
Polysaccharide from Atractylodes macrocephala Koidz Binding with Zinc Oxide Nanoparticles as a Novel Mucosal Immune Adjuvant for H9N2 Inactivated Vaccine


 ,
,
Abstract
1. Introduction
2. Results
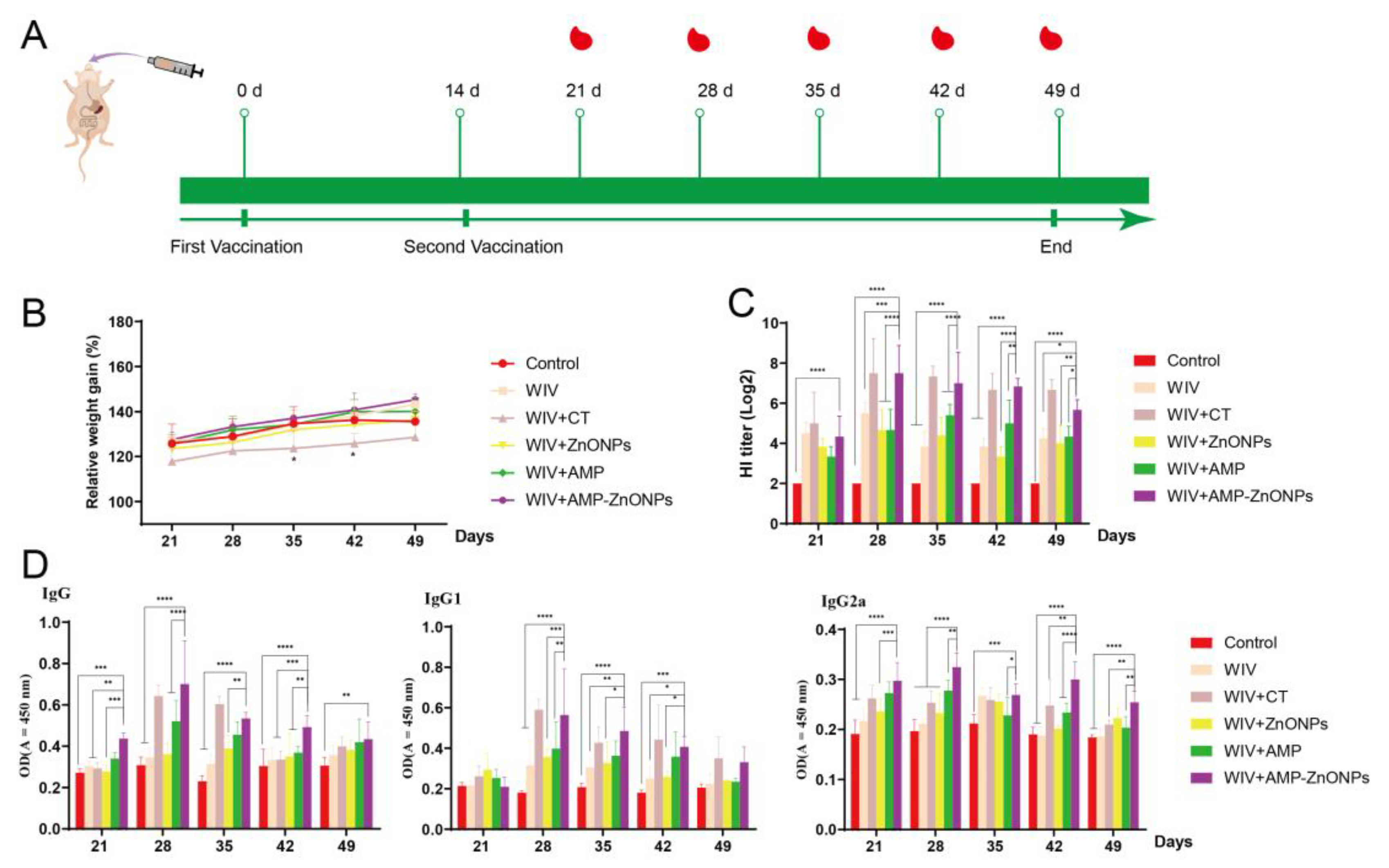
2.1. Specific HI, IgG, IgG1 and IgG2a Antibody Levels
2.2. Differentiation between CD4+ and CD8+ T Cells
2.3. The mRNA Expression Levels of Th1 and Th2 Cytokine
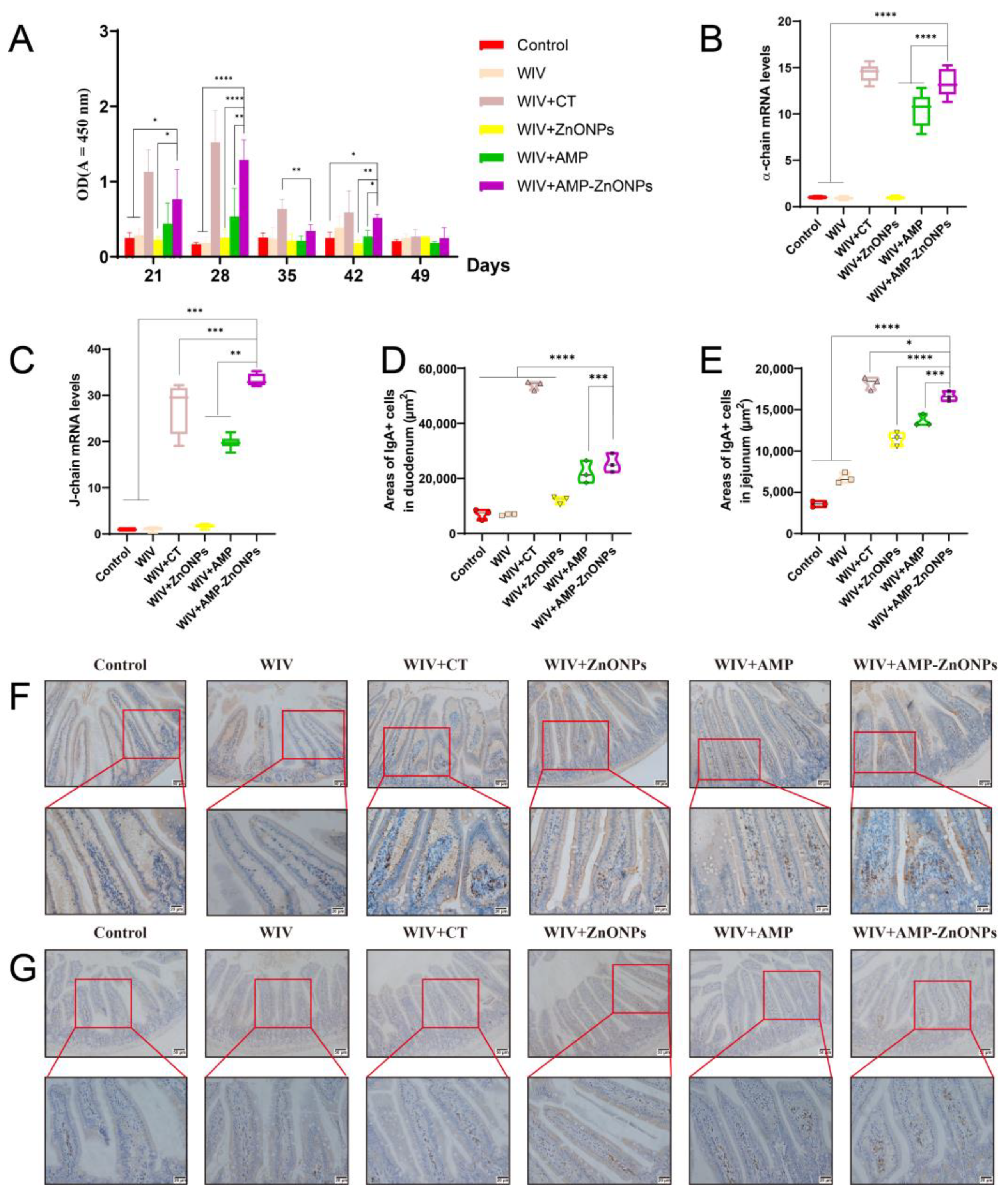
2.4. The sIgA Level at Mucosal Sites
2.5. The Change in IgA-Secreting Cell Areas in the Intestinal Mucosa
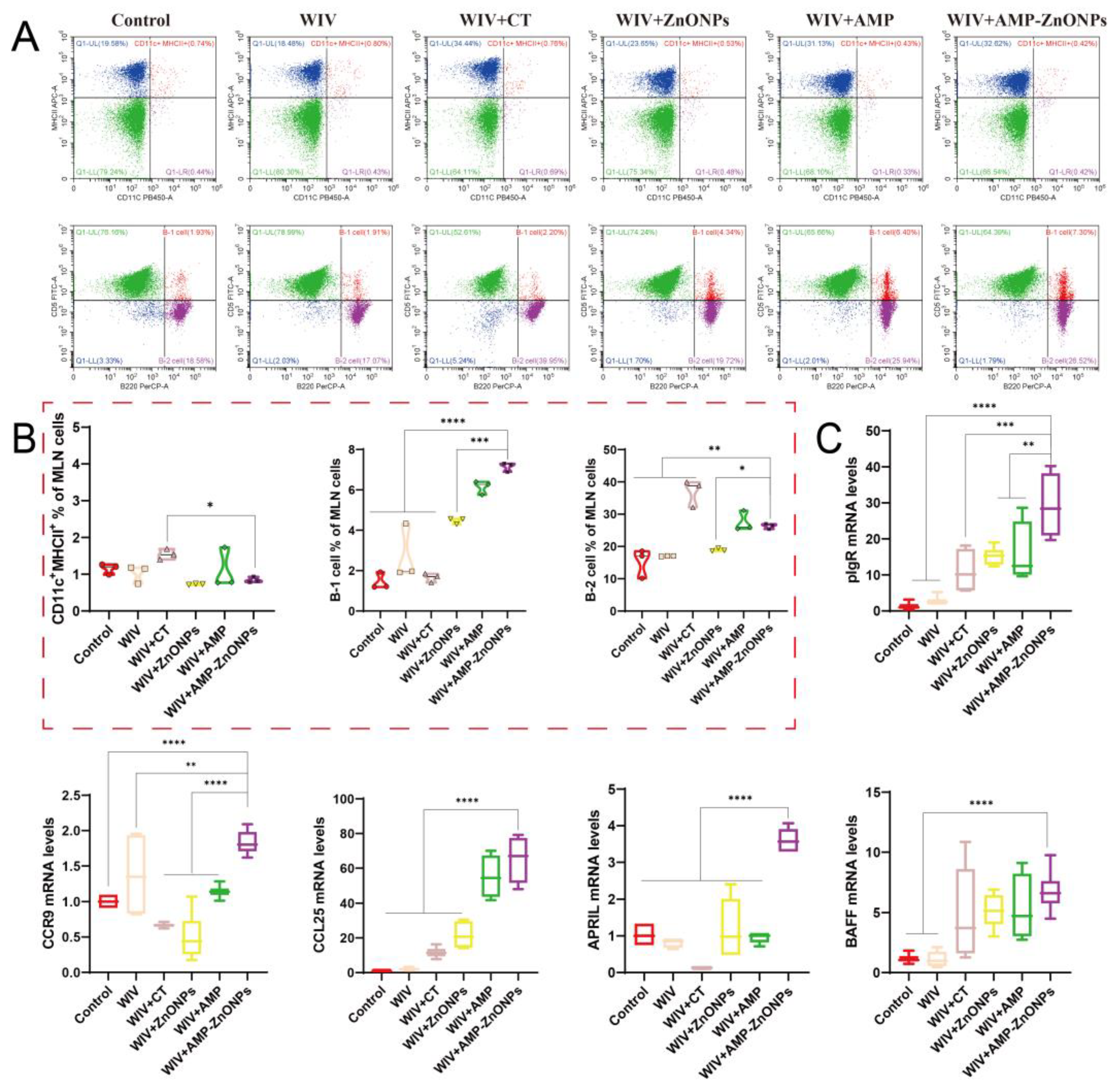
2.6. Effects of Oral Immunization on Dendritic Cells (DCs) or B Cells in the Spleen, Intestinal PP and MLN
2.7. Relative mRNA Expression Levels of Intestinal Mucosal Immune-Related Gene
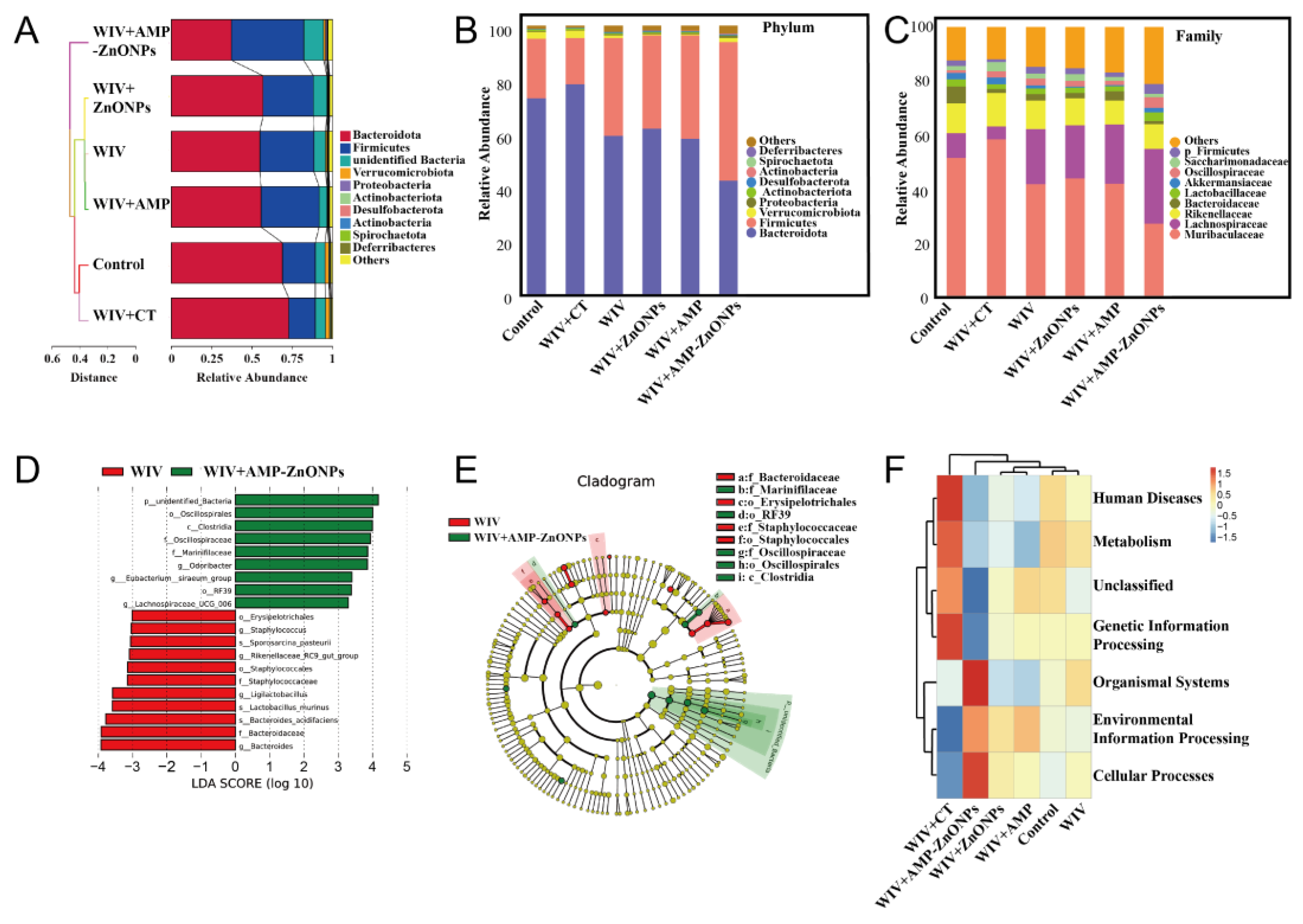
2.8. Effect of AMP-ZnONPs on Intestinal Flora in Mice
2.8.1. Alpha-Diversity and Beta-Diversity
2.8.2. Microbial Community Composition
2.8.3. Differential Intestinal Flora between the Subgroups of WIV and WIV + AMP-ZnONPs
2.8.4. Functional Prediction of Intestinal Microbiota
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Antigens
4.3. Preparation of Vaccines
4.4. Immunization of Animals
4.5. Hemagglutination Inhibition (HI) Assay
4.6. Determination of Anti-H9N2 Specific Secretory Immunoglobulin A (sIgA), IgG, IgG1 and IgG2a by ELISA Assays
4.7. Activation of the CD4+ and CD8+ T Cells in Spleen
4.8. Immunohistochemical Staining for IgA+ Cells
4.9. Analysis of DCs and B-Cell Subsets by Flow Cytometry
4.10. The Expression of mRNA Levels for Cytokines
4.11. Intestinal Microbiota Community Analysis
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, J.; Ma, K.; Li, B.; Chen, Y.; Qiu, Z.; Xing, J.; Huang, J.; Hu, C.; Huang, Y.; Li, H.; et al. A risk marker of tribasic hemagglutinin cleavage site in influenza A (H9N2) virus. Commun. Biol. 2021, 4, 71. [Google Scholar] [CrossRef]
