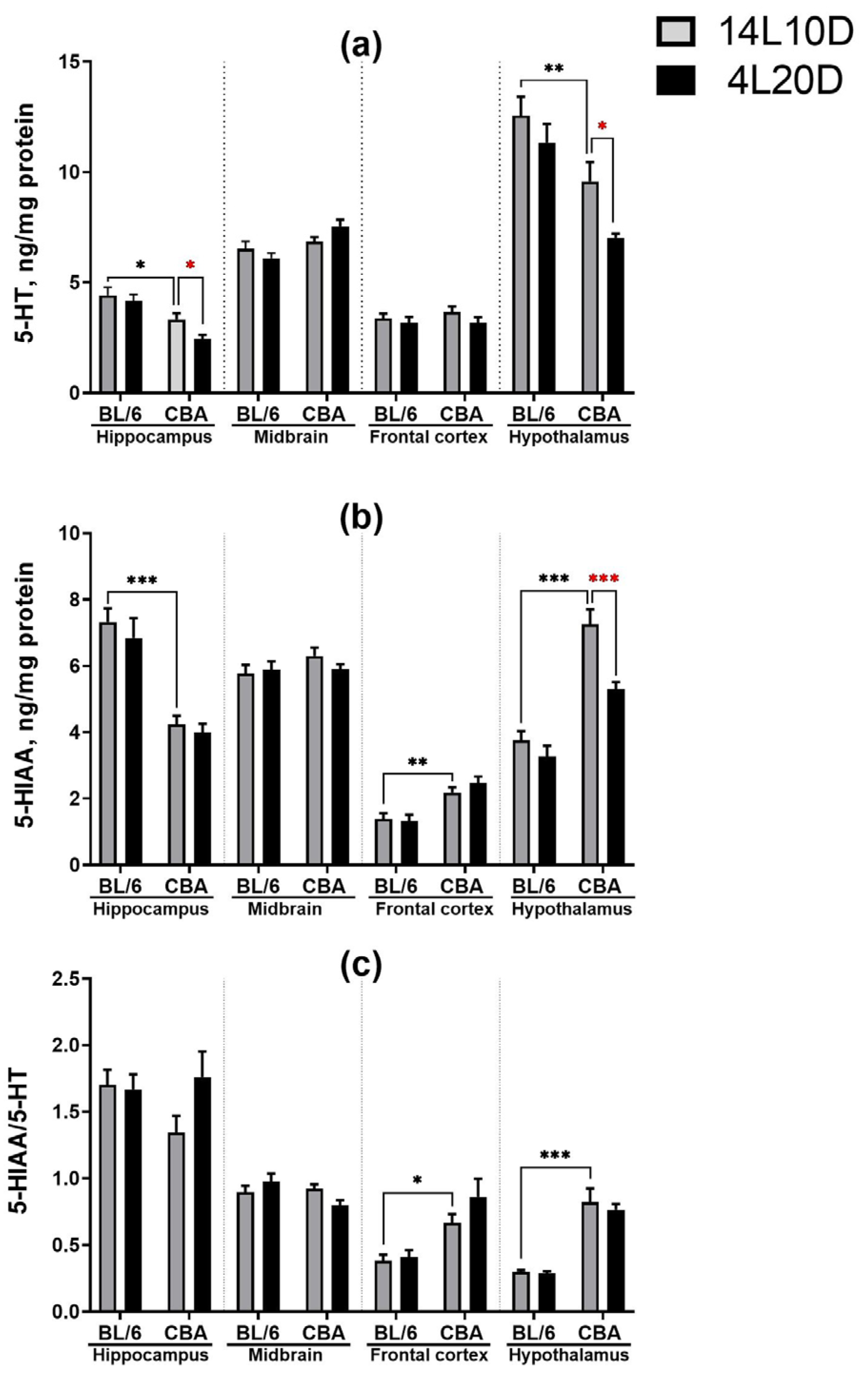
2.3. Effects of Short Photoperiod on Brain 5-HT Metabolism
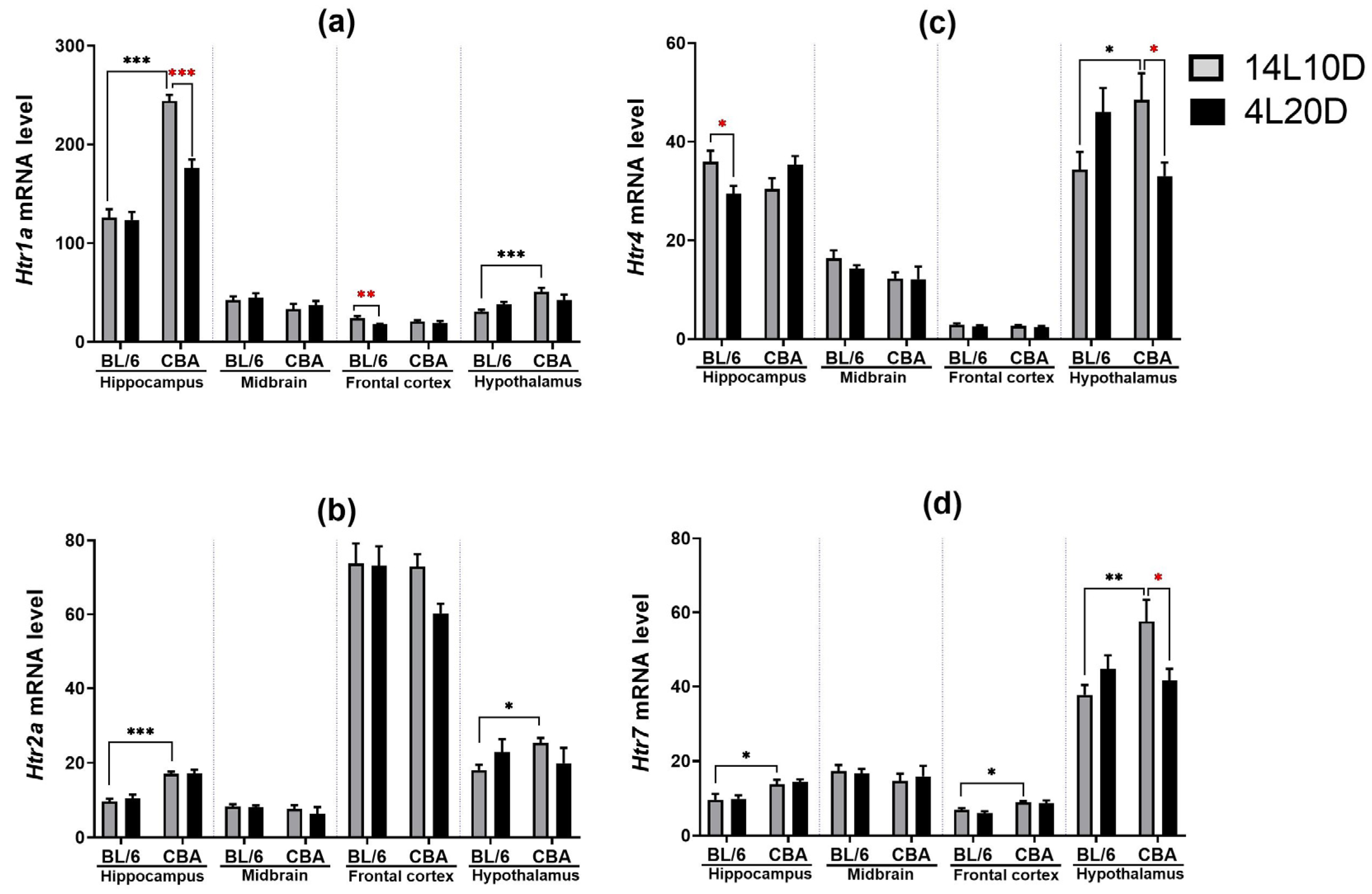
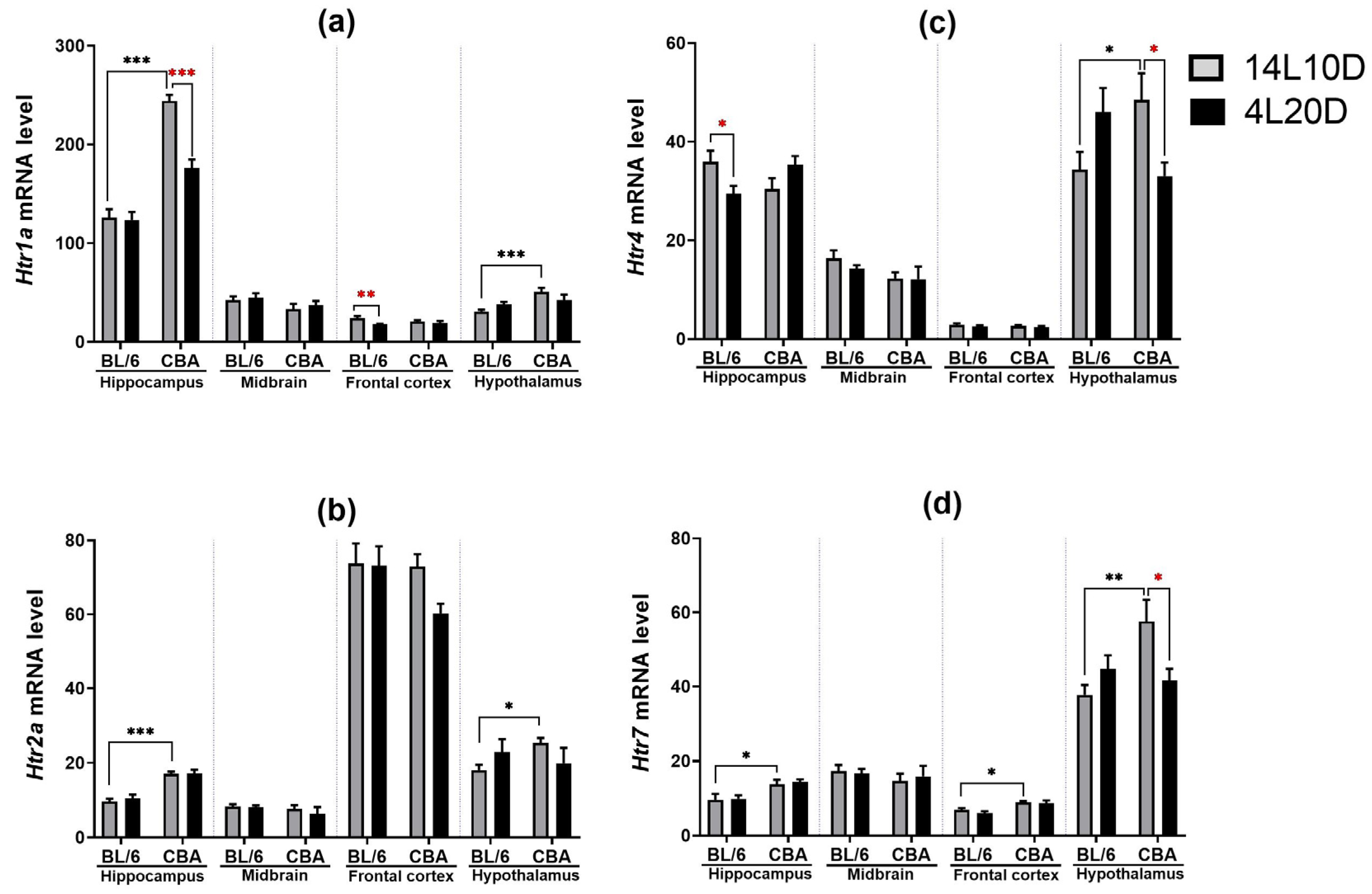
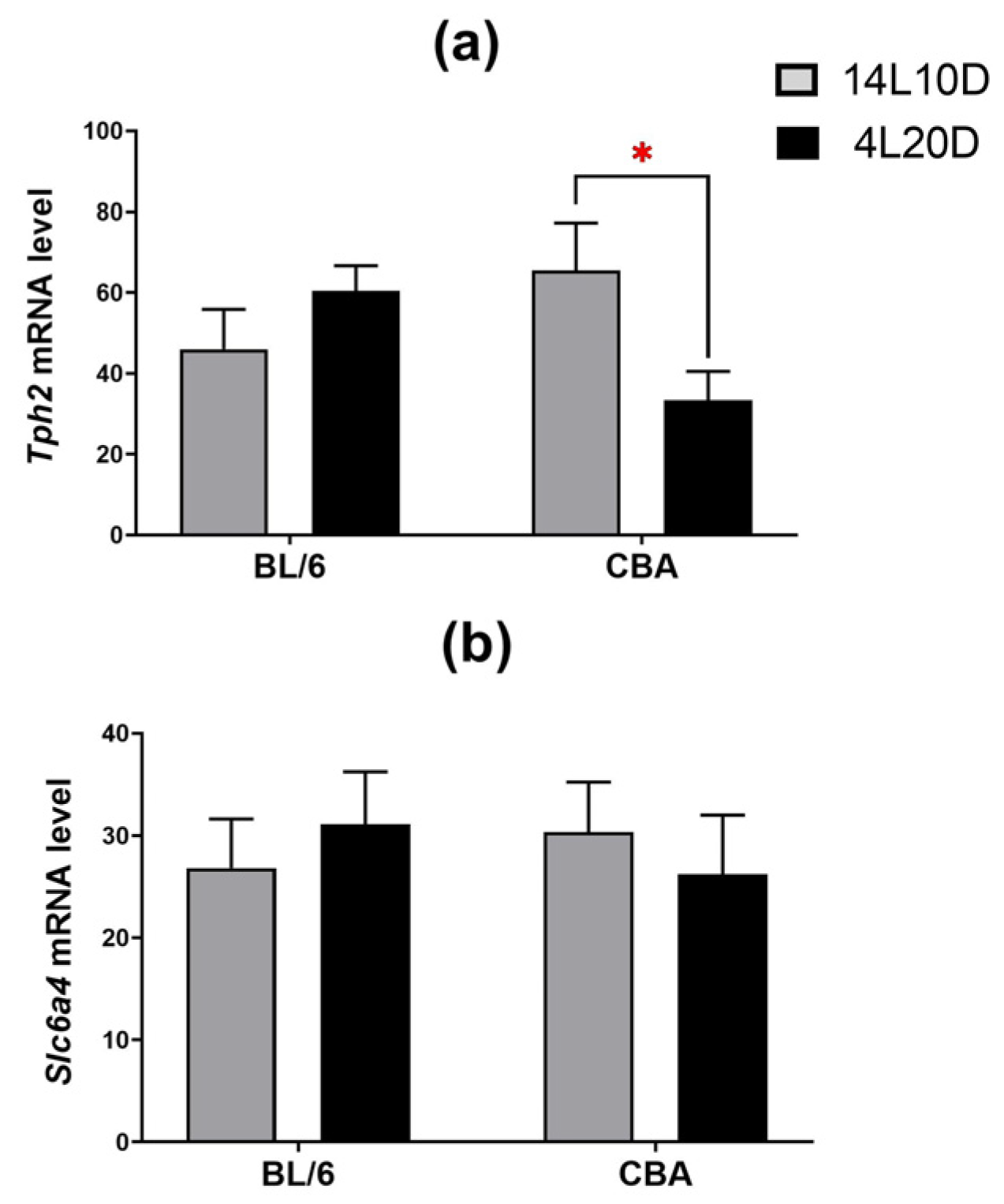
2.4. Effects of Short Photoperiod on mRNA Levels of 5-HT-Related Genes in the Brain
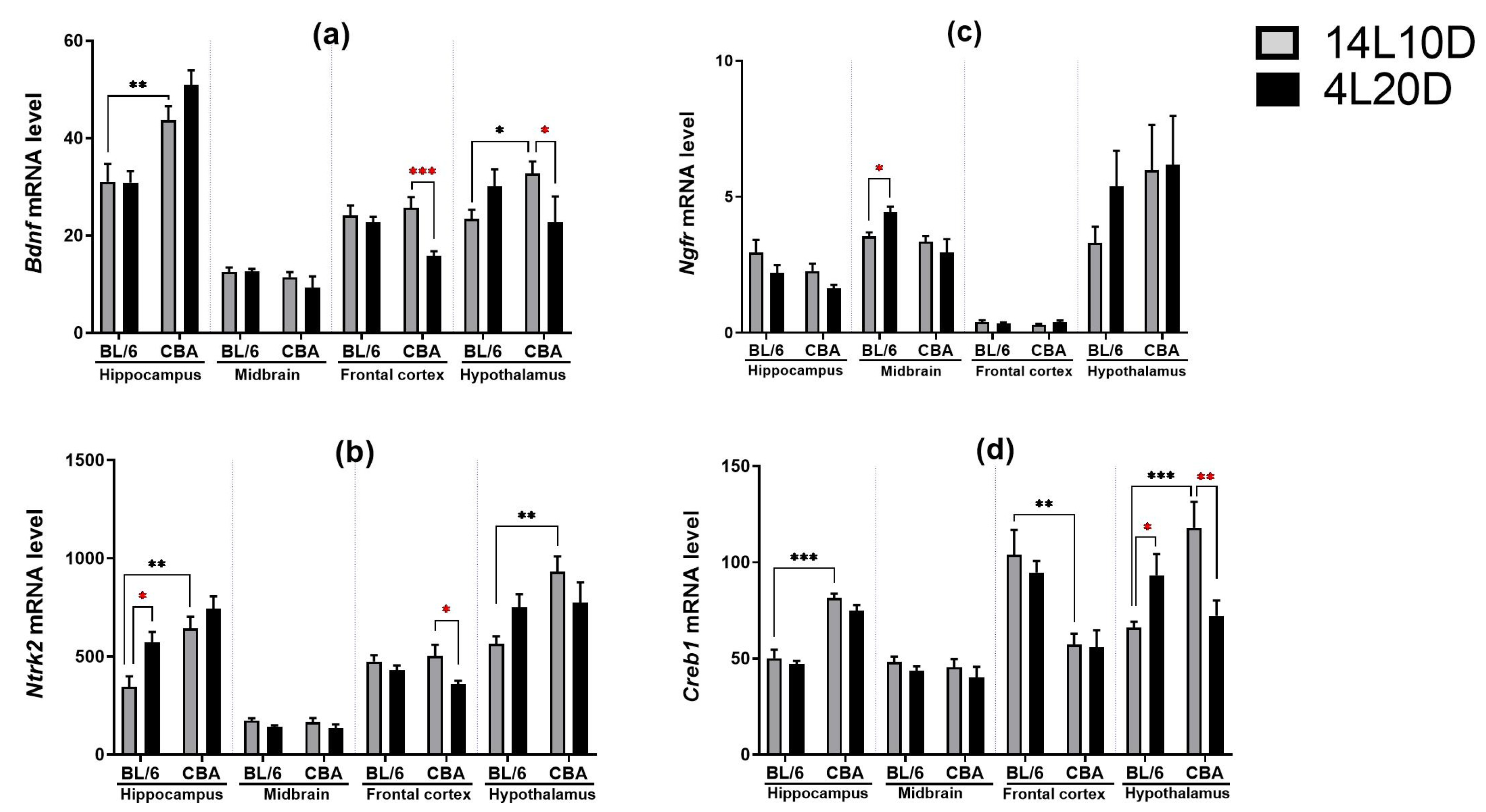
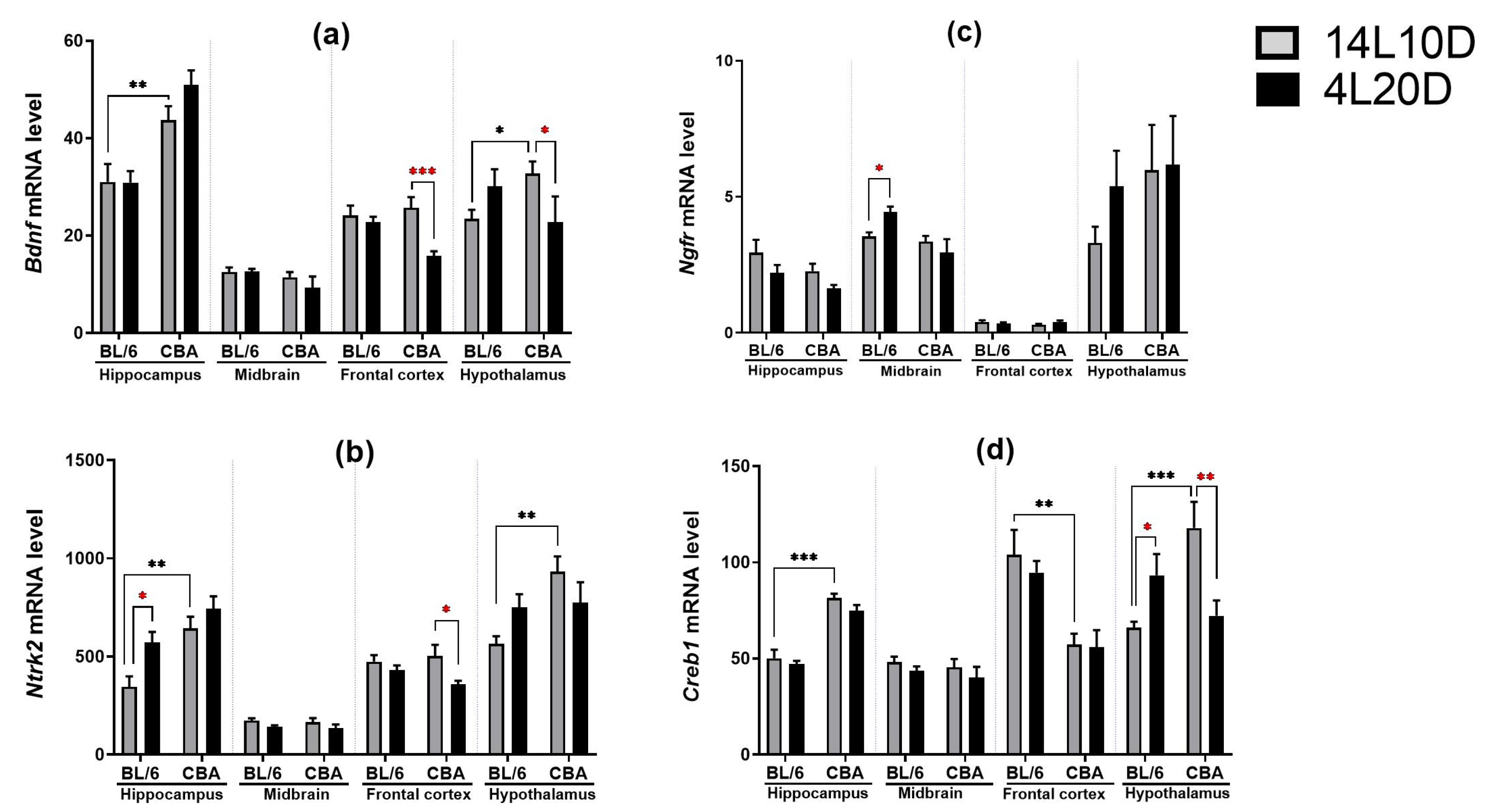
2.5. Effects of Short Photoperiod on mRNA Levels of BDNF-Related Genes in the Brain
3. Discussion
4. Materials and Methods
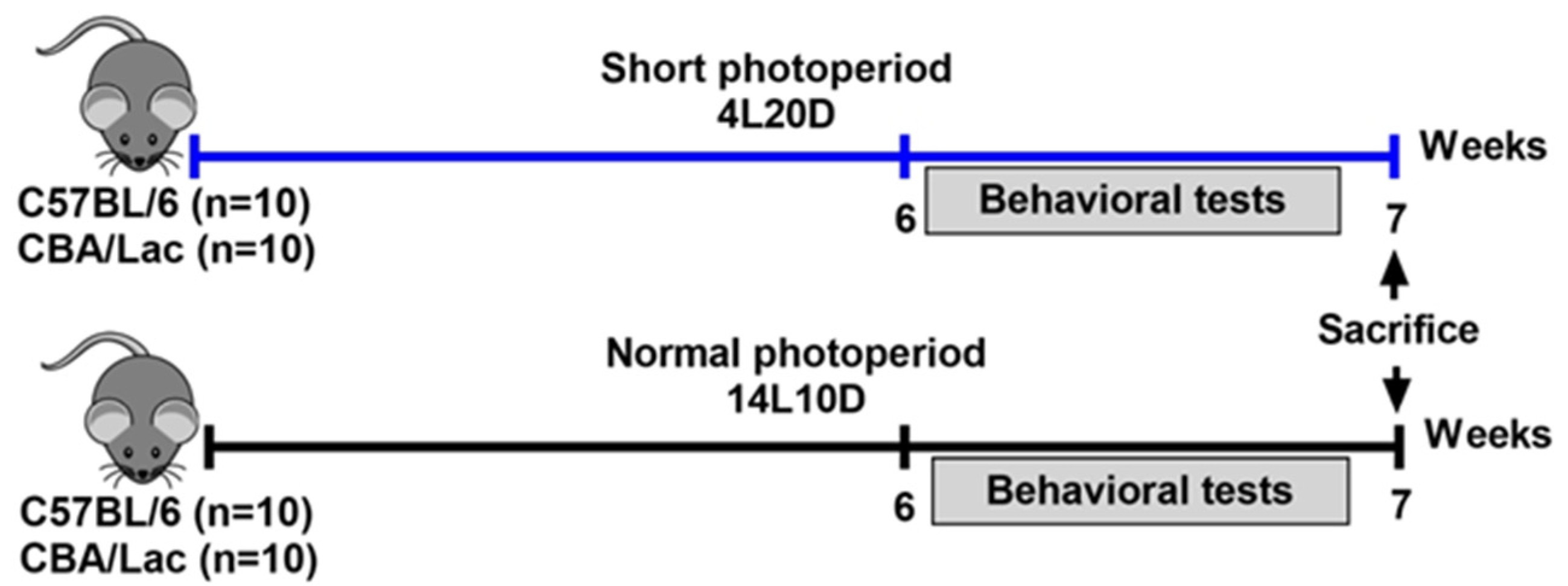
4.1. Animals and Treatment
4.2. Open Field Test
4.3. Forced Swim Test (FST)
4.4. Tail Suspension Test (TST)
4.5. Pinch-Induced Catalepsy
4.6. Tissue Preparation
4.7. Assay of 5-HT and 5-HIAA Levels
4.8. mRNA Level Assay by qPCR
4.9. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rosenthal, N.E.; Sack, D.A.; Gillin, J.C.; Lewy, A.J.; Goodwin, F.K.; Davenport, Y.; Mueller, P.S.; Newsome, D.A.; Wehr, T.A. Seasonal affective disorder. A description of the syndrome and preliminary findings with light therapy. Arch. Gen. Psychiatry 1984, 41, 72–80. [Google Scholar] [CrossRef]
- Magnusson, A.; Boivin, D. Seasonal affective disorder: An overview. Chronobiol. Int. 2003, 20, 189–207. [Google Scholar] [CrossRef]
- Levitan, R.D. The chronobiology and neurobiology of winter seasonal affective disorder. Dialogues Clin. Neurosci. 2007, 9, 315–324. [Google Scholar] [CrossRef]
- Meesters, Y.; Gordijn, M.C. Seasonal affective disorder, winter type: Current insights and treatment options. Psychol. Res. Behav. Manag. 2016, 9, 317–327. [Google Scholar] [CrossRef]
- Wirz-Justice, A. Seasonality in affective disorders. Gen. Comp. Endocrinol. 2018, 258, 244–249. [Google Scholar] [CrossRef]
- Popova, N.K.; Naumenko, V.S. Neuronal and behavioral plasticity: The role of serotonin and BDNF systems tandem. Expert Opin. Ther. Targets 2019, 23, 227–239. [Google Scholar] [CrossRef]
- Sharp, T.; Barnes, N.M. Central 5-HT receptors and their function; present and future. Neuropharmacology 2020, 177, 108155. [Google Scholar] [CrossRef]
- Popova, N.K.; Naumenko, V.S. 5-HT1A receptor as a key player in the brain 5-HT system. Rev. Neurosci. 2013, 24, 191–204. [Google Scholar] [CrossRef]
- Gupta, A.; Sharma, P.K.; Garg, V.K.; Singh, A.K.; Mondal, S.C. Role of serotonin in seasonal affective disorder. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 49–55. [Google Scholar]
- Kulikov, A.V.; Popova, N.K. Tryptophan hydroxylase 2 in seasonal affective disorder: Underestimated perspectives? Rev. Neurosci. 2015, 26, 679–690. [Google Scholar] [CrossRef]
- Luykx, J.J.; Bakker, S.C.; van Geloven, N.; Eijkemans, M.J.; Horvath, S.; Lentjes, E.; Boks, M.P.; Strengman, E.; DeYoung, J.; Buizer-Voskamp, J.E.; et al. Seasonal variation of serotonin turnover in human cerebrospinal fluid, depressive symptoms and the role of the 5-HTTLPR. Transl. Psychiatry 2013, 3, e311. [Google Scholar] [CrossRef]
