
Exosomes from Adipose-Tissue-Derived Stem Cells Induce Proapoptotic Gene Expression in Breast Tumor Cell Line

 , ,
, , 

Abstract
1. Introduction
2. Results
2.1. Exosome Detection
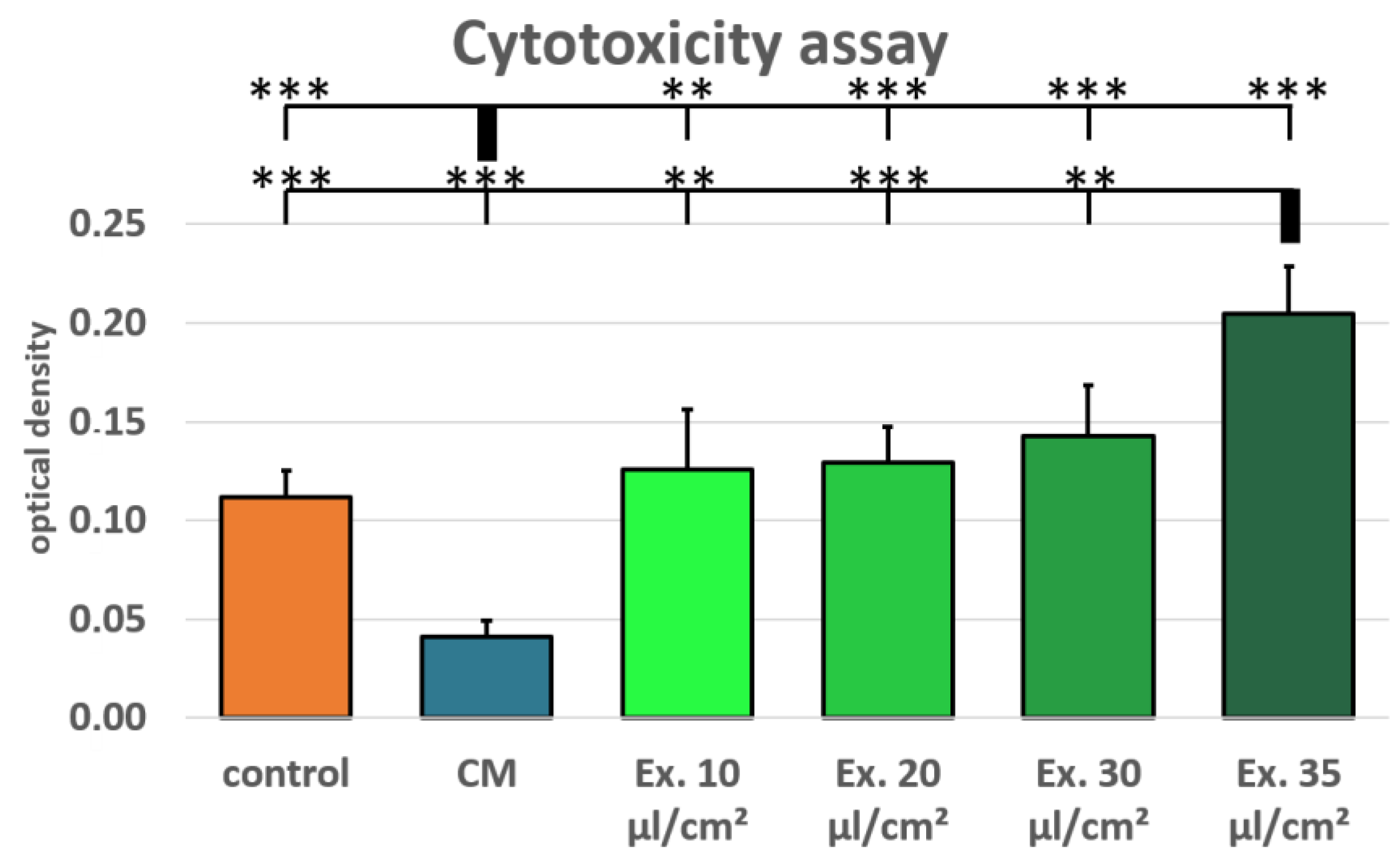
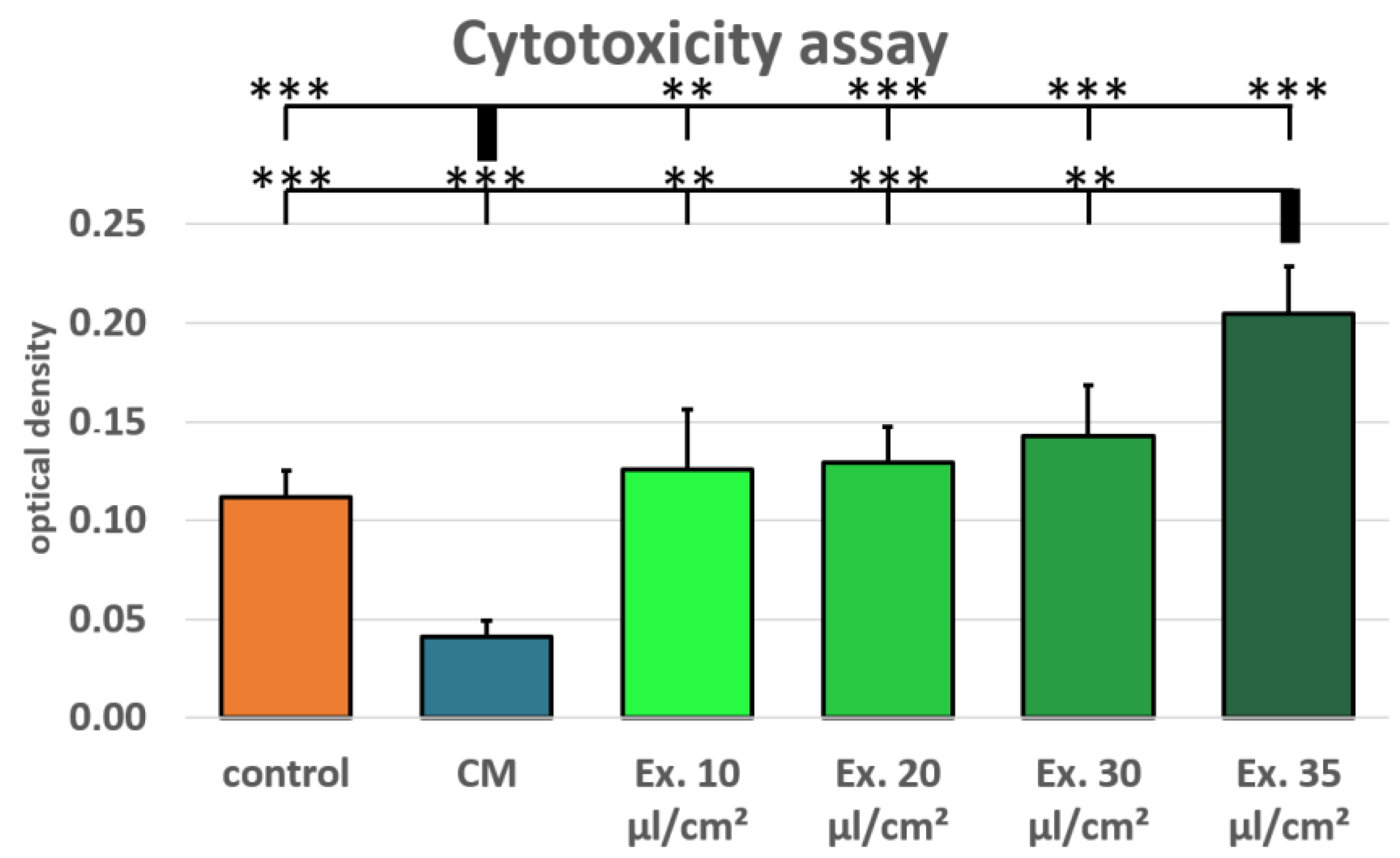
2.2. Cell Viability and Cytotoxicity
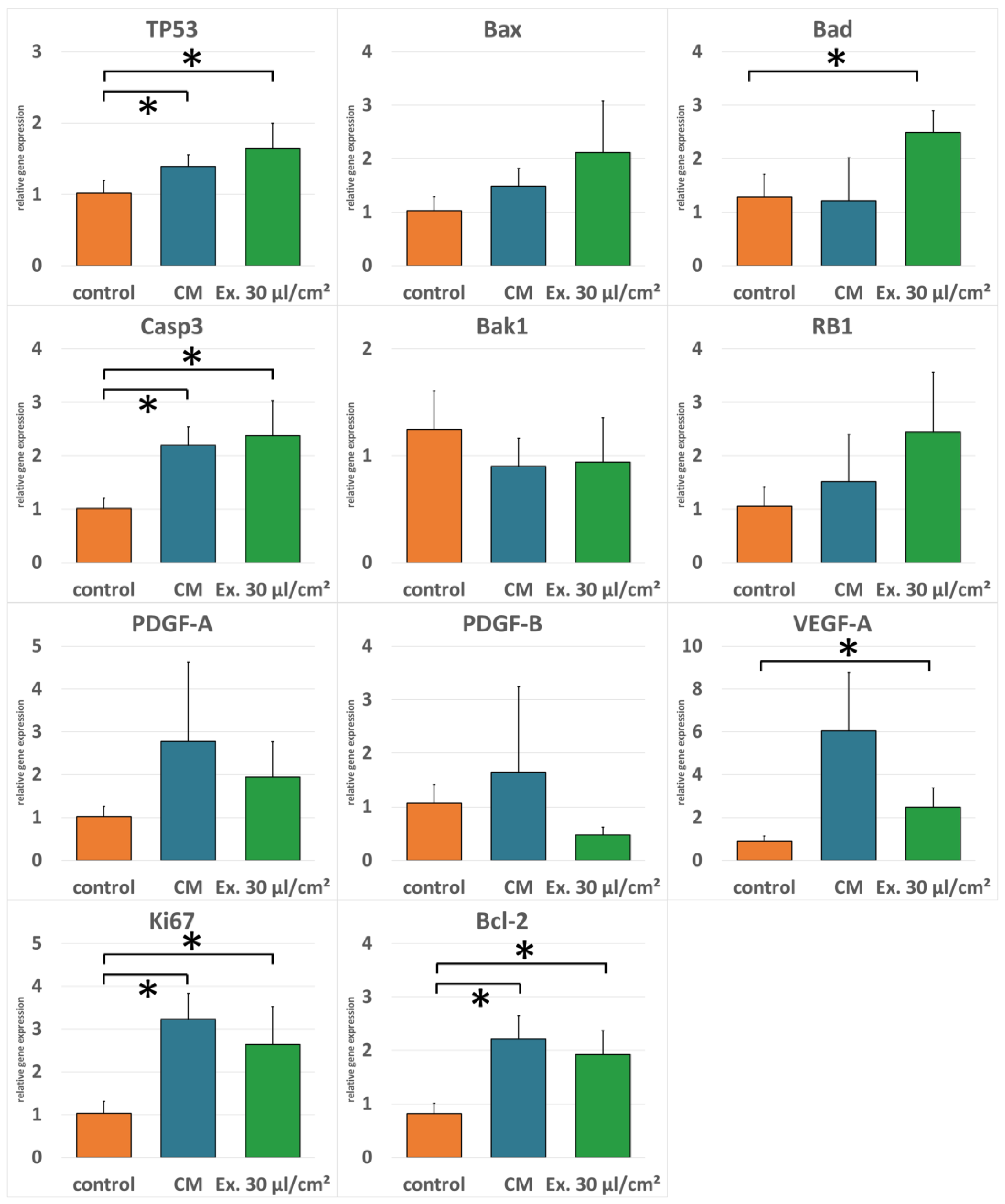
2.3. Gene Expression
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Medium Harvesting, Exosome Isolation, and Exosome Detection
4.3. Viability and Cytotoxicity Assays
4.4. RNA Isolation and RT-PCR
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ciuba, A.; Wnuk, K.; Nitsch-Osuch, A.; Kulpa, M. Health Care Accessibility and Breast Cancer Mortality in Europe. Int. J. Environ. Res. Public Health 2022, 19, 13605. [Google Scholar] [CrossRef]
- Sant, M.; Francisci, S.; Capocaccia, R.; Verdecchia, A.; Allemani, C.; Berrino, F. Time Trends of Breast Cancer Survival in Europe in Relation to Incidence and Mortality. Int. J. Cancer 2006, 119, 2417–2422. [Google Scholar] [CrossRef]
- Canelo-Aybar, C.; Ferreira, D.S.; Ballesteros, M.; Posso, M.; Montero, N.; Solà, I.; Saz-Parkinson, Z.; Lerda, D.; Rossi, P.G.; Duffy, S.W.; et al. Benefits and Harms of Breast Cancer Mammography Screening for Women at Average Risk of Breast Cancer: A Systematic Review for the European Commission Initiative on Breast Cancer. J. Med. Screen. 2021, 28, 389–404. [Google Scholar] [CrossRef]
- Jonczyk, M.M.; Jean, J.; Graham, R.; Chatterjee, A. Surgical Trends in Breast Cancer: A Rise in Novel Operative Treatment Options over a 12 Year Analysis. Breast Cancer Res. Treat. 2019, 173, 267–274. [Google Scholar] [CrossRef]
- Zhu, H.; Doğan, B.E. American Joint Committee on Cancer’s Staging System for Breast Cancer, Eighth Edition: Summary for Clinicians. Eur. J. Breast Health 2021, 17, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Burguin, A.; Diorio, C.; Durocher, F. Breast Cancer Treatments: Updates and New Challenges. J. Pers. Med. 2021, 11, 808. [Google Scholar] [CrossRef]
