
Nicotinic Receptors in Human Chromaffin Cells: Characterization, Functional and Physical Interactions between Subtypes and Regulation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
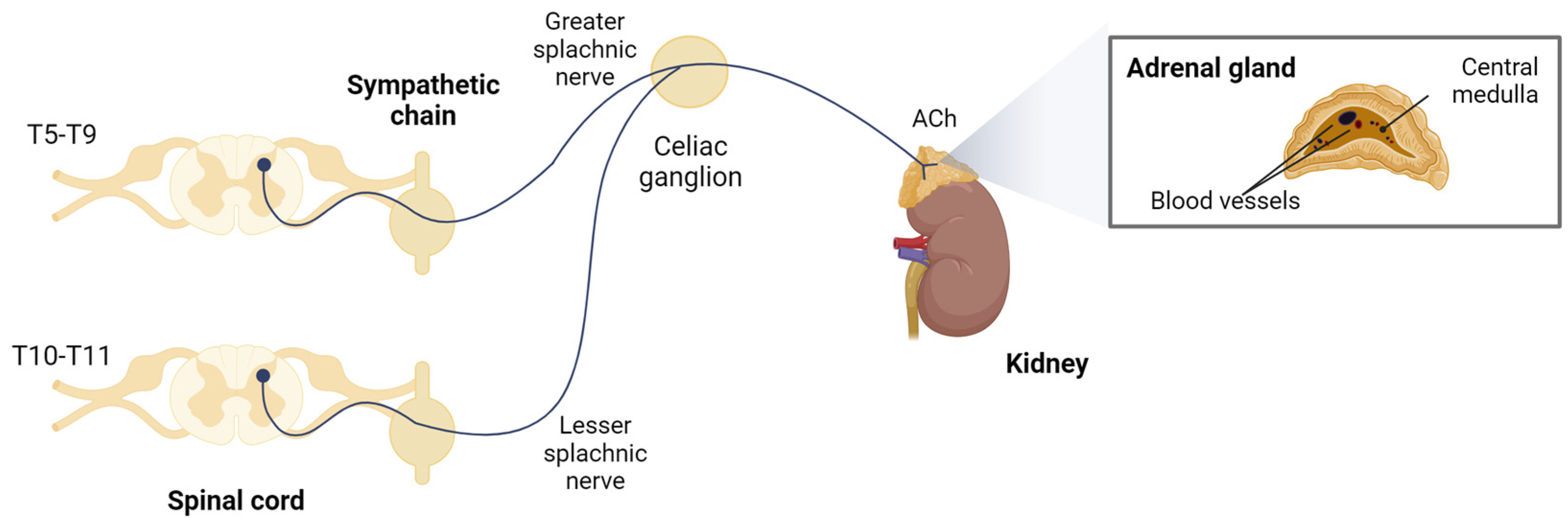
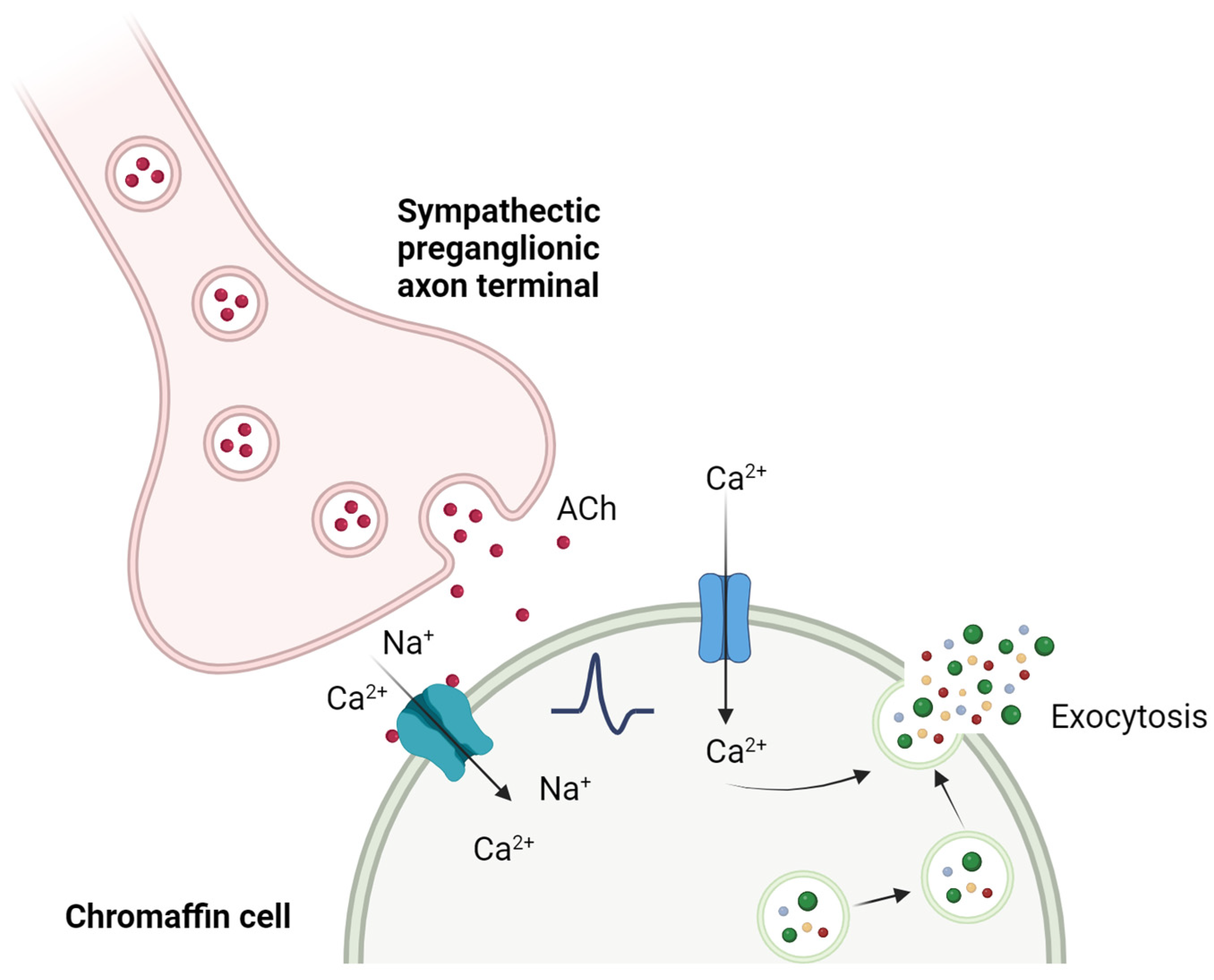
2. nAChRs in Chromaffin Cells of the Adrenal Gland
2.1. Non-Human Species
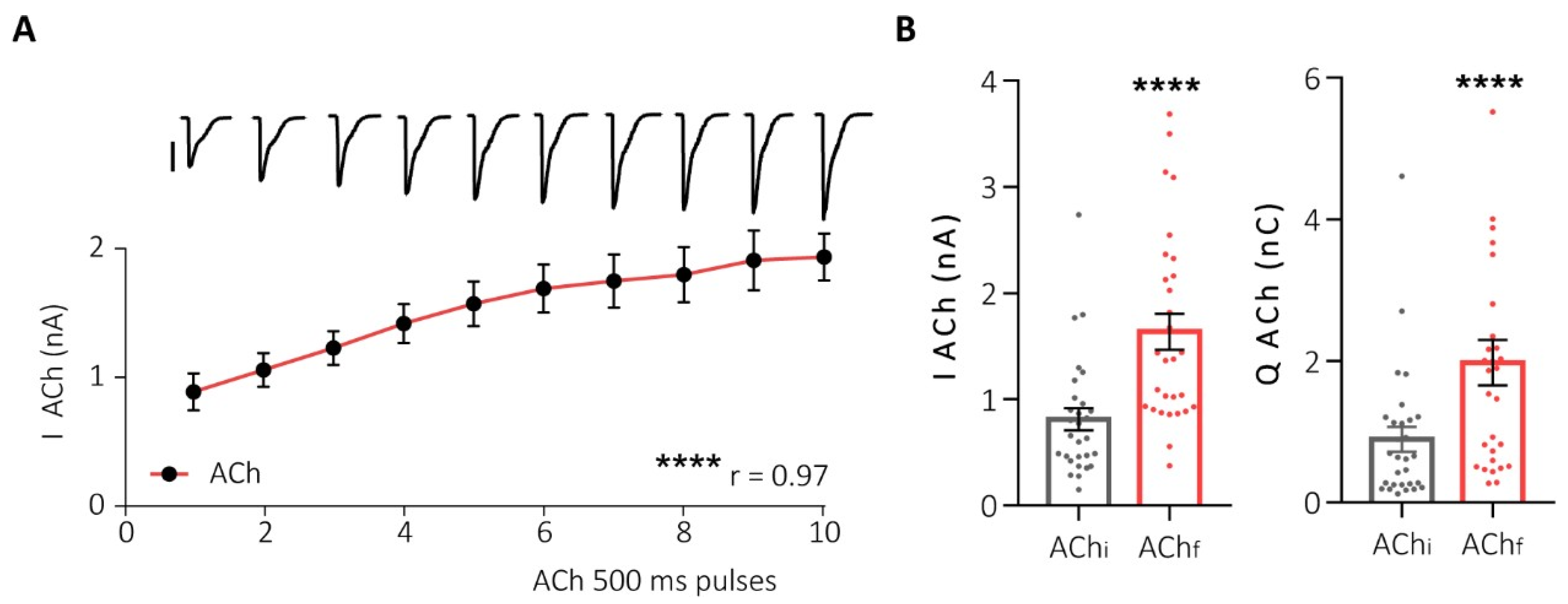
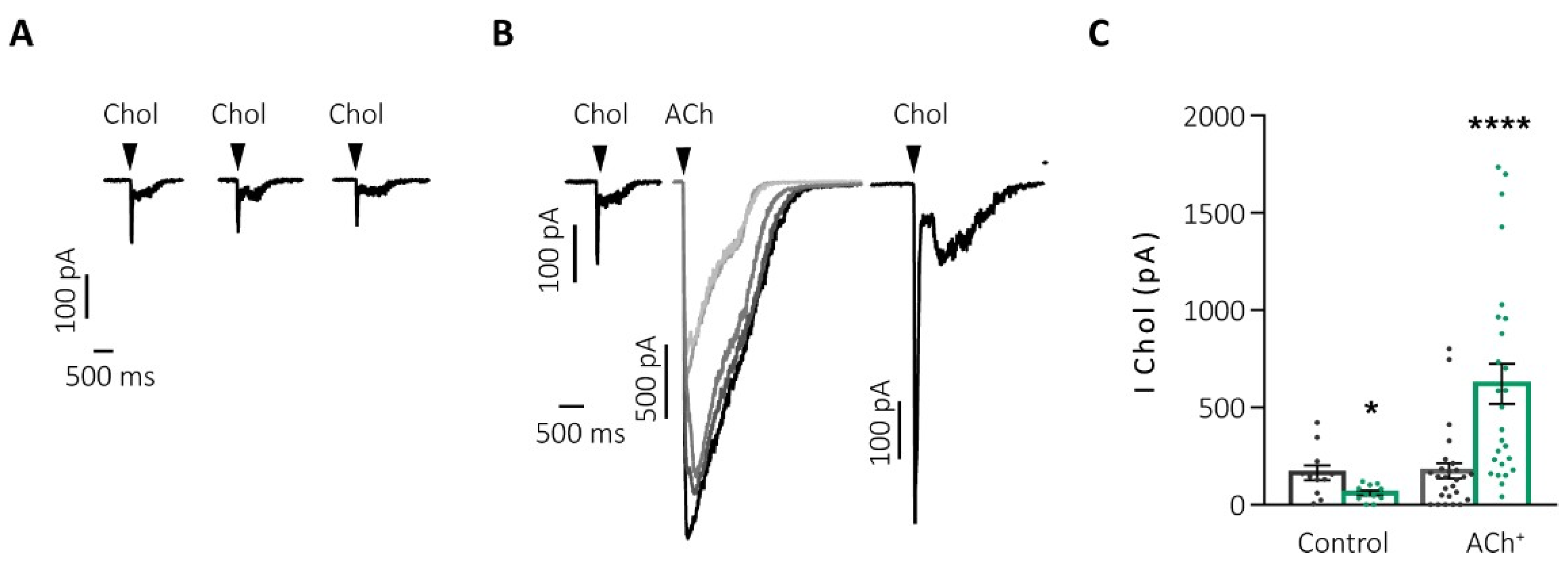
2.2. Human Species
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Gotti, C.; Zoli, M.; Clementi, F. Brain nicotinic acetylcholine receptors: Native subtypes and their relevance. Trends Pharmacol. Sci. 2006, 27, 482–491. [Google Scholar] [CrossRef]
- Gotti, C.; Clementi, F. Neuronal nicotinic receptors: From structure to pathology. Prog. Neurobiol. 2004, 74, 363–396. [Google Scholar] [CrossRef]
- Briggs, C.A.; Gopalakrishnan, M. 2.22—Ion Channels–Ligand Gated. In Comprehensive Medicinal Chemistry II; Taylor, J.B., Triggle, D.J., Eds.; Elsevier: Oxford, UK, 2007; pp. 877–918. [Google Scholar]
- Albuquerque, E.X.; Pereira, E.F.; Alkondon, M.; Rogers, S.W. Mammalian nicotinic acetylcholine receptors: From structure to function. Physiol. Rev. 2009, 89, 73–120. [Google Scholar] [CrossRef]
- Corradi, J.; Bouzat, C. Understanding the Bases of Function and Modulation of alpha7 Nicotinic Receptors: Implications for Drug Discovery. Mol. Pharmacol. 2016, 90, 288–299. [Google Scholar] [CrossRef]
- Dani, J.A. Overview of nicotinic receptors and their roles in the central nervous system. Biol. Psychiatry. 2001, 49, 166–174. [Google Scholar] [CrossRef]
- King, J.R.; Nordman, J.C.; Bridges, S.P.; Lin, M.; Kabbani, N. Identification and characterization of a G protein-binding cluster in α7 nicotinic acetylcholine receptors. J. Biol. Chem. 2015, 290, 20060–20070. [Google Scholar] [CrossRef]
- Moretti, M.; Zoli, M.; George, A.A.; Lukas, R.J.; Pistillo, F.; Maskos, U.; Whiteaker, P.; Gotti, C. The novel α7β2-nicotinic acetylcholine receptor subtype is expressed in mouse and human basal forebrain: Biochemical and pharmacological characterization. Mol. Pharmacol. 2014, 86, 306–317. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Zwart, R.; Ursu, D.; Jensen, M.M.; Pinborg, L.H.; Gilmour, G.; Wu, J.; Sher, E.; Mikkelsen, J.D. α7 and β2 nicotinic acetylcholine receptor subunits form heteromeric receptor complexes that are expressed in the human cortex and display distinct pharmacological properties. PLoS ONE 2015, 10, e0130572. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, Q.; Tang, P.; Mikkelsen, J.D.; Shen, J.; Whiteaker, P.; Yakel, J.L. Heteromeric α7β2 nicotinic acetylcholine receptors in the brain. Trends Pharmacol. Sci. 2016, 37, 562–574. [Google Scholar] [CrossRef] [PubMed]
- Araud, T.; Graw, S.; Berger, R.; Lee, M.; Neveu, E.; Bertrand, D.; Leonard, S. The chimeric gene CHRFAM7A, a partial duplication of the CHRNA7 gene, is a dominant negative regulator of α7* nAChR function. Biochem. Pharmacol. 2011, 82, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Martín-Sánchez, C.; Alés, E.; Balseiro-Gómez, S.; Atienza, G.; Arnalich, F.; Bordas, A.; Cedillo, J.L.; Extremera, M.; Chávez-Reyes, A.; Montiel, C. The human-specific duplicated α7 gene inhibits the ancestral α7, negatively regulating nicotinic acetylcholine receptor-mediated transmitter release. J. Biol. Chem. 2021, 296, 100341. [Google Scholar] [CrossRef]
