
Multi-Omics Approaches for Liver Reveal the Thromboprophylaxis Mechanism of Aspirin Eugenol Ester in Rat Thrombosis Model
 ,
,  and
and
Abstract
1. Introduction
2. Results
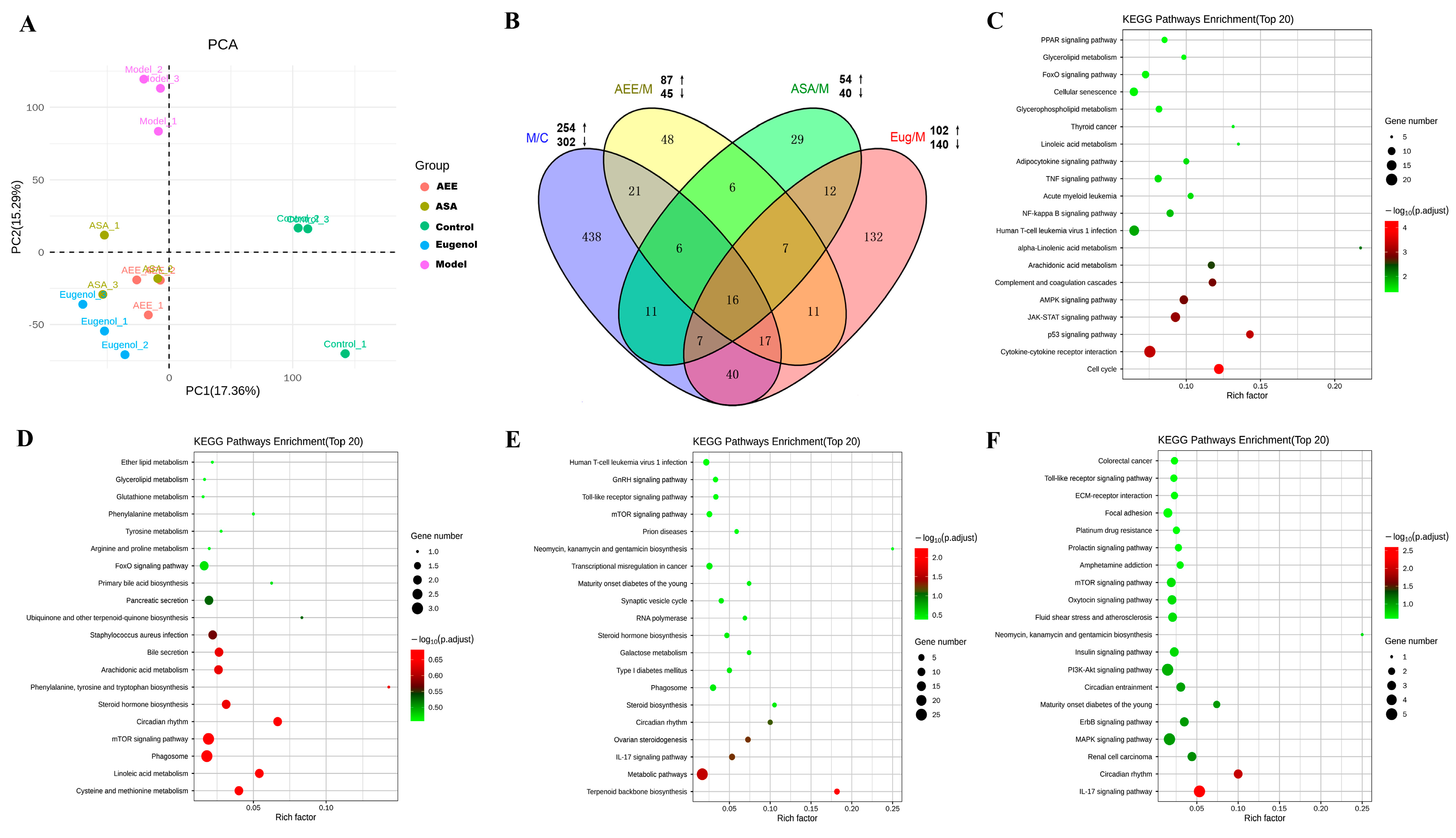
2.1. Transcriptome Sequencing and Gene Expression
2.1.1. Transcriptome Sequencing and Identification of DEGs
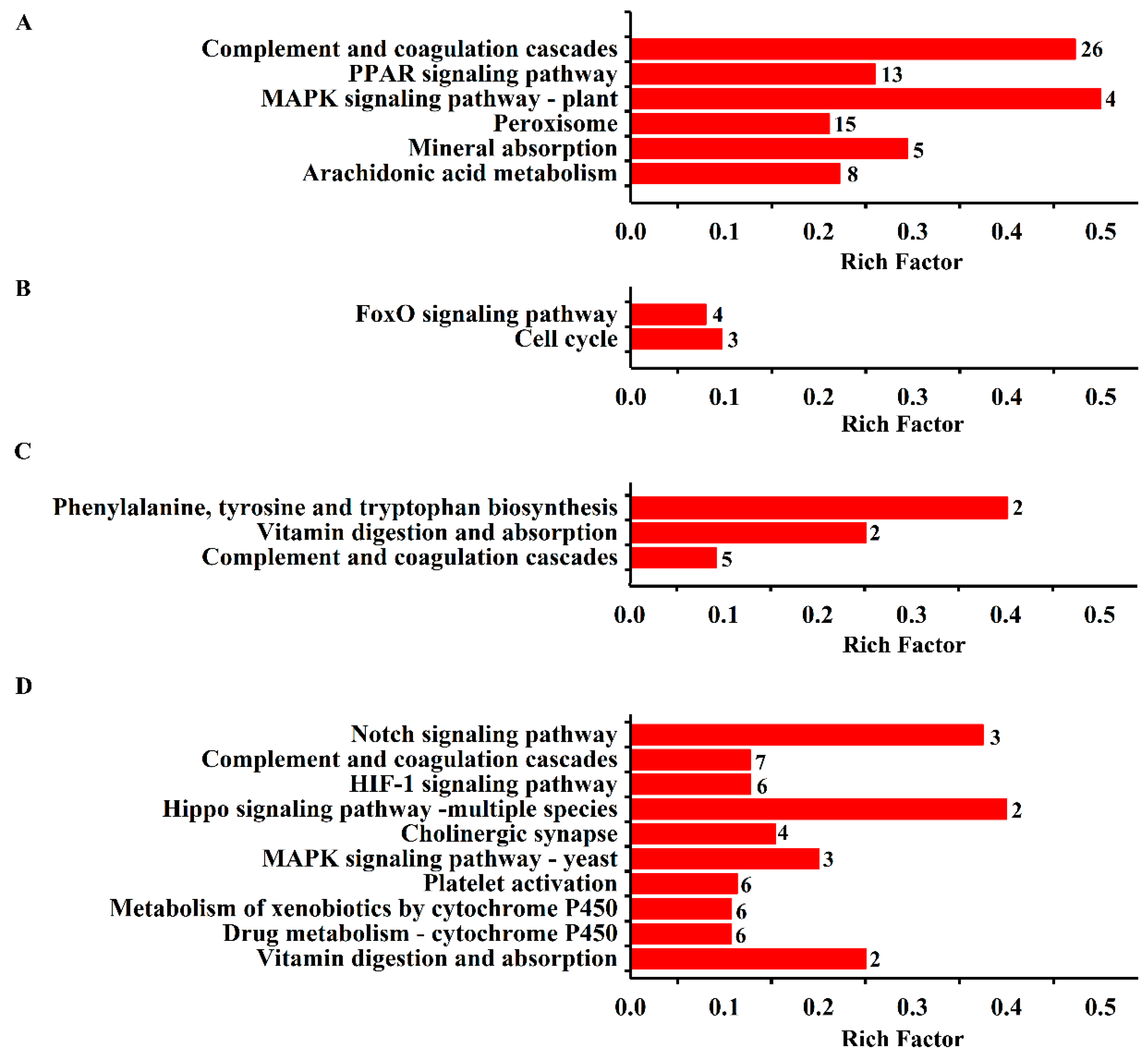
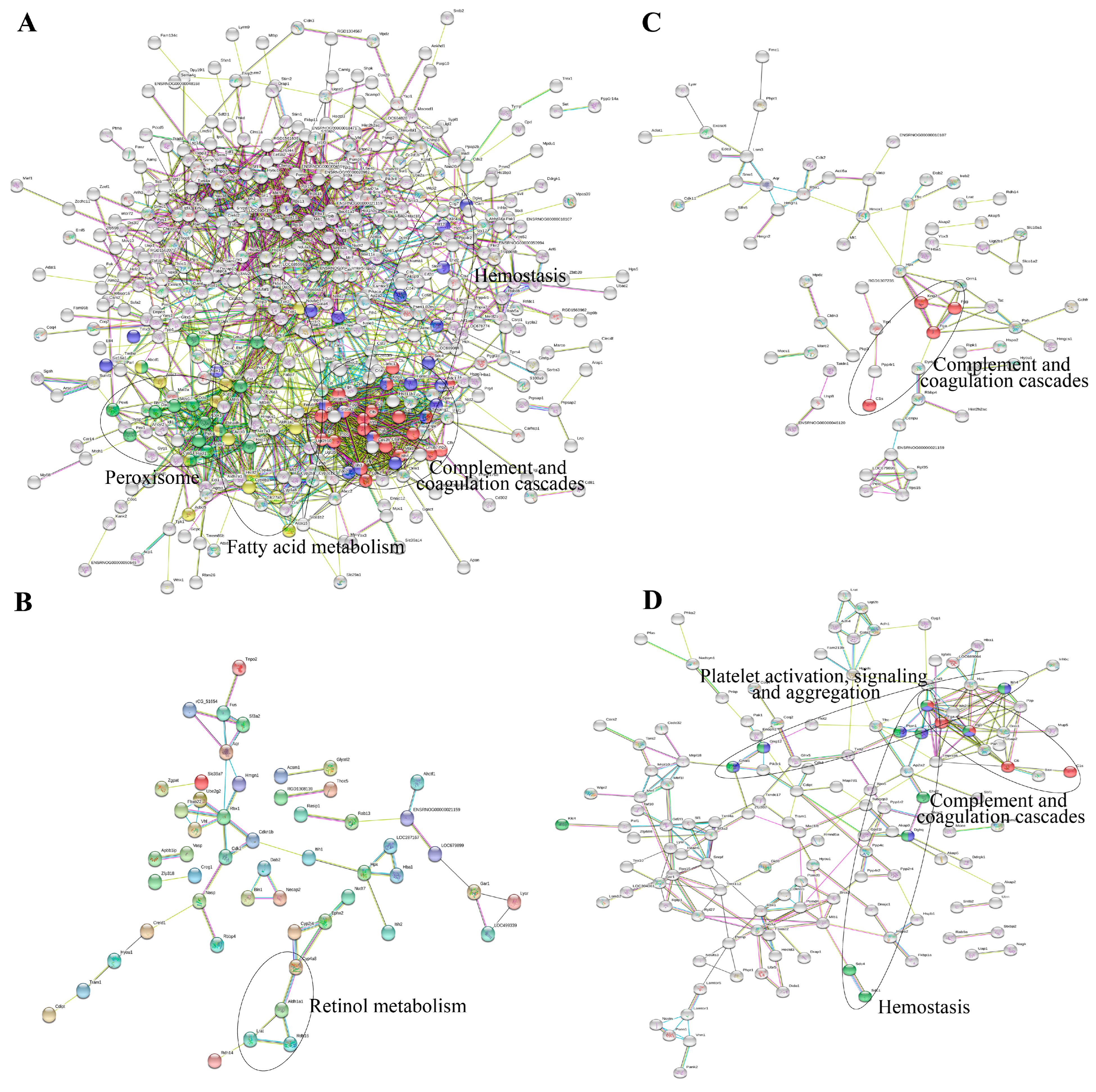
2.1.2. Functional Enrichment of DEGs
2.2. Proteomics Analysis Results
2.2.1. iTRAQ Proteomics Analysis of the Liver
2.2.2. Molecular Function and Pathway Analysis
2.2.3. PRM Validation
2.3. Metabonomics Analysis Results
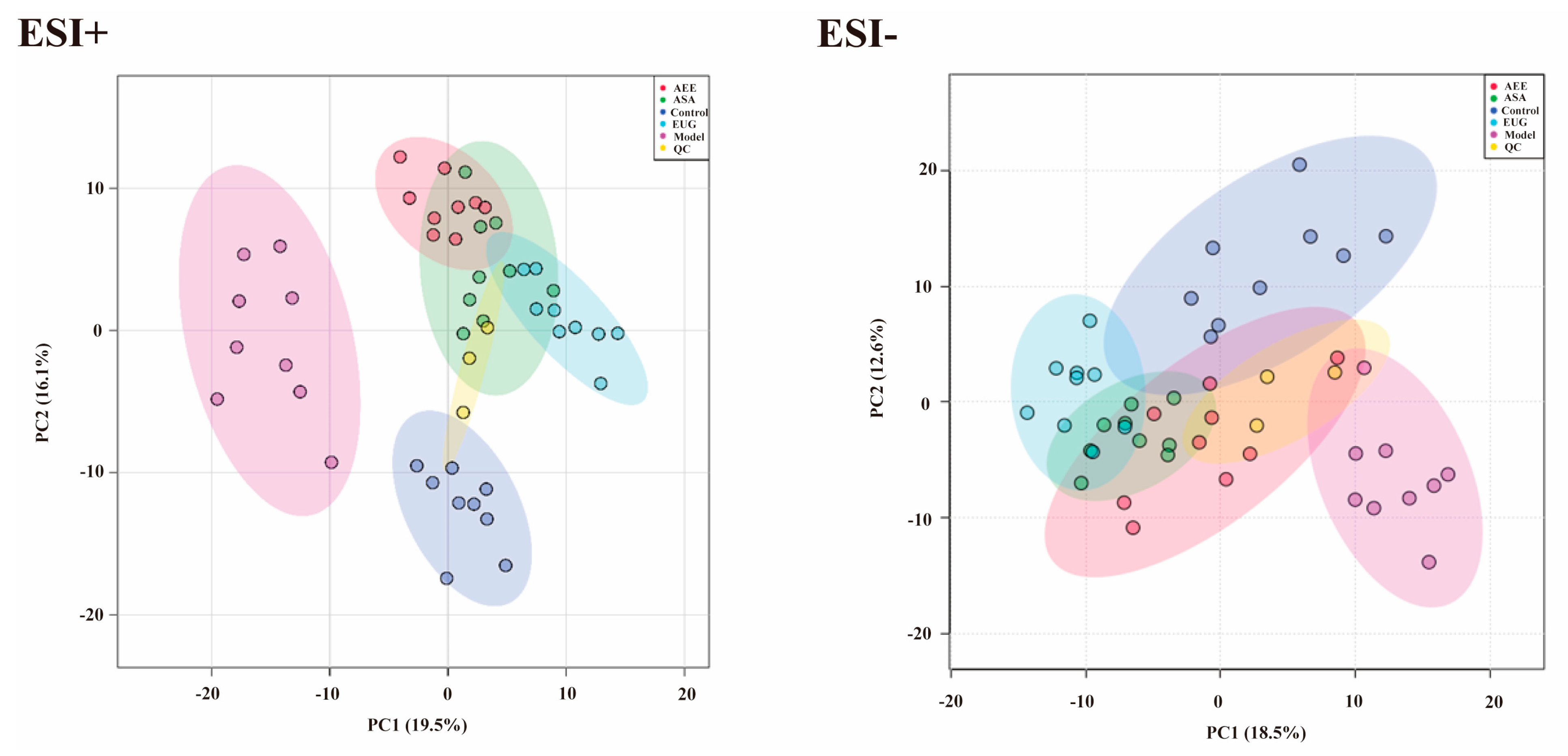
2.3.1. Multivariate Statistical Analysis and Metabolites Identification
2.3.2. Metabolic Pathway and Function Analysis
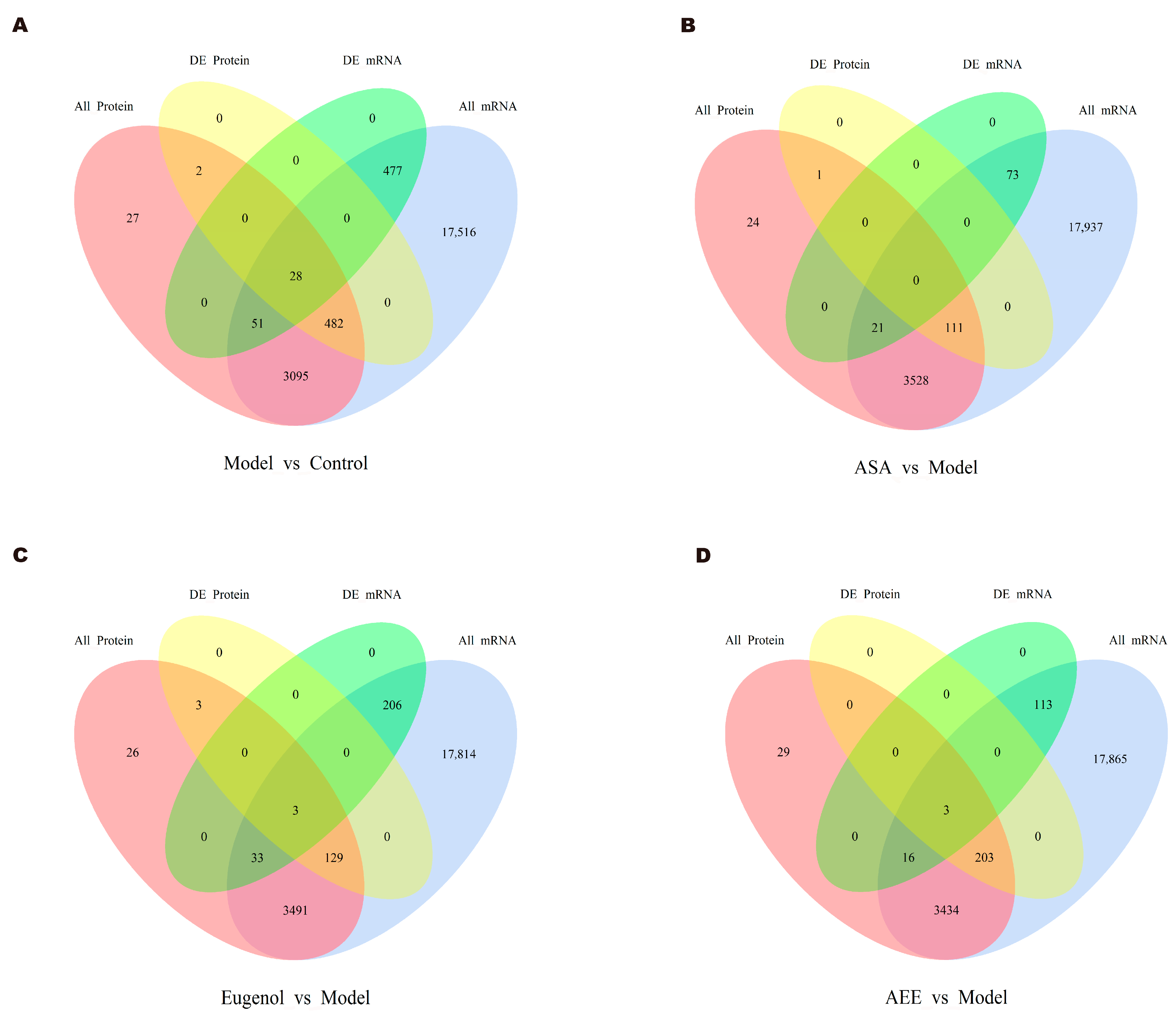
2.4. Integrative Analysis of Transcriptomics and Proteomics Data
3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. Animals and Sampling
4.3. Transcriptomics for RNA Isolation, Library Preparation, Sequencing, and Enrichment Analysis
4.4. ITRAQ Proteomics
4.4.1. Protein Exaction and Labeling
4.4.2. LC-MS/MS Analysis of Proteins
4.4.3. Protein Identification and Bioinformatics Analysis
4.4.4. Verification Analysis by Parallel Reaction Monitoring
4.5. Metabonomics Analysis
4.5.1. Metabonomics Analysis Platform
4.5.2. Multivariate Statistical Analysis
4.5.3. Identification and Pathway Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AEE | Aspirin eugenol ester. |
| ASA | Acetylsalicylic acid. |
| EUG | Eugenol. |
| DEGs | Differentially expressed genes. |
| DEPs | Differentially expressed proteins. |
| TCA | Tricarboxylic acid cycle. |
| CMC-Na | Carboxymethylcellulose. |
| NASID | Non-steroidal anti-inflammatory drug. |
| LC-MS | Liquid chromatography–mass spectrometry. |
| PCA | Principal component analysis. |
| OPLS-DA | Orthogonal partial-least squares discriminant analysis. |
| VIP | Variable importance for projection. |
| BP | Biological process. |
| PRM | Parallel reaction monitoring. |
References
- Stoll, G.; Nieswandt, B. Thrombo-inflammation in acute ischaemic stroke—Implications for treatment. Nat. Rev. Neurol. 2019, 15, 473–481. [Google Scholar] [CrossRef]
- Siscovick, D.S.; Schwartz, S.M.; Rosendaal, F.R.; Psaty, B.M. Thrombosis in the young: Effect of atherosclerotic risk factors on the risk of myocardial infarction associated with prothrombotic factors. Thromb. Haemost. 1997, 78, 7–12. [Google Scholar] [CrossRef]
- Otsuka, F.; Yasuda, S.; Noguchi, T.; Ishibashi-Ueda, H. Pathology of coronary atherosclerosis and thrombosis. Cardiovasc. Diagn. Ther. 2016, 6, 396–408. [Google Scholar] [CrossRef]
- Griesshammer, M.; Kiladjian, J.J.; Besses, C. Thromboembolic events in polycythemia vera. Ann. Hematol. 2019, 98, 1071–1082. [Google Scholar] [CrossRef] [PubMed]
- Fernández, A.G.; Salcedo, C.; Palacios, J.M. Aspirin, salicylate and gastrointestinal injury. Nat. Med. 1995, 1, 602–603. [Google Scholar] [CrossRef] [PubMed]