- Vieira, R.A.d.C.; Bailão-Junior, A.; de Oliveira-Junior, I. Does Breast Oncoplastic Surgery Improve Quality of Life? Front. Oncol. 2022, 12, 1099125. [Google Scholar] [CrossRef] [PubMed]
- Djohan, R.; Gage, E.; Bernard, S. Breast Reconstruction Options Following Mastectomy. Cleve Clin. J. Med. 2008, 75 (Suppl. S1), S17–S23. [Google Scholar] [CrossRef] [PubMed]
- Atisha, D.; Alderman, A.K.; Lowery, J.C.; Kuhn, L.E.; Davis, J.; Wilkins, E.G. Prospective Analysis of Long-Term Psychosocial Outcomes in Breast Reconstruction: Two-Year Postoperative Results from the Michigan Breast Reconstruction Outcomes Study. Ann. Surg. 2008, 247, 1019–1028. [Google Scholar] [CrossRef]
- Krastev, T.K.; Schop, S.J.; Hommes, J.; Piatkowski, A.A.; Heuts, E.M.; van der Hulst, R.R.W.J. Meta-Analysis of the Oncological Safety of Autologous Fat Transfer after Breast Cancer. Br. J. Surg. 2018, 105, 1082–1097. [Google Scholar] [CrossRef]
- Wederfoort, J.L.M.; Kleeven, A.; Hommes, J.E.; Van Kuijk, S.M.J.; van der Hulst, R.R.W.J.; Piatkowski, A. Aesthetic Evaluation of Breast Reconstruction with Autologous Fat Transfer vs. Implants. Aesthetic Plast. Surg. 2023, 47, 593–604. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human Adipose Tissue Is a Source of Multipotent Stem Cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- Kern, S.; Eichler, H.; Stoeve, J.; Klüter, H.; Bieback, K. Comparative Analysis of Mesenchymal Stem Cells from Bone Marrow, Umbilical Cord Blood, or Adipose Tissue. Stem Cells 2006, 24, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Bunnell, B.A. Adipose Tissue-Derived Mesenchymal Stem Cells. Cells 2021, 10, 3433. [Google Scholar] [CrossRef] [PubMed]