- Whiteaker, P.; Peterson, C.G.; Xu, W.; McIntosh, J.M.; Paylor, R.; Beaudet, A.L.; Collins, A.C.; Marks, M.J. Involvement of the α3 subunit in central nicotinic binding populations. J. Neurosci. 2002, 22, 2522–2529. [Google Scholar] [CrossRef]
- Hernández, S.C.; Vicini, S.; Xiao, Y.; Dávila-García, M.I.; Yasuda, R.P.; Wolfe, B.B.; Kellar, K.J. The nicotinic receptor in the rat pineal gland is an α3β4 subtype. Mol. Pharmacol. 2004, 66, 978–987. [Google Scholar] [CrossRef]
- Ramírez-Latorre, J.; Yu, C.R.; Qu, X.; Perin, F.; Karlin, A.; Role, L. Functional contributions of α5 subunit to neuronal acetylcholine receptor channels. Nature 1996, 380, 347–351. [Google Scholar] [CrossRef]
- Gotti, C.; Moretti, M.; Clementi, F.; Riganti, L.; McIntosh, J.M.; Collins, A.C.; Marks, M.J.; Whiteaker, P. Expression of nigrostriatal α6-containing nicotinic acetylcholine receptors is selectively reduced, but not eliminated, by β3 subunit gene deletion. Mol. Pharmacol. 2005, 67, 2007–2015. [Google Scholar] [CrossRef] [PubMed]
- Quik, M.; McIntosh, J.M. Striatal α6* nicotinic acetylcholine receptors: Potential targets for Parkinson’s disease therapy. J. Pharmacol. Exp. Ther. 2006, 316, 481–489. [Google Scholar] [CrossRef]
- McIntosh, J.M.; Absalom, N.; Chebib, M.; Elgoyhen, A.B.; Vincler, M. Alpha9 nicotinic acetylcholine receptors and the treatment of pain. Biochem. Pharmacol. 2009, 78, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Colomer, C.; Olivos-Oré, L.A.; Vincent, A.; McIntosh, J.M.; Artalejo, A.R.; Guérineau, N.C. Functional characterization of alpha9-containing cholinergic nicotinic receptors in the rat adrenal medulla: Implication in stress-induced functional plasticity. J. Neurosci. 2010, 30, 6732–6742. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.L. Physiology of the Adrenal Gland. Br. Med. J. 1937, 1, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T. Neural regulation of adrenal chromaffin cell function in the mouse—Stress effect on the distribution of [3H] dopamine in denervated adrenal medulla. Brain Res. 1982, 238, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Kajiwara, R.; Sand, O.; Kidokoro, Y.; Barish, M.E.; Iijima, T. Functional organization of chromaffin cells and cholinergic synaptic transmission in rat adrenal medulla. Jpn. J. Physiol. 1997, 47, 449–464. [Google Scholar] [CrossRef]
- Douglas, W.W.; Rubin, R.P. The role of calcium in the secretory response of the adrenal medulla to acetylcholine. J. Physiol. 1961, 159, 40–57. [Google Scholar] [CrossRef]
- Sandow, A. Excitation-contraction coupling in muscular response. Yale J. Biol. Med. 1952, 25, 176. [Google Scholar]
- Norman, A.W.; Henry, H.L. Chapter 11—Hormones of the Adrenal Medulla. In Hormones, 3rd ed.; Norman, A.W., Henry, H.L., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 239–253. [Google Scholar]
- Feldberg, W.; Minz, B.; Tsudzimura, H. The mechanism of the nervous discharge of adrenaline. J. Physiol. 1934, 81, 286–304. [Google Scholar] [CrossRef] [PubMed]
