
Intravenous Administration of Mesenchymal Stem Cell-Derived Exosome Alleviates Spinal Cord Injury by Regulating Neutrophil Extracellular Trap Formation through Exosomal miR-125a-3p

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
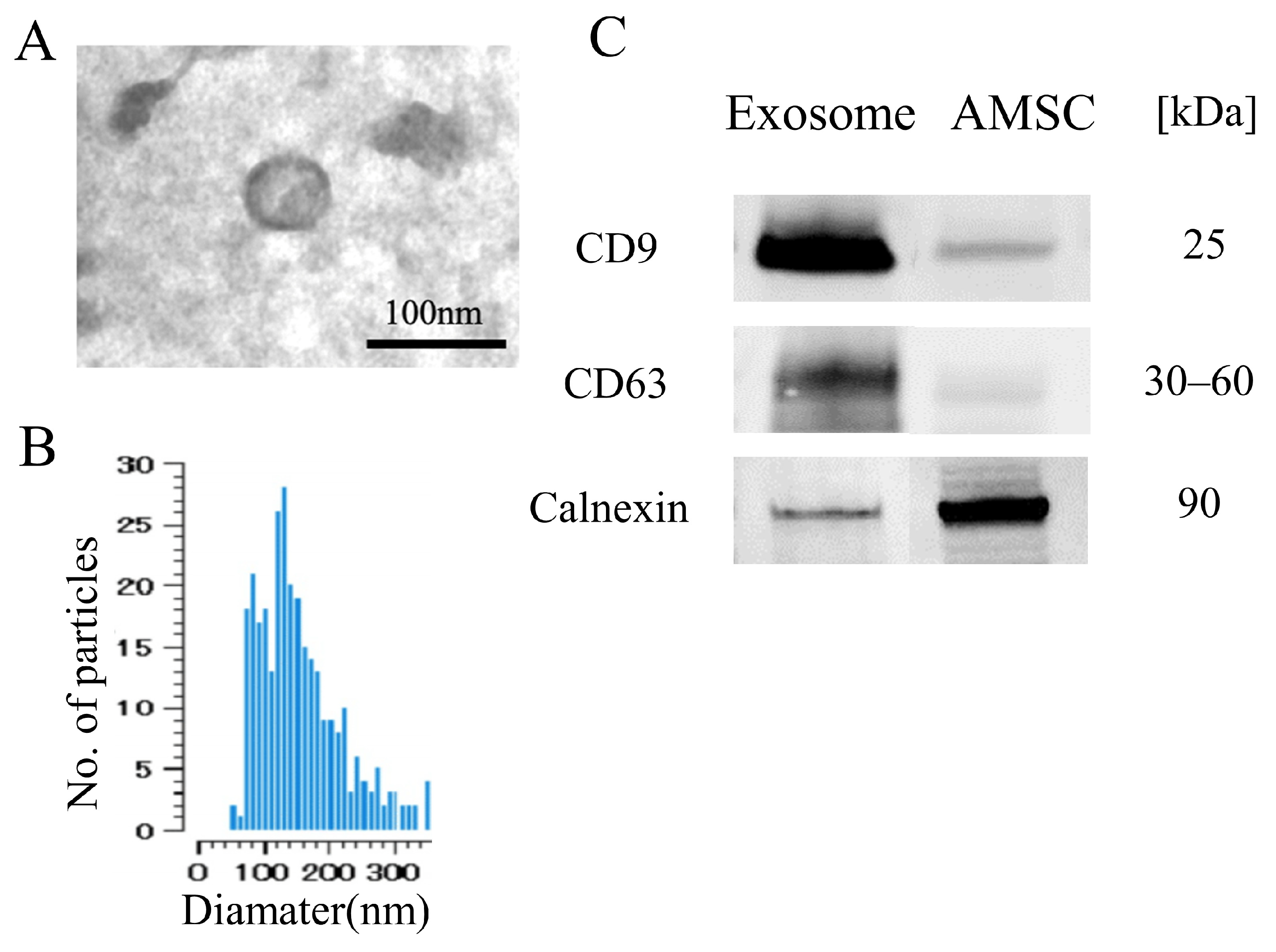
2.1. Exosome Characterization
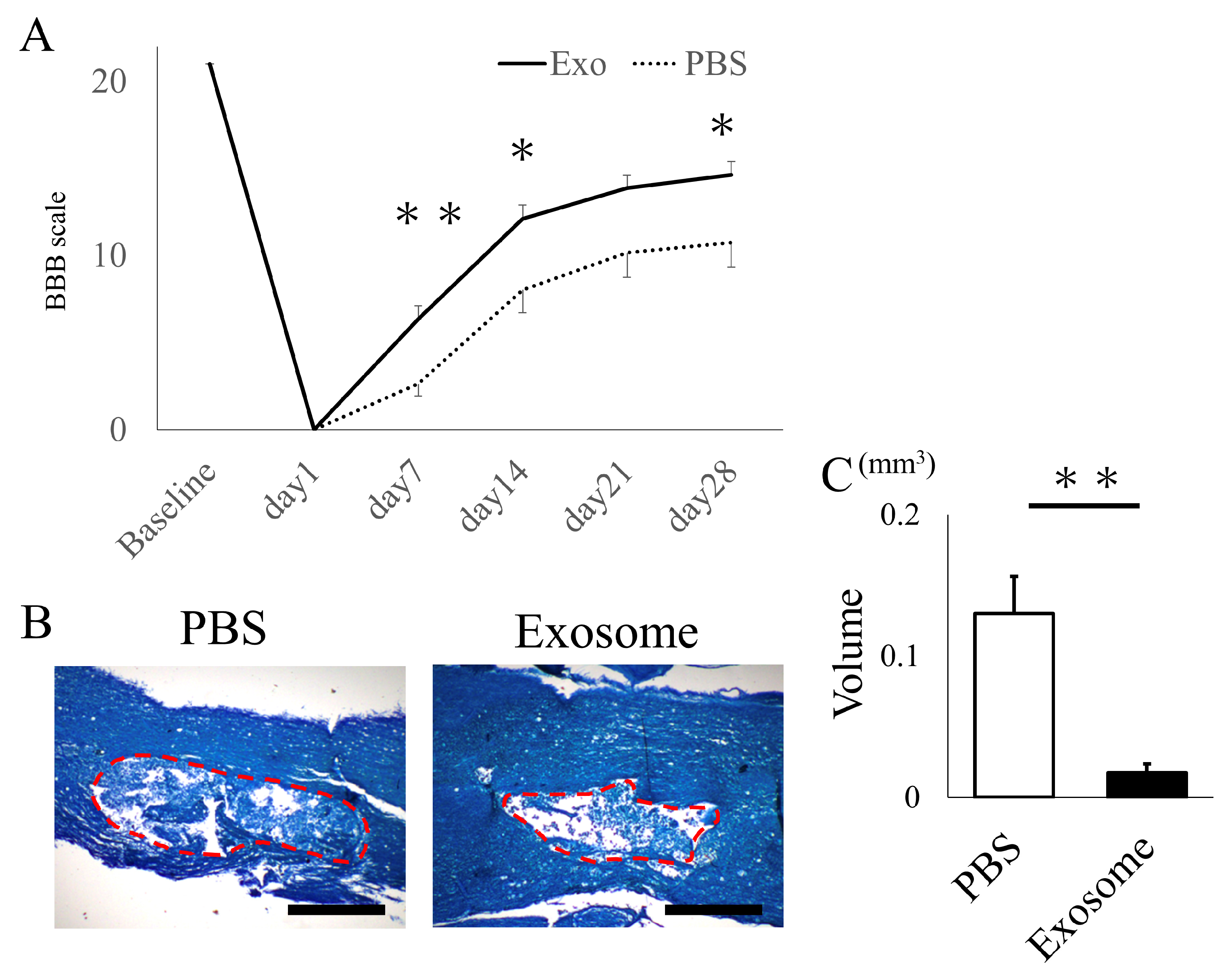
2.2. Intravenous Exosome Administration Ameliorates SCI Damage in Mice
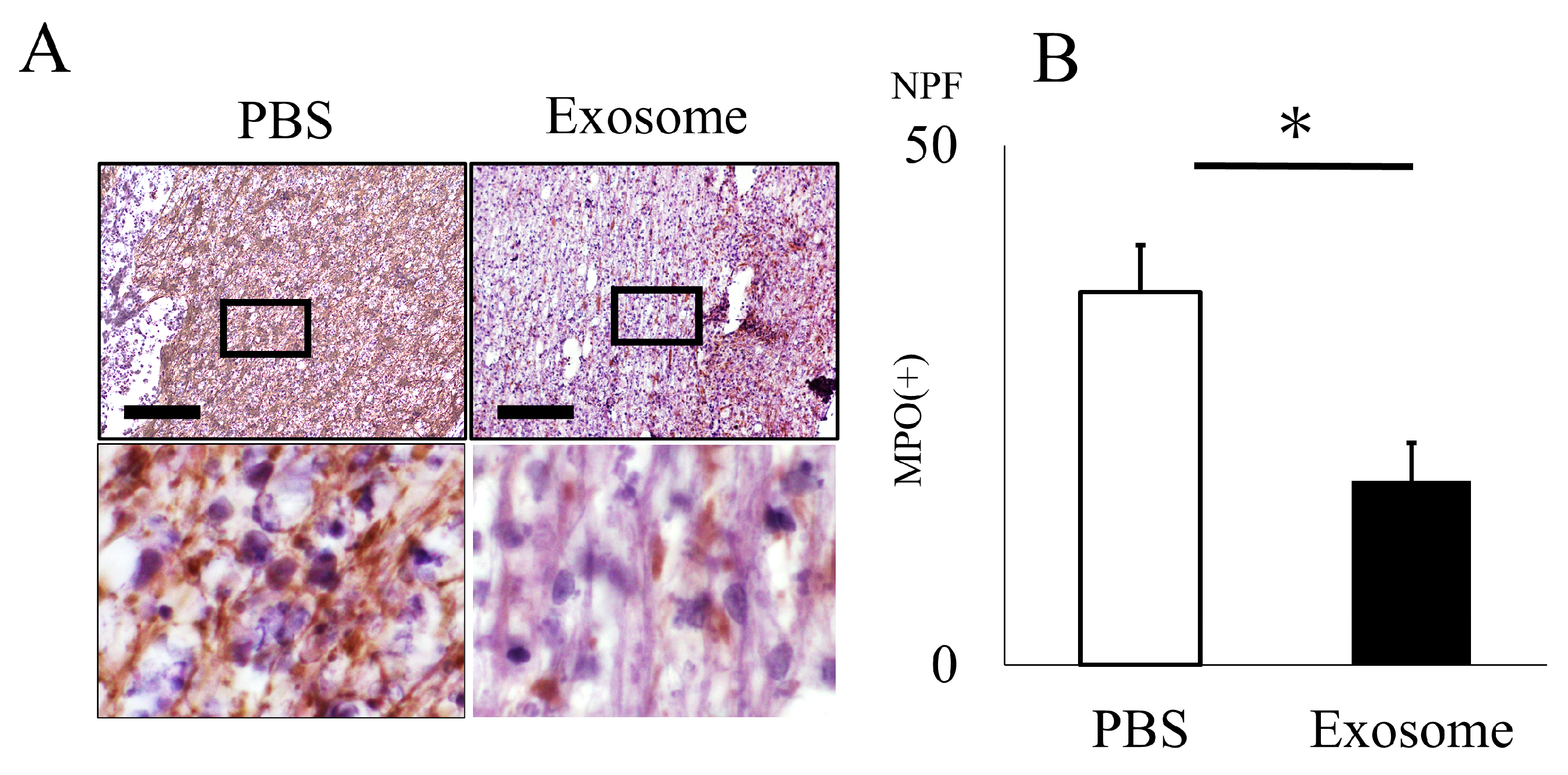
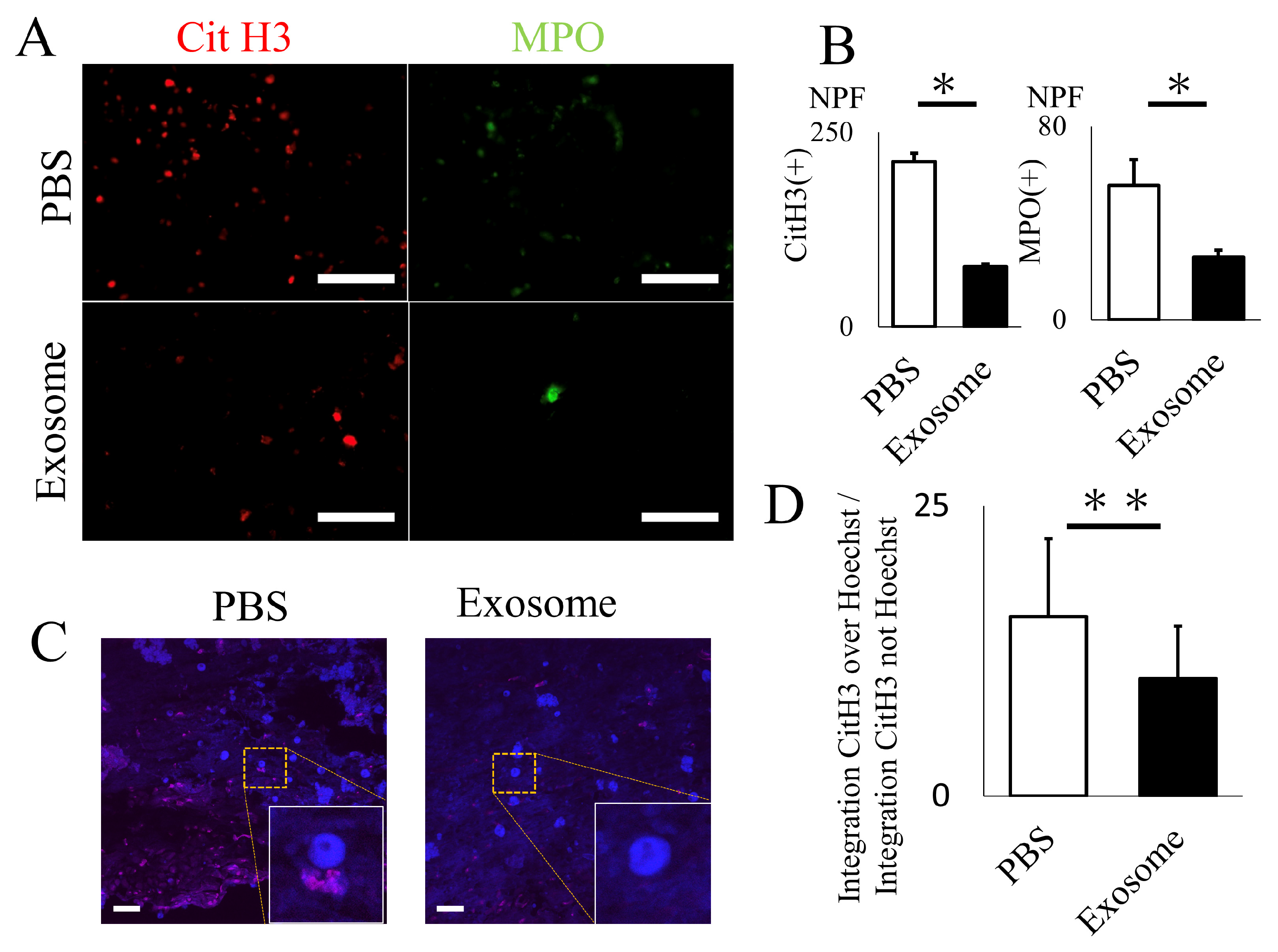
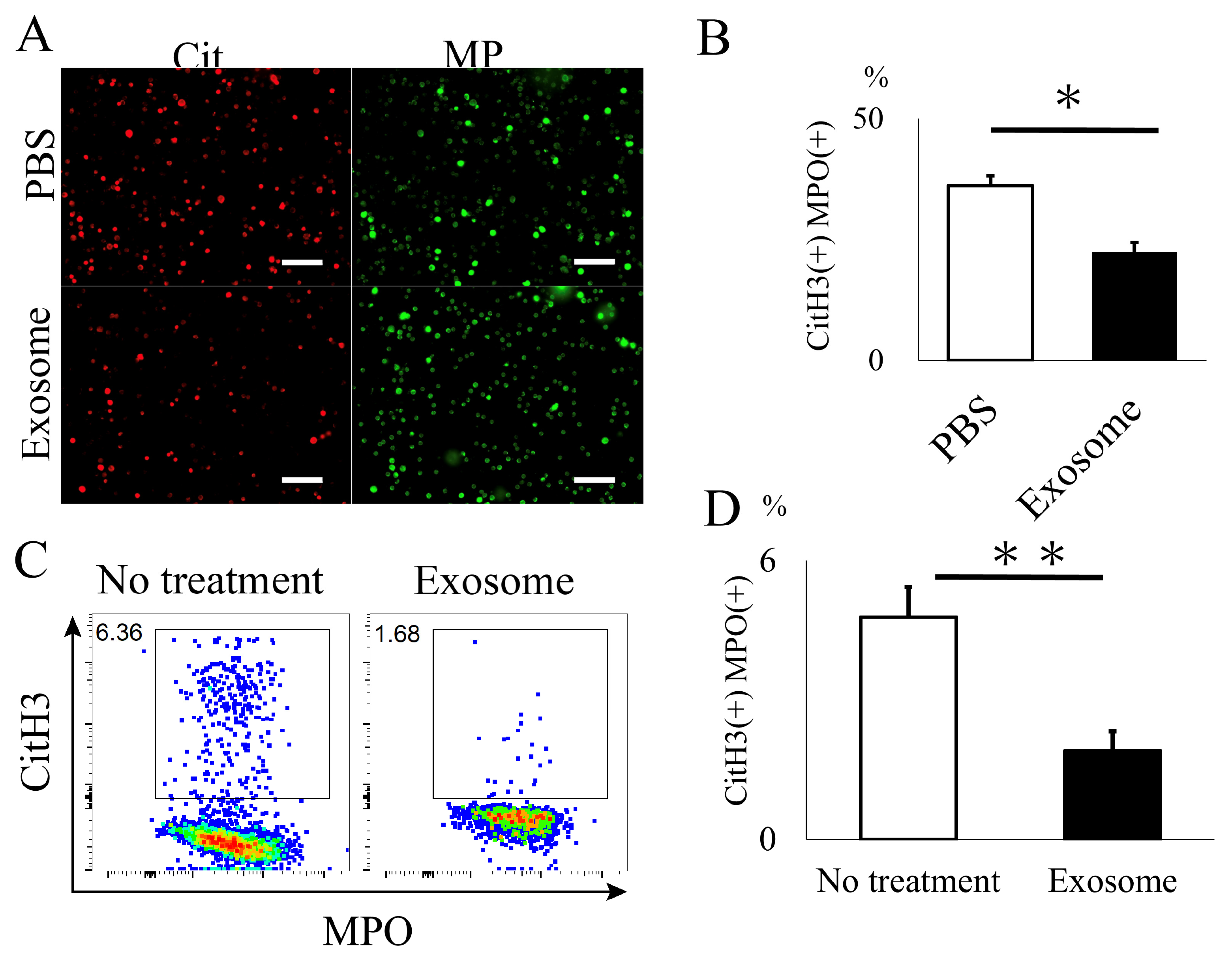
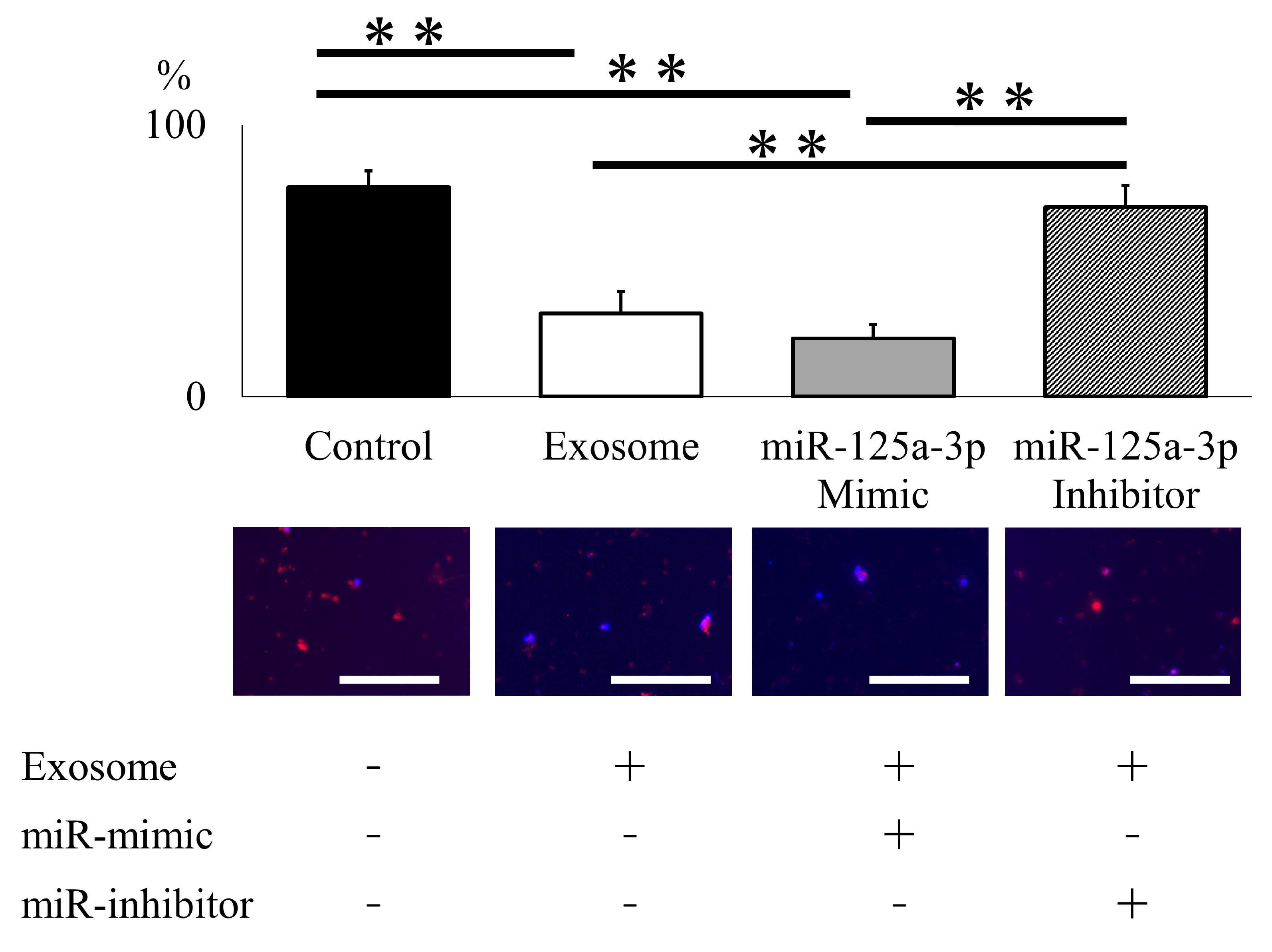
2.3. Ex Vivo Analysis of Exosomes for NET Formation
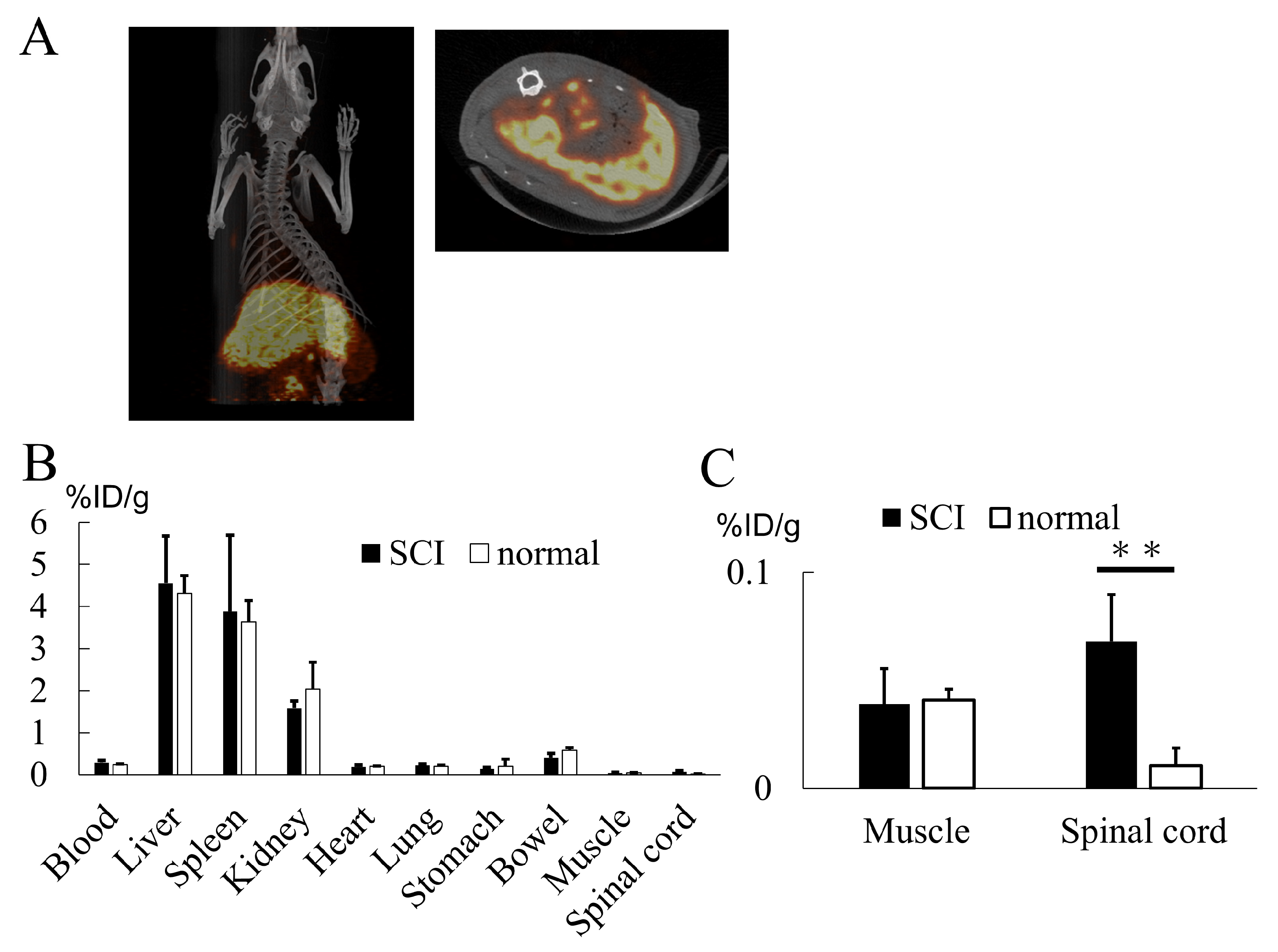
2.4. Biodistribution of Exosomes
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Methods
4.2.1. Culturing and Isolation of AMSC Exosomes
4.2.2. SCI Model and Exosome Administration
4.2.3. Histological Analysis
4.2.4. Ex Vivo Assessment of Neutrophil and NET Formation
4.2.5. Biodistribution
4.2.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bickenbach, J.; Officer, A.; Shakespeare, T.; von Groote, P. International Perspectives on Spinal Cord Injury, 1st ed.; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Feng, Z.; Min, L.; Liang, L.; Chen, B.; Chen, H.; Zhou, Y.; Deng, W.; Liu, H.; Hou, J. Neutrophil Extracellular Traps Exacerbate Secondary Injury via Promoting Neuroinflammation and Blood–Spinal Cord Barrier Disruption in Spinal Cord Injury. Front. Immunol. 2021, 12, 698249. [Google Scholar] [CrossRef]
- Shafqat, A.; Eddin, A.N.; Adi, G.; Al-Rimawi, M.; Rab, S.A.; Abu-Shaar, M.; Adi, K.; Alkattan, K.; Yaqinuddin, A. Neutrophil extracellular traps in central nervous system pathologies: A mini review. Front. Med. 2023, 10, 1083242. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, H.; Hu, X.; Cai, W.; Ni, W.; Zhou, K. Role of NETosis in Central Nervous System Injury. Oxidative Med. Cell. Longev. 2022, 2022, 3235524. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Yu, H.; Yang, X.; Zhu, Y.; Bai, X.; Wang, R.; Cao, Y.; Xu, H.; Luo, H.; Lu, L.; et al. Neutrophil extracellular traps released by neutrophils impair revascularization and vascular remodeling after stroke. Nat. Commun. 