
Antibiotic Combination to Effectively Postpone or Inhibit the In Vitro Induction and Selection of Levofloxacin-Resistant Mutants in Elizabethkingia anophelis

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Clones and MICs in E. anophelis
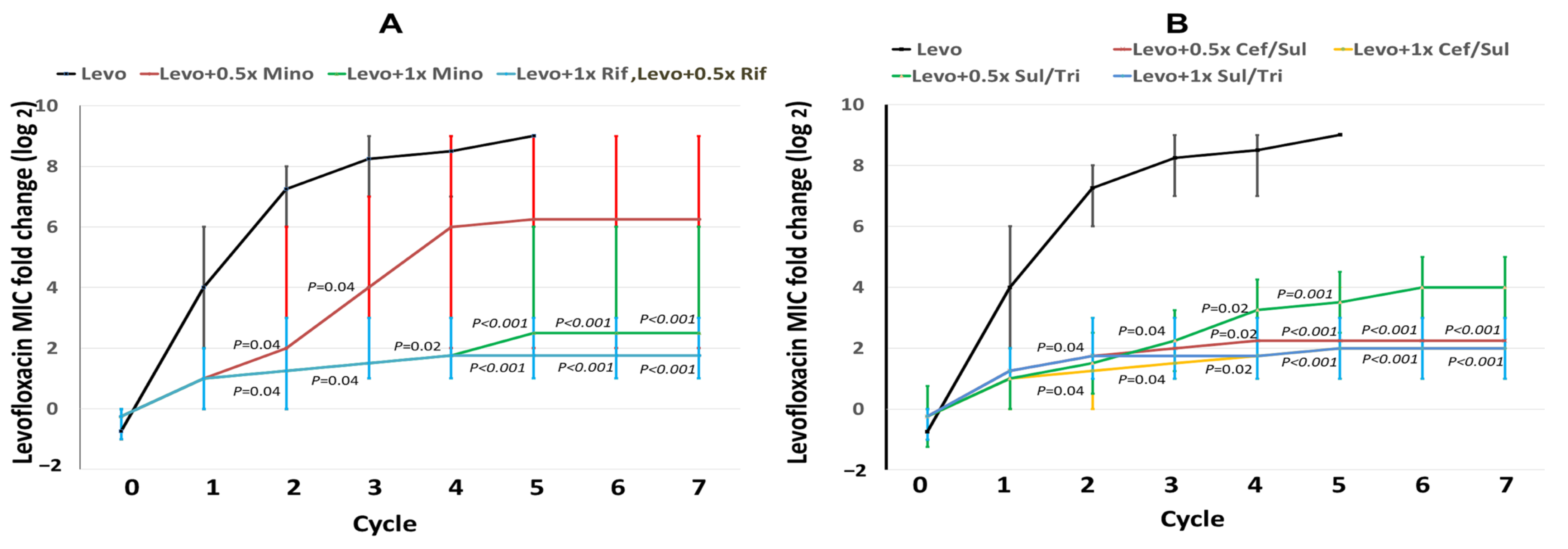
2.2. Changes in Levofloxacin MICs during Induction Cycles
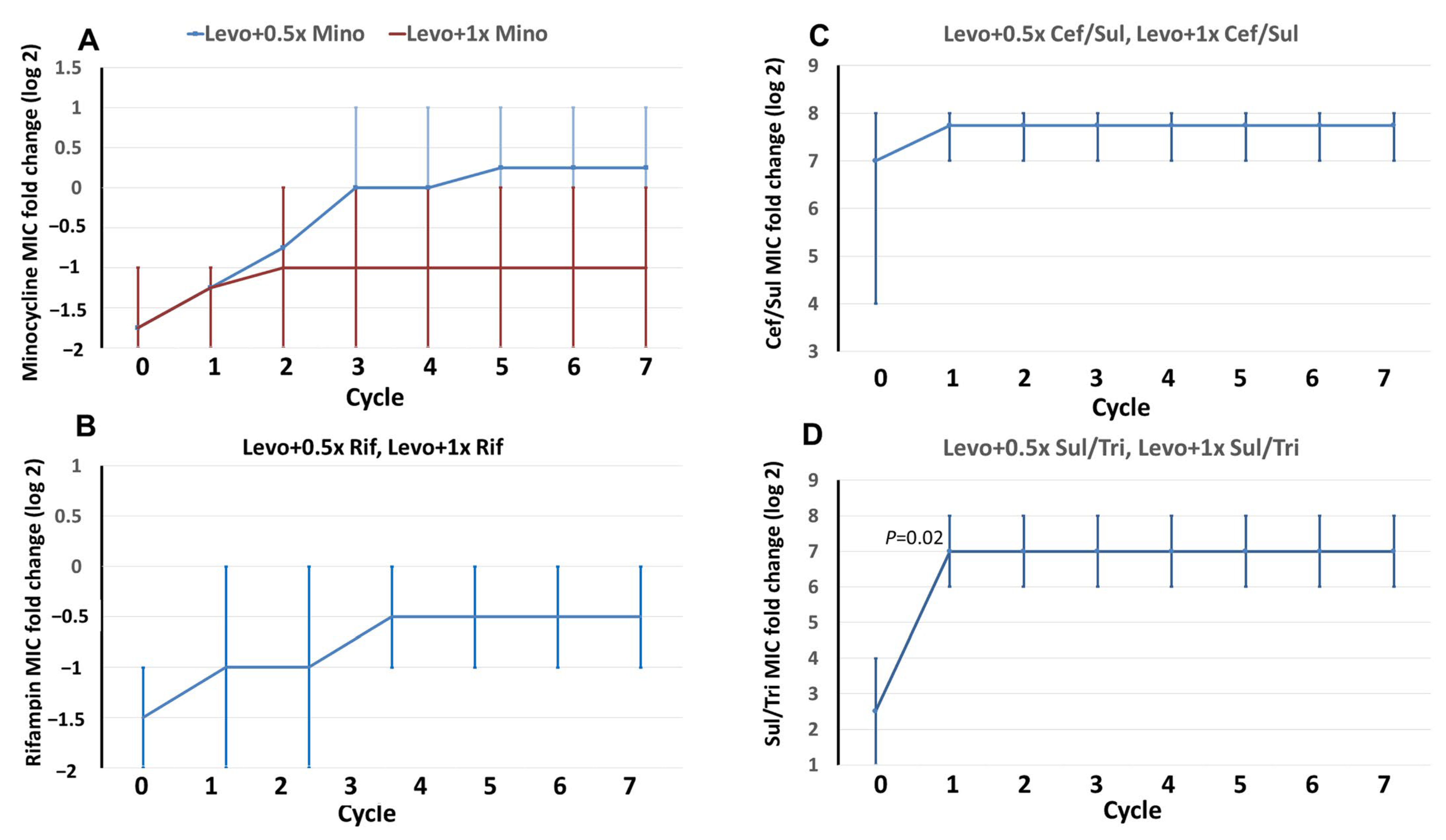
2.3. MIC Changes in the Combined Antibiotics in Induction Cycles
2.4. Mutations in QRDRs during Induction Cycles
3. Discussion
4. Methods
4.1. Study Setting and Bacterial Isolates
4.2. PFGE
4.3. Determination of the Minimum Inhibitory Concentrations
4.4. Multicycle Induction and Selection of Resistant Mutants
4.5. Amplification and Sequencing to Identify Mutations in QRDRs
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Janda, J.M.; Lopez, D.L. Mini review: New pathogen profiles: Elizabethkingia anophelis. Diag. Microbiol. Infect. Dis. 2017, 88, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.; Chow, W.N.; Foo, C.H.; Curreem, S.O.; Lo, G.C.S.; Teng, J.L.; Chen, J.H.; Ng, R.H.; Wu, A.K.; Cheung, I.Y. Elizabethkingia anophelis bacteremia is associated with clinically significant infections and high mortality. Sci. Rep. 2016, 6, 26045. [Google Scholar] [CrossRef] [PubMed]
- Chew, K.L.; Cheng, B.; Lin, R.T.; Teo, J.W. Elizabethkingia anophelis is the dominant Elizabethkingia species found in blood cultures in Singapore. J. Clin. Microbiol. 2018, 56, e01445-17. [Google Scholar] [CrossRef] [PubMed]
- Evans, L.; Rhodes, A.; Alhazzani, W.; Antonelli, M.; Coopersmith, C.M.; French, C.; Machado, F.R.; Mcintyre, L.; Ostermann, M.; Prescott, H.C. Surviving sepsis campaign: International guidelines for management of sepsis and septic shock 2021. Intens. Care Med. 2021, 47, 1181–1247. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.H.; Perng, C.L.; Jian, M.J.; Lee, S.Y.; Sun, J.R.; Shang, H.S. Multicentre study evaluating matrix-assisted laser desorption ionization–time of flight mass spectrometry for identification of clinically isolated Elizabethkingia species and analysis of antimicrobial susceptibility. Clin. Microbiol. Infect. 