Isolation and Identification of a Tibetan Pig Porcine Epidemic Diarrhoea Virus Strain and Its Biological Effects on IPEC-J2 Cells

Abstract
1. Introduction
2. Results
2.1. Microscopic Observation of the Effect of Viruses on Cell Morphology
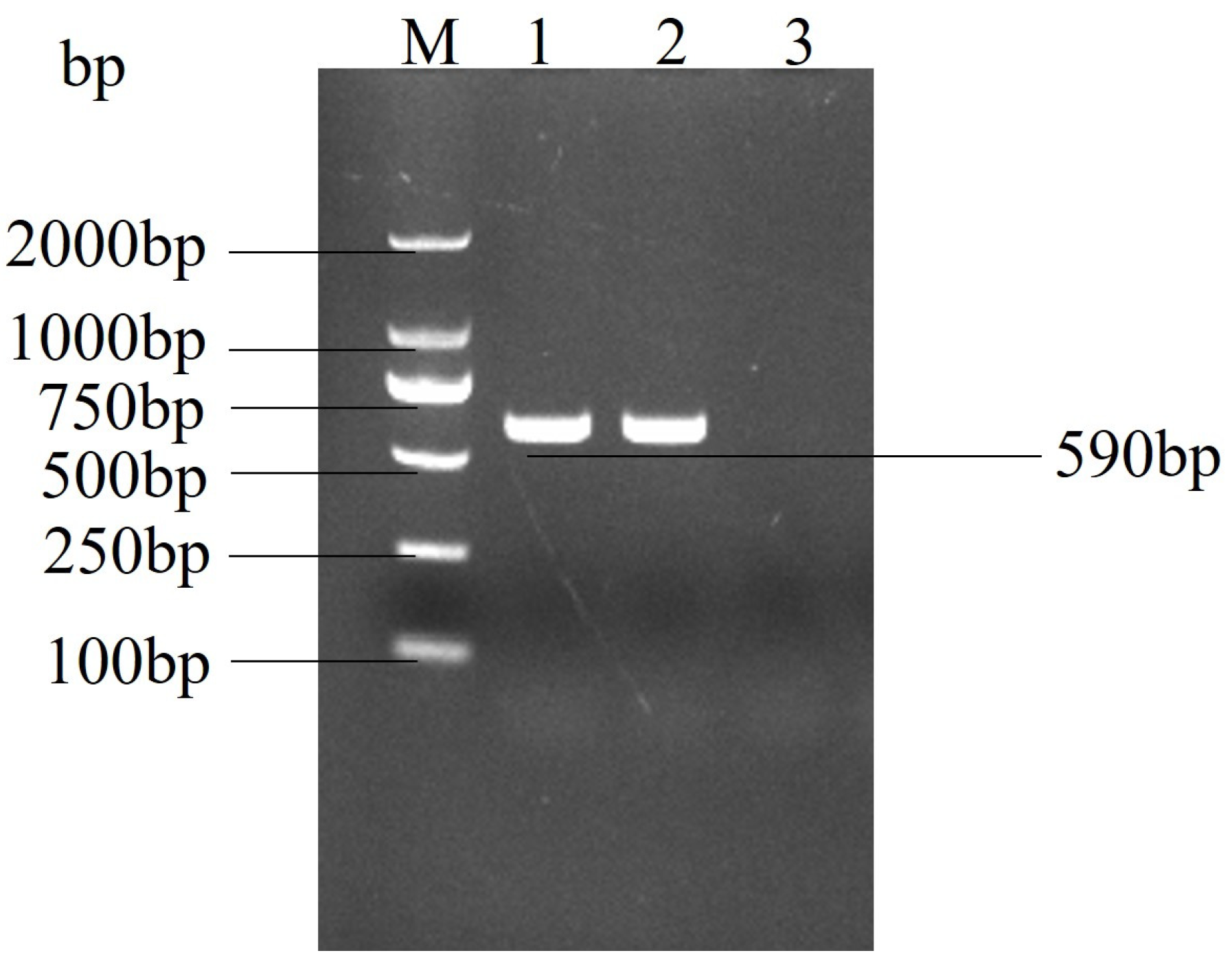
2.2. Real Time PCR Confirmed the Identity of the Suspected Virus as PEDV
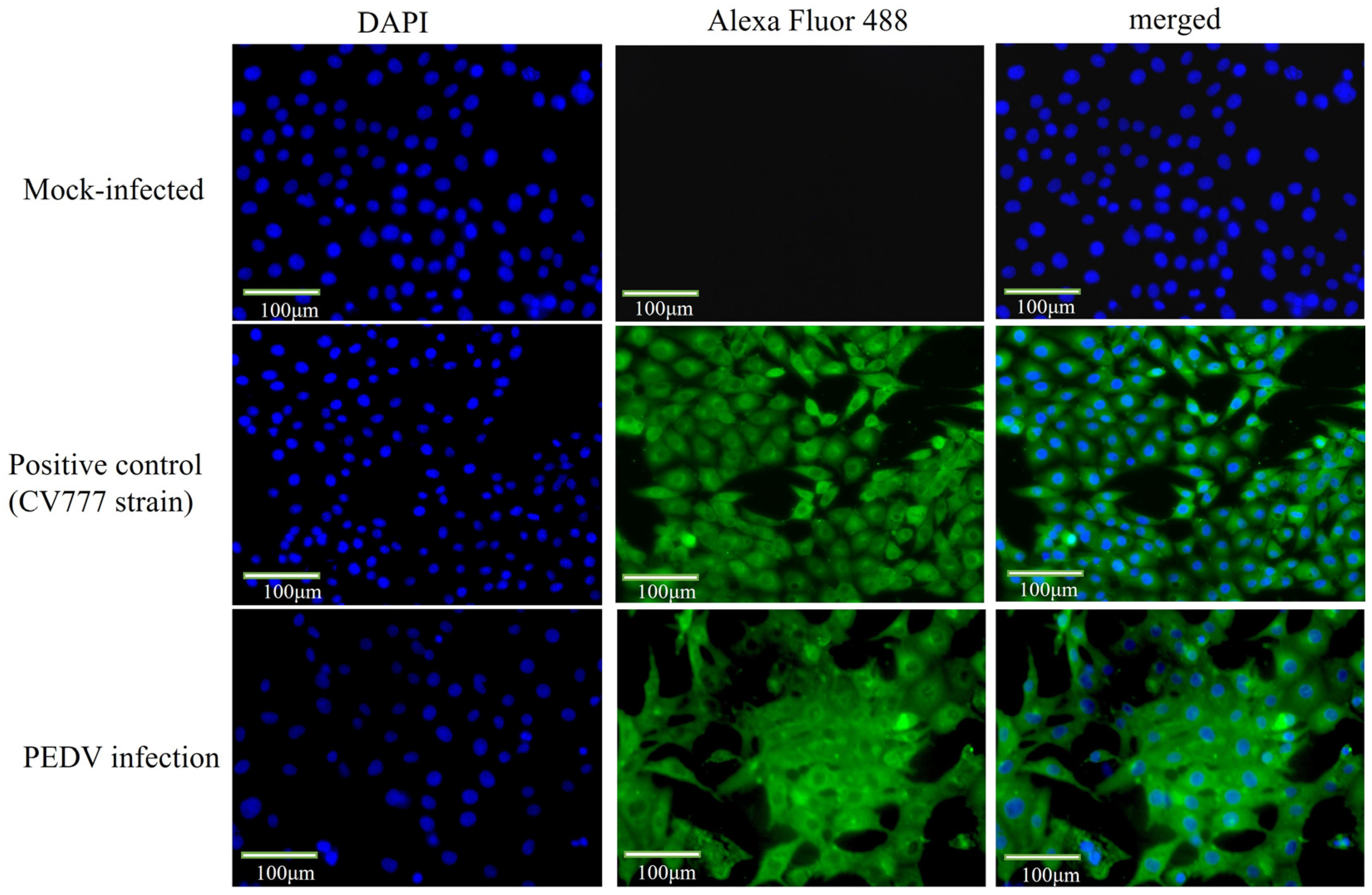
2.3. Indirect Immunofluorescence Identified the Virus as PEDV
2.4. Observation of Viral Particles in Ultra-Thin Sections of Cells
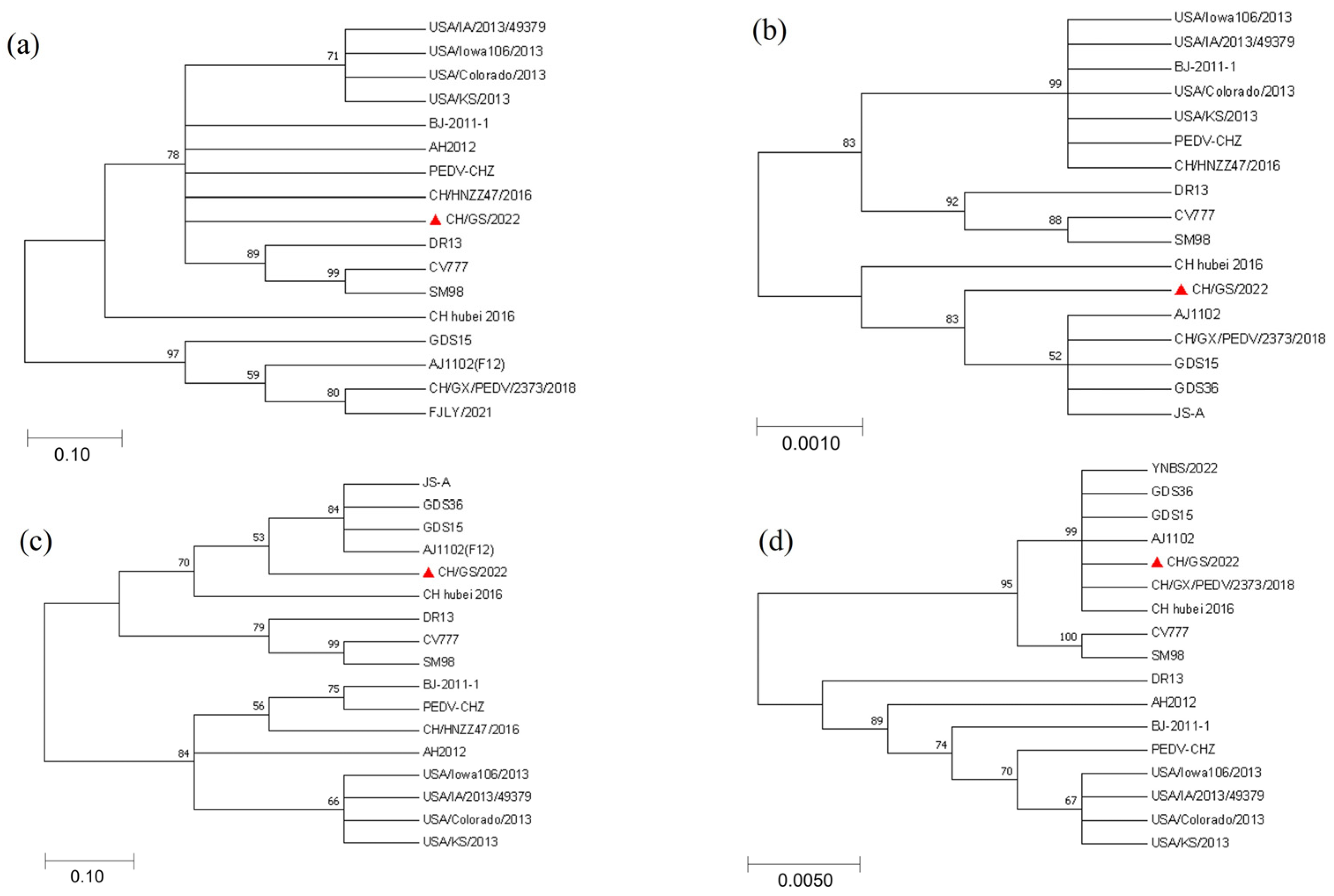
2.5. Results of Phylogenetic Evolutionary Analysis of CH/GS/2022 Isolate Genes
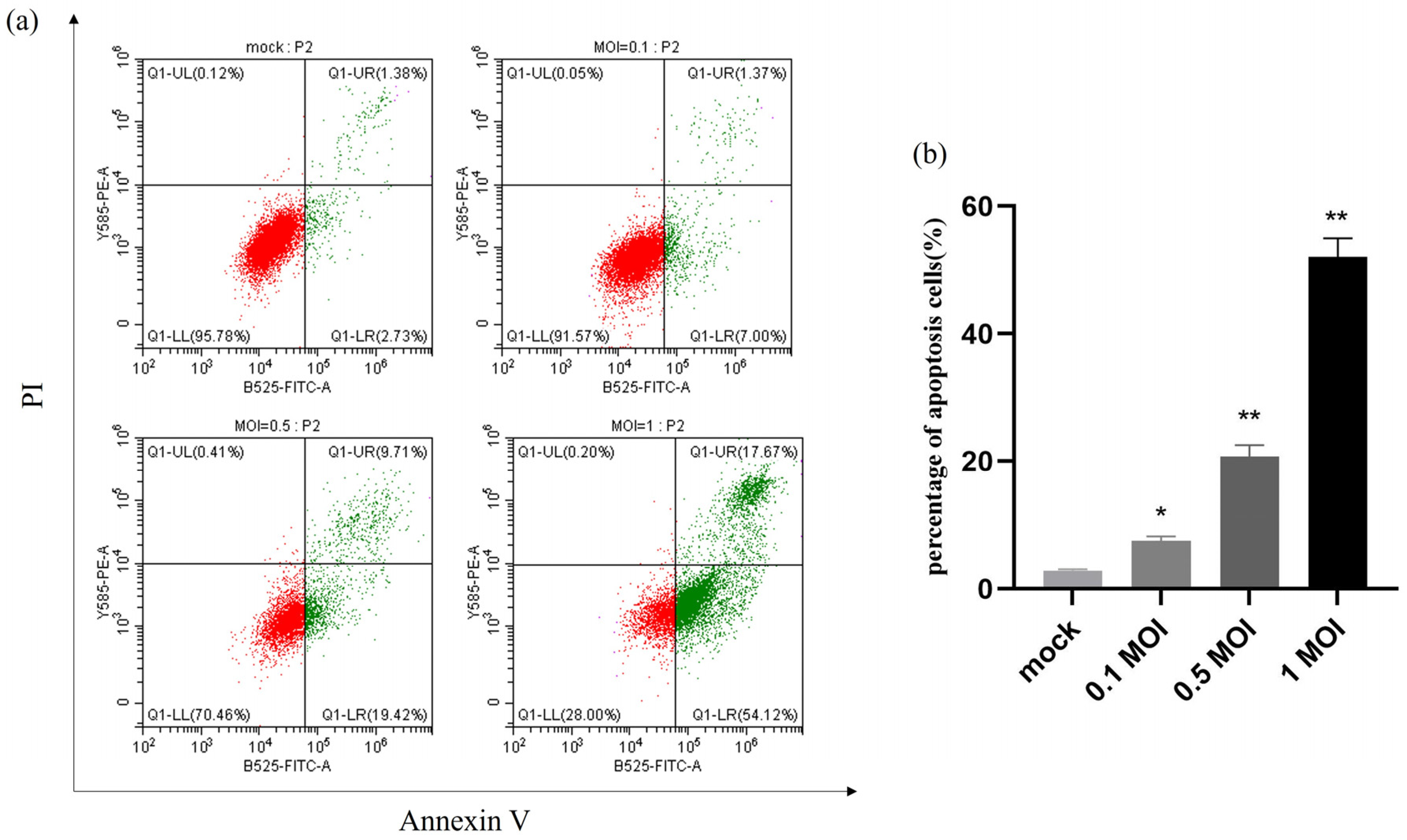
2.6. Apoptosis Is Induced by PEDV Infection in a Dose-Dependent Manner
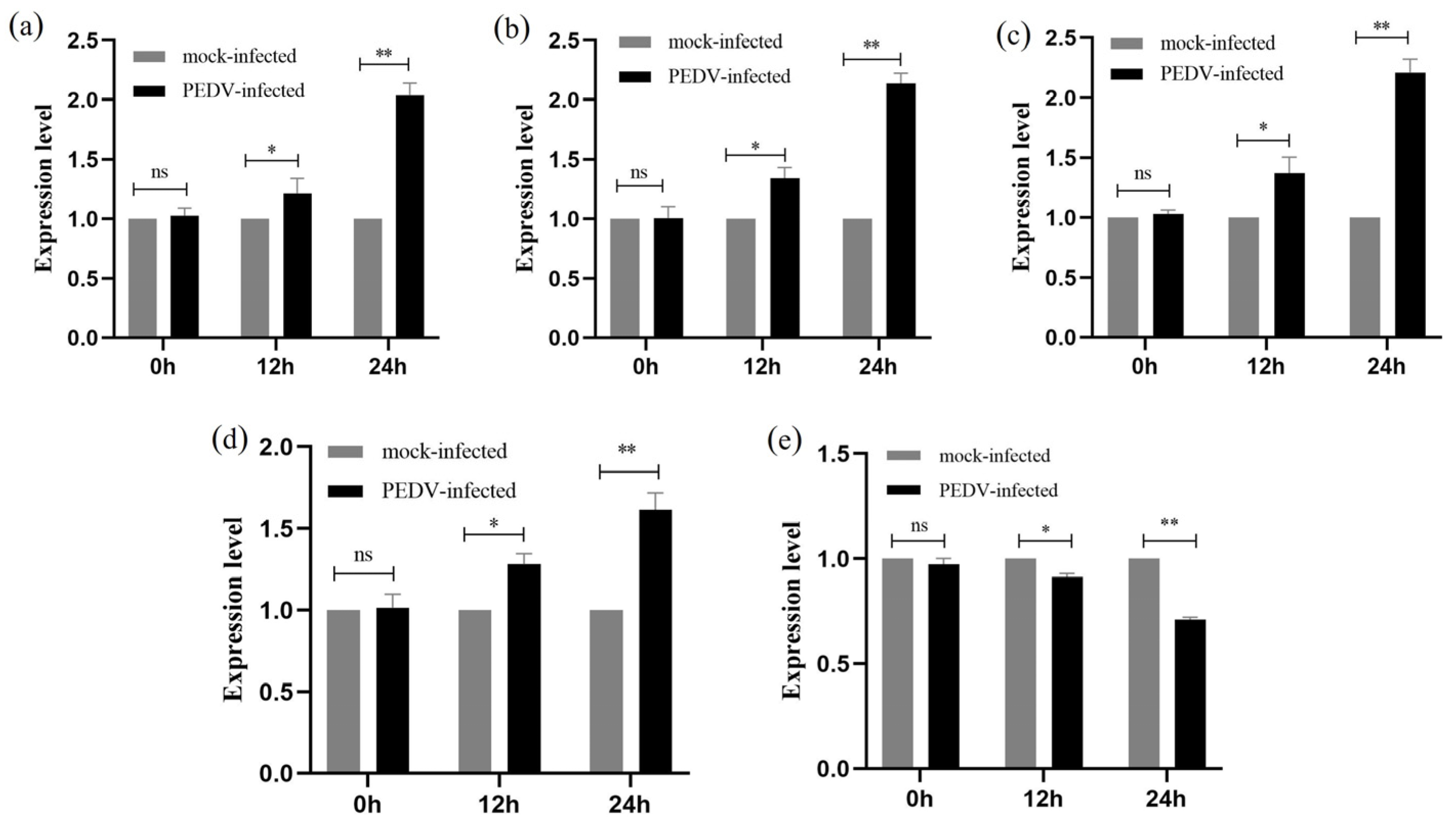
2.7. qRT-PCR to Detect the Relative Expression of Genes at Various Stages of PEDV Infection of IPEC-J2 Cells
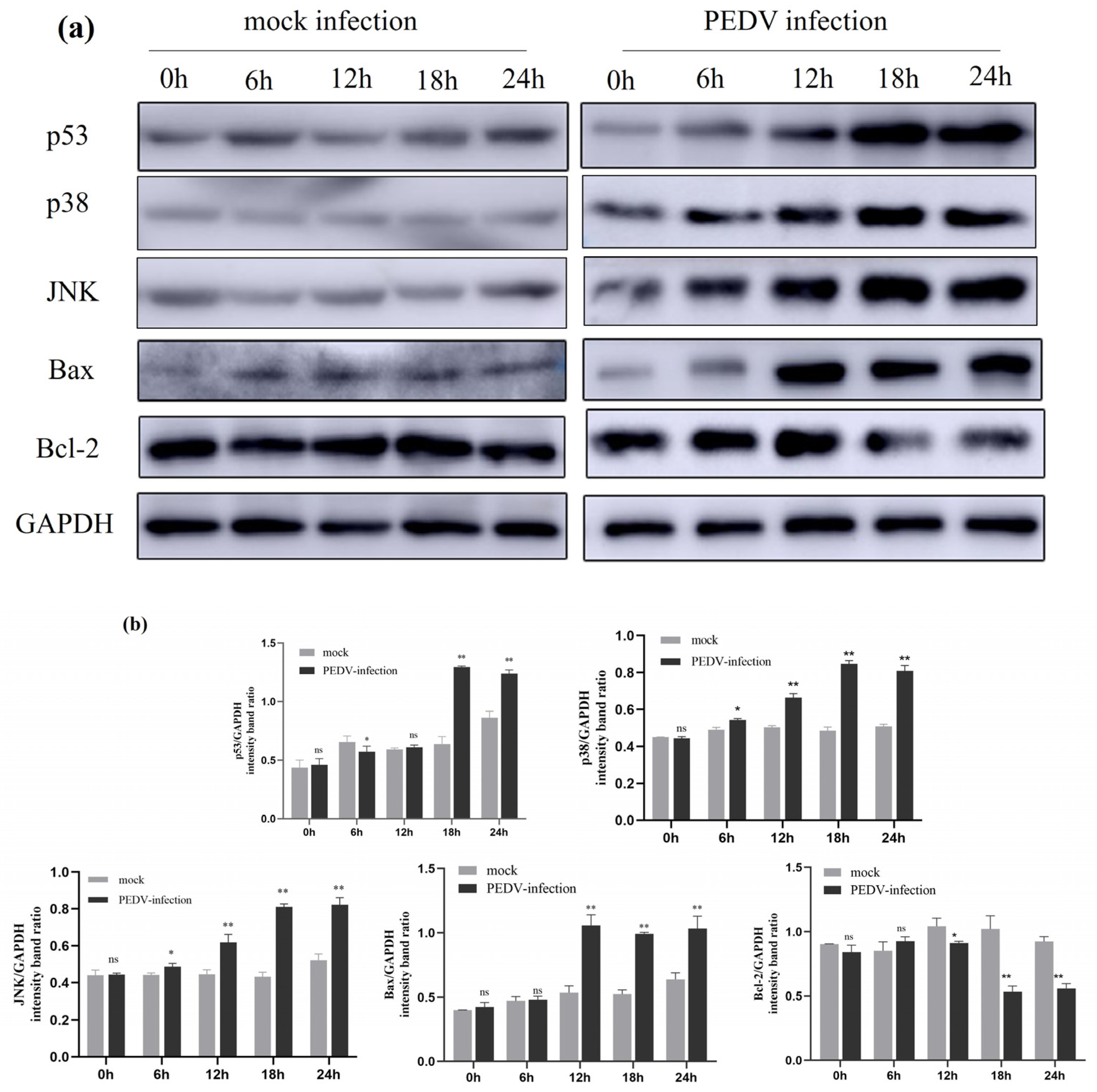
2.8. Expression of Activation-Associated Signalling Proteins by PEDV Infection on IPEC-J2 Cells
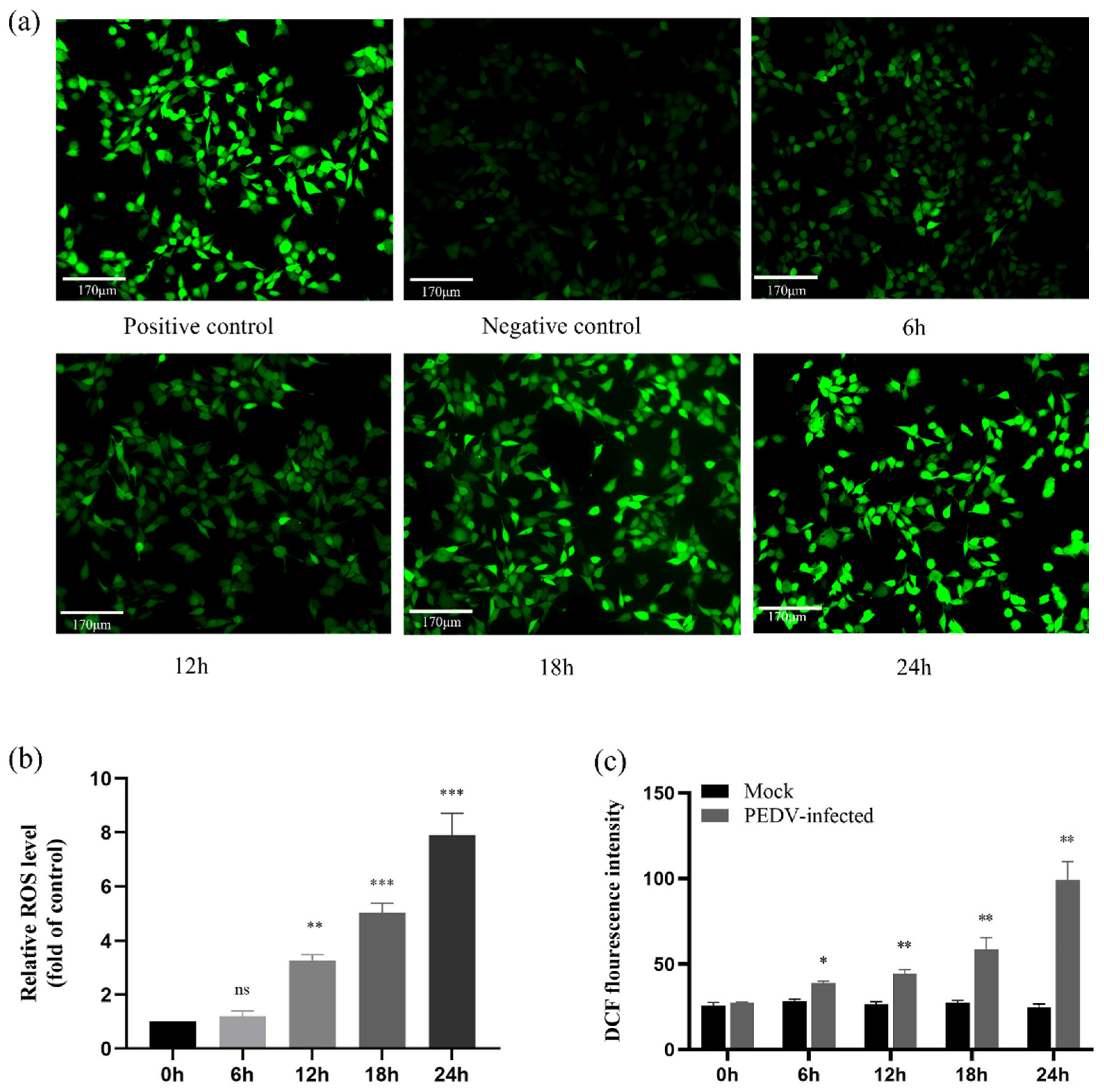
2.9. ROS Accumulate in PEDV-Infected IPEC-J2 Cells in a Time-Dependent Manner
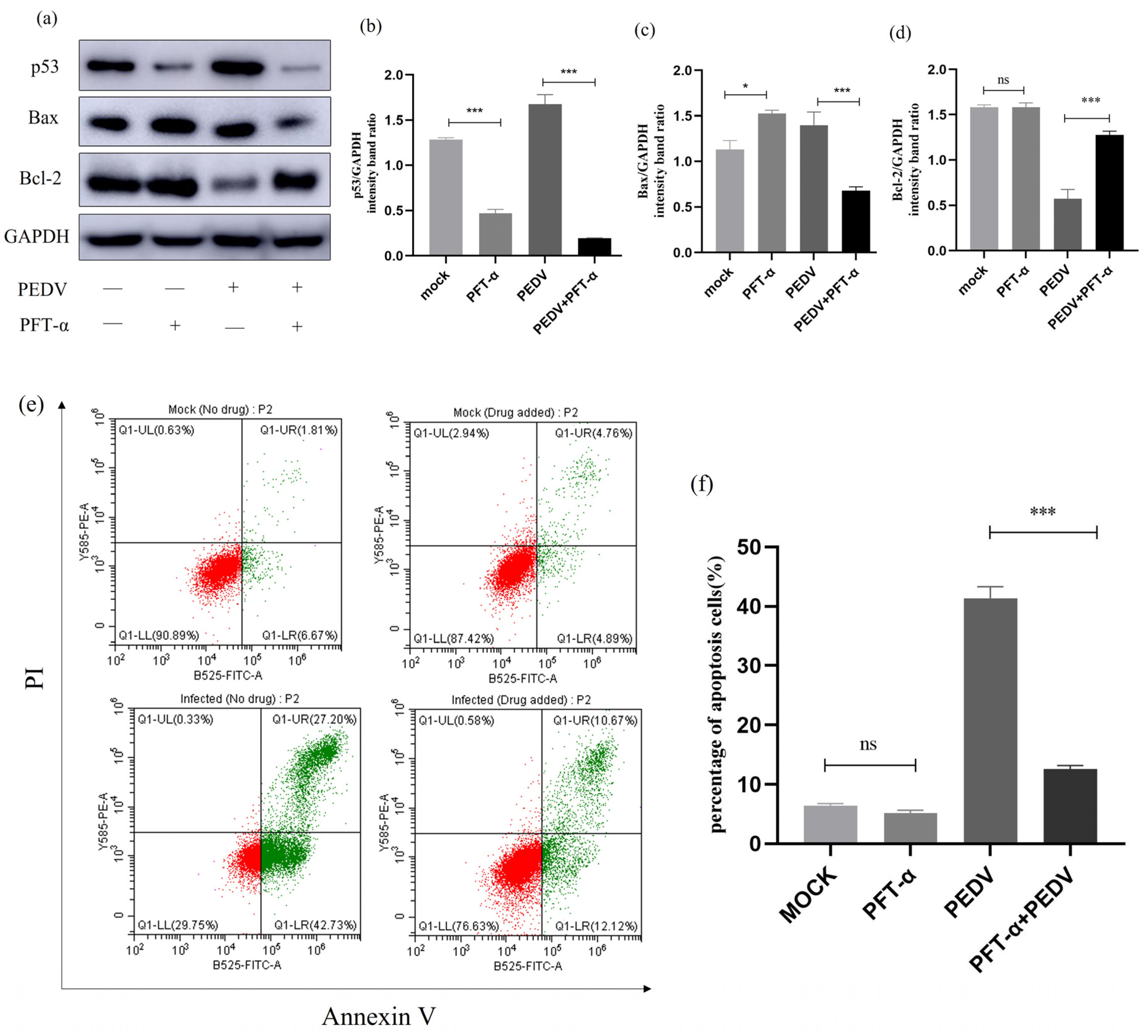
2.10. Effect of Inhibition of p53 Signalling Pathway on Apoptosis and Apoptotic Proteins
2.11. Effects of p38 MAPK Inhibition on Apoptosis and Apoptotic Proteins
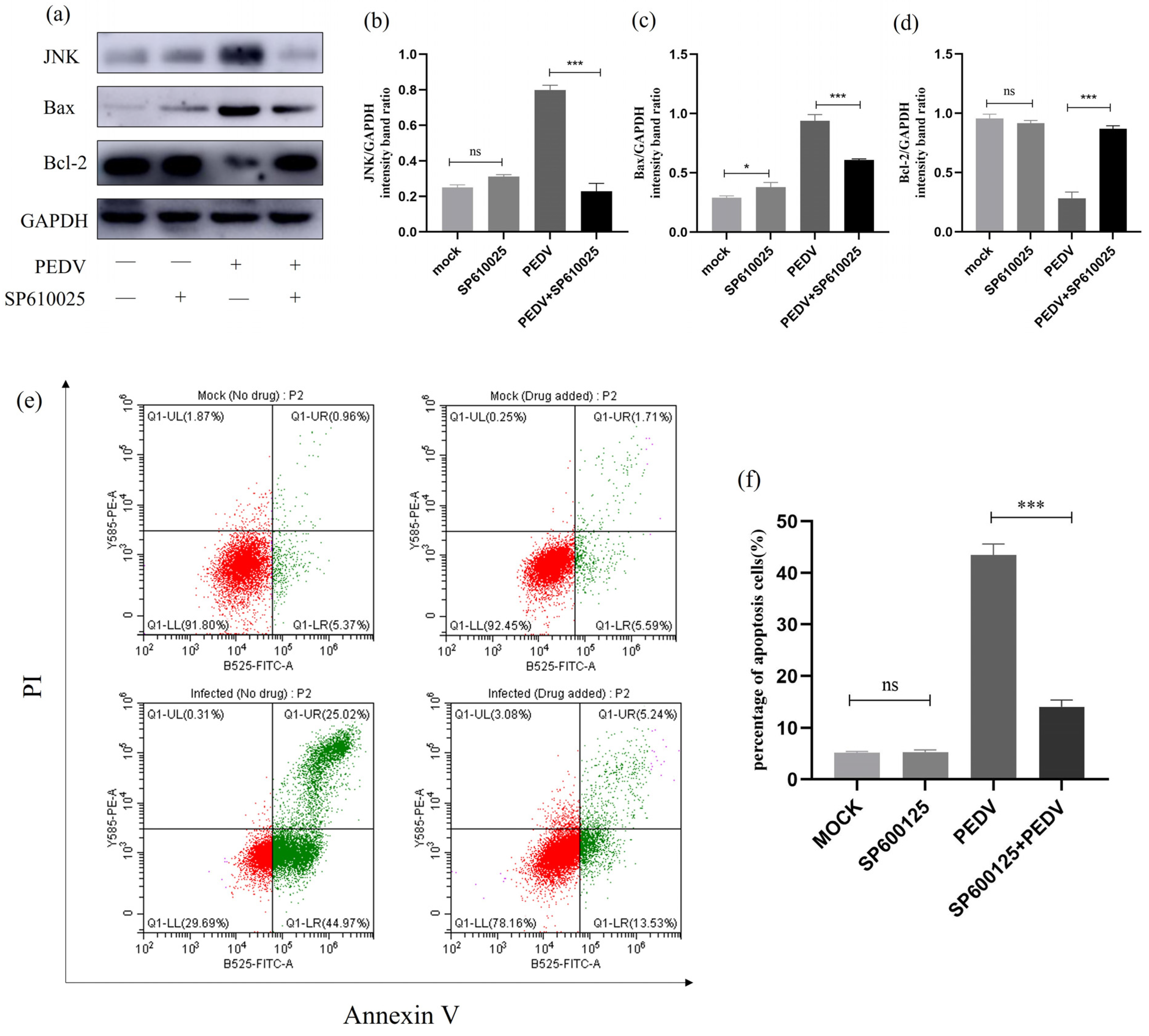
2.12. Effect of JNK Inhibition on Apoptosis and Apoptotic Proteins
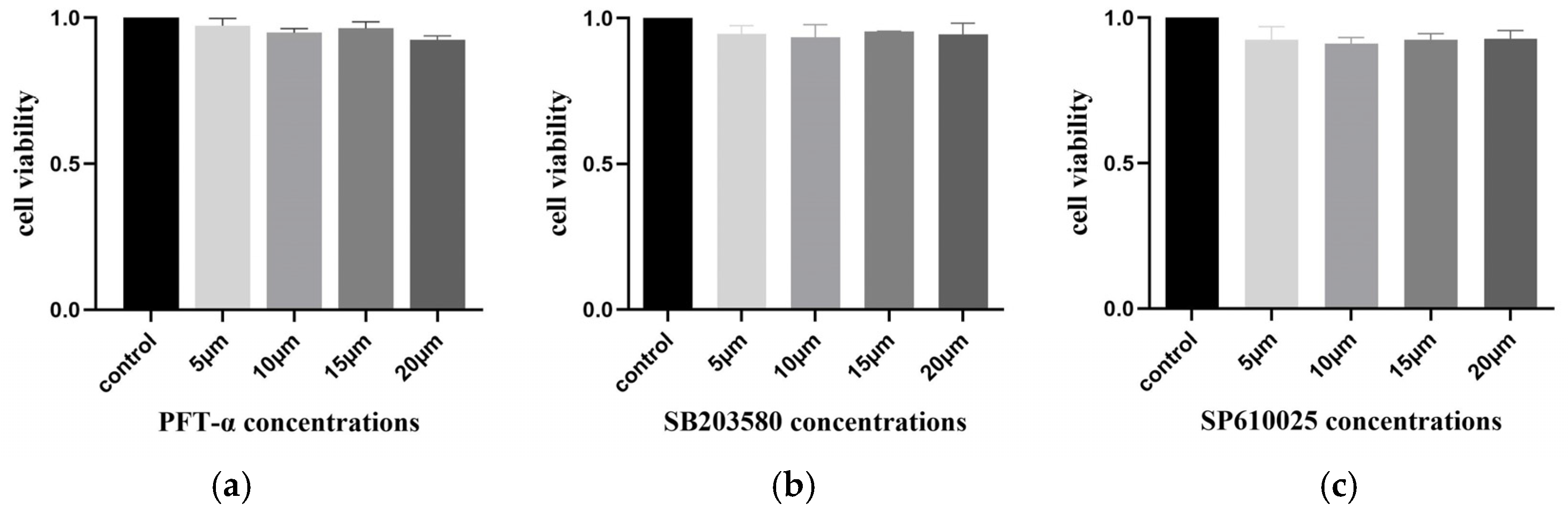
2.13. No Inhibitory Effect of Inhibitors on Cell Activity
3. Discussion
4. Materials and Methods
4.1. Cells
4.2. Inhibitors and Antibodies
4.3. Disease Material Collection and Handling
4.4. Virus Isolation and Culture
4.5. Virus Purification by Density Gradient Centrifugation
4.6. Detection of Viral Titres by the TCID50 Method
4.7. Identification of Viruses
4.7.1. Real-Time PCR Assay
4.7.2. Indirect Immunofluorescence for PEDV
4.7.3. Cell Ultra-Thin Section Observation
4.7.4. Analysis of PEDV Genetic Evolution