- Pawar, S.D.; Tandale, B.V.; Raut, C.G.; Parkhi, S.S.; Barde, T.D.; Gurav, Y.K.; Kode, S.S.; Mishra, A.C. Avian Influenza H9N2 Seroprevalence among Poultry Workers in Pune, India, 2010. PLoS ONE 2012, 7, e36374. [Google Scholar] [CrossRef] [PubMed]
- Coman, A.; Maftei, D.N.; Krueger, W.S.; Heil, G.L.; Friary, J.A.; Chereches, R.M.; Sirlincan, E.; Bria, P.; Dragnea, C.; Kasler, I.; et al. Serological evidence for avian H9N2 influenza virus infections among Romanian agriculture workers. J. Infect. Public Health 2018, 11, 902. [Google Scholar] [CrossRef] [PubMed]
- Nagy, A.; Mettenleiter, T.C.; Abdelwhab, E.M. A brief summary of the epidemiology and genetic relatedness of avian influenza H9N2 virus in birds and mammals in the Middle East and North Africa. Epidemiol. Infect. 2017, 145, 3320–3333. [Google Scholar] [CrossRef] [PubMed]
- Lycke, N. Recent progress in mucosal vaccine development: Potential and limitations. Nat. Rev. Immunol. 2012, 12, 592–605. [Google Scholar] [CrossRef]
- Santos, F.D.S.; Mazzoli, A.; Maia, A.R.; Saggese, A.; Isticato, R.; Leite, F.; Iossa, S.; Ricca, E.; Baccigalupi, L. A probiotic treatment increases the immune response induced by the nasal delivery of spore-adsorbed TTFC. Microb. Cell Fact. 2020, 19, 42. [Google Scholar] [CrossRef] [PubMed]
- Georgousakis, M.M.; McMillan, D.J.; Batzloff, M.R.; Sriprakash, K.S. Moving forward: A mucosal vaccine against group A streptococcus. Expert Rev. Vaccines 2009, 8, 747–760. [Google Scholar] [CrossRef]
- Koussoulas, K.; Gwynne, R.M.; Foong, J.P.P.; Bornstein, J.C. Cholera Toxin Induces Sustained Hyperexcitability in Myenteric, but Not Submucosal, AH Neurons in Guinea Pig Jejunum. Front. Physiol. 2017, 8, 254. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhang, Q.; Taha, R.; Abdelmotalab, M.I.; Wen, Q.; Yuan, Y.; Zhao, Y.; Li, Q.; Liao, C.; Huang, X.; et al. Polysaccharide from Atractylodes macrocephala Koidz. ameliorates DSS-induced colitis in mice by regulating the Th17/Treg cell balance. Front. Immunol. 2022, 13, 1021695. [Google Scholar] [CrossRef]