- Molnar, E.; Lazary, J.; Benko, A.; Gonda, X.; Pap, D.; Mekli, K.; Juhasz, G.; Kovacs, G.; Kurimay, T.; Rihmer, Z.; et al. Seasonality and winter-type seasonal depression are associated with the rs731779 polymorphism of the serotonin-2A receptor gene. Eur. Neuropsychopharmacol. 2010, 20, 655–662. [Google Scholar] [CrossRef]
- Willeit, M.; Praschak-Rieder, N.; Neumeister, A.; Zill, P.; Leisch, F.; Stastny, J.; Hilger, E.; Thierry, N.; Konstantinidis, A.; Winkler, D.; et al. A polymorphism (5-HTTLPR) in the serotonin transporter promoter gene is associated with DSM-IV depression subtypes in seasonal affective disorder. Mol. Psychiatry 2003, 8, 942–946. [Google Scholar] [CrossRef]
- Gonzalez, A.; Moya-Alvarado, G.; Gonzalez-Billaut, C.; Bronfman, F.C. Cellular and molecular mechanisms regulating neuronal growth by brain-derived neurotrophic factor. Cytoskeleton 2016, 73, 612–628. [Google Scholar] [CrossRef]
- Sasi, M.; Vignoli, B.; Canossa, M.; Blum, R. Neurobiology of local and intercellular BDNF signaling. Pflug. Arch. 2017, 469, 593–610. [Google Scholar] [CrossRef]
- Givalois, L.; Naert, G.; Rage, F.; Ixart, G.; Arancibia, S.; Tapia-Arancibia, L. A single brain-derived neurotrophic factor injection modifies hypothalamo-pituitary-adrenocortical axis activity in adult male rats. Mol. Cell Neurosci. 2004, 27, 280–295. [Google Scholar] [CrossRef]
- Popova, N.K.; Ilchibaeva, T.V.; Antonov, E.V.; Pershina, A.V.; Bazovkina, D.V.; Naumenko, V.S. On the interaction between BDNF and serotonin systems: The effects of long-term ethanol consumption in mice. Alcohol 2020, 87, 1–15. [Google Scholar] [CrossRef]
- Arévalo, J.C.; Deogracias, R. Mechanisms Controlling the Expression and Secretion of BDNF. Biomolecules 2023, 13, 789. [Google Scholar] [CrossRef]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef]
- Zagrebelsky, M.; Tacke, C.; Korte, M. BDNF signaling during the lifetime of dendritic spines. Cell Tissue Res. 2020, 382, 185–199. [Google Scholar] [CrossRef]
- Molendijk, M.L.; Haffmans, J.P.; Bus, B.A.; Spinhoven, P.; Penninx, B.W.; Prickaerts, J.; Oude Voshaar, R.C.; Elzinga, B.M. Serum BDNF concentrations show strong seasonal variation and correlations with the amount of ambient sunlight. PLoS ONE 2012, 7, e48046. [Google Scholar] [CrossRef]
- Vinnik, T.; Kirby, M.; Bairachnaya, M.; Koman, I.; Tarkina, T.; Sadykova, G.; Abildinova, G.; Batpenova, G.; Pinhasov, A. Seasonality and BDNF polymorphism influences depression outcome in patients with atopic dermatitis and psoriasis. World J. Biol. Psychiatry 2017, 18, 604–614. [Google Scholar] [CrossRef]
