
Knockdown of DNMT1 Induces SLCO3A1 to Promote Follicular Growth by Enhancing the Proliferation of Granulosa Cells in Mammals

 , ,
, ,
Abstract
1. Introduction
2. Results
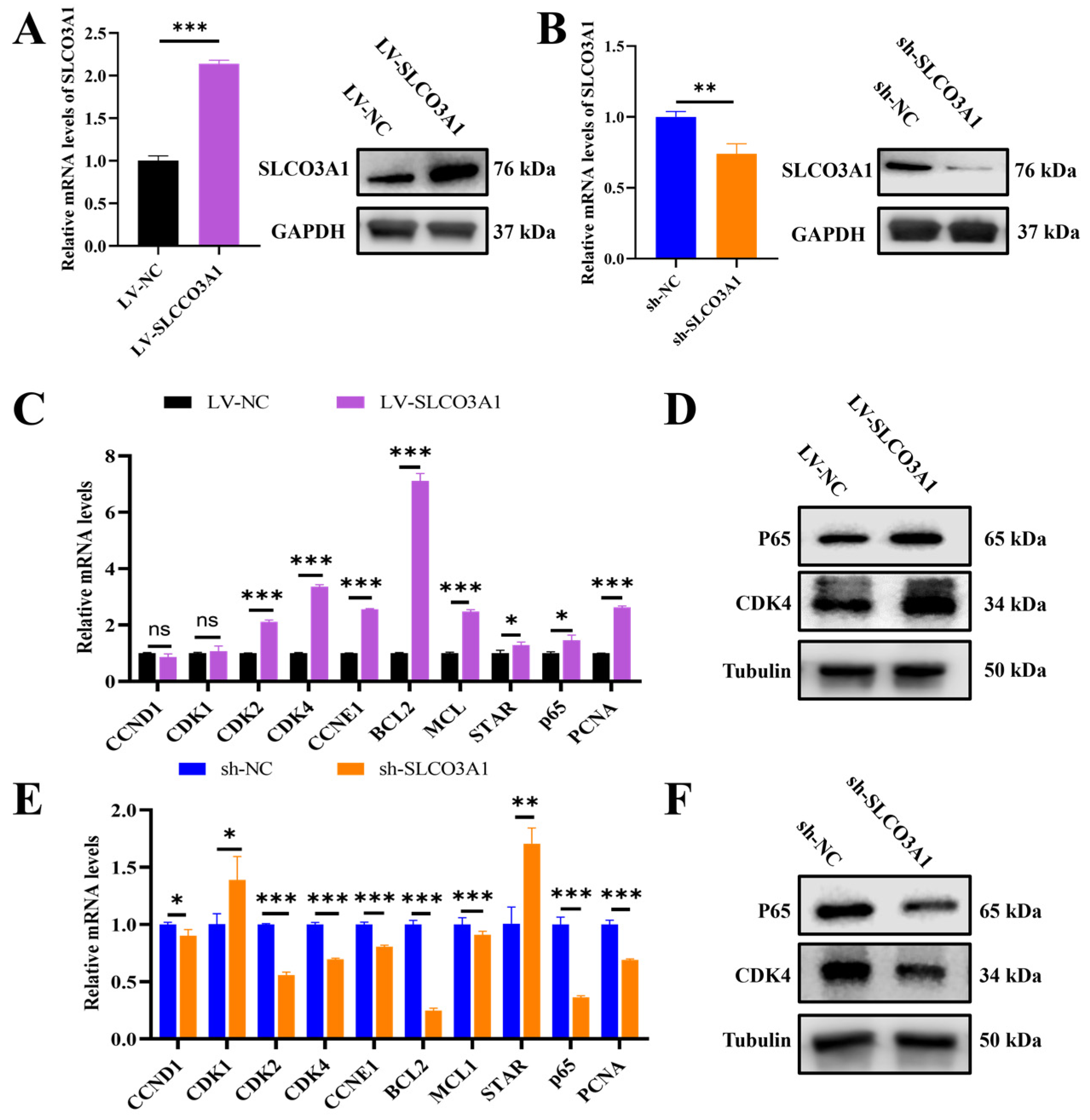
2.1. SLCO3A1 Promotes the Proliferation of GCs
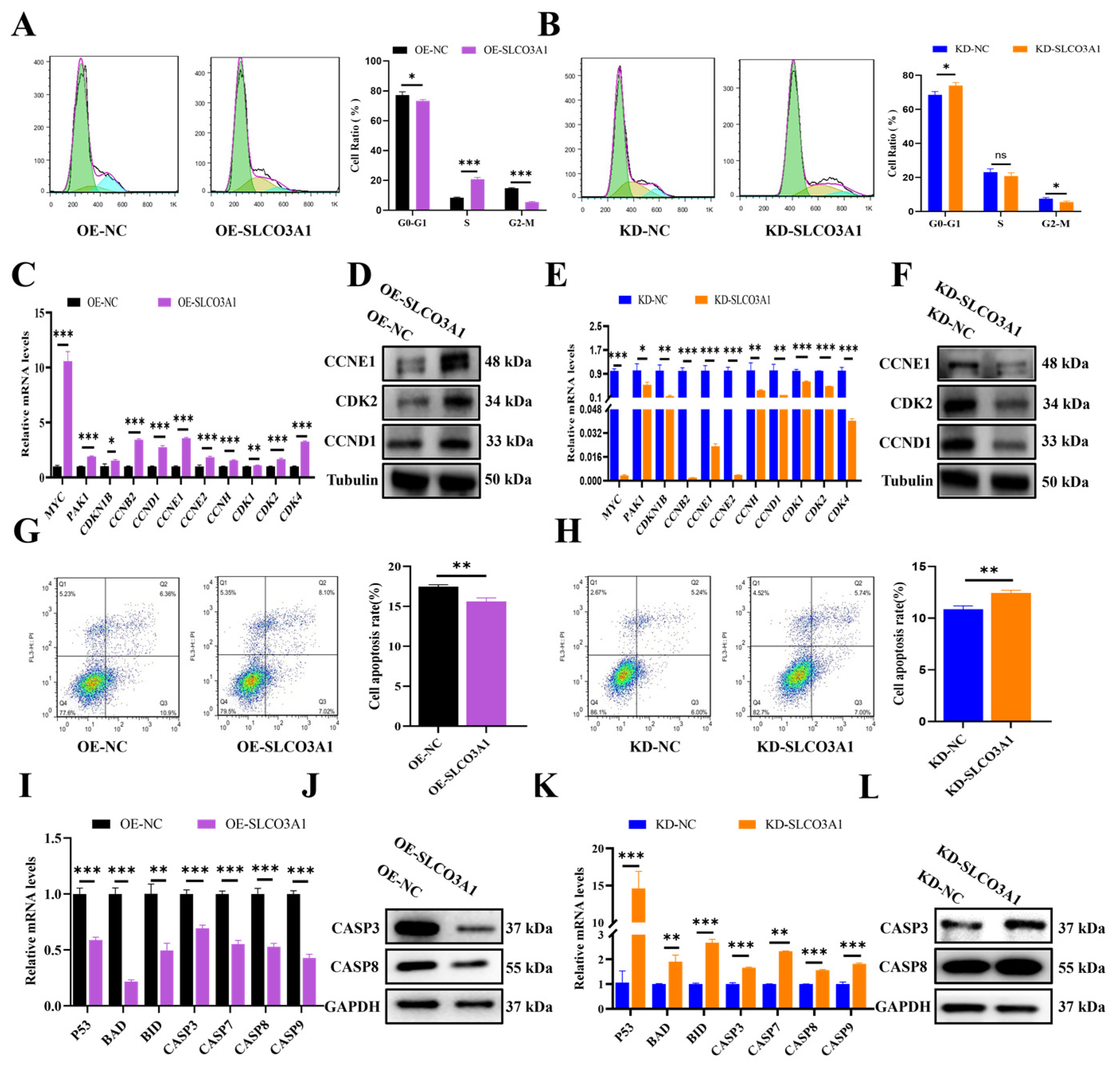
2.2. SLCO3A1 Promotes the Cell Cycle-Related Processes but Inhibits the Apoptosis of GCs
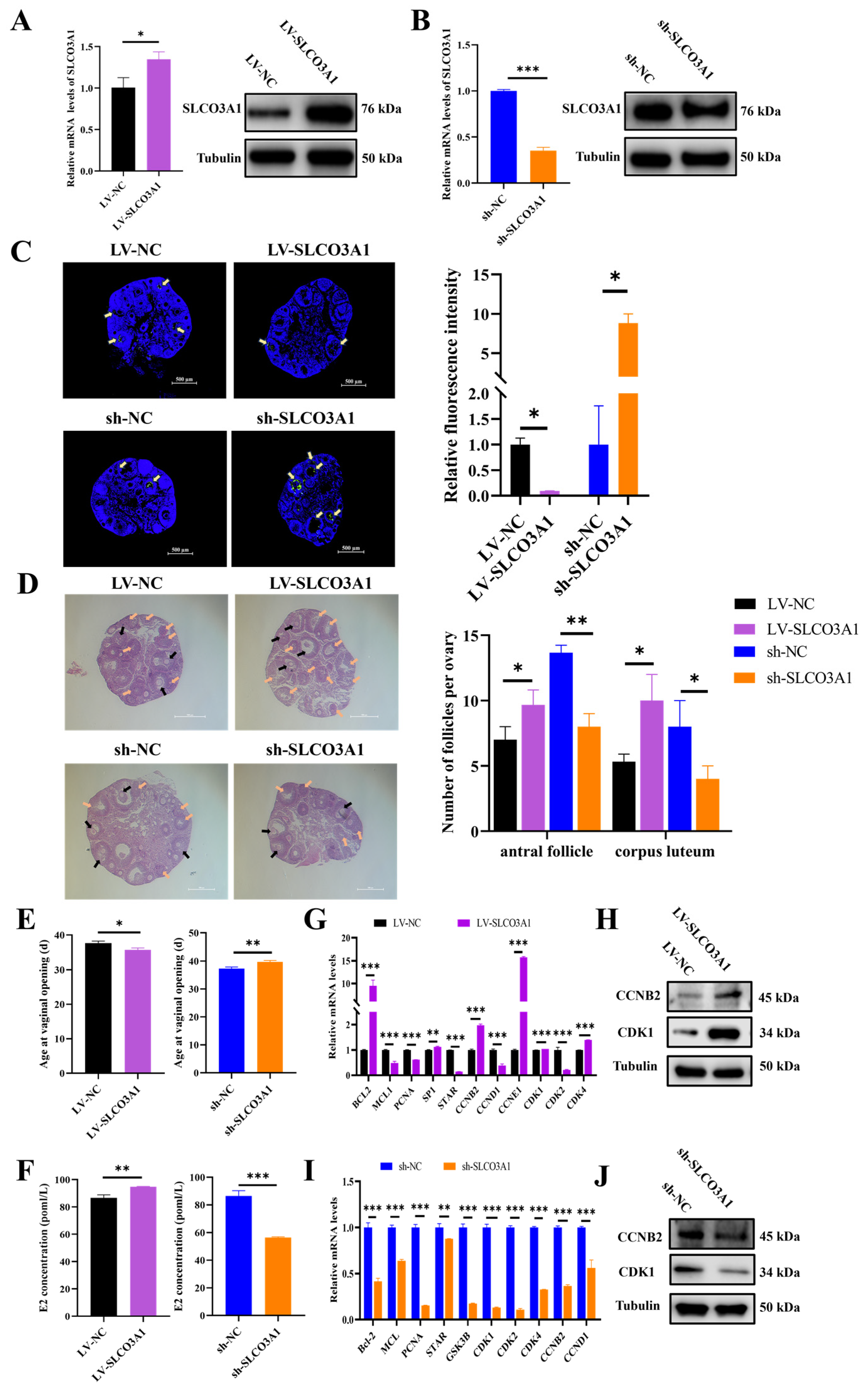
2.3. SLCO3A1 Promotes the Growth of Porcine Follicles
2.4. SLCO3A1 Promotes the Follicular Growth in Mice
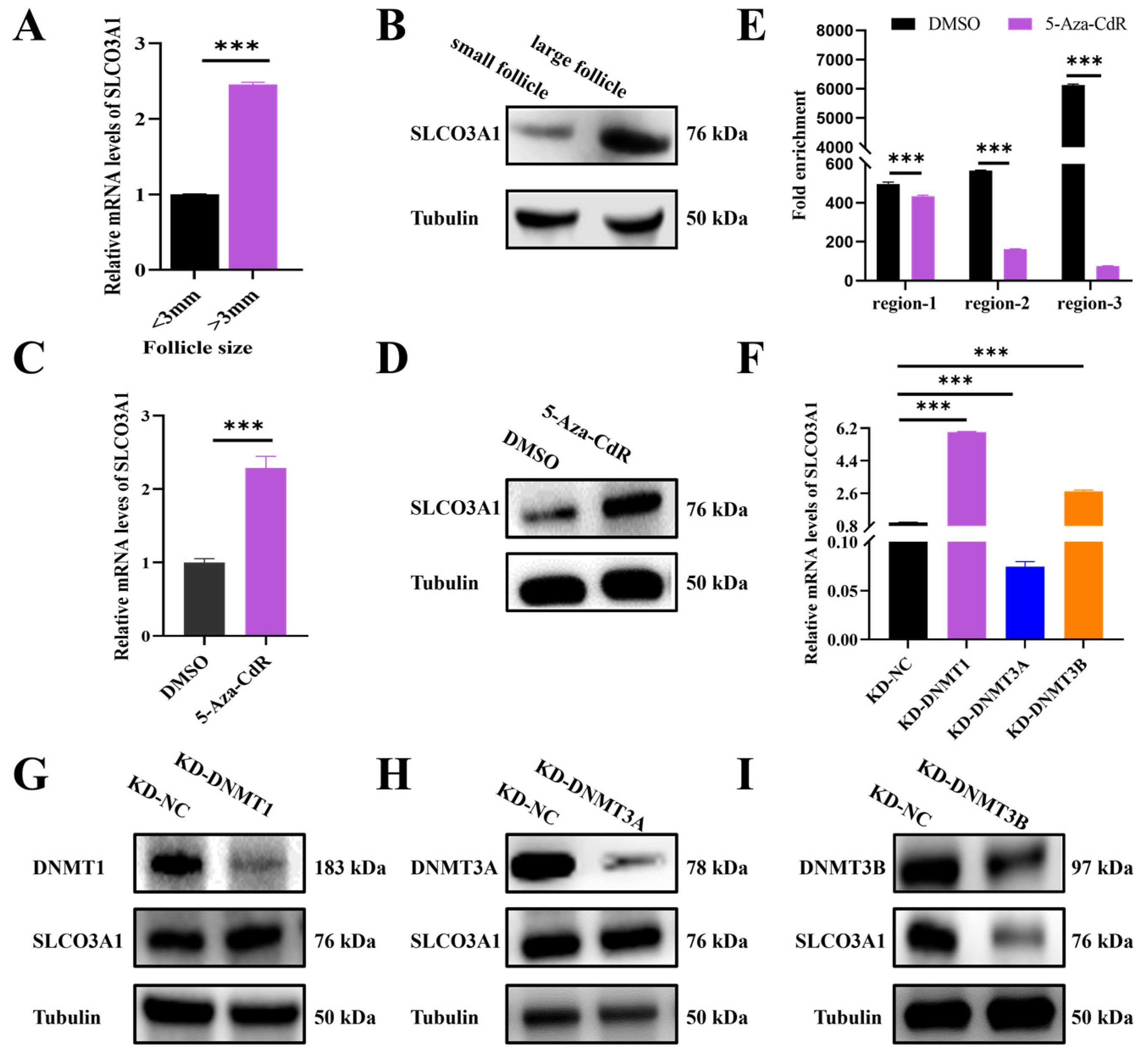
2.5. Knockdown of DNMT1 Upregulates the mRNA and Protein Levels of SLCO3A1
3. Discussion
4. Materials and Methods
4.1. Cells Experiments
4.2. Animal Experiments
4.3. Follicles Culture
4.4. RNA Isolation and Reverse Transcription-Quantitative PCR (RT-qPCR)
4.5. Western Blot Analysis
4.6. Proliferation Assay
4.7. Flow Cytometry Assay
4.8. HE Staining
4.9. TUNEL Assay
4.10. Enzyme-Linked Immunosorbent Assay
4.11. Chromatin Accessibility Assay
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murase, T.; Iwase, A.; Komatsu, K.; Bayasula; Nakamura, T.; Osuka, S.; Takikawa, S.; Goto, M.; Kotani, T.; Kikkawa, F. Follicle Dynamics: Visualization and Analysis of Follicle Growth and Maturation Using Murine Ovarian Tissue Culture. J. Assist. Reprod. Genet. 2018, 35, 339–343. [Google Scholar] [CrossRef]
- Li, C.; Liu, Z.; Wu, G.; Zang, Z.; Zhang, J.-Q.; Li, X.; Tao, J.; Shen, M.; Liu, H. FOXO1 Mediates Hypoxia-Induced G0/G1 Arrest in Ovarian Somatic Granulosa Cells by Activating the TP53INP1-P53-CDKN1A Pathway. Development 2021, 148, dev199453. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Xie, Y.; Liu, Y.; Long, S.; Mo, Z. Polycystic Ovarian Syndrome: Correlation between Hyperandrogenism, Insulin Resistance and Obesity. Clin. Chim. Acta 2020, 502, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Cockrem, J.F.; Han, K.-H.; Kim, D.-H.; Jung, M.-H.; Chu, J.-P. Stress-Induced Activation of Ovarian Heat Shock Protein 90 in a Rat Model of Polycystic Ovary Syndrome. J. Obstet. Gynaecol. Res. 2012, 38, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Yuan, Z.; Li, M.; Zhang, Y.; Nan, F. Decreased Oocyte Quality in Patients with Endometriosis Is Closely Related to Abnormal Granulosa Cells. Front. Endocrinol. 2023, 14, 1226687. [Google Scholar] [CrossRef]