- Fenwick, E.M.; Marty, A.; Neher, E. A patch-clamp study of bovine chromaffin cells and of their sensitivity to acetylcholine. J. Physiol. 1982, 331, 577–597. [Google Scholar] [CrossRef] [PubMed]
- Role, L.W.; Perlman, R.L. Both nicotinic and muscarinic receptors mediate catecholamine secretion by isolated guinea-pig chromaffin cells. Neuroscience 1983, 10, 979–985. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Kuriyama, H. Muscarine induces two distinct current responses in adrenal chromaffin cells of the guinea-pig. Jpn. J. Physiol. 1990, 40, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Douglas, W.W.; Poisner, A.M. Preferential release of adrenaline from the adrenal medulla by muscarine and pilocarpine. Nature 1965, 208, 1102–1103. [Google Scholar] [CrossRef] [PubMed]
- Biales, B.; Dichter, M.; Tischler, A. Electrical excitability of cultured adrenal chromaffin cells. J. Physiol. 1976, 262, 743–753. [Google Scholar] [CrossRef] [PubMed]
- Brandt, B.L.; Hagiwara, S.; Kidokoro, Y.; Miyazaki, S. Action potentials in the rat chromaffin cell and effects of acetylcholine. J. Physiol. 1976, 263, 417–439. [Google Scholar] [CrossRef] [PubMed]
- Kidokoro, Y.; Ritchie, A.K. Chromaffin cell action potentials and their possible role in adrenaline secretion from rat adrenal medulla. J. Physiol. 1980, 307, 199–216. [Google Scholar] [CrossRef] [PubMed]
- Fenwick, E.M.; Marty, A.; Neher, E. Sodium and calcium channels in bovine chromaffin cells. J. Physiol. 1982, 331, 599–635. [Google Scholar] [CrossRef] [PubMed]
- Marty, A.; Neher, E. Potassium channels in cultured bovine adrenal chromaffin cells. J. Physiol. 1985, 367, 117–141. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Shimizu, T.; Nakamura, K.; Yokotani, K. Brain neuronal/inducible nitric oxide synthases and cyclooxygenase-1 are involved in the bombesin-induced activation of central adrenomedullary outflow in rats. Eur. J. Pharmacol. 2008, 590, 177–184. [Google Scholar] [CrossRef]
- Krieglstein, K.; Deimling, F.; Suter-Crazzolara, C.; Unsicker, K. Expression and localization of GDNF in developing and adult adrenal chromaffin cells. Cell Tissue Res. 1996, 286, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Unsicker, K.; Krieglstein, K. Growth factors in chromaffin cells. Prog. Neurobiol. 1996, 48, 307–324. [Google Scholar] [CrossRef]
- Bormann, J.; Clapham, D.E. gamma-Aminobutyric acid receptor channels in adrenal chromaffin cells: A patch-clamp study. Proc. Natl. Acad. Sci. USA 1985, 82, 2168–2172. [Google Scholar] [CrossRef]
- Marley, P.; Livett, B.G. Neuropeptides in the autonomic nervous system. CRC Crit. Rev. Clin. Neurobiol. 1985, 1, 201–283. [Google Scholar]
- Bornstein, S.R.; Ehrhart-Bornstein, M.; Androutsellis-Theotokis, A.; Eisenhofer, G.; Vukicevic, V.; Licinio, J.; Wong, M.; Calissano, P.; Nistico, G.; Preziosi, P.; et al. Chromaffin cells: The peripheral brain. Mol. Psychiatry 2012, 17, 354–358. [Google Scholar] [CrossRef]
- Tischler, A.S. Chromaffin cells as models of endocrine cells and neurons. Ann. N. Y. Acad. 2002, 971, 366–370. [Google Scholar] [CrossRef]
- Campos-Caro, A.; Smillie, F.I.; Dominguez del Toro, E.; Rovira, J.C.; Vicente-Agullo, F.; Chapuli, J.; Juiz, J.M.; Sala, S.; Sala, F.; Ballesta, J.J.; et al. Neuronal nicotinic acetylcholine receptors on bovine chromaffin cells: Cloning, expression, and genomic organization of receptor subunits. J. Neurochem. 1997, 68, 488–497. [Google Scholar] [CrossRef]