- Stark, R.Y.; Mirzabeigi, M.N.; Vonderhaar, R.J.; Bucky, L.P. Utilizing Large Volume Fat Grafting in Breast Reconstruction after Nipple Sparing Mastectomies. Gland. Surg. 2018, 7, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Weinzierl, A.; Schmauss, D.; Brucato, D.; Harder, Y. Implant-Based Breast Reconstruction after Mastectomy, from the Subpectoral to the Prepectoral Approach: An Evidence-Based Change of Mind? J. Clin. Med. 2022, 11, 3079. [Google Scholar] [CrossRef] [PubMed]
- Piccotti, F.; Rybinska, I.; Scoccia, E.; Morasso, C.; Ricciardi, A.; Signati, L.; Triulzi, T.; Corsi, F.; Truffi, M. Lipofilling in Breast Oncological Surgery: A Safe Opportunity or Risk for Cancer Recurrence? Int. J. Mol. Sci. 2021, 22, 3737. [Google Scholar] [CrossRef] [PubMed]
- Ohno, S.; Takanashi, M.; Sudo, K.; Ueda, S.; Ishikawa, A.; Matsuyama, N.; Fujita, K.; Mizutani, T.; Ohgi, T.; Ochiya, T.; et al. Systemically Injected Exosomes Targeted to EGFR Deliver Antitumor microRNA to Breast Cancer Cells. Mol. Ther. 2013, 21, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Rezaie, Z.; Ardeshirylajimi, A.; Ashkezari, M.D.; Seifati, S.M. Antitumoral Potential of Microvesicles Extracted from Human Adipose-Derived Mesenchymal Stem Cells on Human Breast Cancer Cells. J. Cancer Res. Ther. 2019, 15, 1114–1119. [Google Scholar] [CrossRef]
- Trzyna, A.; Banaś-Ząbczyk, A. Adipose-Derived Stem Cells Secretome and Its Potential Application in “Stem Cell-Free Therapy”. Biomolecules 2021, 11, 878. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Alonso-Alonso, M.L.; García-Posadas, L.; Diebold, Y. Extracellular Vesicles from Human Adipose-Derived Mesenchymal Stem Cells: A Review of Common Cargos. Stem Cell Rev. Rep. 2022, 18, 854–901. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-B.; Zhang, Z.-R.; Schluesener, H.J.; Xu, S.