- Ribic, C.; Crowther, M. Thrombosis and anticoagulation in the setting of renal or liver disease. Hematol. Am. Soc. Hematol. Educ. Program 2016, 2016, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Handin, R.I. The History of Antithrombotic Therapy: The Discovery of Heparin, the Vitamin K Antagonists, and the Utility of Aspirin. Hematol./Oncol. Clin. N. Am. 2016, 30, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Schade, D.S.; Burchiel, S.; Eaton, R.P. A Pathophysiologic Primary Prevention Review of Aspirin Administration to Prevent Cardiovascular Thrombosis. Endocr. Pract. 2020, 26, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Lekstrom, J.A.; Bell, W.R. Aspirin in the prevention of thrombosis. Medicine 1991, 70, 161–178. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, D.P.; Militão, G.C.G.; de Morais, M.C.; de Sousa, D.P. The Dual Antioxidant/Prooxidant Effect of Eugenol and Its Action in Cancer Development and Treatment. Nutrients 2017, 9, 1367. [Google Scholar] [CrossRef]
- Mnafgui, K.; Hajji, R.; Derbali, F.; Gammoudi, A.; Khabbabi, G.; Ellefi, H.; Allouche, N.; Kadri, A.; Gharsallah, N. Anti-inflammatory, Antithrombotic and Cardiac Remodeling Preventive Effects of Eugenol in Isoproterenol-Induced Myocardial Infarction in Wistar Rat. Cardiovasc. Toxicol. 2016, 16, 336–344. [Google Scholar] [CrossRef]
- Yogalakshmi, B.; Viswanathan, P.; Anuradha, C.V. Investigation of antioxidant, anti-inflammatory and DNA-protective properties of eugenol in thioacetamide-induced liver injury in rats. Toxicology 2010, 268, 204–212. [Google Scholar] [CrossRef]
- Li, J.; Yu, Y.; Wang, Q.; Zhang, J.; Yang, Y.; Li, B.; Zhou, X.; Niu, J.; Wei, X.; Liu, X.; et al. Synthesis of aspirin eugenol ester and its biological activity. Med. Chem. Res. 2011, 21, 995–999. [Google Scholar] [CrossRef]
- Li, J.; Yu, Y.; Yang, Y.; Liu, X.; Zhang, J.; Li, B.; Zhou, X.; Niu, J.; Wei, X.; Liu, Z. A 15-day oral dose toxicity study of aspirin eugenol ester in Wistar rats. Food Chem. Toxicol. 2012, 50, 1980–1985. [Google Scholar] [CrossRef]
- Shen, Y.; Liu, X.; Yang, Y.; Li, J.; Ma, N.; Li, B. In vivo and in vitro metabolism of aspirin eugenol ester in dog by liquid chromatography tandem mass spectrometry. Biomed. Chromatogr. 2015, 29, 129–137. [Google Scholar] [CrossRef]
- Karczewski, K.J.; Snyder, M.P. Integrative omics for health and disease. Nat. Rev. Genet. 2018, 19, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J. Systems Biology of Metabolism. Annu. Rev. Biochem. 2017, 86, 245–275. [Google Scholar] [CrossRef] [PubMed]