- García-Guzmán, M.; Sala, F.; Sala, S.; Campos-Caro, A.; Stuhmer, W.; Gutierrez, L.M.; Criado, M. alpha-Bungarotoxin-sensitive nicotinic receptors on bovine chromaffin cells: Molecular cloning, functional expression and alternative splicing of the alpha 7 subunit. Eur. J. Neurosci. 1995, 7, 647–655. [Google Scholar] [CrossRef]
- Quik, M.; Geertsen, S.; Trifaró, J.M. Marked up-regulation of the beta-bungarotoxin site in adrenal chromaffin cells by specific nicotinic antagonists. Mol. Pharmacol. 1987, 31, 385–391. [Google Scholar]
- Wilson, S.P.; Kirshner, N. The acetylcholine receptor of the adrenal medulla. J. Neurochem. 1977, 28, 687–695. [Google Scholar] [CrossRef]
- El-Hajj, R.A.; McKay, S.B.; McKay, D.B. Pharmacological and immunological identification of native alpha7 nicotinic receptors: Evidence for homomeric and heteromeric alpha7 receptors. Life Sci. 2007, 81, 1317–1322. [Google Scholar] [CrossRef] [PubMed]
- Blumenthal, E.M.; Conroy, W.G.; Romano, S.J.; Kassner, P.D.; Berg, D.K. Detection of functional nicotinic receptors blocked by alpha-bungarotoxin on PC12 cells and dependence of their expression on post-translational events. J. Neurosci. 1997, 17, 6094–6104. [Google Scholar] [CrossRef] [PubMed]
- López, M.G.; Montiel, C.; Herrero, C.J.; García-Palomero, E.; Mayorgas, I.; Hernández-Guijo, J.M.; Villarroya, M.; Olivares, R.; Gandía, L.; McIntosh, J.M.; et al. Unmasking the functions of the chromaffin cell α7 nicotinic receptor by using short pulses of acetylcholine and selective blockers. Proc. Natl. Acad. Sci. USA 1998, 95, 14184–14189. [Google Scholar] [CrossRef] [PubMed]
- Del Barrio, L.; Egea, J.; Leon, R.; Romero, A.; Ruiz, A.; Montero, M.; Alvarez, J.; López, M.G. Calcium signalling mediated through alpha7 and non-alpha7 nAChR stimulation is differentially regulated in bovine chromaffin cells to induce catecholamine release. Br. J. Pharmacol. 2011, 162, 94–110. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, D.L.; Slepetis, R.; Kirshner, N. Inhibition of catecholamine secretion from adrenal medulla cells by neurotoxins and cholinergic antagonists. J. Neurochem. 1981, 37, 125–131. [Google Scholar] [CrossRef]
- Kumakura, K.; Karoum, F.; Guidotti, A.; Costa, E. Modulation of nicotinic receptors by opiate receptor agonists in cultured adrenal chromaffin cells. Nature 1980, 283, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Tachikawa, E.; Mizuma, K.; Kudo, K.; Kashimoto, T.; Yamato, S.; Ohta, S. Characterization of the functional subunit combination of nicotinic acetylcholine receptors in bovine adrenal chromaffin cells. Neurosci. Lett. 2001, 312, 161–164. [Google Scholar] [CrossRef]
- Trifaró, J.M.; Lee, R.W. Morphological characteristics and stimulus-secretion coupling in bovine adcrenal chromaffin cell cultures. Neuroscience 1980, 5, 1533–1546. [Google Scholar] [CrossRef] [PubMed]
- Di Angelantonio, S.; Matteoni, C.; Fabbretti, E.; Nistri, A. Molecular biology and electrophysiology of neuronal nicotinic receptors of rat chromaffin cells. Eur. J. Neurosci. 2003, 17, 2313–2322. [Google Scholar] [CrossRef] [PubMed]