-Q. Role of Exosomes in Immune Regulation. J. Cell Mol. Med. 2006, 10, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Raychaudhuri, S. A Minimal Model of Signaling Network Elucidates Cell-to-Cell Stochastic Variability in Apoptosis. PLoS ONE 2010, 5, e11930. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, L.; Yuan, X.; Ou, Y.; Zhu, X.; Cheng, Z.; Zhang, P.; Wu, X.; Meng, Y.; Zhang, L. The Relationship between the Bcl-2/Bax Proteins and the Mitochondria-Mediated Apoptosis Pathway in the Differentiation of Adipose-Derived Stromal Cells into Neurons. PLoS ONE 2016, 11, e0163327. [Google Scholar] [CrossRef] [PubMed]
- Sanwlani, R.; Gangoda, L. Role of Extracellular Vesicles in Cell Death and Inflammation. Cells 2021, 10, 2663. [Google Scholar] [CrossRef] [PubMed]
- Rohrich, R.J. The American Society of Plastic Surgeons’ Procedural Statistics: What They Really Mean. Plast. Reconstr. Surg. 2003, 112, 1389–1392. [Google Scholar] [CrossRef] [PubMed]
- Gir, P.; Brown, S.A.; Oni, G.; Kashefi, N.; Mojallal, A.; Rohrich, R.J. Fat Grafting: Evidence-Based Review on Autologous Fat Harvesting, Processing, Reinjection, and Storage. Plast. Reconstr. Surg. 2012, 130, 249–258. [Google Scholar] [CrossRef]
- Simonacci, F.; Bertozzi, N.; Grieco, M.P.; Grignaffini, E.; Raposio, E. Procedure, Applications, and Outcomes of Autologous Fat Grafting. Ann. Med. Surg. 2017, 20, 49–60. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage Cells from Human Adipose Tissue: Implications for Cell-Based Therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [PubMed]