4.8. Apoptosis Assay
4.9. Real-Time Quantitative PCR to Detect the Expression of Related Genes
4.10. Western Blot Assay Analysis
4.11. ROS Detection
4.12. Inhibitor Assay
4.13. MTT Assay
4.14. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lin, F.; Zhang, H.; Li, L.; Yang, Y.; Zou, X.; Chen, J.; Tang, X. PEDV: Insights and Advances into Types, Function, Structure, and Receptor Recognition. Viruses 2022, 14, 1744. [Google Scholar] [CrossRef]
- Shibata, I.; Tsuda, T.; Mori, M.; Ono, M.; Sueyoshi, M.; Uruno, K. Isolation of porcine epidemic diarrhea virus in porcine cell cultures and experimental infection of pigs of different ages. Vet. Microbiol. 2000, 72, 173–182. [Google Scholar] [CrossRef]
- Debouck, P.; Pensaert, M. Experimental infection of pigs with a new porcine enteric coronavirus, CV777. Am. J. Vet. Res. 1980, 41, 219–223. [Google Scholar]
- Fan, B.; Jiao, D.; Zhang, R.; Zhou, J.; Guo, R.; Yu, Z.; Shi, D.; Zhao, Y.; Gu, J.; Niu, B.; et al. Origin and epidemic status of porcine epidemic diarrhea virus variants in China. Transbound. Emerg. Dis. 2020, 67, 1364–1370. [Google Scholar] [CrossRef]
- Kirchdoerfer, R.N.; Bhandari, M.; Martini, O.; Sewall, L.M.; Bangaru, S.; Yoon, K.J.; Ward, A.B. Structure and immune recognition of the porcine epidemic diarrhea virus spike protein. Structure 2021, 29, 385–392. [Google Scholar] [CrossRef]
- Su, M.; Li, C.; Qi, S.; Yang, D.; Jiang, N.; Yin, B.; Guo, D.; Kong, F.; Yuan, D.; Feng, L.; et al. A molecular epidemiological investigation of PEDV in China: Characterization of co-infection and genetic diversity of S1-based genes. Transbound. Emerg. Dis. 2020, 67, 1129–1140. [Google Scholar] [CrossRef]
- Wang, D.; Fang, L.; Xiao, S. Porcine epidemic diarrhea in China. Virus Res. 2016, 226, 7–13. [Google Scholar] [CrossRef]
- Brian, D.A.; Baric, R.S. Coronavirus genome structure and replication. Curr. Top. Microbiol. Immunol. 2005, 287, 1–30. [Google Scholar]