2020, 11, 2488. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Guo, P.; Zhou, J.; Zhang, J.; Zhang, B.; Lan, C.; Xian, J.; Ge, M.; Feng, H.; Chen, Z. Targeting neutrophil extracellular traps enhanced tPA fibrinolysis for experimental intracerebral hemorrhage. Transl. Res. 2019, 211, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Vaibhav, K.; Braun, M.; Alverson, K.; Khodadadi, H.; Kutiyanawalla, A.; Ward, A.; Banerjee, C.; Sparks, T.; Malik, A.; Rashid, M.H.; et al. Neutrophil extracellular traps exacerbate neurological deficits after traumatic brain injury. Sci. Adv. 2020, 6, eaax8847. [Google Scholar] [CrossRef]
- Yamazaki, K.; Kawabori, M.; Seki, T.; Houkin, K. Clinical Trials of Stem Cell Treatment for Spinal Cord Injury. Int. J. Mol. Sci. 2020, 21, 3994. [Google Scholar] [CrossRef] [PubMed]
- Takamiya, S.; Kawabori, M.; Yamazaki, K.; Yamaguchi, S.; Tanimori, A.; Yamamoto, K.; Ohnishi, S.; Seki, T.; Konno, K.; Tha, K.K.; et al. Intravenous transplantation of amnion-derived mesenchymal stem cells promotes functional recovery and alleviates intestinal dysfunction after spinal cord injury. PLoS ONE 2022, 17, e0270606. [Google Scholar] [CrossRef] [PubMed]
- Paul, C.; Samdani, A.F.; Betz, R.R.; Fischer, I.; Neuhuber, B. Grafting of Human Bone Marrow Stromal Cells into Spinal Cord Injury: A Comparison of Delivery Methods. Spine 2009, 34, 328–334. [Google Scholar] [CrossRef]
- Shin, D.A.; Kim, J.-M.; Kim, H.-I.; Yi, S.; Ha, Y.; Yoon, D.H.; Kim, K.N. Comparisonof functional and histological outcomes after intralesional, intracisternal, and intravenous transplantation of human bone marrow-derived mesenchymal stromal cells in a rat model of spinal cord injury. Acta Neurochir. 2013, 155, 1943–1950. [Google Scholar] [CrossRef]
- Hwang, J.; Jang, S.; Kim, C.; Lee, S.; Jeong, H.-S. Role of Stem Cell-Derived Exosomes and microRNAs in Spinal Cord Injury. Int. J. Mol. Sci. 2023, 24, 13849. [Google Scholar] [CrossRef]
- Kosaka, N.; Iguchi, H.; Ochiya, T. Circulating microRNA in body fluid: A new potential biomarker for cancer diagnosis and prognosis. Cancer Sci. 2010, 101, 2087–2092. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Tkach, M.; Théry, C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef]