- Otsuka, T.; Kawai, M.; Togo, Y.; Goda, R.; Kawase, T.; Matsuo, H.; Iwamoto, A.; Nagasawa, M.; Furuse, M.; Yasuo, S. Photoperiodic responses of depression-like behavior, the brain serotonergic system, and peripheral metabolism in laboratory mice. Psychoneuroendocrinology 2014, 40, 37–47. [Google Scholar] [CrossRef]
- Bazhenova, E.Y.; Fursenko, D.V.; Kulikova, E.A.; Khotskin, N.V.; Sinyakova, N.A.; Kulikov, A.A. Effect of photoperiodic alterations on depression-like behavior and the brain serotonin system in mice genetically different in tryptophan hydroxylase 2 activity. Neurosci. Lett. 2019, 699, 91–96. [Google Scholar] [CrossRef]
- Goda, R.; Otsuka, T.; Iwamoto, A.; Kawai, M.; Shibata, S.; Furuse, M.; Yasuo, S. Serotonin levels in the dorsal raphe nuclei of both chipmunks and mice are enhanced by long photoperiod, but brain dopamine level response to photoperiod is species-specific. Neurosci. Lett. 2015, 593, 95–100. [Google Scholar] [CrossRef]
- Weder, N.D.; Muralee, S.; Penland, H.; Tampi, R.R. Catatonia: A review. Ann. Clin. Psychiatry 2008, 20, 97–107. [Google Scholar] [CrossRef]
- Daniels, J. Catatonia: Clinical aspects and neurobiological correlates. J. Neuropsychiatry Clin. Neurosci. 2009, 21, 371–380. [Google Scholar] [CrossRef]
- Cattarinussi, G.; Gugliotta, A.A.; Hirjak, D.; Wolf, R.C.; Sambataro, F. Brain mechanisms underlying catatonia: A systematic review. Schizophr. Res. 2024, 263, 194–207. [Google Scholar] [CrossRef]
- Kulikov, A.V.; Kozlachkova, E.Y.; Maslova, G.B.; Popova, N.K. Inheritance of predisposition to catalepsy in mice. Behav. Genet. 1993, 23, 379–384. [Google Scholar] [CrossRef]
- Tikhonova, M.A.; Kulikov, A.V.; Bazovkina, D.V.; Kulikova, E.A.; Tsybko, A.S.; Bazhenova, E.Y.; Naumenko, V.S.; Akulov, A.E.; Moshkin, M.P.; Popova, N.K. Hereditary catalepsy in mice is associated with the brain dysmorphology and altered stress response. Behav. Brain Res. 2013, 243, 53–60. [Google Scholar] [CrossRef]
- Tikhonova, M.A.; Alperina, E.L.; Tolstikova, T.G.; Bazovkina, D.V.; Di, V.Y.; Idova, G.V.; Kulikov, A.V.; Popova, N.K. Effects of chronic fluoxetine treatment on catalepsy and the immune response in mice with a genetic predisposition to freezing reactions: The roles of types 1A and 2A serotonin receptors and the tph2 and SERT genes. Neurosci. Behav. Physiol. 2010, 40, 521–527. [Google Scholar] [CrossRef]
- Naumenko, V.S.; Kondaurova, E.M.; Bazovkina, D.V.; Tsybko, A.S.; Tikhonova, M.A.; Kulikov, A.V.; Popova, N.K. Effect of brain-derived neurotrophic factor on behavior and key members of the brain serotonin system in genetically predisposed to behavioral disorders mouse strains. Neuroscience 2012, 214, 59–67. [Google Scholar] [CrossRef]
- Otsuka, T.; Goda, R.; Iwamoto, A.; Kawai, M.; Shibata, S.; Oka, Y.; Mizunoya, W.; Furuse, M.; Yasuo, S. Dietary protein ingested before and during short photoperiods makes an impact on affect-related behaviours and plasma composition of amino acids in mice. Br. J. Nutr. 2015, 114, 1734–1743. [Google Scholar] [CrossRef]