- Jung, K.; Saif, L.J. Porcine epidemic diarrhea virus infection: Etiology, epidemiology, pathogenesis and immunoprophylaxis. Vet. J. 2015, 204, 134–143. [Google Scholar] [CrossRef]
- Li, Z.; Ma, Z.; Li, Y.; Gao, S.; Xiao, S. Porcine epidemic diarrhea virus: Molecular mechanisms of attenuation and vaccines. Microb. Pathog. 2020, 149, 104553. [Google Scholar] [CrossRef]
- Tian, Y.; Yang, X.; Li, H.; Ma, B.; Guan, R.; Yang, J.; Chen, D.; Han, X.; Zhou, L.; Song, Z.; et al. Molecular characterization of porcine epidemic diarrhea virus associated with outbreaks in southwest China during 2014–2018. Transbound. Emerg. Dis. 2021, 68, 3482–3497. [Google Scholar] [CrossRef]
- Schierack, P.; Nordhoff, M.; Pollmann, M.; Weyrauch, K.D.; Amasheh, S.; Lodemann, U.; Jores, J.; Tachu, B.; Kleta, S.; Blikslager, A.; et al. Characterization of a porcine intestinal epithelial cell line for in vitro studies of microbial pathogenesis in swine. Histochem. Cell Biol. 2006, 125, 293–305. [Google Scholar] [CrossRef]
- Curry, S.M.; SchwaReal-timez, K.J.; Yoon, K.J.; Gabler, N.K.; Burrough, E.R. Effects of porcine epidemic diarrhea virus infection on nursery pig intestinal function and barrier integrity. Vet. Microbiol. 2017, 211, 58–66. [Google Scholar] [CrossRef]
- Li, Z.; Ma, Z.; Dong, L.; Yang, T.; Li, Y.; Jiao, D.; Han, W.; Zheng, H.; Xiao, S. Molecular Mechanism of Porcine Epidemic Diarrhea Virus Cell Tropism. MBio 2022, 13, e0373921. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, H.; Yu, J.; Feng, R.; Chen, Z.; Zhang, X.; Ren, Y.; Yang, G.; Huang, X.; Li, G. Comparative transcriptomic analysis of porcine epidemic diarrhea virus epidemic and classical strains in IPEC-J2 cells. Vet. Microbiol. 2022, 273, 109540. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lee, C. Porcine epidemic diarrhea virus induces caspase-independent apoptosis through activation of mitochondrial apoptosis-inducing factor. Virology 2014, 460–461, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Yin, L.; Pan, X.; Zhao, R.; Zhang, D. Porcine epidemic diarrhea virus infection blocks cell cycle and induces apoptosis in pig intestinal epithelial cells. Microb. Pathog. 