- Hipp, H.S.; Kawwass, J.F. Discordant Ovarian Reserve Testing: What Matters Most? Fertil. Steril. 2019, 112, 34. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, F.; Inoue, N.; Manabe, N.; Ohkura, S. Follicular Growth and Atresia in Mammalian Ovaries: Regulation by Survival and Death of Granulosa Cells. J. Reprod. Dev. 2012, 58, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Skinner, M.K. Regulation of Primordial Follicle Assembly and Development. Human Reprod. Update 2005, 11, 461–471. [Google Scholar] [CrossRef]
- Vendola, K.; Zhou, J.; Wang, J.; Famuyiwa, O.A.; Bievre, M.; Bondy, C.A. Androgens Promote Oocyte Insulin-Like Growth Factor I Expression and Initiation of Follicle Development in the Primate Ovary1. Biol. Reprod. 1999, 61, 353–357. [Google Scholar] [CrossRef]
- Gougeon, A. Regulation of Ovarian Follicular Development in Primates: Facts and Hypotheses. Endocr. Rev. 1996, 17, 121–155. [Google Scholar] [CrossRef]
- Grant, S.A.; Hunter, M.G.; Foxcroft, G.R. Morphological and Biochemical Characteristics during Ovarian Follicular Development in the Pig. J. Reprod. Fertil. 1989, 86, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, H.D.; Cooper, B.S.; Welch, G.R.; Zakaria, A.D.; Johnson, L.A. Atresia in Follicles Grown after Ovulation in the Pig: Measurement of Increased Apoptosis in Granulosa Cells and Reduced Follicular Fluid Estradiol-17 Beta. Biol. Reprod. 1995, 52, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Veazey, K.J.; Wang, H.; Bedi, Y.S.; Skiles, W.M.; Chang, R.C.-A.; Golding, M.C. Disconnect between Alcohol-Induced Alterations in Chromatin Structure and Gene Transcription in a Mouse Embryonic Stem Cell Model of Exposure. Alcohol 2017, 60, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, B.E.; Meissner, A.; Lander, E.S. The Mammalian Epigenome. Cell 2007, 128, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, A.D.; Allis, C.D.; Bernstein, E. Epigenetics: A Landscape Takes Shape. Cell 2007, 128, 635–638. [Google Scholar] [CrossRef]
- LaVoie, H.A. Epigenetic Control of Ovarian Function: The Emerging Role of Histone Modifications. Mol. Cell. Endocrinol. 2005, 243, 12–18. [Google Scholar] [CrossRef]
- Pan, Z.; Zhang, J.; Li, Q.; Li, Y.; Shi, F.; Xie, Z.; Liu, H. Current Advances in Epigenetic Modification and Alteration during Mammalian Ovarian Folliculogenesis. J. Genet. Genom. 2012, 39, 111–123. [Google Scholar] [CrossRef]
- Zhou, X.; He, Y.; Li, N.; Bai, G.; Pan, X.; Zhang, Z.; Zhang, H.; Li, J.; Yuan, X. DNA Methylation Mediated RSPO2 to Promote Follicular Development in Mammals. Cell Death Dis. 2021, 12, 653. [Google Scholar] [CrossRef]