- Cui, Y.; Zhang, X.; Jiao, L.; Tao, X.; Li, M.; Lu, J.; Jin, M.; Zhang, L.; Zhou, Q. The regulation effect of Atractylodis macrocephalae polysaccharides on the growth performance, antioxidant capacity and immune function in Litopenaeus vannamei. Aquac. Rep. 2023, 31, 101641. [Google Scholar] [CrossRef]
- Feng, Z.; Yang, R.; Wu, L.; Tang, S.; Wei, B.; Guo, L.; He, L.; Feng, Y. Atractylodes macrocephala polysaccharides regulate the innate immunity of colorectal cancer cells by modulating the TLR4 signaling pathway. OncoTargets Ther. 2019, 12, 7111–7121. [Google Scholar] [CrossRef]
- Heng, B.C.; Zhao, X.; Tan, E.C.; Khamis, N.; Assodani, A.; Xiong, S.; Ruedl, C.; Ng, K.W.; Loo, J.S. Evaluation of the cytotoxic and inflammatory potential of differentially shaped zinc oxide nanoparticles. Arch. Toxicol. 2011, 85, 1517–1528. [Google Scholar] [CrossRef]
- Sharma, P.; Jang, N.Y.; Lee, J.W.; Park, B.C.; Kim, Y.K.; Cho, N.H. Application of ZnO-Based Nanocomposites for Vaccines and Cancer Immunotherapy. Pharmaceutics 2019, 11, 493. [Google Scholar] [CrossRef]
- Bo, R.; Liu, X.; Wang, J.; Wei, S.; Wu, X.; Tao, Y.; Xu, S.; Liu, M.; Li, J.; Pang, H. Polysaccharide from Atractylodes macrocephala Koidz binding with zinc oxide nanoparticles: Characterization, immunological effect and mechanism. Front. Nutr. 2022, 9, 992502. [Google Scholar] [CrossRef]
- Yu, J.; Shi, F.S.; Hu, S. Improved immune responses to a bivalent vaccine of Newcastle disease and avian influenza in chickens by ginseng stem-leaf saponins. Vet. Immunol. Immunopathol. 2015, 167, 147–155. [Google Scholar] [CrossRef]
- Liu, J.; Wan, R.; Xu, X.F.; Wang, X.P.; Yang, W.J.; Xia, Y.J.; Liu, H.; Yan, Q.L.; Yan, D.X.; Guo, C.Y. Effect of Lianshu preparation on lipopolysaccharide-induced diarrhea in rats. World J. Gastroenterol. 2009, 15, 2009–2015. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zuo, Z.; Zhu, P.; Zheng, Z.; Peng, X.; Fang, J.; Cui, H.; Zhou, Y.; Ouyang, P.; Geng, Y.; et al. Sodium selenite prevents suppression of mucosal humoral response by AFB(1) in broiler’s cecal tonsil. Oncotarget 2017, 8, 54215–54226. [Google Scholar] [CrossRef]