2020, 147, 104378. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Xu, Y.; Zhang, Q.; Yang, F.; Yin, Z.; Wang, L.; Li, Q. Porcine epidemic diarrhea virus infections induce apoptosis in Vero cells via a reactive oxygen species (ROS)/p53, but not p38 MAPK and SAPK/JNK signalling pathways. Vet. Microbiol. 2019, 232, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, C.; Shu, J.; Feng, H.; He, Y.; Chen, J.; Shu, J. Porcine Epidemic Diarrhea Virus Induces Vero Cell Apoptosis via the p53-PUMA Signaling Pathway. Viruses 2021, 13, 1218. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewski, S.S.; Richter, J.F.; Krug, S.M.; Jebautzke, B.; Lee, I.-F.M.; Rieger, J.; Sachtleben, M.; Bondzio, A.; Schulzke, J.D.; Fromm, M.; et al. Improved cell line IPEC-J2, characterized as a model for porcine jejunal epithelium. PLoS ONE 2013, 8, e79643. [Google Scholar] [CrossRef] [PubMed]
- Altawaty, T.; Liu, L.; Zhang, H.; Tao, C.; Hou, S.; Li, K.; Wang, Y. Lack of LTβR Increases Susceptibility of IPEC-J2 Cells to Porcine Epidemic Diarrhea Virus. Cells 2018, 7, 222. [Google Scholar] [CrossRef]
- Lin, H.; Li, B.; Chen, L.; Ma, Z.; He, K.; Fan, H. Differential Protein Analysis of IPEC-J2 Cells Infected with Porcine Epidemic Diarrhea Virus Pandemic and Classical Strains Elucidates the Pathogenesis of Infection. J. Proteome Res. 2017, 16, 2113–2120. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, Z.; Li, J.; Gao, Y.; Zhou, L.; Ge, X.; Han, J.; Guo, X.; Yang, H. Porcine epidemic diarrhea virus S1 protein is the critical inducer of apoptosis. Virol. J. 2018, 15, 170. [Google Scholar] [CrossRef]
- Wang, L.; Wang, X.; Chang, J.; Wang, P.; Liu, C.; Yuan, L.; Yin, Q.; Zhu, Q.; Lu, F. Effect of the Combined Compound Probiotics with Glycyrrhinic Acid on Alleviating Cytotoxicity of IPEC-J2 Cells Induced by Multi-Mycotoxins. Toxins 2022, 14, 670. [Google Scholar] [CrossRef]