- Han, C.; Wei, Y.; Geng, Y.; Cui, Y.; Li, S.; Bao, Y.; Shi, W. Bisphenol A in Utero Exposure Induces Ovary Dysfunction in Mice Offspring and the Ameliorating Effects of Cuscuta Chinensis Flavonoids. Environ. Sci. Pollut. Res. Int. 2020, 27, 31357–31368. [Google Scholar] [CrossRef]
- Zhou, X.; He, Y.; Pan, X.; Quan, H.; He, B.; Li, Y.; Bai, G.; Li, N.; Zhang, Z.; Zhang, H.; et al. DNMT1-Mediated lncRNA IFFD Controls the Follicular Development via Targeting GLI1 by Sponging miR-370. Cell Death Differ. 2023, 30, 576–588. [Google Scholar] [CrossRef]
- Adachi, H.; Suzuki, T.; Abe, M.; Asano, N.; Mizutamari, H.; Tanemoto, M.; Nishio, T.; Onogawa, T.; Toyohara, T.; Kasai, S.; et al. Molecular Characterization of Human and Rat Organic Anion Transporter OATP-D. Am. J. Physiol. Ren. Physiol. 2003, 285, F1188–F1197. [Google Scholar] [CrossRef]
- Nozawa, T.; Suzuki, M.; Yabuuchi, H.; Irokawa, M.; Tsuji, A.; Tamai, I. Suppression of Cell Proliferation by Inhibition of Estrone-3-Sulfate Transporter in Estrogen-Dependent Breast Cancer Cells. Pharm. Res. 2005, 22, 1634–1641. [Google Scholar] [CrossRef]
- Yi, W.; Li, X.; Chen, K.; Li, J.; Chen, K.; Pan, A. Effect of rNA Interference on Oatp3a1 Gene Expression on Biological Characteristics and Immune Factors of Ovarian Granulosa Cells in Rats with PCOS. Am. J. Transl. Res. 2020, 12, 4659–4668. [Google Scholar]
- Byers, S.L.; Wiles, M.V.; Dunn, S.L.; Taft, R.A. Mouse Estrous Cycle Identification Tool and Images. PLoS ONE 2012, 7, e35538. [Google Scholar] [CrossRef]
- Zhang, Q.; Lei, Y.; Deng, Y.; Ma, R.; Ding, X.; Xue, W.; Sun, A. Treatment Progress in Diminished Ovarian Reserve: Western and Chinese Medicine. Chin. J. Integr. Med. 2023, 29, 361–367. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, J.; Wang, W.; Zhao, H.; Liu, H.; Liu, X.; Liu, J.; Sun, Y.; Dunaif, A.; Du, Y.; et al. Long Non-Coding RNA LINC-01572:28 Inhibits Granulosa Cell Growth via a Decrease in P27 (Kip1) Degradation in Patients with Polycystic Ovary Syndrome. EBioMedicine 2018, 36, 526–538. [Google Scholar] [CrossRef]
- Geng, X.; Zhao, J.; Huang, J.; Li, S.; Chu, W.; Wang, W.-S.; Chen, Z.-J.; Du, Y. Lnc-MAP3K13-7:1 Inhibits Ovarian GC Proliferation in PCOS via DNMT1 Downregulation-Mediated CDKN1A Promoter Hypomethylation. Mol. Ther. 2021, 29, 1279–1293. [Google Scholar] [CrossRef]