- Getahun, A.; Beavers, N.A.; Larson, S.R.; Shlomchik, M.J.; Cambier, J.C. Continuous inhibitory signaling by both SHP-1 and SHIP-1 pathways is required to maintain unresponsiveness of anergic B cells. J. Exp. Med. 2016, 213, 751–769. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Wang, L.; An, S.; Wang, C.; Jiang, Q.; Li, X. Fabrication of quercetin-loaded nanoparticles based on Hohenbuehelia serotina polysaccharides and their modulatory effects on intestinal function and gut microbiota in vivo. Innov. Food Sci. Emerg. 2022, 78, 102993. [Google Scholar] [CrossRef]
- Han, M.; Dsouza, M.; Zhou, C.; Li, H.; Zhang, J.; Chen, C.; Yao, Q.; Zhong, C.; Zhou, H.; Gilbert, J.A.; et al. Agricultural Risk Factors Influence Microbial Ecology in Honghu Lake. Genom. Proteom. Bioinf. 2019, 17, 76–90. [Google Scholar] [CrossRef] [PubMed]
- Echeverry, A.; Saijo, S.; Schesser, K.; Adkins, B. Yersinia enterocolitica promotes robust mucosal inflammatory T-cell immunity in murine neonates. Infect. Immun. 2010, 78, 3595–3608. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Li, M.; Zhang, Z.; Gong, T.; Sun, X. Induction of HIV-1 gag specific immune responses by cationic micelles mediated delivery of gag mRNA. Drug Deliv. 2016, 23, 2596–2607. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Su, G.; Wu, A.; Chen, D.; Yu, B.; Huang, Z.; Luo, Y.; Mao, X.; Zheng, P.; Yu, J.; et al. Bombyx mori gloverin A2 alleviates enterotoxigenic Escherichia coli-induced inflammation and intestinal mucosa disruption. Antimicrob. Resist. Infect. Control 2019, 8, 189. [Google Scholar] [CrossRef] [PubMed]