- Arslan, R.; Bor, Z.; Bektas, N.; Meriçli, A.H.; Ozturk, Y. Antithrombotic effects of ethanol extract of Crataegus orientalis in the carrageenan-induced mice tail thrombosis model. Thromb. Res. 2011, 127, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Chen, Y.; Zhao, D.; Yang, S.; Zhang, S.; Wei, Z.; Wang, Y.; Qian, K.; Zhao, B.; Zhu, Y.; et al. LongShengZhi Capsule reduces carrageenan-induced thrombosis by reducing activation of platelets and endothelial cells. Pharmacol. Res. 2019, 144, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Shui, S.; Shen, S.; Huang, R.; Xiao, B.; Yang, J. Metabonomic analysis of biochemical changes in the plasma and urine of carrageenan-induced rats after treatment with Yi-Guan-Jian decoction. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1033–1034, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Liu, X.W.; Yang, Y.J.; Li, J.Y.; Mohamed, I.; Liu, G.R.; Zhang, J.Y. Preventive Effect of Aspirin Eugenol Ester on Thrombosis in κ-Carrageenan-Induced Rat Tail Thrombosis Model. PLoS ONE 2015, 10, e0133125. [Google Scholar] [CrossRef]
- Ma, N.; Liu, X.W.; Yang, Y.J.; Shen, D.S.; Zhao, X.L.; Mohamed, I.; Kong, X.J.; Li, J.Y. Evaluation on antithrombotic effect of aspirin eugenol ester from the view of platelet aggregation, hemorheology, TXB2/6-keto-PGF1α and blood biochemistry in rat model. BMC Vet. Res. 2016, 12, 108. [Google Scholar] [CrossRef]
- Ma, N.; Yang, Y.; Liu, X.; Li, S.; Qin, Z.; Li, J. Plasma metabonomics and proteomics studies on the anti-thrombosis mechanism of aspirin eugenol ester in rat tail thrombosis model. J. Proteom. 2020, 215, 103631. [Google Scholar] [CrossRef]
- Watt, M.J.; Miotto, P.M.; De Nardo, W.; Montgomery, M.K. The Liver as an Endocrine Organ-Linking NAFLD and Insulin Resistance. Endocr. Rev. 2019, 40, 1367–1393. [Google Scholar] [CrossRef]
- Foley, J.H. Examining coagulation-complement crosstalk: Complement activation and thrombosis. Thromb. Res. 2016, 141 (Suppl. 2), S50–S54. [Google Scholar] [CrossRef]
- Sztolsztener, K.; Chabowski, A.; Harasim-Symbor, E.; Bielawiec, P.; Konstantynowicz-Nowicka, K. Arachidonic Acid as an Early Indicator of Inflammation during Non-Alcoholic Fatty Liver Disease Development. Biomolecules 2020, 10, 1133. [Google Scholar] [CrossRef]
- Yang, Q.; Cao, W.; Zhou, X.; Cao, W.; Xie, Y.; Wang, S. Anti-thrombotic effects of α-linolenic acid isolated from Zanthoxylum bungeanum Maxim seeds. BMC Complement. Altern. Med. 2014, 14, 348. [Google Scholar] [CrossRef] [PubMed]
- Spector, A.A.; Kim, H.Y. Discovery of essential fatty acids. J. Lipid Res. 2015, 56, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Nagamine, R.; Tanaka, Y.; Tsai, W.T.; Jiang, Z.; Takeyama, A.; Imaizumi, K.; Sato, M. The effects of dietary linoleic acid on reducing serum cholesterol and atherosclerosis development are nullified by a high-cholesterol diet in male and female apoE-deficient mice. Br. J. Nutr. 2023, 129, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Azemi, N.A.; Azemi, A.K.; Abu-Bakar, L.; Sevakumaran, V.; Muhammad, T.S.T.; Ismail, N. Effect of Linoleic Acid on Cholesterol Levels in a High-Fat Diet-Induced Hypercholesterolemia Rat Model. Metabolites 2022, 13, 53. [Google Scholar] [CrossRef] [PubMed]
- Shui, S.; Cai, X.; Huang, R.; Xiao, B.; Yang, J. The investigation of anti-inflammatory activity of Yi Guanjian decoction by serum metabonomics approach. J. Pharm. Biomed. Anal. 2017, 133, 41–48. [Google Scholar] [CrossRef]