- Maalouf, S.W.; Liu, W.S.; Pate, J.L. MicroRNA in Ovarian Function. Cell Tissue Res. 2016, 363, 7–18. [Google Scholar] [CrossRef]
- Emmen, J.M.A.; Couse, J.F.; Elmore, S.A.; Yates, M.M.; Kissling, G.E.; Korach, K.S. In Vitro Growth and Ovulation of Follicles from Ovaries of Estrogen Receptor (ER)α and ERβ Null Mice Indicate a Role for ERβ in Follicular Maturation. Endocrinology 2005, 146, 2817–2826. [Google Scholar] [CrossRef]
- Khristi, V.; Chakravarthi, V.P.; Singh, P.; Ghosh, S.; Pramanik, A.; Ratri, A.; Borosha, S.; Roby, K.F.; Wolfe, M.W.; Rumi, M.A.K. ESR2 Regulates Granulosa Cell Genes Essential for Follicle Maturation and Ovulation. Mol. Cell Endocrinol. 2018, 474, 214–226. [Google Scholar] [CrossRef]
- Zhang, S.; Tu, H.; Zhu, J.; Liang, A.; Huo, P.; Shan, K.; He, J.; Zhao, M.; Chen, X.; Lei, X. Dendrobium Nobile Lindl. Polysaccharides Improve Follicular Development in PCOS Rats. Int. J. Biol. Macromol. 2020, 149, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Alenzi, F.Q.B. Links between Apoptosis, Proliferation and the Cell Cycle. Br. J. Biomed. Sci. 2004, 61, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Farshadi, E.; van der Horst, G.T.J.; Chaves, I. Molecular Links between the Circadian Clock and the Cell Cycle. J. Mol. Biol. 2020, 432, 3515–3524. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.-L.; Wu, Q.-F.; Xie, Q.; Tan, J.; Shu, K.-Y. PATL2 Regulated the Apoptosis of Ovarian Granulosa Cells in Patients with PCOS. Gynecol. Endocrinol. 2021, 37, 629–634. [Google Scholar] [CrossRef]
- Gong, Y.; Luo, S.; Fan, P.; Zhu, H.; Li, Y.; Huang, W. Growth Hormone Activates PI3K/Akt Signaling and Inhibits ROS Accumulation and Apoptosis in Granulosa Cells of Patients with Polycystic Ovary Syndrome. Reprod. Biol. Endocrinol. 2020, 18, 121. [Google Scholar] [CrossRef]
- Honnma, H.; Endo, T.; Henmi, H.; Nagasawa, K.; Baba, T.; Yamazaki, K.; Kitajima, Y.; Hayashi, T.; Manase, K.; Saito, T. Altered Expression of Fas/Fas Ligand/Caspase 8 and Membrane Type 1-Matrix Metalloproteinase in Atretic Follicles within Dehydroepiandrosterone-Induced Polycystic Ovaries in Rats. Apoptosis 2006, 11, 1525–1533. [Google Scholar] [CrossRef]
- Yuan, X.; Li, Z.; Kong, Y.; Zhong, Y.; He, Y.; Zhang, A.; Zhou, X.; Jiang, Y.; Zhang, Z.; Zhang, H.; et al. P65 Targets FGFR1 to Regulate the Survival of Ovarian Granulosa Cells. Cells 2019, 8, 1334. [Google Scholar] [CrossRef]