- Uchiwa, T.; Takai, Y.; Tashiro, A.; Furuse, M.; Yasuo, S. Exposure of C57BL/6J mice to long photoperiod during early life stages increases body weight and alters plasma metabolomic profiles in adulthood. Physiol. Rep. 2016, 4, e12974. [Google Scholar] [CrossRef]
- Taniguchi, E.; Tashiro, A.; Hattori, A.; Furuse, M.; Yasuo, S. Photoperiodic changes in hippocampal neurogenesis and plasma metabolomic profiles in relation to depression-like behavior in mice. Behav. Brain Res. 2021, 403, 113136. [Google Scholar] [CrossRef]
- Fursenko, D.V.; Bazhenova, E.Y.; Khotskin, N.V.; Sorokin, I.E.; Kulikova, E.A.; Kulikov, A.V. Effect of Photoperiod and Lethal Yellow Mutation on Depression-Like Behavior and Expression of Proinflammatory Cytokines in the Hypothalamus in Mice. Bull. Exp. Biol. Med. 2019, 167, 100–103. [Google Scholar] [CrossRef]
- Porsolt, R.D.; Brossard, G.; Hautbois, C.; Roux, S. Rodent models of depression: Forced swimming and tail suspension behavioral de spair tests in rats and mice. Curr. Protoc. Neurosci. 2001, 14, 8.10A.1–8.10A.10. [Google Scholar] [CrossRef]
- Powell, T.R.; Fernandes, C.; Schalkwyk, L.C. Depression-Related Behavioral Tests. Curr. Protoc. Mouse Biol. 2012, 2, 119–127. [Google Scholar] [CrossRef]
- Workman, J.L.; Nelson, R.J. Potential animal models of seasonal affective disorder. Neurosci. Biobehav. Rev. 2011, 35, 669–679. [Google Scholar] [CrossRef]
- Edinoff, A.N.; Raveendran, K.; Colon, M.A.; Thomas, B.H.; Trettin, K.A.; Hunt, G.W.; Kaye, A.M.; Cornett, E.M.; Kaye, A.D. Selective Serotonin Reuptake Inhibitors and Associated Bleeding Risks: A Narrative and Clinical Review. Health Psychol. Res. 2022, 10, 39580. [Google Scholar] [CrossRef]
- Hamon, M.; Blier, P. Monoamine neurocircuitry in depression and strategies for new treatments. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 45, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.L.W.; Harmer, C.J.; Cowen, P.J.; Murphy, S.E. The Serotonin 1A (5-HT1A) Receptor as a Pharmacological Target in Depression. CNS Drugs 2023, 37, 571–585. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, L.; Korkutata, M.; Vimal, S.K.; Yadav, M.K.; Bhattacharyya, S.; Shiga, T. Therapeutic potential of serotonin 4 receptor for chronic depression and its associated comorbidity in the gut. Neuropharmacology 2020, 166, 107969. [Google Scholar] [CrossRef] [PubMed]
- Stiedl, O.; Pappa, E.; Konradsson-Geuken, Å.; Ögren, S.O. The role of the serotonin receptor subtypes 5-HT1A and 5-HT7 and its interaction in emotional learning and memory. Front. Pharmacol. 2015, 6, 162. [Google Scholar] [CrossRef] [PubMed]
- Bombardi, C.; Grandis, A.; Pivac, N.; Sagud, M.; Lucas, G.; Chagraoui, A.; Lemaire-Mayo, V.; De Deurwaerdère, P.; Di Giovanni, G. Serotonin modulation of hippocampal functions: From anatomy to neurotherapeutics. Prog. Brain Res. 2021, 261, 83–158. [Google Scholar]