- Hone, A.J.; Rueda-Ruzafa, L.; Gordon, T.J.; Gajewiak, J.; Christensen, S.; Dyhring, T.; Albillos, A.; McIntosh, J.M. Expression of α3β2β4 nicotinic acetylcholine receptors by rat adrenal chromaffin cells determined using novel conopeptide antagonists. J. Neurochem. 2020, 154, 158–176. [Google Scholar] [CrossRef]
- Luo, S.; Zhangsun, D.; Zhu, X.; Wu, Y.; Hu, Y.; Christensen, S.; Harvey, P.J.; Akcan, M.; Craik, D.J.; McIntosh, J.M. Characterization of a novel α-conotoxin TxID from Conus textile that potently blocks rat α3β4 nicotinic acetylcholine receptors. J. Med. Chem. 2013, 56, 9655–9663. [Google Scholar] [CrossRef] [PubMed]
- Hone, A.J.; Michael McIntosh, J.; Rueda-Ruzafa, L.; Passas, J.; de Castro-Guerín, C.; Blázquez, J.; González-Enguita, C.; Albillos, A. Therapeutic concentrations of varenicline in the presence of nicotine increase action potential firing in human adrenal chromaffin cells. J. Neurochem. 2017, 140, 37–52. [Google Scholar] [CrossRef]
- Rust, G.; Burgunder, J.M.; Lauterburg, T.E.; Cachelin, A.B. Expression of neuronal nicotinic acetylcholine receptor subunit genes in the rat autonomic nervous system. Eur. J. Neurosci. 1994, 6, 478–485. [Google Scholar] [CrossRef]
- Mousavi, M.; Hellström-Lindahl, E.; Guan, Z.; Bednar, I.; Nordberg, A. Expression of nicotinic acetylcholine receptors in human and rat adrenal medulla. Life Sci. 2001, 70, 577–590. [Google Scholar] [CrossRef]
- Pérez-Alvarez, A.; Hernández-Vivanco, A.; Alonso y Gregorio, S.; Tabernero, A.; McIntosh, J.M.; Albillos, A. Pharmacological characterization of native α7 nicotinic ACh receptors and their contribution to depolarization-elicited exocytosis in human chromaffin cells. Br. J. Pharmacol. 2012, 165, 908–921. [Google Scholar] [CrossRef]
- Jiménez-Pompa, A.; Sanz-Lázaro, S.; Omodolor, R.E.; Medina-Polo, J.; González-Enguita, C.; Blázquez, J.; McIntosh, J.M.; Albillos, A. Cross talk between α7 and α3β4 nicotinic receptors prevents their desensitization in human chromaffin cells. J. Neurosci. 2022, 42, 1173–1183. [Google Scholar] [CrossRef]
- Azam, L.; Maskos, U.; Changeux, J.; Dowell, C.D.; Christensen, S.; De Biasi, M.; McIntosh, J.M. α-Conotoxin BuIA [T5A; P6O]: A novel ligand that discriminates between α6β4 and α6β2 nicotinic acetylcholine receptors and blocks nicotine-stimulated norepinephrine release. FASEB J. 2010, 24, 5113. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Alvarez, A.; Hernández-Vivanco, A.; McIntosh, J.M.; Albillos, A. Native α6β4∗ nicotinic receptors control exocytosis in human chromaffin cells of the adrenal gland. FASEB J. 2012, 26, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Hone, A.J.; McIntosh, J.M.; Azam, L.; Lindstrom, J.; Lucero, L.; Whiteaker, P.; Passas, J.; Blazquez, J.; Albillos, A. alpha-Conotoxins Identify the alpha3beta4* Subtype as the Predominant Nicotinic Acetylcholine Receptor Expressed in Human Adrenal Chromaffin Cells. Mol. Pharmacol. 2015, 88, 881–893. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Alvarez, A.; Albillos, A. Key role of the nicotinic receptor in neurotransmitter exocytosis in human chromaffin cells. J. Neurochem. 2007, 103, 2281–2290. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Coggan, J.S.; Berg, D.K. Synaptic currents generated by neuronal acetylcholine receptors sensitive to α-bungarotoxin. Neuron 1996, 17, 1231–1240. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ullian, E.M.; McIntosh, J.M.; Sargent, P.B. Rapid synaptic transmission in the avian ciliary ganglion is mediated by two distinct classes of nicotinic receptors. J. Neurosci. 1997, 17, 7210–7219. [Google Scholar] [CrossRef]