- Li, N.; Zhou, Y.; Cai, J.; Wang, Y.; Zhou, X.; Hu, M.; Li, Y.; Zhang, H.; Li, J.; Cai, B.; et al. A Novel Trans-Acting lncRNA of ACTG1 That Induces the Remodeling of Ovarian Follicles. Int. J. Biol. Macromol. 2023, 242, 125170. [Google Scholar] [CrossRef]
- Plaza-Parrochia, F.; Poblete, C.; Gabler, F.; Carvajal, R.; Romero, C.; Valladares, L.; Vega, M. Expression of Steroid Sulfated Transporters and 3β-HSD Activity in Endometrium of Women Having Polycystic Ovary Syndrome. Steroids 2015, 104, 189–195. [Google Scholar] [CrossRef]
- Guo, X.; Puttabyatappa, M.; Thompson, R.C.; Padmanabhan, V. Developmental Programming: Contribution of Epigenetic Enzymes to Antral Follicular Defects in the Sheep Model of PCOS. Endocrinology 2019, 160, 2471–2484. [Google Scholar] [CrossRef]
- Fang, Y.; Deng, S.; Zhang, J.; Liu, H.; Li, Y.; Zhang, X.; Liu, Y. Melatonin-Mediated Development of Ovine Cumulus Cells, Perhaps by Regulation of DNA Methylation. Molecules 2018, 23, 494. [Google Scholar] [CrossRef] [PubMed]
- He, Z.-Y.; Wang, H.-Y.; Zhou, X.; Liang, X.-Y.; Yan, B.; Wang, R.; Ma, L.-H.; Wang, Y.-L. Evaluation of Vitrification Protocol of Mouse Ovarian Tissue by Effect of DNA Methyltransferase-1 and Paternal Imprinted Growth Factor Receptor-Binding Protein 10 on Signaling Pathways. Cryobiology 2018, 80, 89–95. [Google Scholar] [CrossRef] [PubMed]
- do Amaral, G.C.L.S.; Planello, A.C.; Borgato, G.; de Lima, D.G.; Guimarães, G.N.; Marques, M.R.; de Souza, A.P. 5-Aza-CdR Promotes Partial MGMT Demethylation and Modifies Expression of Different Genes in Oral Squamous Cell Carcinoma. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2019, 127, 425–432. [Google Scholar] [CrossRef]
- Nikbakht Dastjerdi, M.; Azarnezhad, A.; Hashemibeni, B.; Salehi, M.; Kazemi, M.; Babazadeh, Z. An Effective Concentration of 5-Aza-CdR to Induce Cell Death and Apoptosis in Human Pancreatic Cancer Cell Line through Reactivating RASSF1A and Up-Regulation of Bax Genes. Iran. J. Med. Sci. 2018, 43, 533–540. [Google Scholar]
- Baylin, S.B.; Herman, J.G. DNA Hypermethylation in Tumorigenesis: Epigenetics Joins Genetics. Trends Genet. 2000, 16, 168–174. [Google Scholar] [CrossRef]
- Liu, H.; Zhou, J.; Tian, W.; Luo, C.; Bartlett, A.; Aldridge, A.; Lucero, J.; Osteen, J.K.; Nery, J.R.; Chen, H.; et al. DNA Methylation Atlas of the Mouse Brain at Single-Cell Resolution. Nature 2021, 598, 120–128. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence Number | Target Sequence (5′→3′) |
|---|---|
| SLCO3A1-siRNA # 1 | GCCCTGAACTCAAGTCTTA |
| SLCO3A1-siRNA # 2 | CCCACCAGTACAAGTACGA |