- Carlsson, A.; Svennerholm, L.; Winblad, B. Seasonal and circadian monoamine variations in human brains examined post mortem. Acta Psychiatr. Scand. 1980, 61, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Casey, A.B.; Cui, M.; Booth, R.G.; Canal, C.E. “Selective” serotonin 5-HT2A receptor antagonists. Biochem. Pharmacol. 2022, 200, 115028. [Google Scholar] [CrossRef]
- Kasahara, T.; Abe, K.; Mekada, K.; Yoshiki, A.; Kato, T. Genetic variation of melatonin productivity in laboratory mice under domestication. Proc. Natl. Acad. Sci. USA 2010, 107, 6412–6417. [Google Scholar] [CrossRef]
- Goto, M.; Oshima, I.; Tomita, T.; Ebihara, S. Melatonin content of the pineal gland in different mouse strains. J. Pineal. Res. 1989, 7, 195–204. [Google Scholar] [CrossRef]
- Kennaway, D.J. Melatonin research in mice: A review. Chronobiol. Int. 2019, 36, 1167–1183. [Google Scholar] [CrossRef]
- Sakamoto, K.; Norona, F.E.; Alzate-Correa, D.; Scarberry, D.; Hoyt, K.R.; Obrietan, K. Clock and light regulation of the CREB coactivator CRTC1 in the suprachiasmatic circadian clock. J. Neurosci. 2013, 33, 9021–9027. [Google Scholar] [CrossRef]
- Zanesco, A.M.; Mendes, N.F.; Engel, D.F.; Gaspar, R.S.; Sidarta-Oliveira, D.; Donato, J., Jr.; Velloso, L.A. Hypothalamic CREB Regulates the Expression of Pomc-Processing Enzyme Pcsk2. Cells 2022, 11, 1996. [Google Scholar] [CrossRef]
- Yang, Y.; Cong, W.; Liu, J.; Zhao, M.; Xu, P.; Han, W.; Wang, D.; Zhao, R. Constant light in early life induces fear-related behavior in chickens with suppressed melatonin secretion and disrupted hippocampal expression of clock- and BDNF-associated genes. J. Anim. Sci. Biotechnol. 2022, 13, 67. [Google Scholar] [CrossRef] [PubMed]
- Carboni, L.; Rullo, L.; Caputi, F.F.; Stamatakos, S.; Candeletti, S.; Romualdi, P. Chronic Trazodone and Citalopram Treatments Increase Trophic Factor and Circadian Rhythm Gene Expression in Rat Brain Regions Relevant for Antidepressant Efficacy. Int. J. Mol. Sci. 2022, 23, 14041. [Google Scholar] [CrossRef]
- Amigó, J.; Díaz, A.; Pilar-Cuéllar, F.; Vidal, R.; Martín, A.; Compan, V.; Pazos, A.; Castro, E. The absence of 5-HT4 receptors modulates depression- and anxiety-like responses and influences the response of fluoxetine in olfactory bulbectomised mice: Adaptive changes in hippocampal neuroplasticity markers and 5-HT1A autoreceptor. Neuropharmacology 2016, 111, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Brazo, J.; Castro, E.; Díaz, A.; Valdizán, E.M.; Pilar-Cuéllar, F.; Vidal, R.; Treceño, B.; Pazos, A. Modulation of neuroplasticity pathways and antidepressant-like behavioural responses following the short-term (3 and 7 days) administration of the 5-HT4 receptor agonist RS67333. Int. J. Neuropsychopharmacol. 2012, 15, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Slotnick, B.M.; Leonard, C.M. A Stereotaxic Atlas of the Albino Mouse Forebrain; The United States Department of Health, Education and Welfare: Rockville, MD, USA, 1975; 174p. [Google Scholar]