- Nazim, U.M.; Park, S.-Y. Attenuation of autophagy flux by 6-shogaol sensitizes human liver cancer cells to TRAIL-induced apoptosis via p53 and ROS. Int. J. Mol. Med. 2019, 43, 701–708. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, S. Mitogen-activated protein kinase cascades in signaling plant growth and development. Trends Plant Sci. 2015, 20, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, G.W.; Hoang, H.; SchwaReal-timez, K.J.; Burrough, E.R.; Sun, D.; Madson, D.; Cooper, V.L.; Pillatzki, A.; Gauger, P.; Schmitt, B.J.; et al. Emergence of Porcine epidemic diarrhea virus in the United States: Clinical signs, lesions, and viral genomic sequences. J. Vet. Diagn. Investig. 2013, 25, 649–654. [Google Scholar] [CrossRef]
- Sun, D.; Wang, X.; Wei, S.; Chen, J.; Feng, L. Epidemiology and vaccine of porcine epidemic diarrhea virus in China: A mini-review. J. Vet. Med. Sci. 2016, 78, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Si, F.; Hu, X.; Wang, C.; Chen, B.; Wang, R.; Dong, S.; Yu, R.; Li, Z. Porcine Epidemic Diarrhea Virus (PEDV) ORF3 Enhances Viral Proliferation by Inhibiting Apoptosis of Infected Cells. Viruses 2020, 12, 214. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Kepp, O.; Trojel-Hansen, C.; Kroemer, G. Mitochondrial control of cellular life, stress, and death. Circ. Res. 2012, 111, 1198–1207. [Google Scholar] [CrossRef]
- Kvansakul, M. Viral Infection and Apoptosis. Viruses 2017, 9, 356. [Google Scholar] [CrossRef]
- Ampomah, P.B.; Lim, L.H.K. Influenza A virus-induced apoptosis and virus propagation. Apoptosis 2020, 25, 1–11. [Google Scholar] [CrossRef]
- Verburg, S.G.; Lelievre, R.M.; Westerveld, M.J.; Inkol, J.M.; Sun, Y.L.; Workenhe, S.T. Viral-mediated activation and inhibition of programmed cell death. PLoS Pathog. 2022, 18, e1010718. [Google Scholar] [CrossRef]
- Danthi, P. Viruses and the Diversity of Cell Death. Annu. Rev. Virol. 2016, 3, 533–553. [Google Scholar] [CrossRef] [PubMed]