- Gu, P.F.; Wusiman, A.; Zhang, Y.; Cai, G.F.; Xu, S.W.; Zhu, S.W.; Liu, Z.G.; Hu, Y.L.; Liu, J.G.; Wang, D.Y. Polyethylenimine-coated PLGA nanoparticles-encapsulated Angelica sinensis polysaccharide as an adjuvant for H9N2 vaccine to improve immune responses in chickens compared to Alum and oil-based adjuvants. Vet. Microbiol. 2020, 251, 108894. [Google Scholar] [CrossRef] [PubMed]
- Li, S.L.; Xue, G.H.; Zhao, H.Q.; Feng, Y.L.; Yan, C.; Cui, J.H.; Sun, H.M. The Mycoplasma pneumoniae HapE alters the cytokine profile and growth of human bronchial epithelial cells. Biosci. Rep. 2019, 39, BSR20182201. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.; Akiyama, Y.; Takimoto, H.; Kumazawa, Y. Mechanism of up-regulation of immunoglobulin A production in the intestine of mice unresponsive to lipopolysaccharide. Immunology 2005, 116, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Mkaddem, S.B.; Christou, I.; Rossato, E.; Berthelot, L.; Lehuen, A.; Monteiro, R.C. IgA, IgA Receptors, and Their Anti-inflammatory Properties. Curr. Top. Microbiol. Immunol. 2014, 382, 221–235. [Google Scholar] [CrossRef]
- Li, Q.; Wang, S.; Poungchawanwong, S.; Hou, H. Effect of Peptides from Alaska Pollock on Intestinal Mucosal Immunity Function and Purification of Active Fragments. Nutrients 2019, 11, 2517. [Google Scholar] [CrossRef]
- Block, M.S.; Dietz, A.B.; Gustafson, M.P.; Kalli, K.R.; Erskine, C.L.; Youssef, B.; Vijay, G.V.; Allred, J.B.; Pavelko, K.D.; Strausbauch, M.A.; et al. Th17-inducing autologous dendritic cell vaccination promotes antigen-specific cellular and humoral immunity in ovarian cancer patients. Nat. Commun. 2020, 11, 5173. [Google Scholar] [CrossRef]
- Mora, J.R.; von Andrian, U.H. Differentiation and homing of IgA-secreting cells. Mucosal Immunol. 2008, 1, 96–109. [Google Scholar] [CrossRef]