- Varghese, J.; Griffin, M.; Mosahebi, A.; Butler, P. Systematic Review of Patient Factors Affecting Adipose Stem Cell Viability and Function: Implications for Regenerative Therapy. Stem Cell Res. Ther. 2017, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Wang, W.; Kratz, K.; Xu, X.; Nie, Y.; Ma, N.; Lendlein, A. Evaluation of Human Mesenchymal Stem Cell Senescence, Differentiation and Secretion Behavior Cultured on Polycarbonate Cell Culture Inserts. Clin. Hemorheol. Microcirc. 2018, 70, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Krastev, T.K.; Jonasse, Y.; Kon, M. Oncological Safety of Autologous Lipoaspirate Grafting in Breast Cancer Patients: A Systematic Review. Ann. Surg. Oncol. 2013, 20, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Sekigami, Y.; Schwartz, T.; Losken, A.; Margenthaler, J.; Chatterjee, A. Lipofilling after Breast Conserving Surgery: A Comprehensive Literature Review Investigating Its Oncologic Safety. Gland. Surg. 2019, 8, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Stam, J.; Bartel, S.; Bischoff, R.; Wolters, J.C. Isolation of Extracellular Vesicles with Combined Enrichment Methods. J. Chromatogr. B Analyt Technol. Biomed. Life Sci. 2021, 1169, 122604. [Google Scholar] [CrossRef] [PubMed]
- Crescitelli, R.; Lässer, C.; Szabó, T.G.; Kittel, A.; Eldh, M.; Dianzani, I.; Buzás, E.I.; Lötvall, J. Distinct RNA Profiles in Subpopulations of Extracellular Vesicles: Apoptotic Bodies, Microvesicles and Exosomes. J. Extracell. Vesicles 2013, 2, 20677. [Google Scholar] [CrossRef]
- Tsai, Y.-C.; Cheng, T.-S.; Liao, H.-J.; Chuang, M.-H.; Chen, H.-T.; Chen, C.-H.; Zhang, K.-L.; Chang, C.-H.; Lin, P.-C.; Huang, C.-Y.F. Mesenchymal Stem Cell Secreted-Extracellular Vesicles Are Involved in Chondrocyte Production and Reduce Adipogenesis during Stem Cell Differentiation. Tissue Eng. Regen. Med. 2022, 19, 1295–1310. [Google Scholar] [CrossRef]
- Oksvold, M.P.; Neurauter, A.; Pedersen, K.W. Magnetic Bead-Based Isolation of Exosomes. Methods Mol. Biol. 2015, 1218, 465–481. [Google Scholar] [CrossRef]