- Barber, G.N. Host defense, viruses and apoptosis. Cell Death Differ. 2001, 8, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Laptenko, O.; Prives, C. Transcriptional regulation by p53: One protein, many possibilities. Cell Death Differ. 2006, 13, 951–961. [Google Scholar] [CrossRef]
- Hafner, A.; Bulyk, M.L.; Jambhekar, A.; Lahav, G. The multiple mechanisms that regulate p53 activity and cell fate. Nat. Rev. Mol. Cell Biol. 2019, 20, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Wu, H.; Huang, J.; Xu, Y.; Yang, F.; Zhang, Q.; Xu, X. Porcine epidemic diarrhea virus through p53-dependent pathway causes cell cycle arrest in the G0/G1 phase. Virus Res. 2018, 253, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Huang, Y.; Du, Q.; Luo, X.; Zhang, L.; Zhao, X.; Tong, D. Porcine parvovirus infection induces apoptosis in PK-15 cells through activation of p53 and mitochondria-mediated pathway. Biochem. Biophys. Res. Commun. 2015, 456, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhu, Y.; Yan, S.; Xiao, H.; Yi, J.; Li, R.; Wu, J.; Wen, L. Phenethyl isothiocyanate induces IPEC-J2 cells cytotoxicity and apoptosis via S-G2/M phase arrest and mitochondria-mediated Bax/Bcl-2 pathway. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 226, 108574. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Tang, L. Bcl-2 Family Proteins Regulate Apoptosis and Epithelial to Mesenchymal Transition by Calcium Signals. Curr. Pharm. Des. 2016, 22, 4700–4704. [Google Scholar] [CrossRef]
- Moldoveanu, T.; Czabotar, P.E. BAX, BAK, and BOK: A Coming of Age for the BCL-2 Family Effector Proteins. Cold Spring Harb. Perspect. Biol. 2020, 12, a036319. [Google Scholar] [CrossRef]
- Carpio, M.A.; Michaud, M.; Zhou, W.; Fisher, J.K.; Walensky, L.D.; Katz, S.G. BCL-2 family member BOK promotes apoptosis in response to endoplasmic reticulum stress. Proc. Natl. Acad. Sci. USA 2015, 112, 7201–7206. [Google Scholar] [CrossRef] [PubMed]
- Czabotar, P.E.; Garcia-Saez, A.J. Mechanisms of BCL-2 family proteins in mitochondrial apoptosis. Nat. Rev. Mol. Cell Biol. 2023, 24, 732–748. [Google Scholar] [CrossRef]