- Manganelli, V.; Longo, A.; Mattei, V.; Recalchi, S.; Riitano, G.; Caissutti, D.; Capozzi, A.; Sorice, M.; Misasi, R.; Garofalo, T. Role of ERLINs in the Control of Cell Fate through Lipid Rafts. Cells 2021, 10, 2408. [Google Scholar] [CrossRef]
- Cronin, A.; Mowbray, S.; Dürk, H.; Homburg, S.; Fleming, I.; Fisslthaler, B.; Oesch, F.; Arand, M. The N-terminal domain of mammalian soluble epoxide hydrolase is a phosphatase. Proc. Natl. Acad. Sci. USA 2003, 100, 1552–1557. [Google Scholar] [CrossRef]
- Lin, H.L.; Yen, H.W.; Hsieh, S.L.; An, L.M.; Shen, K.P. Low-dose aspirin ameliorated hyperlipidemia, adhesion molecule, and chemokine production induced by high-fat diet in Sprague-Dawley rats. Drug Dev. Res. 2014, 75, 97–106. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, M.; Zhou, T.; Shen, Q.; Qin, X. Exendin-4 promotes the vascular smooth muscle cell re-differentiation through AMPK/SIRT1/FOXO3a signaling pathways. Atherosclerosis 2018, 276, 58–66. [Google Scholar] [CrossRef]
- Lotfimehr, H.; Mardi, N.; Narimani, S.; Nasrabadi, H.T.; Karimipour, M.; Sokullu, E.; Rahbarghazi, R. mTOR signalling pathway in stem cell bioactivities and angiogenesis potential. Cell Prolif. 2023, 56, e13499. [Google Scholar] [CrossRef] [PubMed]
- Kordjazy, N.; Haj-Mirzaian, A.; Haj-Mirzaian, A.; Rohani, M.M.; Gelfand, E.W.; Rezaei, N.; Abdolghaffari, A.H. Role of toll-like receptors in inflammatory bowel disease. Pharmacol. Res. 2018, 129, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Luo, T.; Yuan, D.; Liu, J.; Fu, Y.; Yuan, J. Qilongtian ameliorate bleomycin-induced pulmonary fibrosis in mice via inhibiting IL-17 signal pathway. Sci. Rep. 2023, 13, 6002. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Yang, B.; Zhao, M.; Li, P.; Fan, J.; Chang, M.; Pan, Z.; Zhang, Z.; Xue, S.; Zhang, Y. Effects of modified Huangqi Chifeng decoction on the IL-17 signaling pathway in an IgA nephropathy rat model. J. Ethnopharmacol. 2023, 307, 116220. [Google Scholar] [CrossRef]
- Shi, Q.; Lang, W.; Wang, S.; Li, G.; Bai, X.; Yan, X.; Zhang, H. Echinacea polysaccharide attenuates lipopolysaccharide-induced acute kidney injury via inhibiting inflammation, oxidative stress and the MAPK signaling pathway. Int. J. Mol. Med. 2021, 47, 243–255. [Google Scholar] [CrossRef]
- Cao, X.; Fu, M.; Bi, R.; Zheng, X.; Fu, B.; Tian, S.; Liu, C.; Li, Q.; Liu, J. Cadmium induced BEAS-2B cells apoptosis and mitochondria damage via MAPK signaling pathway. Chemosphere 2021, 263, 128346. [Google Scholar] [CrossRef]
- Zang, P.; Yang, C.; Lei, H.; Guo, Q.; Wang, W.; Shao, J. Ghrelin regulates the proliferation and apoptosis of high glucose-induced islet cells through the PI3K-Akt signaling pathway. Cell Biol. Int. 2023, 47, 768–775. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.R.; Gallant, R.C.; Ni, H. Platelets, immune-mediated thrombocytopenias, and fetal hemorrhage. Thromb. Res. 2016, 141 (Suppl. 2), S76–S79. [Google Scholar] [CrossRef] [PubMed]