- Tezuka, H.; Ohteki, T. Regulation of intestinal homeostasis by dendritic cells. Immunol. Rev. 2010, 234, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, A.; Rescigno, M. The biology of intestinal immunoglobulin A responses. Immunity 2008, 28, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, E.; Asanuma, H.; Momose, H.; Furuhata, K.; Mizukami, T.; Matsumura, T.; Takahashi, Y.; Hamaguchi, I. Systemically inoculated adjuvants stimulate pDC-dependent IgA response in local site. Mucosal Immunol. 2023, 16, 275–286. [Google Scholar] [CrossRef]
- Mora, J.R.; Iwata, M.; Eksteen, B.; Song, S.Y.; Junt, T.; Senman, B.; Otipoby, K.L.; Yokota, A.; Takeuchi, H.; Ricciardi-Castagnoli, P.; et al. Generation of gut-homing IgA-secreting B cells by intestinal dendritic cells. Science 2006, 314, 1157–1160. [Google Scholar] [CrossRef]
- Hardenberg, G.; Planelles, L.; Schwarte, C.M.; van Bostelen, L.; Le Huong, T.; Hahne, M.; Medema, J.P. Specific TLR ligands regulate APRIL secretion by dendritic cells in a PKR-dependent manner. Eur. J. Immunol. 2007, 37, 2900–2911. [Google Scholar] [CrossRef]
- Foxx, C.L.; Nagy, M.R.; King, A.E.; Albin, D.; DeKrey, G.K. TCDD exposure alters fecal IgA concentrations in male and female mice. BMC Pharmacol. Toxicol. 2022, 23, 25. [Google Scholar] [CrossRef] [PubMed]
- Grasset, E.K.; Chorny, A.; Casas-Recasens, S.; Gutzeit, C.; Bongers, G.; Thomsen, I.; Chen, L.; He, Z.; Matthews, D.B.; Oropallo, M.A.; et al. Gut T cell-independent IgA responses to commensal bacteria require engagement of the TACI receptor on B cells. Sci. Immunol. 2020, 5, eaat7117. [Google Scholar] [CrossRef]
- Zhao, T.; Li, J.; Fu, Y.T.; Ye, H.; Liu, X.C.; Li, G.L.; Yang, X.L.; Yang, J.S. Influence of gut microbiota on mucosal IgA antibody response to the polio vaccine. NPJ Vaccines 2020, 5, 47. [Google Scholar] [CrossRef]
- Hutin, D.; Hagen, K.A.; Shao, P.; Sugamori, K.; Grant, D.M.; Matthews, J. Reduced Colonic Mucosal Injury in 2,3,7,8-Tetrachlorodibenzo-p-Dioxin Poly ADP-Ribose Polymerase (TIPARP/PARP7)-Deficient Mice. Int. J. Mol. Sci. 2022, 23, 920. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Steed, H.; Macfarlane, S. Bacterial metabolism and health-related effects of galacto-oligosaccharides and other prebiotics. J. Appl. Microbiol. 2008, 104, 305–344. [Google Scholar] [CrossRef]