- Kowal, E.J.K.; Ter-Ovanesyan, D.; Regev, A.; Church, G.M. Extracellular Vesicle Isolation and Analysis by Western Blotting. Methods Mol. Biol. 2017, 1660, 143–152. [Google Scholar] [CrossRef]
- Gebremeskel, S.; Gencarelli, J.; Gareau, A.J.; Levatte, T.; Dugandzic B Sc, A.; Johnston, B.; Bezuhly, M. Promotion of Primary Murine Breast Cancer Growth and Metastasis by Adipose-Derived Stem Cells Is Reduced in the Presence of Autologous Fat Graft. Plast. Reconstr. Surg. 2019, 143, 137–147. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Takahara, K.; Ii, M.; Inamoto, T.; Nakagawa, T.; Ibuki, N.; Yoshikawa, Y.; Tsujino, T.; Uchimoto, T.; Saito, K.; Takai, T.; et al. microRNA-145 Mediates the Inhibitory Effect of Adipose Tissue-Derived Stromal Cells on Prostate Cancer. Stem Cells Dev. 2016, 25, 1290–1298. [Google Scholar] [CrossRef] [PubMed]
- Barczyk, K.; Kreuter, M.; Pryjma, J.; Booy, E.P.; Maddika, S.; Ghavami, S.; Berdel, W.E.; Roth, J.; Los, M. Serum Cytochrome c Indicates in Vivo Apoptosis and Can Serve as a Prognostic Marker during Cancer Therapy. Int. J. Cancer 2005, 116, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Vitagliano, O.; Addeo, R.; D’Angelo, V.; Indolfi, C.; Indolfi, P.; Casale, F. The Bcl-2/Bax and Ras/Raf/MEK/ERK Signaling Pathways: Implications in Pediatric Leukemia Pathogenesis and New Prospects for Therapeutic Approaches. Expert. Rev. Hematol. 2013, 6, 587–597. [Google Scholar] [CrossRef]
- Ge, Y.-L.; Zhang, X.; Zhang, J.-Y.; Hou, L.; Tian, R.-H. The Mechanisms on Apoptosis by Inhibiting VEGF Expression in Human Breast Cancer Cells. Int. Immunopharmacol. 2009, 9, 389–395. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, J.; Chai, K.; Ying, X.; Zhou, B.P. The Role of Snail in EMT and Tumorigenesis. Curr. Cancer Drug Targets 2013, 13, 963–972. [Google Scholar] [CrossRef]