- Matsusaka, K.; Fujiwara, Y.; Pan, C.; Esumi, S.; Saito, Y.; Bi, J.; Nakamura, Y.; Mukunoki, A.; Takeo, T.; Nakagata, N.; et al. α(1)-Acid Glycoprotein Enhances the Immunosuppressive and Protumor Functions of Tumor-Associated Macrophages. Cancer Res. 2021, 81, 4545–4559. [Google Scholar] [CrossRef] [PubMed]
- Schaer, D.J.; Vinchi, F.; Ingoglia, G.; Tolosano, E.; Buehler, P.W. Haptoglobin, hemopexin, and related defense pathways-basic science, clinical perspectives, and drug development. Front. Physiol. 2014, 5, 415. [Google Scholar] [CrossRef]
- Chavakis, T.; Preissner, K.T. Potential pharmacological applications of the antithrombotic molecule high molecular weight kininogen. Curr. Vasc. Pharmacol. 2003, 1, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Ammar, N.M.; Hassan, H.A.; Abdallah, H.M.I.; Afifi, S.M.; Elgamal, A.M.; Farrag, A.R.H.; El-Gendy, A.E.G.; Farag, M.A.; Elshamy, A.I. Protective Effects of Naringenin from Citrus sinensis (var. Valencia) Peels against CCl(4)-Induced Hepatic and Renal Injuries in Rats Assessed by Metabolomics, Histological and Biochemical Analyses. Nutrients 2022, 14, 841. [Google Scholar] [CrossRef] [PubMed]
- Martínez, Y.; Li, X.; Liu, G.; Bin, P.; Yan, W.; Más, D.; Valdivié, M.; Hu, C.A.; Ren, W.; Yin, Y. The role of methionine on metabolism, oxidative stress, and diseases. Amino Acids 2017, 49, 2091–2098. [Google Scholar] [CrossRef]
- Weaver, Z.; Difilippantonio, S.; Carretero, J.; Martin, P.L.; El Meskini, R.; Iacovelli, A.J.; Gumprecht, M.; Kulaga, A.; Guerin, T.; Schlomer, J.; et al. Temporal molecular and biological assessment of an erlotinib-resistant lung adenocarcinoma model reveals markers of tumor progression and treatment response. Cancer Res. 2012, 72, 5921–5933. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Map ID | Pathway Name | Proteins Accession ID | Count | RF | p-Value |
|---|---|---|---|---|---|
| Control vs. Model | |||||
| map04610 | Complement and coagulation cascades | P08932 P19999 Q3KR94 D4A1S0 Q62930 Q5PQU1 P06399 P02680 A0A096P6L9 P14480 Q6MG74 Q5M7T5 D3ZWD6 A0A0G2JY31 G3V836 Q9EQV9 F1M983 Q6MG73 Q6P734 B5DEH7 P08649 P16296 F1M7F7 M0R5R0 G3V843 P31394 | 26 | 0.47 | 0.0000 |
| map03320 | PPAR signaling pathway | P20817 P07379 P55051 G3V8J2 Q9ES38 P24464 P07896 P07308 F1LNW3 F1LQC1 P55053 D3ZJX6 A0A0G2K8Q1 | 13 | 0.26 | 0.0022 |
| map04016 | MAPK signaling pathway—plant | P19804 P62161 P04762 G3V816 | 4 | 0.50 | 0.0070 |
| map04146 | Peroxisome | P07895 P56574 F1LNG8 Q8CHM7 P04762 B0BNF9 P07896 F1LNW3 F1LQC1 D3ZDM7 D3ZZB2 Q5I0K1 P54777 D3ZHR2 O35078 | 15 | 0.21 | 0.0089 |
| map04978 | Mineral absorption | Q9WUC4 Q7M082 D3ZHV3 P12346 Q66HI5 | 5 | 0.29 | 0.0316 |
| map00590 | Arachidonic acid metabolism | O35543 P20817 P24464 P11510 Q5EB99 D3ZJX6 Q02759 B2GV28 | 8 | 0.22 | 0.0379 |
| AEE vs. Model | |||||
| map04330 | Notch signaling pathway | P97887 B3DM90 Q2LC86 | 3 | 0.38 | 0.0041 |
| map04610 | Complement and coagulation cascades | F1M7F7 Q5PQU1 P06399 P02680 A0A0G2JY31 P14480 D4A1S0 | 7 | 0.13 | 0.0102 |