| SLCO3A1-siRNA # 3 | ACCTGCTCTCAAACCCTGT |
| siRNA-DNMT1 | GGAAGAAGAGUUACUAUAA |
| siRNA-DNMT3A | GCCUCAGAGCUAUUACCCA |
| siRNA-DNMT3B | GAAGAUCAAGCUCGCGACU |
| Gene Name | Primer Sequences (5′ to 3′) | Size (bp) | Accession Number |
|---|---|---|---|
| SLCO3A1 | F: GTGGGGTGGCTTTCTGCTCT | 145 | NM_013272.4 |
| R: GGGTCTCTCGTATTCTCTTTCGG | |||
| PCNA | F: CAAGTAATGTCGATAAAGAGGAGG | 126 | NM_182649.2 |
| R: GTGTCACCGTTGAAGAGAGTGG | |||
| BCL-2 | F: ATCGCCCTGTGGATGACTGAGT | 127 | NM_000633.3 |
| R: GCCAGGAGAAATCAAACAGAGGC | |||
| IKBA | F: TCCACTCCATCCTGAAGGCTAC | 101 | NM_020529.3 |
| R: CAAGGACACCAAAAGCTCCACG | |||
| STAR | F: TACGTGGCTACTCAGCATCGAC | 142 | NM_000349.3 |
| R: TCAACACCTGGCTTCAGAGGCA | |||
| MCL1 | F: CCAAGAAAGCTGCATCGAACCAT | 151 | NM_001197320.2 |
| R: CAGCACATTCCTGATGCCACCT | |||
| ESR1 | F: GCTTACTGACCAACCTGGCAGA | 129 | XM_054354493.1 |
| R: GGATCTCTAGCCAGGCACATTC | |||
| ESR2 | F: ATGGAGTCTGGTCGTGTGAAGG | 148 | NM_001437.3 |
| R: TAACACTTCCGAAGTCGGCAGG | |||
| FSHR | F: GGTTTGTCCTCACCAAGCTTCG | 126 | NM_000145.4 |
| R: GGTTGGAGAACACATCTGCCTC | |||
| CYP1A1 | F: GATTGAGCACTGTCAGGAGAAGC | 138 | NM_000499.5 |
| R: ATGAGGCTCCAGGAGATAGCAG | |||
| CYP17A1 | F: GCACACCAACTATCAGTGACCG | 147 | NM_000102.4 |
| R: CCTTGTCCACAGCAAACTCACC | |||
| CYP19A1 | F: GACGCAGGATTTCCACAGAAGAG | 145 | NM_001347252.2 |
| R: ATGGTGTCAGGAGCTGCGATCA | |||
| HSD17B1 | F: TTCCTGCCAGACATGAAGAGGC | 143 | NM_001330219.3 |
| R: AGAACCGCCAGACTCTCGCATA | |||
| MYC | F: CCTGGTGCTCCATGAGGAGAC | 128 | NM_002467.6 |
| R: CAGACTCTGACCTTTTGCCAGG | |||
| PAK1 | F: GTGAAGGCTGTGTCTGAGACTC | 149 | NM_001376274.1 |
| R: GGAAGTGGTTCAATCACAGACCG | |||
| CDKN1B | F: ATAAGGAAGCGACCTGCAACCG | 119 | NM_004064.5 |
| R: TTCTTGGGCGTCTGCTCCACAG | |||
| CCNB2 | F: CAACCAGAGCAGCACAAGTAGC | 136 | NM_004701.4 |
| R: GGAGCCAACTTTTCCATCTGTAC | |||
| CCND1 | F: TCTACACCGACAACTCCATCCG | 133 | NM_053056.3 |
| R: TCTGGCATTTTGGAGAGGAAGTG | |||
| CCNE1 | F: TGTGTCCTGGATGTTGACTGCC | 123 | NM_001322262.2 |
| R: CTCTATGTCGCACCACTGATACC | |||
| CCNE2 | F: CTTACGTCACTGATGGTGCTTGC | 126 | NM_057749.3 |
| R: CTTGGAGAAAGAGATTTAGCCAGG | |||
| CCNH | F: CGATGTCATTCTGCTGAGCTTGC | 128 | NM_001199189.2 |
| R: TCTACCAGGTCGTCATCAGTCC | |||
| CDK1 | F: GGAAACCAGGAAGCCTAGCATC | 124 | NM_001320918.1 |
| R: GGATGATTCAGTGCCATTTTGCC | |||
| CDK2 | F: ATGGATGCCTCTGCTCTCACTG | 97 | NM_052827.4 |
| R: CCCGATGAGAATGGCAGAAAGC | |||
| CDK4 | F: CCATCAGCACAGTTCGTGAGGT | 103 | NM_000075.4 |