2019, 25, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Han, M.S.; Kim, H.; Lee, Y.; Kim, M.; Ku, N.S.; Choi, J.Y.; Yong, D.; Jeong, S.H.; Lee, K.; Chong, Y. Relative prevalence and antimicrobial susceptibility of clinical isolates of Elizabethkingia species based on 16S rRNA gene sequencing. J. Clin. Microbiol. 2017, 55, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Perrin, A.; Larsonneur, E.; Nicholson, A.C.; Edwards, D.J.; Gundlach, K.M.; Whitney, A.M.; Gulvik, C.A.; Bell, M.E.; Rendueles, O.; Cury, J. Evolutionary dynamics and genomic features of the Elizabethkingia anophelis 2015 to 2016 Wisconsin outbreak strain. Nat. Commun. 2017, 8, 15483. [Google Scholar] [CrossRef] [PubMed]
- Aldred, K.J.; Kerns, R.J.; Osheroff, N. Mechanism of quinolone action and resistance. Biochemistry 2014, 53, 1565–1574. [Google Scholar] [CrossRef] [PubMed]
- Jian, M.J.; Cheng, Y.H.; Chung, H.Y.; Cheng, Y.H.; Yang, H.Y.; Hsu, C.S.; Perng, C.L.; Shang, H.S. Fluoroquinolone resistance in carbapenem-resistant Elizabethkingia anophelis: Phenotypic and genotypic characteristics of clinical isolates with topoisomerase mutations and comparative genomic analysis. J. Antimicrob. Chemother. 2019, 74, 1503–1510. [Google Scholar] [CrossRef] [PubMed]
- Lin, I.F.; Lai, C.H.; Lin, S.Y.; Lee, C.C.; Lee, N.Y.; Liu, P.Y.; Yang, C.H.; Huang, Y.H.; Lin, J.N. Mutant Prevention Concentrations of Ciprofloxacin and Levofloxacin and Target Gene Mutations of Fluoroquinolones in Elizabethkingia anophelis. Antimicrob. Agents Chemother. 2022, 66, e00301-22. [Google Scholar] [CrossRef] [PubMed]
- Mehta, K.C.; Dargad, R.R.; Borade, D.M.; Swami, O.C. Burden of antibiotic resistance in common infectious diseases: Role of antibiotic combination therapy. J Clin. Diag. Res. 2014, 8, ME05. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.N.; Lai, C.H.; Huang, Y.H.; Yang, C.H. Antimicrobial effects of minocycline, tigecycline, ciprofloxacin, and levofloxacin against Elizabethkingia anophelis using in vitro time-kill assays and in vivo zebrafish animal models. Antibiotics 2021, 10, 285. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.J.; Lin, Y.T.; Chen, C.C.; Chen, C.W.; Lu, Y.C.; Ko, W.C.; Chen, H.J.; Su, B.A.; Chang, P.C.; Chuang, Y.C. Molecular characteristics and in vitro effects of antimicrobial combinations on planktonic and biofilm forms of Elizabethkingia anophelis. J. Antimicrob. Chemother. 2021, 76, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Burnard, D.; Gore, L.; Henderson, A.; Ranasinghe, A.; Bergh, H.; Cottrell, K.; Sarovich, D.S.; Price, E.P.; Paterson, D.L.; Harris, P.N. Comparative genomics and antimicrobial resistance profiling of Elizabethkingia isolates reveal nosocomial transmission and in vitro susceptibility to fluoroquinolones, tetracyclines, and trimethoprim-sulfamethoxazole. J. Clin. Microbiol. 2020, 58, e00730-20. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.; Chong, C.Y.; Thoon, K.C.; Tee, N.W.S.; Maiwald, M.; Lam, J.C.M.; Bhattacharya, R.; Chandran, S.; Yung, C.F.; Tan, N.W.H. Invasive paediatric Elizabethkingia meningoseptica infections are best treated with a combination of piperacillin/tazobactam and trimethoprim/sulfamethoxazole or fluoroquinolone. J. Med. Microbiol. 2019, 68, 1167. [Google Scholar] [CrossRef] [PubMed]
- Seong, H.; Kim, J.H.; Kim, J.H.; Lee, W.J.; Ahn, J.Y.; Ku, N.S.; Choi, J.Y.; Yeom, J.S.; Song, Y.G.; Jeong, S.J. Risk factors for mortality in patients with elizabethkingia infection and the clinical impact of the antimicrobial susceptibility patterns of elizabethkingia species. J. Clin. Microbiol. 2020, 9, 1431. [Google Scholar] [CrossRef] [PubMed]