| map04723 | Retrograde endocannabinoid signaling | G3V6P8 Q45QM4 Q45QM8 Q5XI64 | 4 | 0.19 | 0.0125 |
| map04066 | HIF-1 signaling pathway | Q498D8 O08769 P12346 G3V679 E9PTN6 M0RC47 | 6 | 0.13 | 0.0168 |
| map04392 | Hippo signaling pathway—multiple species | P35465 R9PXS9 | 2 | 0.40 | 0.0179 |
| map04725 | Cholinergic synapse | G3V6P8 Q45QM4 Q45QM8 M0RC47 | 4 | 0.15 | 0.0263 |
| map04011 | MAPK signaling pathway—yeast | P35465 D3ZAP9 Q4V886 | 3 | 0.20 | 0.0265 |
| map04611 | Platelet activation | Q45QM4 Q45QM8 M0RC47 P06399 P02680 P14480 | 6 | 0.11 | 0.0289 |
| map00980 | Metabolism of xenobiotics by CYP450 | A1L128 P04903 O35543 P06757 A9EEP5 F1LM22 | 6 | 0.11 | 0.0366 |
| map00982 | Drug metabolism—CYP450 | A1L128 P04903 O35543 P06757 A9EEP5 F1LM22 | 6 | 0.11 | 0.0366 |
| map04977 | Vitamin digestion and absorption | Q5FVF9 Q9JI61 | 2 | 0.25 | 0.0461 |
| ASA vs. Model | |||||
| map04068 | FoxO signaling pathway | Q811R1 O08769 Q63699 Q9Z2X5 | 4 | 0.08 | 0.0306 |
| map04110 | Cell cycle | Q498D8 O08769 Q63699 | 3 | 0.10 | 0.0367 |
| Eug vs. Model | |||||
| map00400 | Phenylalanine, tyrosine, and tryptophan biosynthesis | P04176 P04694 | 2 | 0.40 | 0.0080 |
| map04977 | Vitamin digestion and absorption | Q5FVF9 Q9JI61 | 2 | 0.25 | 0.0211 |
| map04610 | Complement and coagulation cascades | F7FEU1 D4A1S0 Q5PQU1 P06399 P02680 | 5 | 0.09 | 0.0214 |
| Protein ID | Gene | Description | M/C | ASA/M | Eug/M | AEE/M | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| iTRAQ | PRM | iTRAQ | PRM | iTRAQ | PRM | iTRAQ | PRM | |||
| P06399 | Fga | Fibrinogen alpha chain | 1.71 ** | 1.17 | 0.90 | 0.96 | 0.81 * | 0.47 * | 0.77 * | 0.81 |
| P02680 | Fgg | Fibrinogen gamma chain | 1.69 ** | 1.33 | 0.88 | 0.85 | 0.82 * | 0.43 ** | 0.76 * | 0.79 |
| P14480 | Fgb | Fibrinogen beta chain | 1.58 ** | 1.33 | 0.90 | 0.83 | 0.86 | 0.37 ** | 0.81 * | 0.76 |
| P02764 | Orm1 | Alpha-1-acid glycoprotein | 2.52 ** | 3.84 ** | 0.96 | 1.16 | 0.81 * | 0.80 | 0.67 ** | 0.70 |
| P20059 | Hpx | Hemopexin | 1.62 ** | 1.32 | 0.83 * | 0.94 | 0.82 * | 0.79 | 0.80 * | 0.80 |
| Q5PQU1 | Kng2 | Kininogen 2 | 2.08 ** | 5.72 ** | 1.00 | 1.11 | 0.81 ** | 0.68 | 0.83 * | 0.82 |
| No. | Metabolites | Formula | Ion Mode | m/z | RT (min) | Fold Change | |||
|---|---|---|---|---|---|---|---|---|---|
| Mod/C | ASA/Mod | Eug/Mod | AEE/Mod | ||||||
| 1 | Cinnamic acid | C9H8O2 | ESI− | 147.0465 | 5.36 | 0.56 ** | 1.30 ** | 1.42 ** | 1.13 |
| 2 | Xanthine | C5H4N4O2 | ESI− | 151.0275 | 2.39 | 0.64 ** | 1.47 * | 1.40 | 1.26 * |
| 3 | Cholic acid | C24H40O5 | ESI− | 407.2843 | 14.47 | 0.37 ** | 0.98 | 1.47 | 2.18 |
| 4 | Palmitic acid | C16H32O2 | ESI− | 255.2352 | 23.64 | 0.41 ** | 2.51 ** | 1.91 ** | 2.00 ** |
| 5 | Malic acid | C4H6O5 | ESI− | 133.0153 | 2.01 | 0.39 ** | 3.34 ** | 1.46 | 3.89 ** |
| 6 | Geranylgeranylcysteine | C23H37NO3S | ESI− | 406.2457 | 21.05 | 1.38 | 0.58 * | 1.10 | 0.42 ** |
| 7 | Thymine | C5H6N2O2 | ESI− | 125.0366 | 3.94 | 0.44 ** | 1.83 ** | 2.36 ** | 1.39 ** |