- Lu, X.; Xu, G.; Lin, Z.; Zou, F.; Liu, S.; Zhang, Y.; Fu, W.; Jiang, J.; Ma, X.; Song, J. Engineered exosomes enriched in netrin-1 modRNA promote axonal growth in spinal cord injury by attenuating inflammation and pyroptosis. Biomater. Res. 2023, 27, 3. [Google Scholar] [CrossRef]
- Shao, Y.; Wang, Q.; Liu, L.; Wang, J.; Wu, M. Alleviation of Spinal Cord Injury by MicroRNA 137-Overexpressing Bone Marrow Mesenchymal Stem Cell-Derived Exosomes. Tohoku J. Exp. Med. 2023, 259, 237–246. [Google Scholar] [CrossRef]
- Gao, X.; Gao, L.F.; Zhang, Y.N.; Kong, X.Q.; Jia, S.; Meng, C.Y. Huc-Mscs-Derived Exosomes Attenuate Neuropathic Pain by Inhibiting Activation of the Tlr2/Myd88/Nf-Kappab Signaling Pathway in the Spinal Microglia by Targeting Rsad2. Int. Immunopharmacol. 2023, 114, 109505. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.; Wang, Y.; Wang, X.; Liu, B.; Rong, L. Mir-146a-5p-Modified Hucmsc-Derived Exosomes Facilitate Spinal Cord Function Recovery by Targeting Neurotoxic Astrocytes. Stem Cell Res. Ther. 2022, 13, 487. [Google Scholar] [CrossRef]
- Shao, Y.; Wang, Q.; Liu, L.; Wang, J.; Wu, M. Exosomes from microRNA 146a overexpressed bone marrow mesenchymal stem cells protect against spinal cord injury in rats. J. Orthop. Sci. 2023, 28, 1149–1156. [Google Scholar] [CrossRef]
- He, X.; Yang, L.; Dong, K.; Zhang, F.; Liu, Y.; Ma, B.; Chen, Y.; Hai, J.; Zhu, R.; Cheng, L. Biocompatible exosome-modified fibrin gel accelerates the recovery of spinal cord injury by VGF-mediated oligodendrogenesis. J. Nanobiotechnol. 2022, 20, 360. [Google Scholar] [CrossRef]
- Zhang, Z.G.; Buller, B.; Chopp, M. Exosomes—Beyond stem cells for restorative therapy in stroke and neurological injury. Nat. Rev. Neurol. 2019, 15, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-Z.; Ma, Z.-J.; Li, J.-R.; Kang, X.-W. Mesenchymal stem cell-derived exosomes: Therapeutic opportunities and challenges for spinal cord injury. Stem Cell Res. Ther. 2021, 12, 102. [Google Scholar] [CrossRef] [PubMed]