- Borda-Molina, D.; Vital, M.; Sommerfeld, V.; Rodehutscord, M.; Camarinha-Silva, A. Insights into Broilers’ Gut Microbiota Fed with Phosphorus, Calcium, and Phytase Supplemented Diets. Front. Microbiol. 2016, 7, 2033. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.J.; Xue, J.; Bai, Y.; Wang, J.D.; Zhang, Y.L.; Zhou, D.Y. Pathogenicty and immune prophylaxis of cag pathogenicity island gene knockout homogenic mutants. World J. Gastroenterol. 2004, 10, 3289–3291. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Raw PE | Q20 | Q30 | Effective % |
|---|---|---|---|---|
| Control 1 | 93,386 | 97.67 | 92.91 | 80.65 |
| Control 2 | 82,334 | 97.77 | 93.14 | 75.82 |
| Control 3 | 95,407 | 97.64 | 92.65 | 74.14 |
| WIV 1 | 84,102 | 97.72 | 92.98 | 73.55 |
| WIV 2 | 77,857 | 97.69 | 92.93 | 77.64 |
| WIV 3 | 54,435 | 97.98 | 93.54 | 76.30 |
| WIV + CT 1 | 83,070 | 97.71 | 92.91 | 71.07 |
| WIV + CT 2 | 79,503 | 97.90 | 93.51 | 70.07 |
| WIV + CT 3 | 85,281 | 97.97 | 93.68 | 77.07 |
| WIV + ZnONPs 1 | 88,067 | 97.68 | 92.95 | 75.21 |
| WIV + ZnONPs 2 | 84,474 | 97.50 | 92.44 | 75.76 |
| WIV + ZnONPs 3 | 84,056 | 97.62 | 92.73 | 77.16 |
| WIV + AMP 1 | 88,093 | 97.64 | 92.80 | 76.04 |
| WIV + AMP 2 | 81,627 | 97.70 | 92.95 | 76.70 |
| WIV + AMP 3 | 86,940 | 97.74 | 92.99 | 71.95 |
| WIV + AMP-ZnONPs 1 | 87,192 | 97.76 | 93.08 | 83.56 |
| WIV + AMP-ZnONPs 2 | 86,835 | 97.83 | 93.34 | 86.75 |
| WIV + AMP-ZnONPs 3 | 87,073 | 97.73 | 93.07 | 75.05 |
| Group | Vaccination on d0 and d14 | Dose (Per Mouse) |
|---|---|---|
| Control | PBS | 0.2 mL |
| WIV + ZnONPs | WIV + ZnONPs | 0.2 mL |
| WIV + AMP | WIV + AMP | 0.2 mL |
| WIV + AMP-ZnONPs | WIV + AMP-ZnONPs | 0.2 mL |
| WIV | WIV | 0.2 mL |
| WIV + CT | WIV + CT | 0.2 mL |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| GAPDH | ATGGTGAAGGTCGGTGTGAA | CCTTGACTGTGCCGTTGAAT |