- Alkondon, M.; Pereira, E.F.; Albuquerque, E.X. α-Bungarotoxin-and methyllycaconitine-sensitive nicotinic receptors mediate fast synaptic transmission in interneurons of rat hippocampal slices. Brain Res. 1998, 810, 257–263. [Google Scholar] [CrossRef]
- Frazier, C.J.; Buhler, A.V.; Weiner, J.L.; Dunwiddie, T.V. Synaptic potentials mediated via α-bungarotoxin-sensitive nicotinic acetylcholine receptors in rat hippocampal interneurons. J. Neurosci. 1998, 18, 8228–8235. [Google Scholar] [CrossRef]
- Liu, Z.; Tearle, A.W.; Nai, Q.; Berg, D.K. Rapid activity-driven SNARE-dependent trafficking of nicotinic receptors on somatic spines. J. Neurosci. 2005, 25, 1159–1168. [Google Scholar] [CrossRef]
- Harkness, P.C.; Millar, N.S. Changes in conformation and subcellular distribution of α4β2 nicotinic acetylcholine receptors revealed by chronic nicotine treatment and expression of subunit chimeras. J. Neurosci. 2002, 22, 10172–10181. [Google Scholar] [CrossRef]
- Buisson, B.; Bertrand, D. Chronic exposure to nicotine upregulates the human α4β2 nicotinic acetylcholine receptor function. J. Neurosci. 2001, 21, 1819–1829. [Google Scholar] [CrossRef]
- St. John, P.A. Cellular trafficking of nicotinic acetylcholine receptors. Acta Pharmacol. Sin. 2009, 30, 656–662. [Google Scholar] [CrossRef]
- Palma, E.; Maggi, L.; Barabino, B.; Eusebi, F.; Ballivet, M. Nicotinic acetylcholine receptors assembled from the α7 and β3 subunits. J. Biol. Chem. 1999, 274, 18335–18340. [Google Scholar] [CrossRef]
- Khiroug, S.S.; Harkness, P.C.; Lamb, P.W.; Sudweeks, S.N.; Khiroug, L.; Millar, N.S.; Yakel, J.L. Rat nicotinic ACh receptor α7 and β2 subunits co-assemble to form functional heteromeric nicotinic receptor channels. J. Physiol. 2002, 540, 425–434. [Google Scholar] [CrossRef]
- Criado, M.; Valor, L.M.; Mulet, J.; Gerber, S.; Sala, S.; Sala, F. Expression and functional properties of alpha7 acetylcholine nicotinic receptors are modified in the presence of other receptor subunits. J. Neurochem. 2012, 123, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Maneu, V.; Rojo, J.; Mulet, J.; Valor, L.M.; Sala, F.; Criado, M.; García, A.G.; Gandía, L. A single neuronal nicotinic receptor α3α7β4* is present in the bovine chromaffin cell. Ann. N. Y. Acad. Sci. 2002, 971, 165–167. [Google Scholar] [CrossRef] [PubMed]
- González-Rubio, J.M.; Rojo, J.; Tapia, L.; Maneu, V.; Mulet, J.; Valor, L.M.; Criado, M.; Sala, F.; García, A.G.; Gandía, L. Activation and blockade by choline of bovine α7 and α3β4 nicotinic receptors expressed in oocytes. Eur. J. Pharmacol. 2006, 535, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Mazzaferro, S.; Benallegue, N.; Carbone, A.; Gasparri, F.; Vijayan, R.; Biggin, P.C.; Moroni, M.; Bermudez, I. Additional acetylcholine (ACh) binding site at α4/α4 interface of (α4β2) 2α4 nicotinic receptor influences agonist sensitivity. J. Biol. Chem. 2011, 286, 31043–31054. [Google Scholar] [CrossRef] [PubMed]