| R: TCAGTTCGGGATGTGGCACAGA | |||
| P53 | F: CCTCAGCATCTTATCCGAGTGG | 128 | NM_001407269.1 |
| R: TGGATGGTGGTACAGTCAGAGC | |||
| BAD | F: CCAACCTCTGGGCAGCACAGC | 126 | NM_032989.3 |
| R: TTTGCCGCATCTGCGTTGCTGT | |||
| BID | F: TGGGACACTGTGAACCAGGAGT | 125 | NM_197966.3 |
| R: GAGGAAGCCAAACACCAGTAGG | |||
| CASP3 | F: GGAAGCGAATCAATGGACTCTGG | 146 | NM_001354783.2 |
| R: GCATCGACATCTGTACCAGACC | |||
| CASP7 | F: CGGAACAGACAAAGATGCCGAG | 143 | NM_033338.6 |
| R: AGGCGGCATTTGTATGGTCCTC | |||
| CASP8 | F: GCTGACTTTCTGCTGGGGAT | 112 | NM_033355.4 |
| R: GACATCGCTCTCTCAGGCTC | |||
| CASP9 | F: GTTTGAGGACCTTCGACCAGCT | 129 | NM_001278054.2 |
| R: CAACGTACCAGGAGCCACTCTT | |||
| GAPDH | F: GTCTCCTCTGACTTCAACAGCG | 131 | NM_002046.7 |
| R: ACCACCCTGTTGCTGTAGCCAA |
| Gene Name | Primer Sequences (5′ to 3′) | Size (bp) | Accession Number |
|---|---|---|---|
| SLCO3A1 | F: GTGGGGTGGCTTTCTGCTCT | 145 | XM_021099372.1 |
| R: GGGTCTCTCTATTCTCTTTCGG | |||
| CCND1 | F: TTCATTTCCAACCCGCCCTC | 185 | XM_021082686.1 |
| R: TCCAGAAGGGCTTCGATCTG | |||
| CDK1 | F: AAGTGTGGCCAGAAGTGGAG | 157 | XM_005671016.3 |
| R: CCAGAAATTCGCTTGGCAGG | |||
| CDK2 | F: TTTGCTGAGATGGTGACCCG | 254 | NM_001285465.1 |
| R: GCTGAAATCCGCTTGTTGGG | |||
| CDK4 | F: TTGTCCGGCTGATGGATGTC | 255 | NM_001123097.1 |
| R: GCTTGACTGTCCCACCACTT | |||
| CCNE1 | F: ACTGATGTCTCTGTTCGCTCC | 175 | XM_005653265.2 |
| R: TGTCAGGTGTGGGAATGAAGG | |||
| BCL-2 | F: GAGTTCGGTGGGGTCATGTG | 152 | XM_021099593.1 |
| R: TACAGCTCCACAAAGGCATCC | |||
| MCL1 | F: GGAAGGCGTTAGAGACCCTG | 178 | NM_001348806.1 |
| R: GTCACAATCCTGCCCCAGTT | |||
| STAR | F: AGACTTTGTGAGTGTGCGCT | 254 | NM_213755.2 |
| R: AGTCCACCTGGGTCTGTGAT | |||
| P65 | F: CATGCGCTTCCGCTACAAG | 284 | NM_001114281.1 |
| R: GGTCCCGCTTCTTTACACAC | |||
| PCNA | F: AAGAGGAGGAAGCAGTTACCA | 205 | NM_001291925.1 |
| R: TCATCTTCGATCTTGGGAGCC | |||
| GAPDH | F: GGACTCATGACCACGGTCCAT | 220 | XM_021091114.1 |
| R: TCAGATCCACAACCGACACGT |
| Gene Name | Primer Sequences (5′ to 3′) | Size (bp) | Accession Number |
|---|---|---|---|
| SLCO3A1 | F: CTGCTCCTTCGTTTGTTGGG | 279 | XM_021167570.2 |
| R: AGAGGCAAAGAACTCACTGGT | |||
| BCL-2 | F: CCTGTGGATGACTGAGTACCTG | 123 | XM_029538378.1 |
| R: AGCCAGGAGAAATCAAACAGAGG | |||
| MCL1 | F: AGCTTCATCGAACCATTAGCAGAA | 125 | NM_008562.3 |
| R: CCTTCTAGGTCCTGTACGTGGA | |||
| PCNA | F: CAAGTGGAGAGCTTGGCAATGG | 112 | NM_011045.2 |
| R: GCAAACGTTAGGTGAACAGGCTC | |||
| SP1 | F: CTCCAGACCATTAACCTCAGTGC | 133 | XM_029548031.1 |