| α-chain | TGAGCGCTGGAACAGTGGCG | TCAGGGCCAGCTCCTCCGAC |
| J-chain | GGATCCTAATGAGGACATTGTGGAG | CTGGGTGGCAGTAACAACCTGA |
| pIgR | TCGATGTCAGCCTGGAGGTC | AGGGCATTCAATGGTCACATTTC |
| IL-6 | TTCCATCCAGTTGCCTTCTTG | AATTAAGCCTCCGACTTGTGAA |
| TNF-α | ATGAGCACAGAAAGCATGATCCGC | AAAGTAGACCTGCCCGGACTC |
| CCL25 | CCGGCATGCTAGGAATTATCA | GGCACTCCTCACGCTTGTACT |
| CCR9 | CTTCAGCTATGACTCCACTGC | CAAGGTGCCCACAATGAACA |
| APRIL | CTTTCGGTTGCTCTTTGGTTG | CGACAGCACAAGTCACAGC |
| BAFF | CAGCGACACGCCGACTATAC | CCTCCAAGGCATTTCCTCTTTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Lin, X.; Hong, H.; Wang, J.; Tao, Y.; Huai, Y.; Pang, H.; Liu, M.; Li, J.; Bo, R. Polysaccharide from Atractylodes macrocephala Koidz Binding with Zinc Oxide Nanoparticles as a Novel Mucosal Immune Adjuvant for H9N2 Inactivated Vaccine. Int. J. Mol. Sci. 2024, 25, 2132. https://doi.org/10.3390/ijms25042132
Liu X, Lin X, Hong H, Wang J, Tao Y, Huai Y, Pang H, Liu M, Li J, Bo R. Polysaccharide from Atractylodes macrocephala Koidz Binding with Zinc Oxide Nanoparticles as a Novel Mucosal Immune Adjuvant for H9N2 Inactivated Vaccine. International Journal of Molecular Sciences. 2024; 25(4):2132. https://doi.org/10.3390/ijms25042132
Chicago/Turabian StyleLiu, Xiaopan, Xinyi Lin, Hailong Hong, Jing Wang, Ya Tao, Yuying Huai, Huan Pang, Mingjiang Liu, Jingui Li, and Ruonan Bo. 2024. "Polysaccharide from Atractylodes macrocephala Koidz Binding with Zinc Oxide Nanoparticles as a Novel Mucosal Immune Adjuvant for H9N2 Inactivated Vaccine" International Journal of Molecular Sciences 25, no. 4: 2132. https://doi.org/10.3390/ijms25042132
APA StyleLiu, X., Lin, X., Hong, H., Wang, J., Tao, Y., Huai, Y., Pang, H., Liu, M., Li, J., & Bo, R. (2024). Polysaccharide from Atractylodes macrocephala Koidz Binding with Zinc Oxide Nanoparticles as a Novel Mucosal Immune Adjuvant for H9N2 Inactivated Vaccine. International Journal of Molecular Sciences, 25(4), 2132. https://doi.org/10.3390/ijms25042132