- Cekanova, M.; Fernando, R.I.; Siriwardhana, N.; Sukhthankar, M.; de la Parra, C.; Woraratphoka, J.; Malone, C.; Ström, A.; Baek, S.J.; Wade, P.A.; et al. BCL-2 Family Protein, BAD Is down-Regulated in Breast Cancer and Inhibits Cell Invasion. Exp. Cell Res. 2015, 331, 1–10. [Google Scholar] [CrossRef]
- Huun, J.; Lønning, P.E.; Knappskog, S. Effects of Concomitant Inactivation of P53 and pRb on Response to Doxorubicin Treatment in Breast Cancer Cell Lines. Cell Death Discov. 2017, 3, 17026. [Google Scholar] [CrossRef]
- Engeland, K. Cell Cycle Regulation: P53-P21-RB Signaling. Cell Death Differ. 2022, 29, 946–960. [Google Scholar] [CrossRef] [PubMed]
- Manning, A.L.; Benes, C.; Dyson, N.J. Whole Chromosome Instability Resulting from the Synergistic Effects of pRB and P53 Inactivation. Oncogene 2014, 33, 2487–2494. [Google Scholar] [CrossRef] [PubMed]
- Kempa, S.; Brix, E.; Heine, N.; Hösl, V.; Strauss, C.; Eigenberger, A.; Brébant, V.; Seitz, S.; Prantl, L. Autologous Fat Grafting for Breast Reconstruction after Breast Cancer: A 12-Year Experience. Arch. Gynecol. Obstet. 2022, 305, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Toyserkani, N.M.; Quaade, M.L.; Sørensen, J.A. Cell-Assisted Lipotransfer: A Systematic Review of Its Efficacy. Aesthetic Plast. Surg. 2016, 40, 309–318. [Google Scholar] [CrossRef]
- van Dongen, J.A.; van Boxtel, J.; Harmsen, M.C.; Stevens, H.P. The Development of Facial Lipofilling from a Historical Point of View. Facial Plast. Surg. 2019, 35, 358–367. [Google Scholar] [CrossRef]
- Schreml, S.; Babilas, P.; Fruth, S.; Orsó, E.; Schmitz, G.; Mueller, M.B.; Nerlich, M.; Prantl, L. Harvesting Human Adipose Tissue-Derived Adult Stem Cells: Resection versus Liposuction. Cytotherapy 2009, 11, 947–957. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence | Reverse Primer Sequence |
|---|---|---|
| Bad | 5′-TCACAAATCCTCCCCAAGTGG-3′ | 5′-GAATGCGCCCTAAATCACTGA-3′ |