| 8 | Tetracosahexaenoic acid | C24H36O2 | ESI− | 355.2675 | 22.99 | 0.41 ** | 1.63 ** | 1.92 ** | 1.33 * |
| 9 | Leucine | C6H13NO2· | ESI− | 130.0885 | 2.86 | 0.57 ** | 1.59 ** | 1.61 * | 1.39 * |
| 10 | Aspartic acid | C4H7NO4 | ESI− | 132.0315 | 1.35 | 0.42 ** | 2.20 ** | 2.61 | 1.56 ** |
| 11 | Stearic acid | C18H36O2 | ESI− | 283.2668 | 25.55 | 0.26 ** | 3.93 ** | 5.24 ** | 4.88 ** |
| 12 | Succinic acid | C4H6O4 | ESI− | 117.0202 | 2.62 | 0.51 ** | 2.08 ** | 2.10 ** | 1.71 ** |
| 13 | Linoleic acid | C18H32O2 | ESI− | 279.2355 | 22.55 | 0.63 ** | 1.90 ** | 1.82 ** | 1.51 * |
| 14 | Uracil | C4H4N2O2 | ESI− | 111.0210 | 2.18 | 0.71 ** | 2.62 ** | 2.82 ** | 1.95 ** |
| 15 | Ornithine | C5H12N2O2 | ESI− | 131.0837 | 1.27 | 0.69 ** | 1.54 ** | 1.68 ** | 1.38 ** |
| 16 | Adrenic acid | C22H36O2 | ESI− | 331.2672 | 23.52 | 0.70 ** | 1.61 ** | 1.69 ** | 1.25 * |
| 17 | Hypoxanthine | C5H4N4O | ESI− | 135.0322 | 2.24 | 0.75 * | 1.61 * | 2.35 ** | 1.35 * |
| 18 | Histidine | C6H9N3O2 | ESI− | 154.0635 | 1.35 | 0.57 ** | 1.57 ** | 2.02 ** | 1.64 ** |
| 19 | lysine | C6H14N2O2 | ESI− | 145.0995 | 1.28 | 0.68 ** | 1.49 ** | 1.62 ** | 1.32 ** |
| 20 | Phenylalanine | C9H11NO2 | ESI− | 164.0733 | 5.36 | 0.71 ** | 1.47 ** | 1.64 ** | 1.30 ** |
| 21 | Oxidized glutathione | C20H32N6O12S2 | ESI− | 611.1507 | 2.75 | 2.29 * | 1.35 | 2.30 * | 0.45 * |
| 22 | Proline | C5H9NO2 | ESI+ | 116.0709 | 1.51 | 0.67 ** | 1.33 ** | 1.72 ** | 1.19 ** |
| 23 | Valine | C5H11NO2 | ESI+ | 118.0864 | 2.14 | 0.67 ** | 1.52 ** | 1.81 ** | 1.28 ** |
| 24 | Niacinamide | C6H6N2O | ESI+ | 123.0552 | 2.22 | 0.66 ** | 1.35 ** | 1.55 ** | 1.15 ** |
| 25 | Pipecolic acid | C6H11NO2 | ESI+ | 130.0864 | 1.27 | 0.60 ** | 1.08 | 1.39 ** | 1.11 * |
| 26 | Isoleucine | C6H13NO2 | ESI+ | 132.1018 | 2.45 | 0.53 ** | 1.62 * | 1.84 * | 1.28 |
| 27 | Urocanic acid | C6H6N2O2 | ESI+ | 139.0499 | 4.14 | 0.49 | 4.33 ** | 6.60 ** | 2.03 |
| 28 | Spermidine | C7H19N3 | ESI+ | 146.1649 | 1.20 | 0.76 ** | 1.57 ** | 1.59 ** | 1.42 ** |
| 29 | Glutamic acid | C5H9NO4 | ESI+ | 148.0606 | 1.45 | 0.76 ** | 1.32 ** | 1.50 ** | 1.17 |
| 30 | Methionine | C5H11NO2S | ESI+ | 150.0583 | 1.68 | 0.69 ** | 1.29 ** | 1.66 ** | 1.13 * |
| 31 | Phenylpyruvic acid | C9H8O3 | ESI+ | 165.0543 | 1.87 | 0.57 ** | 1.19 | 1.13 | 1.31 ** |
| 32 | Methylphenylalanine | C10H13NO2 | ESI+ | 180.1015 | 7.19 | 0.75 ** | 1.83 ** | 2.62 ** | 1.84 ** |
| 33 | Leucylproline | C11H20N2O3 | ESI+ | 229.1545 | 6.48 | 0.47 ** | 1.28 | 1.75 ** | 1.82 ** |
| 34 | Palmitic amide | C16H33NO | ESI+ | 256.2634 | 24.32 | 1.29 | 0.78 | 0.67 ** | 0.54 ** |
| 35 | Octadecylamine | C18H39N | ESI+ | 270.3158 | 17.57 | 1.25 | 0.57 ** | 0.81 | 0.53 ** |
| 36 | Linolenic acid | C18H30O2 | ESI+ | 279.2321 | 21.35 | 1.19 | 0.65 ** | 0.78 * | 0.60 ** |
| 37 | Linoleamide | C18H33NO | ESI+ | 280.2640 | 20.87 | 1.69 ** | 0.65 ** | 0.66 ** | 0.64 ** |
| 38 | Oleamide | C18H35NO | ESI+ | 282.2799 | 15.37 | 0.67 ** | 0.97 | 0.95 | 1.15 * |