- Kulikov, A.V.; Tikhonova, M.A.; Kulikov, V.A. Automated measurement of spatial preference in the open field test with transmitted lighting. J. Neurosci. Methods 2008, 170, 345–351. [Google Scholar] [CrossRef]
- Kulikov, A.V.; Morozova, M.V.; Kulikov, V.A.; Kirichuk, V.S.; Popova, N.K. Automated analysis of antidepressants’ effect in the forced swim test. J. Neurosci. Methods 2010, 191, 26–31. [Google Scholar] [CrossRef]
- Bazovkina, D.; Naumenko, V.; Bazhenova, E.; Kondaurova, E. Effect of Central Administration of Brain-Derived Neurotrophic Factor (BDNF) on Behavior and Brain Monoamine Metabolism in New Recombinant Mouse Lines Differing by 5-HT1A Receptor Functionality. Int. J. Mol. Sci. 2021, 22, 11987. [Google Scholar] [CrossRef]
- Khotskin, N.V.; Plyusnina, A.V.; Kulikova, E.A.; Bazhenova, E.Y.; Fursenko, D.V.; Sorokin, I.E.; Kolotygin, I.; Mormede, P.; Terenina, E.E.; Shevelev, O.B.; et al. On association of the lethal yellow (AY) mutation in the agouti gene with the alterations in mouse brain and behavior. Behav. Brain Res. 2019, 359, 446–456. [Google Scholar] [CrossRef]
- Naumenko, V.S.; Kulikov, A.V. Quantitative assay of 5-HT(1A) serotonin receptor gene expression in the brain. Mol. Biol. 2006, 40, 37–44. [Google Scholar] [CrossRef]
- Naumenko, V.S.; Osipova, D.V.; Kostina, E.V.; Kulikov, A.V. Utilization of a two-standard system in real-time PCR for quantification of gene expression in the brain. J. Neurosci. Methods 2008, 170, 197–203. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect of Genotype | Effect of Photoperiod | Effect of Interaction | |
|---|---|---|---|
| 5-HT | |||
| Hippocampus | F(1,36) = 22.81 *** | F(1,36) = 3.97, p = 0.054 | F(1,36) = 1.11, p > 0.05 |
| Midbrain | F(1,36) = 10.12 ** | F(1,36) < 1 | F(1,36) = 4.01, p = 0.053 |
| Frontal cortex | F (1,35) < 1 | F(1,35) = 1.93, p > 0.05 | F(1,35) < 1 |
| Hypothalamus | F (1,35) = 24.30 *** | F(1,35) = 6.52 * | F(1,35) < 1 |
| 5-HIAA | |||
| Hippocampus | F(1,36) = 53.49 *** | F(1,36) < 1 | F(1,36) < 1 |
| Midbrain | F(1,36) = 1.37, p > 0.05 | F(1,36) < 1 | F(1,36) = 1.20, p > 0.05 |
| Frontal cortex | F(1,35) = 28.50 *** | F(1,35) < 1 | F(1,35) < 1 |
| Hypothalamus | F(1,35) = 68.02 *** | F(1,35) = 13.21 *** | F(1,35) = 4.74 * |
| 5-HIAA/5-HT | |||
| Hippocampus | F(1,36) < 1 | F(1,36) = 1.83, p > 0.05 | F(1,36) = 2.57, p > 0.05 |
| Midbrain | F(1,36) = 2.83, p > 0.05 | F(1,36) < 1 | F(1,36) = 4.99 * |
| Frontal cortex | F(1,35) = 19.03 *** | F(1,35) = 1.74, p > 0.05 | F(1,35) < 1 |
| Hypothalamus | F(1,35) = 72.89 *** | F(1,35) < 1 | F(1,35) < 1 |