- Colombo, S.F.; Mazzo, F.; Pistillo, F.; Gotti, C. Biogenesis, trafficking and up-regulation of nicotinic ACh receptors. Biochem. Pharmacol. 2013, 86, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Millar, N.S.; Harkness, P.C. Assembly and trafficking of nicotinic acetylcholine receptors. Mol. Membr. Biol. 2008, 25, 279–292. [Google Scholar] [CrossRef]
- Crespi, A.; Colombo, S.F.; Gotti, C. Proteins and chemical chaperones involved in neuronal nicotinic receptor expression and function: An update. Br. J. Pharmacol. 2018, 175, 1869–1879. [Google Scholar] [CrossRef]
- Wang, F.; Gerzanich, V.; Wells, G.B.; Anand, R.; Peng, X.; Keyser, K.; Lindstrom, J. Assembly of human neuronal nicotinic receptor α5 subunits with α3, β2, and β4 subunits. J. Biol. Chem. 1996, 271, 17656–17665. [Google Scholar] [CrossRef]
- Gerzanich, V.; Wang, F.; Kuryatov, A.; Lindstrom, J. α5 subunit alters desensitization, pharmacology, Ca permeability and Ca modulation of human neuronal α3 nicotinic receptors. J. Pharmacol. Exp. Ther. 1998, 286, 311–320. [Google Scholar]
- Boorman, J.P.; GrootKormelink, P.J.; Sivilotti, L.G. Stoichiometry of human recombinant neuronal nicotinic receptors containing the β3 subunit expressed in Xenopus oocytes. J. Physiol. 2000, 529, 565–577. [Google Scholar] [CrossRef]
- Nelson, M.E.; Kuryatov, A.; Choi, C.H.; Zhou, Y.; Lindstrom, J. Alternate stoichiometries of α4β2 nicotinic acetylcholine receptors. Mol. Pharmacol. 2003, 63, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Moroni, M.; Zwart, R.; Sher, E.; Cassels, B.K.; Bermudez, I. α4β2 nicotinic receptors with high and low acetylcholine sensitivity: Pharmacology, stoichiometry, and sensitivity to long-term exposure to nicotine. Mol. Pharmacol. 2006, 70, 755–768. [Google Scholar] [CrossRef]
- Zoli, M.; Pistillo, F.; Gotti, C. Diversity of native nicotinic receptor subtypes in mammalian brain. Neuropharmacology 2015, 96, 302–311. [Google Scholar] [CrossRef]
- Giribaldi, J.; Dutertre, S. α-Conotoxins to explore the molecular, physiological and pathophysiological functions of neuronal nicotinic acetylcholine receptors. Neurosci. Lett. 2018, 679, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Conroy, W.G.; Liu, Z.; Nai, Q.; Coggan, J.S.; Berg, D.K. PDZ-containing proteins provide a functional postsynaptic scaffold for nicotinic receptors in neurons. Neuron 2003, 38, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Parker, M.J.; Zhao, S.; Bredt, D.S.; Sanes, J.R.; Feng, G. PSD93 regulates synaptic stability at neuronal cholinergic synapses. J. Neurosci. 2004, 24, 378–388. [Google Scholar] [CrossRef]
- Conroy, W.G.; Ogden, L.F.; Berg, D.K. Cluster formation of α7-containing nicotinic receptors at interneuronal interfaces in cell culture. Neuropharmacology 2000, 39, 2699–2705. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ford, B.; Mann, M.A.; Fischbach, G.D. Neuregulins increase α7 nicotinic acetylcholine receptors and enhance excitatory synaptic transmission in GABAergic interneurons of the hippocampus. J. Neurosci. 2001, 21, 5660–5669. [Google Scholar] [CrossRef] [PubMed]