| 39 | Sphingosine | C18H37NO2 | ESI+ | 300.2903 | 21.07 | 1.80 ** | 0.44 ** | 0.49 ** | 0.53 ** |
| 40 | Sphinganine | C18H39NO2 | ESI+ | 302.3061 | 14.95 | 2.06 ** | 0.54 * | 0.50 ** | 1.02 |
| 41 | Arachidonic acid | C20H32O2 | ESI+ | 305.2479 | 22.16 | 1.34 | 0.60 | 0.62 ** | 0.57 ** |
| 42 | Linolenoyl ethanolamide | C20H37NO2 | ESI+ | 324.2907 | 20.06 | 1.59 ** | 0.59 ** | 0.71 * | 0.61 ** |
| 43 | Oleoylethanolamide | C20H39NO2 | ESI+ | 326.3062 | 21.62 | 1.70 ** | 0.39 ** | 0.44 ** | 0.44 ** |
| 44 | Stearoylglycine | C20H39NO3 | ESI+ | 342.3009 | 23.09 | 2.32 ** | 0.51 ** | 0.47 ** | 0.62 ** |
| 45 | DGLEA | C22H39NO2 | ESI+ | 350.3067 | 20.84 | 1.56 * | 0.43 ** | 0.51 ** | 0.53 ** |
| 46 | Taurodeoxycholic acid | C26H45NO6S | ESI+ | 500.3052 | 13.72 | 2.39 ** | 0.75 | 0.42 ** | 0.75 |
| 47 | N-Palmitoyl phenylalanine | C25H41NO3 | ESI+ | 404.3140 | 23.02 | 0.30 * | 4.27 ** | 3.70 * | 5.07 ** |
| 48 | LysoPC (O-18:0/0:0) | C26H56NO6P | ESI+ | 510.3935 | 21.12 | 1.79 ** | 0.84 | 0.91 | 0.61* |
| 49 | LysoPC (18:3) | C26H48NO7P | ESI+ | 518.3233 | 16.59 | 0.46 ** | 1.89 ** | 1.93 ** | 2.19 ** |
| 50 | LysoPC (18:0/0:0) | C26H54NO7P | ESI+ | 524.3726 | 21.10 | 2.43 ** | 0.59 ** | 0.52 ** | 0.76 |
| 51 | LysoPC (20:5) | C28H48NO7P | ESI+ | 542.3234 | 16.94 | 0.34 ** | 2.25 ** | 2.19 ** | 3.12 ** |
| 52 | LysoPC (20:4) | C28H50NO7P | ESI+ | 544.3394 | 18.15 | 0.23 ** | 2.61 * | 2.20 ** | 3.60 ** |
| 53 | LysoPC (20:3) | C28H52NO7P | ESI+ | 546.3572 | 20.01 | 0.29 * | 5.79 ** | 5.44 ** | 4.81 ** |
| 54 | LysoPC (20:0/0:0) | C28H58NO7P | ESI+ | 552.4033 | 20.70 | 2.41 ** | 0.40 ** | 0.54 ** | 0.44 ** |
| 55 | LysoPC (22:4) | C30H54NO7P | ESI+ | 572.3703 | 20.44 | 0.51 | 1.64 | 1.74 | 3.70 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, Q.; Ma, N.; Fan, L.; Ge, W.; Zhang, Z.; Liu, X.; Li, J.; Yang, Y. Multi-Omics Approaches for Liver Reveal the Thromboprophylaxis Mechanism of Aspirin Eugenol Ester in Rat Thrombosis Model. Int. J. Mol. Sci. 2024, 25, 2141. https://doi.org/10.3390/ijms25042141
Tao Q, Ma N, Fan L, Ge W, Zhang Z, Liu X, Li J, Yang Y. Multi-Omics Approaches for Liver Reveal the Thromboprophylaxis Mechanism of Aspirin Eugenol Ester in Rat Thrombosis Model. International Journal of Molecular Sciences. 2024; 25(4):2141. https://doi.org/10.3390/ijms25042141
Chicago/Turabian StyleTao, Qi, Ning Ma, Liping Fan, Wenbo Ge, Zhendong Zhang, Xiwang Liu, Jianyong Li, and Yajun Yang. 2024. "Multi-Omics Approaches for Liver Reveal the Thromboprophylaxis Mechanism of Aspirin Eugenol Ester in Rat Thrombosis Model" International Journal of Molecular Sciences 25, no. 4: 2141. https://doi.org/10.3390/ijms25042141
APA StyleTao, Q., Ma, N., Fan, L., Ge, W., Zhang, Z., Liu, X., Li, J., & Yang, Y. (2024). Multi-Omics Approaches for Liver Reveal the Thromboprophylaxis Mechanism of Aspirin Eugenol Ester in Rat Thrombosis Model. International Journal of Molecular Sciences, 25(4), 2141. https://doi.org/10.3390/ijms25042141