- Gootz, T.D.; Zaniewski, R.; Haskell, S.; Schmieder, B.; Tankovic, J.; Girard, D.; Courvalin, P.; Polzer, R.J. Activity of the new fluoroquinolone trovafloxacin (CP-99,219) against DNA gyrase and topoisomerase IV mutants of Streptococcus pneumoniae selected in vitro. Antimicrob. Agents Chemother. 1996, 40, 2691–2697. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, L.; Cameron, B.; Crouzet, J. Analysis of gyrA and grlA mutations in stepwise-selected ciprofloxacin-resistant mutants of Staphylococcus aureus. Antimicrob. Agents Chemother. 1995, 39, 1554–1558. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.L.; Liu, K.M.; Chang, H.L.; Liao, Y.C.; Lin, J.S.; Kung, F.Y.; Ho, C.M.; Lin, K.H.; Chen, Y.T. The evolutionary trend and genomic features of an emerging lineage of elizabethkingia anophelis strains in Taiwan. Microbiol. Spectr. 2022, 10, e01682-21. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; Approved standard. 32th informational supplement. CLSI document M100-S32; CLSI: Wayne, PA, USA, 2022. [Google Scholar]

{kind=link}
{kind=link}
| Levofloxacin + 1 × MIC Minocycline | 1 × Levofloxacin + 0.5 × MIC Minocycline | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Strain | Cycle | Levofloxacin MIC (mg/L) | Point Mutation in Gyr A (gyrA) | Point Mutation in GyrB (gyrB) | Strain | Cycle | Levofloxacin MIC (mg/L) | Point Mutation in Gyr A (gyrA) | Point Mutation in GyrB (gyrB) |
| No. 1 | 0 | 0.5 | 83 Ser (AGC) | N/D | No. 1 | 0 | 0.5 | 83 Ser (AGC) | 470 Glu (GAG) |
| 1 | 4 | N/D | N/D | 1 | 4 | N/D | N/D | ||
| 2 | 8 | N/D | N/D | 2 | 64 | 83 Arg (AGG) | 470 Glu (GAG) | ||
| 3 | 8 | N/D | N/D | 3 | 128 | 83 Arg (AGG) | 470 Glu (GAG) | ||
| 4 | 8 | N/D | N/D | 4 | >256 | 83 Arg (AGG) | 470 Lys (AAG) | ||
| 5 | 64 | 83 Ile (ATC) | N/D | - | - | - | - | ||
| 6 | 64 | 83 Ile (ATC) | N/D | - | - | - | - | ||
| 7 | 64 | 83 Ile (ATC) | N/D | - | - | - | - | ||
| No. 2 | 0 | 1 | N/D | N/D | No. 2 | 0 | 1 | N/D | N/D |
| 1 | 2 | N/D | N/D | 1 | 2 | N/D | N/D | ||
| 2 | 2 | N/D | N/D | 2 | 2 | N/D | N/D | ||
| 3 | 2 | N/D | N/D | 3 | 2 | N/D | N/D | ||
| 4 | 4 | N/D | N/D | 4 | 4 | N/D | N/D | ||
| 5 | 4 | N/D | N/D | 5 | 4 | N/D | N/D | ||
| 6 | 4 | N/D | N/D | 6 | 4 | N/D | N/D | ||
| 7 | 4 | N/D | N/D | 7 | 4 | N/D | N/D | ||