| R: CACCACCAGATCCATGAAGACC | |||
| STAR | F: GTGCTTCATCCACTGGCTGGAA | 113 | NM_011485.5 |
| R: GTCTGCGATAGGACCTGGTTGA | |||
| CCNB2 | F: GCACTACCATCCTTCTCAGGTG | 137 | NM_007630.2 |
| R: TGTGCTGCATGACTTCCAGGAC | |||
| CCND1 | F: GCAGAAGGAGATTGTGCCATCC | 123 | NM_001379248.1 |
| R: AGGAAGCGGTCCAGGTAGTTCA | |||
| CCNE1 | F: AAGCCCTCTGACCATTGTGTCC | 155 | XM_021201435.2 |
| R: CTAAGCAGCCAACATCCAGGAC | |||
| CDK1 | F: CATGGACCTCAAGAAGTACCTGG | 136 | NM_007659.4 |
| R: CAAGTCTCTGTGAAGAACTCGCC | |||
| CDK2 | F: TCATGGATGCCTCTGCTCTCAC | 106 | NM_016756.4 |
| R: TGAAGGACACGGTGAGAATGGC | |||
| CDK4 | F: CATACCTGGACAAAGCACCTCC | 135 | XM_029482281.1 |
| R: GAATGTTCTCTGGCTTCAGGTCC | |||
| GAPDH | F: GGACTCATGACCACGGTCCAT | 220 | NM_008084.4 |
| R: TCAGATCCACAACCGACACGT |
| Name | Primer Sequences (5′ to 3′) | Size (bp) |
|---|---|---|
| region-1 | F: GGGGAGTAATTAGGAGCGGC | 282 |
| R: ACCGAAAGGACAGAAAGGGG | ||
| region-2 | F: TCTCCCTTCCTGGGAACAGC | 125 |
| R: GAGACTGGCGGCTCTGC | ||
| region-3 | F: CACCCCCCTTACTCCGCAA | 209 |
| R: GCTCTCACCCCCTCAGCCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Zeng, L.; Miao, F.; Li, N.; Liao, W.; Zhou, X.; Chen, Y.; Quan, H.; He, Y.; Zhang, H.; et al. Knockdown of DNMT1 Induces SLCO3A1 to Promote Follicular Growth by Enhancing the Proliferation of Granulosa Cells in Mammals. Int. J. Mol. Sci. 2024, 25, 2468. https://doi.org/10.3390/ijms25052468
Li S, Zeng L, Miao F, Li N, Liao W, Zhou X, Chen Y, Quan H, He Y, Zhang H, et al. Knockdown of DNMT1 Induces SLCO3A1 to Promote Follicular Growth by Enhancing the Proliferation of Granulosa Cells in Mammals. International Journal of Molecular Sciences. 2024; 25(5):2468. https://doi.org/10.3390/ijms25052468
Chicago/Turabian StyleLi, Shuo, Liqing Zeng, Fen Miao, Nian Li, Weili Liao, Xiaofeng Zhou, Yongcai Chen, Hongyan Quan, Yingting He, Hao Zhang, and et al. 2024. "Knockdown of DNMT1 Induces SLCO3A1 to Promote Follicular Growth by Enhancing the Proliferation of Granulosa Cells in Mammals" International Journal of Molecular Sciences 25, no. 5: 2468. https://doi.org/10.3390/ijms25052468
APA StyleLi, S., Zeng, L., Miao, F., Li, N., Liao, W., Zhou, X., Chen, Y., Quan, H., He, Y., Zhang, H., Li, J., & Yuan, X. (2024). Knockdown of DNMT1 Induces SLCO3A1 to Promote Follicular Growth by Enhancing the Proliferation of Granulosa Cells in Mammals. International Journal of Molecular Sciences, 25(5), 2468. https://doi.org/10.3390/ijms25052468