| Bak1 | data | data |
| Bax | 5′-TTCCGAGTGGCAGCTGAGATGTTT-3′ | 5′-TGCTGGCAAAGTAGAAGAGGGCAA-3′ |
| Bcl-2 | 5′-ATCGCCCTGTGGATGACTGAG-3′ | 5′-CAGCCAGGAGAAATCAAACAGAGG-3′ |
| Casp3 | data | data |
| GAPDH | 5′-GGAGCGAGATCCCTCCAAAAT-3′ | 5′-GGCTGTTGTCATACTTCTCATGG-3′ |
| Ki67 | data | data |
| PDGF-A | 5′-CCCCTGCCCATTCGGAGGAAGAG-3′ | 5′-TTGGCCACCTTGACGCTGCGGTG-3′ |
| PDGF-B | 5′-GATCCGCTCCTTTGATGATC-3′ | 5′-GTCTCACACTTGCATGCCAG-3′ |
| RB1 | 5′-TTGGATCACAGCGATACAAACTT-3′ | 5′-AGCGCACGCCAATAAAGACAT-3′ |
| TP53 | 5′-GGAGTATTTGGATGACAGAAAC-3′ | 5′-GATTACCACTGGAGTCTTC-3′ |
| VEGF-A | 5′-AACCAGCAGAAAGAGGAAAGAGG-3′ | 5′-CCAAAAGCAGGTCACTCACTTTG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Felthaus, O.; Vedlin, S.; Eigenberger, A.; Klein, S.M.; Prantl, L. Exosomes from Adipose-Tissue-Derived Stem Cells Induce Proapoptotic Gene Expression in Breast Tumor Cell Line. Int. J. Mol. Sci. 2024, 25, 2190. https://doi.org/10.3390/ijms25042190
Felthaus O, Vedlin S, Eigenberger A, Klein SM, Prantl L. Exosomes from Adipose-Tissue-Derived Stem Cells Induce Proapoptotic Gene Expression in Breast Tumor Cell Line. International Journal of Molecular Sciences. 2024; 25(4):2190. https://doi.org/10.3390/ijms25042190
Chicago/Turabian StyleFelthaus, Oliver, Simon Vedlin, Andreas Eigenberger, Silvan M. Klein, and Lukas Prantl. 2024. "Exosomes from Adipose-Tissue-Derived Stem Cells Induce Proapoptotic Gene Expression in Breast Tumor Cell Line" International Journal of Molecular Sciences 25, no. 4: 2190. https://doi.org/10.3390/ijms25042190
APA StyleFelthaus, O., Vedlin, S., Eigenberger, A., Klein, S. M., & Prantl, L. (2024). Exosomes from Adipose-Tissue-Derived Stem Cells Induce Proapoptotic Gene Expression in Breast Tumor Cell Line. International Journal of Molecular Sciences, 25(4), 2190. https://doi.org/10.3390/ijms25042190