| No. 3 | 0 | 1 | N/D | N/D | No. 3 | 0 | 1 | 81 Gly (GGT) | N/D |
| 1 | 2 | N/D | N/D | 1 | 2 | N/D | N/D | ||
| 2 | 2 | N/D | N/D | 2 | 2 | N/D | N/D | ||
| 3 | 2 | N/D | N/D | 3 | 16 | N/D | N/D | ||
| 4 | 2 | N/D | N/D | 4 | >256 | 81 Asp (GAT) | N/D | ||
| 5 | 2 | N/D | N/D | - | - | - | - | ||
| 6 | 2 | N/D | N/D | - | - | - | - | ||
| 7 | 2 | N/D | N/D | - | - | - | - | ||
| No. 4 | 0 | 1 | N/D | N/D | No. 4 | 0 | 1 | N/D | N/D |
| 1 | 1 | N/D | N/D | 1 | 1 | N/D | N/D | ||
| 2 | 1 | N/D | N/D | 2 | 1 | N/D | N/D | ||
| 3 | 2 | N/D | N/D | 3 | 16 | N/D | N/D | ||
| 4 | 2 | N/D | N/D | 4 | 16 | N/D | N/D | ||
| 5 | 2 | N/D | N/D | 5 | 32 | N/D | N/D | ||
| 6 | 2 | N/D | N/D | 6 | 32 | N/D | N/D | ||
| 7 | 2 | N/D | N/D | 7 | 32 | N/D | N/D | ||
| Levofloxacin + 1 × MIC Sulfamethoxazole/Trimethoprim | Levofloxacin + 0.5 × MIC Sulfamethoxazole/Trimethoprim | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Strain | Cycle | Levofloxacin MIC (mg/L) | Point Mutation in Gyr A (gyrA) | Point Mutation in GyrB (gyrB) | Strain | Cycle | Levofloxacin MIC (mg/L) | Point Mutation in Gyr A (gyrA) | Point Mutation in GyrB (gyrB) |
| No. 1 | 0 | 0.5 | N/D | N/D | No. 1 | 0 | 0.5 | N/D | N/D |
| 1 | 2 | N/D | N/D | 1 | 2 | N/D | N/D | ||
| 2 | 8 | N/D | N/D | 2 | 8 | N/D | N/D | ||
| 3 | 8 | N/D | N/D | 3 | 8 | N/D | N/D | ||
| 4 | 8 | N/D | N/D | 4 | 8 | N/D | N/D | ||
| 5 | 8 | N/D | N/D | 5 | 8 | N/D | N/D | ||
| 6 | 8 | N/D | N/D | 6 | 8 | N/D | N/D | ||
| 7 | 8 | N/D | N/D | 7 | 8 | N/D | N/D | ||
| No. 2 | 0 | 1 | N/D | N/D | No. 2 | 0 | 1 | N/D | N/D |
| 1 | 4 | N/D | N/D | 1 | 4 | N/D | N/D | ||
| 2 | 4 | N/D | N/D | 2 | 4 | N/D | N/D | ||
| 3 | 4 | N/D | N/D | 3 | 4 | N/D | N/D | ||
| 4 | 4 | N/D | N/D | 4 | 4 | N/D | N/D | ||
| 5 | 8 | N/D | N/D | 5 | 8 | N/D | N/D | ||
| 6 | 8 | N/D | N/D | 6 | 8 | N/D | N/D | ||
| 7 | 8 | N/D | N/D | 7 | 8 | N/D | N/D | ||
| No. 3 | 0 | 1 | N/D | N/D | No. 3 | 0 | 1 | 87 Asp (GAT) | 471 Glu (GAA) |
| 1 | 2 | N/D | N/D | 1 | 2 | N/D | N/D | ||