| Effect of Genotype | Effect of Photoperiod | Effect of Interaction | |
|---|---|---|---|
| Htr1a | |||
| Hippocampus | F(1,35) = 116.43 *** | F(1,35) = 19.56 *** | F(1,35) = 17.05 *** |
| Midbrain | F(1,34) = 3.61, p = 0.066 | F(1,34) < 1 | F(1,34) < 1 |
| Frontal cortex | F(1,36) < 1 | F(1,36) = 5.88 * | F(1,36) = 2.79, p > 0.05 |
| Hypothalamus | F(1,33) = 6.11 * | F(1,33) < 1 | F(1,33) = 4.71 * |
| Htr2a | |||
| Hippocampus | F(1,35) = 64.24 *** | F(1,35) < 1 | F(1,35) < 1 |
| Midbrain | F(1,34) = 1.31, p > 0.05 | F(1,34) < 1 | F(1,34) < 1 |
| Frontal cortex | F(1,36) = 2.36, p > 0.05 | F(1,36) = 2.15, p > 0.05 | F(1,36) = 1.79, p > 0.05 |
| Hypothalamus | F(1,33) < 1 | F(1,33) < 1 | F(1,33) = 4.31 * |
| Htr4 | |||
| Hippocampus | F(1,35) < 1 | F(1,35) < 1 | F(1,35) = 8.42 ** |
| Midbrain | F(1,34) = 1.65, p > 0.05 | F(1,34) < 1 | F(1,34) < 1 |
| Frontal cortex | F(1,36) < 1 | F(1,36) = 1.02, p > 0.05 | F(1,36) < 1 |
| Hypothalamus | F(1,33) < 1 | F(1,33) < 1 | F(1,33) = 7.46 * |
| Htr7 | |||
| Hippocampus | F(1,36) = 13.46 *** | F(1,36) < 1 | F(1,36) < 1 |
| Midbrain | F(1,34) < 1 | F(1,34) < 1 | F(1,34) < 1 |
| Frontal cortex | F(1,36) = 16.44 *** | F(1,36) = 1.14, p > 0.05 | F(1,36) < 1 |
| Hypothalamus | F(1,33) = 3.43, p = 0.073 | F(1,33) < 1 | F(1,33) = 6.40 * |
| Tph2 | |||
| Midbrain | F(1,34) < 1 | F(1,34) = 1.02, p > 0.05 | F(1,34) = 5.16 * |
| Slc6a4 | |||
| Midbrain | F(1,34) < 1 | F(1,34) < 1 | F(1,34) < 1 |
| Effect of Genotype | Effect of Photoperiod | Effect of Interaction | |
|---|---|---|---|
| Bdnf | |||
| Hippocampus | F(1,36) = 31.71 *** | F(1,36) = 1.93, p > 0.05 | F(1,36) < 1 |
| Midbrain | F(1,35) = 2.83, p > 0.05 | F(1,35) < 1 | F(1,35) < 1 |
| Frontal cortex | F(1,36) = 2.63, p > 0.05 | F(1,36) = 11.73 ** | F(1,36) = 6.60 * |
| Hypothalamus | F(1,33) < 1 | F(1,33) < 1 | F(1,33) = 7.29 * |
| Ntrk2 | |||
| Hippocampus | F(1,36) = 6.73 ** | F(1,36) = 4.63 * | F(1,36) = 1.48, p > 0.05 |
| Midbrain | F(1,35) < 1 | F(1,35)= 4.44 * | F(1,35) < 1 |
| Frontal cortex | F(1,35) < 1 | F(1,35) = 6.38 * | F(1,35) = 1.87, p > 0.05 |
| Hypothalamus | F(1,33) = 7.29 * | F(1,33) < 1 | F(1,33) = 5.63 * |
| Ngfr | |||
| Hippocampus | F(1,36) < 1 | F(1,36) = 4.51 * | F(1,36) < 1 |
| Midbrain | F(1,35) = 8.02 ** | F(1,35) < 1 | F(1,35) = 4.86 * |
| Frontal cortex | F(1,36) < 1 | F(1,36) < 1 | F(1,36) = 2.04, p > 0.05 |
| Hypothalamus | F(1,33) = 1.43, p > 0.05 | F(1,33) < 1 | F(1,33) < 1 |
| Creb1 | |||
| Hippocampus | F(1,36) = 94.78 *** | F(1,36) = 2.52, p > 0.05 | F(1,36) < 1 |
| Midbrain | F(1,35) < 1 | F(1,35) = 1.72, p > 0.05 | F(1,35) < 1 |
| Frontal cortex | F(1,35) = 21.98 *** | F(1,35) < 1 | F(1,35) < 1 |
| Hypothalamus | F(1,33) = 4.77 * | F(1,33) = 1.55, p > 0.05 | F(1,33) = 16.81 *** |
| Index | Short Photoperiod Exposure | ||
|---|---|---|---|
| Body weight | BL/6 = | CBA ↑ | |