- Herbert, F.J.; Bharathi, D.; Suresh, S.; David, E.; Kumar, S. Regenerative Potential of Stem Cell-derived Extracellular Vesicles in Spinal Cord Injury (SCI). Curr. Stem Cell Res. Ther. 2022, 17, 280–293. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-H.; Zeng, L.; Wang, G.; Cao, M.; Lu, H.-B. Anti-Apoptotic Effect of MicroRNA-21 after Contusion Spinal Cord Injury in Rats. J. Neurotrauma 2013, 30, 1349–1360. [Google Scholar] [CrossRef]
- Fleming, J.C.; Norenberg, M.D.; Ramsay, D.A.; Dekaban, G.A.; Marcillo, A.E.; Saenz, A.D.; Pasquale-Styles, M.; Dietrich, W.D.; Weaver, L.C. The cellular inflammatory response in human spinal cords after injury. Brain 2006, 129, 3249–3269. [Google Scholar] [CrossRef] [PubMed]
- Stirling, D.P.; Yong, V.W. Dynamics of the inflammatory response after murine spinal cord injury revealed by flow cytometry. J. Neurosci. Res. 2008, 86, 1944–1958. [Google Scholar] [CrossRef]
- Hellenbrand, D.J.; Quinn, C.M.; Piper, Z.J.; Morehouse, C.N.; Fixel, J.A.; Hanna, A.S. Inflammation after spinal cord injury: A review of the critical timeline of signaling cues and cellular infiltration. J. Neuroinflam. 2021, 18, 284. [Google Scholar] [CrossRef]
- Matsushita, T.; Lankford, K.L.; Arroyo, E.J.; Sasaki, M.; Neyazi, M.; Radtke, C.; Kocsis, J.D. Diffuse and persistent blood–spinal cord barrier disruption after contusive spinal cord injury rapidly recovers following intravenous infusion of bone marrow mesenchymal stem cells. Exp. Neurol. 2015, 267, 152–164. [Google Scholar] [CrossRef]
- Nakazaki, M.; Morita, T.; Lankford, K.L.; Askenase, P.W.; Kocsis, J.D. Small Extracellular Vesicles Released by Infused Mesenchymal Stromal Cells Target M2 Macrophages and Promote Tgf-Beta Upregulation, Microvascular Stabilization and Functional Recovery in a Rodent Model of Severe Spinal Cord Injury. J. Extracell. Vesicles 2021, 10, e12137. [Google Scholar] [CrossRef]
- Jin, Z.; Huang, Q.; Peng, J.; Liu, Z.; Hu, R.; Wu, J.; Wang, F. Mir-125a-3p Alleviates Hyperproliferation of Keratinocytes and Psoriasis-Like Inflammation by Targeting Tlr4/Nf-Kappab Pathway. Postepy Dermatol. Alergol. 2023, 40, 447–461. [Google Scholar] [CrossRef]