| 2 | 2 | N/D | N/D | 2 | 2 | N/D | N/D | ||
| 3 | 2 | N/D | N/D | 3 | 8 | N/D | N/D | ||
| 4 | 2 | N/D | N/D | 4 | 128 | 87 Asp (GAT) | 471 Lys (AAA) | ||
| 5 | 2 | N/D | N/D | 5 | 128 | 87 Asp (GAT) | 471 Lys (AAA) | ||
| 6 | 2 | N/D | N/D | 6 | >256 | 87 Asn (AAT) | 471 Lys (AAA) | ||
| 7 | 2 | N/D | N/D | - | - | - | - | ||
| No. 4 | 0 | 1 | N/D | N/D | No. 4 | 0 | 1 | N/D | N/D |
| 1 | 1 | N/D | N/D | 1 | 1 | N/D | N/D | ||
| 2 | 1 | N/D | N/D | 2 | 1 | N/D | N/D | ||
| 3 | 2 | N/D | N/D | 3 | 2 | N/D | N/D | ||
| 4 | 2 | N/D | N/D | 4 | 2 | N/D | N/D | ||
| 5 | 2 | N/D | N/D | 5 | 2 | N/D | N/D | ||
| 6 | 2 | N/D | N/D | 6 | 2 | N/D | N/D | ||
| 7 | 2 | N/D | N/D | 7 | 2 | N/D | N/D | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.-C.; Lai, C.-H.; Yang, C.-H.; Huang, Y.-H.; Lin, J.-N. Antibiotic Combination to Effectively Postpone or Inhibit the In Vitro Induction and Selection of Levofloxacin-Resistant Mutants in Elizabethkingia anophelis. Int. J. Mol. Sci. 2024, 25, 2215. https://doi.org/10.3390/ijms25042215
Lee C-C, Lai C-H, Yang C-H, Huang Y-H, Lin J-N. Antibiotic Combination to Effectively Postpone or Inhibit the In Vitro Induction and Selection of Levofloxacin-Resistant Mutants in Elizabethkingia anophelis. International Journal of Molecular Sciences. 2024; 25(4):2215. https://doi.org/10.3390/ijms25042215
Chicago/Turabian StyleLee, Ching-Chi, Chung-Hsu Lai, Chih-Hui Yang, Yi-Han Huang, and Jiun-Nong Lin. 2024. "Antibiotic Combination to Effectively Postpone or Inhibit the In Vitro Induction and Selection of Levofloxacin-Resistant Mutants in Elizabethkingia anophelis" International Journal of Molecular Sciences 25, no. 4: 2215. https://doi.org/10.3390/ijms25042215
APA StyleLee, C.-C., Lai, C.-H., Yang, C.-H., Huang, Y.-H., & Lin, J.-N. (2024). Antibiotic Combination to Effectively Postpone or Inhibit the In Vitro Induction and Selection of Levofloxacin-Resistant Mutants in Elizabethkingia anophelis. International Journal of Molecular Sciences, 25(4), 2215. https://doi.org/10.3390/ijms25042215