| Behavior | FST depressive-like immobility TST depressive-like immobility | BL/6 = BL/6 ↑ | CBA ↑ CBA = |
| Cataleptic immobility | No | CBA ↑ | |
| 5-HT brain system: metabolism | 5-HT level in Ht and in Hc 5-HIAA level in Ht | BL/6 = | CBA ↓ |
| 5-HT brain system: expression of key genes | Htr1a mRNA in Hc Htr4 and Htr7 mRNA in Ht Tph2 mRNA in Mb | BL/6 = | CBA ↓ |
| Htr1a mRNA in FC Htr4 mRNA in Hc | BL/6 ↓ | CBA = | |
| Expression of BDNF-related genes | Bdnf mRNA in FC and Ht Ntrk2 mRNA in FC | BL/6 = | CBA ↓ |
| Ngfr mRNA in Mb Ntrk2 mRNA in Hc | BL/6 ↑ | CBA = | |
| Creb1 mRNA in Ht | BL/6 ↑ | CBA ↓ | |
| Gene | Sequence | Annealing Temperature, °C | Product Length, bp |
|---|---|---|---|
| Htr1a | F 5′-ctgtgacctgtttatcgccctg-3′ R 5′-gtagtctatagggtcggtgattgc-3′ | 62 | 109 |
| Htr2a | F 5′-agaagccaccttgtgtgtga-3′ R 5′-ttgctcattgctgatggact-3′ | 61 | 169 |
| Htr4 | F 5′-gtggtgtgtcttcatggtcaac-3′ R 5′-ctcctgcccgttgtaacatc-3′ | 62 | 155 |
| Htr7 | F 5′-ggctacacgatctactccaccg-3′ R 5′-cgcacactcttccacctccttc-3′ | 65 | 198 |
| Tph2 | F 5′-cattcctcgcacaattccagtcg-3′ R 5′-cttgacatattcaactagacgctc-3′ | 61 | 239 |
| Scl6a4 | F 5′-cgctctactacctcatctcctcc-3′ R 5′-gtcctgggcgaagtagttgg-3′ | 63 | 101 |
| Bdnf | F 5′- tagcaaaaagagaattggctg-3′ R 5′- tttcaggtcatggatatgtcc-3′ | 59 | 255 |
| Ntrk2 | F 5′-cattcactgtgagaggcaacc-3′ R 5′-atcagggtgtagtctccgttatt-3′ | 63 | 175 |
| Ngfr | F 5′-acaacacccagcacccagga-3′ R 5′-cacaaccacagcagccaaga-3′ | 62 | 171 |
| Creb1 | F 5′-gctggctaacaatggtacggat-3′ R 5′-tggttgctgggcactagaat-3′ | 64 | 140 |
| Polr2a | F 5′-tgtgacaactccatacaatgc-3′ R 5′-ctctcttagtgaatttgcgtact-3′ | 61 | 194 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adonina, S.; Bazhenova, E.; Bazovkina, D. Effect of Short Photoperiod on Behavior and Brain Plasticity in Mice Differing in Predisposition to Catalepsy: The Role of BDNF and Serotonin System. Int. J. Mol. Sci. 2024, 25, 2469. https://doi.org/10.3390/ijms25052469
Adonina S, Bazhenova E, Bazovkina D. Effect of Short Photoperiod on Behavior and Brain Plasticity in Mice Differing in Predisposition to Catalepsy: The Role of BDNF and Serotonin System. International Journal of Molecular Sciences. 2024; 25(5):2469. https://doi.org/10.3390/ijms25052469
Chicago/Turabian StyleAdonina, Svetlana, Ekaterina Bazhenova, and Darya Bazovkina. 2024. "Effect of Short Photoperiod on Behavior and Brain Plasticity in Mice Differing in Predisposition to Catalepsy: The Role of BDNF and Serotonin System" International Journal of Molecular Sciences 25, no. 5: 2469. https://doi.org/10.3390/ijms25052469
APA StyleAdonina, S., Bazhenova, E., & Bazovkina, D. (2024). Effect of Short Photoperiod on Behavior and Brain Plasticity in Mice Differing in Predisposition to Catalepsy: The Role of BDNF and Serotonin System. International Journal of Molecular Sciences, 25(5), 2469. https://doi.org/10.3390/ijms25052469