- Kim, S.W.; Ramasamy, K.; Bouamar, H.; Lin, A.P.; Jiang, D.; Aguiar, R.C. Micrornas Mir-125a and Mir-125b Constitutively Activate the Nf-Kappab Pathway by Targeting the Tumor Necrosis Factor Alpha-Induced Protein 3 (Tnfaip3, A20). Proc. Natl. Acad. Sci. USA 2012, 109, 7865–7870. [Google Scholar] [CrossRef]
- Wang, Z.; Zhu, D.; Zhang, Y.; Xia, F.; Zhu, J.; Dai, J.; Zhuge, X. Extracellular vesicles produced by avian pathogenic Escherichia coli (APEC) activate macrophage proinflammatory response and neutrophil extracellular trap (NET) formation through TLR4 signaling. Microb. Cell Factories 2023, 22, 177. [Google Scholar] [CrossRef]
- Chang, Q.; Hao, Y.; Wang, Y.; Zhou, Y.; Zhuo, H.; Zhao, G. Bone marrow mesenchymal stem cell-derived exosomal microRNA-125a promotes M2 macrophage polarization in spinal cord injury by downregulating IRF5. Brain Res. Bull. 2021, 170, 199–210. [Google Scholar] [CrossRef]
- Elamir, A.; Shaker, O.; Kamal, M.; Khalefa, A.; Abdelwahed, M.; El Reheem, F.A.; Ahmed, T.; Hassan, E.; Ayoub, S. Expression profile of serum LncRNA THRIL and MiR-125b in inflammatory bowel disease. PLoS ONE 2022, 17, e0275267. [Google Scholar] [CrossRef]
- Loh, J.T.; Zhang, B.; Teo, J.K.H.; Lai, R.C.; Choo, A.B.H.; Lam, K.-P.; Lim, S.K. Mechanism for the attenuation of neutrophil and complement hyperactivity by MSC exosomes. Cytotherapy 2022, 24, 711–719. [Google Scholar] [CrossRef]
- Lankford, K.L.; Arroyo, E.J.; Nazimek, K.; Bryniarski, K.; Askenase, P.W.; Kocsis, J.D. Intravenously delivered mesenchymal stem cell-derived exosomes target M2-type macrophages in the injured spinal cord. PLoS ONE 2018, 13, e0190358. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Perets, N.; Betzer, O.; Ben-Shaul, S.; Sheinin, A.; Michaelevski, I.; Popovtzer, R.; Offen, D.; Levenberg, S. Intranasal Delivery of Mesenchymal Stem Cell Derived Exosomes Loaded with Phosphatase and Tensin Homolog siRNA Repairs Complete Spinal Cord Injury. ACS Nano 2019, 13, 10015–10028. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Chen, Z.; Wu, Y.; Shi, Q.; Yang, E.; Zhang, B.; Qian, Y.; Lian, X.; Xu, J. Adsc-Exos Enhance Functional Recovery after Spinal Cord Injury by Inhibiting Ferroptosis and Promoting the Survival and Function of Endothelial Cells through the Nrf2/Slc7a11/Gpx4 Pathway. Biomed. Pharmacother. 2024, 172, 116225. [Google Scholar] [CrossRef] [PubMed]