- Huang, Y.-K.; Chang, K.-C.; Li, C.-Y.; Lieu, A.-S.; Lin, C.-L. AKR1B1 Represses Glioma Cell Proliferation through p38 MAPK-Mediated Bcl-2/BAX/Caspase-3 Apoptotic Signaling Pathways. Curr. Issues Mol. Biol. 2023, 45, 3391–3405. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Xiang, H.; Bai, X.; Fei, N.; Huang, Y.; Song, X.; Zhang, H.; Zhang, L.; Tong, D. Porcine parvovirus infection activates mitochondria-mediated apoptotic signaling pathway by inducing ROS accumulation. Virol. J. 2016, 13, 26. [Google Scholar] [CrossRef]
- Niemietz, I.; Brown, K.L. Hyaluronan promotes intracellular ROS production and apoptosis in TNFα-stimulated neutrophils. Front. Immunol. 2023, 14, 1032469. [Google Scholar] [CrossRef]
- Hao, W.; Wang, S.; Zhou, Z. Tubeimoside-1 (TBMS1) inhibits lung cancer cell growth and induces cells apoptosis through activation of MAPK-JNK pathway. Int. J. Clin. Exp. Pathol. 2015, 8, 12075–12083. [Google Scholar]
- Zhong, W.; Zhu, H.; Sheng, F.; Tian, Y.; Zhou, J.; Chen, Y.; Li, S.; Lin, J. Activation of the MAPK11/12/13/14 (p38 MAPK) pathway regulates the transcription of autophagy genes in response to oxidative stress induced by a novel copper complex in HeLa cells. Autophagy 2014, 10, 1285–1300. [Google Scholar] [CrossRef]
- Miura, H.; Kondo, Y.; Matsuda, M.; Aoki, K. Cell-to-Cell Heterogeneity in p38-Mediated Cross-Inhibition of JNK Causes Stochastic Cell Death. Cell Rep. 2018, 24, 2658–2668. [Google Scholar] [CrossRef] [PubMed]
- Geens, M.M.; Niewold, T.A. Optimizing culture conditions of a porcine epithelial cell line IPEC-J2 through a histological and physiological characterization. Cytotechnology 2011, 63, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Jia, S.; Zhao, H.; Yin, J.; Wang, X.; Yu, M.; Ma, S.; Wu, Y.; Chen, Y.; Fan, W.; et al. Novel Approach for Isolation and Identification of Porcine Epidemic Diarrhea Virus (PEDV) Strain NJ Using Porcine Intestinal Epithelial Cells. Viruses 2017, 9, 19. [Google Scholar] [CrossRef]