- Kawai, H.; Zago, W.; Berg, D.K. Nicotinic α7 receptor clusters on hippocampal GABAergic neurons: Regulation by synaptic activity and neurotrophins. J. Neurosci. 2002, 22, 7903–7912. [Google Scholar] [CrossRef] [PubMed]
- Baer, K.; Bürli, T.; Huh, K.; Wiesner, A.; Erb-Vögtli, S.; Göckeritz-Dujmovic, D.; Moransard, M.; Nishimune, A.; Rees, M.I.; Henley, J.M.; et al. PICK1 interacts with α7 neuronal nicotinic acetylcholine receptors and controls their clustering. Mol. Cell. Neurosci. 2007, 35, 339–355. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wiesner, A.; Fuhrer, C. Regulation of nicotinic acetylcholine receptors by tyrosine kinases in the peripheral and central nervous system: Same players, different roles. Cell. Mol. Life Sci. CMLS 2006, 63, 2818–2828. [Google Scholar] [CrossRef]
- Cho, C.; Song, W.; Leitzell, K.; Teo, E.; Meleth, A.D.; Quick, M.W.; Lester, R.A. Rapid upregulation of α7 nicotinic acetylcholine receptors by tyrosine dephosphorylation. J. Neurosci. 2005, 25, 3712–3723. [Google Scholar] [CrossRef]
- Mamlouk, G.M.; Dorris, D.M.; Barrett, L.R.; Meitzen, J. Sex bias and omission in neuroscience research is influenced by research model and journal, but not reported NIH funding. Front. Neuroendocrinol. 2020, 57, 100835. [Google Scholar] [CrossRef] [PubMed]
- Léna, C.; Changeux, J. Allosteric modulations of the nicotinic acetylcholine receptor. Trends Neurosci. 1993, 16, 181–186. [Google Scholar] [CrossRef]
- Zheng, P. Neuroactive steroid regulation of neurotransmitter release in the central nervous system: Action, mechanism and possible significance. Prog. Neurobiol. 2009, 89, 134–152. [Google Scholar] [CrossRef]
- Ke, L.; Lukas, R.J. Effects of steroid exposure on ligand binding and functional activities of diverse nicotinic acetylcholine receptor subtypes. J. Neurochem. 1996, 67, 1100–1112. [Google Scholar] [CrossRef]
- Stitzel, J.A.; Farnham, D.A.; Collins, A.C. Chronic corticosterone treatment elicits dose-dependent changes in mouse brain α-bungarotoxin binding. Neuroscience 1996, 72, 791–799. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-Pompa, A.; Albillos, A. Nicotinic Receptors in Human Chromaffin Cells: Characterization, Functional and Physical Interactions between Subtypes and Regulation. Int. J. Mol. Sci. 2024, 25, 2304. https://doi.org/10.3390/ijms25042304
Jiménez-Pompa A, Albillos A. Nicotinic Receptors in Human Chromaffin Cells: Characterization, Functional and Physical Interactions between Subtypes and Regulation. International Journal of Molecular Sciences. 2024; 25(4):2304. https://doi.org/10.3390/ijms25042304
Chicago/Turabian StyleJiménez-Pompa, Amanda, and Almudena Albillos. 2024. "Nicotinic Receptors in Human Chromaffin Cells: Characterization, Functional and Physical Interactions between Subtypes and Regulation" International Journal of Molecular Sciences 25, no. 4: 2304. https://doi.org/10.3390/ijms25042304
APA StyleJiménez-Pompa, A., & Albillos, A. (2024). Nicotinic Receptors in Human Chromaffin Cells: Characterization, Functional and Physical Interactions between Subtypes and Regulation. International Journal of Molecular Sciences, 25(4), 2304. https://doi.org/10.3390/ijms25042304