- Romanelli, P.; Bieler, L.; Heimel, P.; Škokić, S.; Jakubecova, D.; Kreutzer, C.; Zaunmair, P.; Smolčić, T.; Benedetti, B.; Rohde, E.; et al. Enhancing Functional Recovery Through Intralesional Application of Extracellular Vesicles in a Rat Model of Traumatic Spinal Cord Injury. Front. Cell. Neurosci. 2022, 15, 795008. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Sun, Y.; Liu, Y.; Zhao, J.; Liu, Q.; Xu, J.; Qin, Y.; He, R.; Yuan, F.; Wu, T.; et al. Targeted Delivery of RGD-CD146+CD271+ Human Umbilical Cord Mesenchymal Stem Cell-Derived Exosomes Promotes Blood–Spinal Cord Barrier Repair after Spinal Cord Injury. ACS Nano 2023, 17, 18008–18024. [Google Scholar] [CrossRef]
- Guo, S.; Betzer, O.; Perets, N.; Landau, S.; Offen, D.; Popovtzer, R.; Levenberg, S. Extracellular Vesicles Tracking and Quantification Using CT and Optical Imaging in Rats. Bio-Protoc. 2020, 10, e3635. [Google Scholar] [CrossRef] [PubMed]
- Soleimani, A.; Yazdani, S.O.; Pedram, M.; Saadinam, F.; Rasaee, M.J.; Soleimani, M. Intrathecal injection of human placental mesenchymal stem cells derived exosomes significantly improves functional recovery in spinal cord injured rats. Mol. Biol. Rep. 2024, 51, 193. [Google Scholar] [CrossRef] [PubMed]
- Yuyama, K.; Sun, H.; Mikami, D.; Mioka, T.; Mukai, K.; Igarashi, Y. Lysosomal-associated transmembrane protein 4B regulates ceramide-induced exosome release. FASEB J. 2020, 34, 16022–16033. [Google Scholar] [CrossRef] [PubMed]
- Yuyama, K.; Takahashi, K.; Usuki, S.; Mikami, D.; Sun, H.; Hanamatsu, H.; Furukawa, J.; Mukai, K.; Igarashi, Y. Plant Sphingolipids Promote Extracellular Vesicle Release and Alleviate Amyloid-Beta Pathologies in a Mouse Model of Alzheimer’s Disease. Sci. Rep. 2019, 9, 16827. [Google Scholar] [CrossRef]
- Yamazaki, K.; Kawabori, M.; Seki, T.; Takamiya, S.; Konno, K.; Watanabe, M.; Houkin, K.; Fujimura, M. Mesenchymal Stem Cell Sheet Promotes Functional Recovery and Palliates Neuropathic Pain in a Subacute Spinal Cord Injury Model. Stem Cells Int. 2021, 2021, 9964877. [Google Scholar] [CrossRef]