- Zhang, X. Isolation and Identification of PEDV Strains and Establishment of Antibody ELISA Detection Methods; Chinese Academy of Agricultural Sciences: Beijing, China, 2021. (In Chinese) [Google Scholar]
- Chen, G.; Yang, K.; Tang, Q. Isolation and culture of a porcine epidemic diarrhoea virus and sequence analysis of its ORF3, sM, and N genes. Hunan Anim. Husb. Vet. Med. 2020, 5, 47–52. (In Chinese) [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers Name | Forward Primer (5′-3′) | Reversed Primer (5′-3′) |
|---|---|---|
| p53 | CCAGATGACGCCTCCAGAGTG | TGAGAAGGGACAAAGGACGACAG |
| p38 | ATCTCATTAACAGGATGCCAAGCC | CCAGCAAGTCAACAGCCAAGG |
| JNK | CCACCACCAAAGATACCTGACAAG | GGTTCTCTCCTCCAAGTCCATAAC |
| BAX | CATCTACCAAGAAGTTGAGCGAGTG | ACGGCTGCGATCATCCTCTG |
| Bcl-2 | CGCAGAGGGGCTACGAGTG | CGGGCTGGGAGGAGAAGATG |
| GAPDH | GCGACTTCAACAGCAACTCCC | CACCCTGTTGCTGTAGCCGTA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Wang, M.; Xi, Y.; Qiu, S.; Zeng, Q.; Pan, Y. Isolation and Identification of a Tibetan Pig Porcine Epidemic Diarrhoea Virus Strain and Its Biological Effects on IPEC-J2 Cells. Int. J. Mol. Sci. 2024, 25, 2200. https://doi.org/10.3390/ijms25042200
Li M, Wang M, Xi Y, Qiu S, Zeng Q, Pan Y. Isolation and Identification of a Tibetan Pig Porcine Epidemic Diarrhoea Virus Strain and Its Biological Effects on IPEC-J2 Cells. International Journal of Molecular Sciences. 2024; 25(4):2200. https://doi.org/10.3390/ijms25042200
Chicago/Turabian StyleLi, Mei, Meng Wang, Yao Xi, Shantong Qiu, Qiaoying Zeng, and Yangyang Pan. 2024. "Isolation and Identification of a Tibetan Pig Porcine Epidemic Diarrhoea Virus Strain and Its Biological Effects on IPEC-J2 Cells" International Journal of Molecular Sciences 25, no. 4: 2200. https://doi.org/10.3390/ijms25042200
APA StyleLi, M., Wang, M., Xi, Y., Qiu, S., Zeng, Q., & Pan, Y. (2024). Isolation and Identification of a Tibetan Pig Porcine Epidemic Diarrhoea Virus Strain and Its Biological Effects on IPEC-J2 Cells. International Journal of Molecular Sciences, 25(4), 2200. https://doi.org/10.3390/ijms25042200