- Seki, T.; Fehlings, M.G. Mechanistic insights into posttraumatic syringomyelia based on a novel in vivo animal model. J. Neurosurg. Spine 2008, 8, 365–375. [Google Scholar] [CrossRef]
- Basso, D.M.; Beattie, M.S.; Bresnahan, J.C. A Sensitive and Reliable Locomotor Rating Scale for Open Field Testing in Rats. J. Neurotrauma 1995, 12, 1–21. [Google Scholar] [CrossRef]
- Masuda, S.; Nakazawa, D.; Shida, H.; Miyoshi, A.; Kusunoki, Y.; Tomaru, U.; Ishizu, A. Netosis Markers: Quest for Specific, Objective, and Quantitative Markers. Clin. Chim. Acta 2016, 459, 89–93. [Google Scholar] [CrossRef]
- Kawabori, M.; Kacimi, R.; Kauppinen, T.; Calosing, C.; Kim, J.Y.; Hsieh, C.L.; Nakamura, M.C.; Yenari, M.A. Triggering Receptor Expressed on Myeloid Cells 2 (TREM2) Deficiency Attenuates Phagocytic Activities of Microglia and Exacerbates Ischemic Damage in Experimental Stroke. J. Neurosci. 2015, 35, 3384–3396. [Google Scholar] [CrossRef]
- Li, R.H.L.; Ng, G.; Tablin, F. Lipopolysaccharide-induced neutrophil extracellular trap formation in canine neutrophils is dependent on histone H3 citrullination by peptidylarginine deiminase. Vet. Immunol. Immunopathol. 2017, 193–194, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, T.; Seino, R.; Umezaki, K.; Shimoda, A.; Ezoe, T.; Ishiyama, M.; Akiyoshi, K. New Lipophilic Fluorescent Dyes for Labeling Extracellular Vesicles: Characterization and Monitoring of Cellular Uptake. Bioconj. Chem. 2021, 32, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, Y.; Kashiwagi, S.; Yoshida, A.; Satoh, A.O.; Fujioka, M.; Amano, M.; Yamauchi, Y.; Ohba, Y. A method for the generation of pseudovirus particles bearing SARS coronavirus spike protein in high yields. Cell Struct. Funct. 2022, 47, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Senjo, H.; Harada, S.; Kubota, S.I.; Tanaka, Y.; Tateno, T.; Zhang, Z.; Okada, S.; Chen, X.; Kikuchi, R.; Miyashita, N.; et al. Calcineurin inhibitor inhibits tolerance induction by suppressing terminal exhaustion of donor T cells after allo-HCT. Blood 2023, 142, 477–492. [Google Scholar] [CrossRef]
- Shi, S.; Li, T.; Wen, X.; Wu, S.Y.; Xiong, C.; Zhao, J.; Lincha, V.R.; Chow, D.S.; Liu, Y.; Sood, A.K.; et al. Copper-64 Labeled PEGylated Exosomes for In Vivo Positron Emission Tomography and Enhanced Tumor Retention. Bioconj. Chem. 2019, 30, 2675–2683. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morishima, Y.; Kawabori, M.; Yamazaki, K.; Takamiya, S.; Yamaguchi, S.; Nakahara, Y.; Senjo, H.; Hashimoto, D.; Masuda, S.; Fujioka, Y.; et al. Intravenous Administration of Mesenchymal Stem Cell-Derived Exosome Alleviates Spinal Cord Injury by Regulating Neutrophil Extracellular Trap Formation through Exosomal miR-125a-3p. Int. J. Mol. Sci. 2024, 25, 2406. https://doi.org/10.3390/ijms25042406
Morishima Y, Kawabori M, Yamazaki K, Takamiya S, Yamaguchi S, Nakahara Y, Senjo H, Hashimoto D, Masuda S, Fujioka Y, et al. Intravenous Administration of Mesenchymal Stem Cell-Derived Exosome Alleviates Spinal Cord Injury by Regulating Neutrophil Extracellular Trap Formation through Exosomal miR-125a-3p. International Journal of Molecular Sciences. 2024; 25(4):2406. https://doi.org/10.3390/ijms25042406
Chicago/Turabian StyleMorishima, Yutaka, Masahito Kawabori, Kazuyoshi Yamazaki, Soichiro Takamiya, Sho Yamaguchi, Yo Nakahara, Hajime Senjo, Daigo Hashimoto, Sakiko Masuda, Yoichiro Fujioka, and et al. 2024. "Intravenous Administration of Mesenchymal Stem Cell-Derived Exosome Alleviates Spinal Cord Injury by Regulating Neutrophil Extracellular Trap Formation through Exosomal miR-125a-3p" International Journal of Molecular Sciences 25, no. 4: 2406. https://doi.org/10.3390/ijms25042406
APA StyleMorishima, Y., Kawabori, M., Yamazaki, K., Takamiya, S., Yamaguchi, S., Nakahara, Y., Senjo, H., Hashimoto, D., Masuda, S., Fujioka, Y., Ohba, Y., Mizuno, Y., Kuge, Y., & Fujimura, M. (2024). Intravenous Administration of Mesenchymal Stem Cell-Derived Exosome Alleviates Spinal Cord Injury by Regulating Neutrophil Extracellular Trap Formation through Exosomal miR-125a-3p. International Journal of Molecular Sciences, 25(4), 2406. https://doi.org/10.3390/ijms25042406