
Populus euphratica GRP2 Interacts with Target mRNAs to Negatively Regulate Salt Tolerance by Interfering with Photosynthesis, Na+, and ROS Homeostasis


Abstract
1. Introduction
2. Results
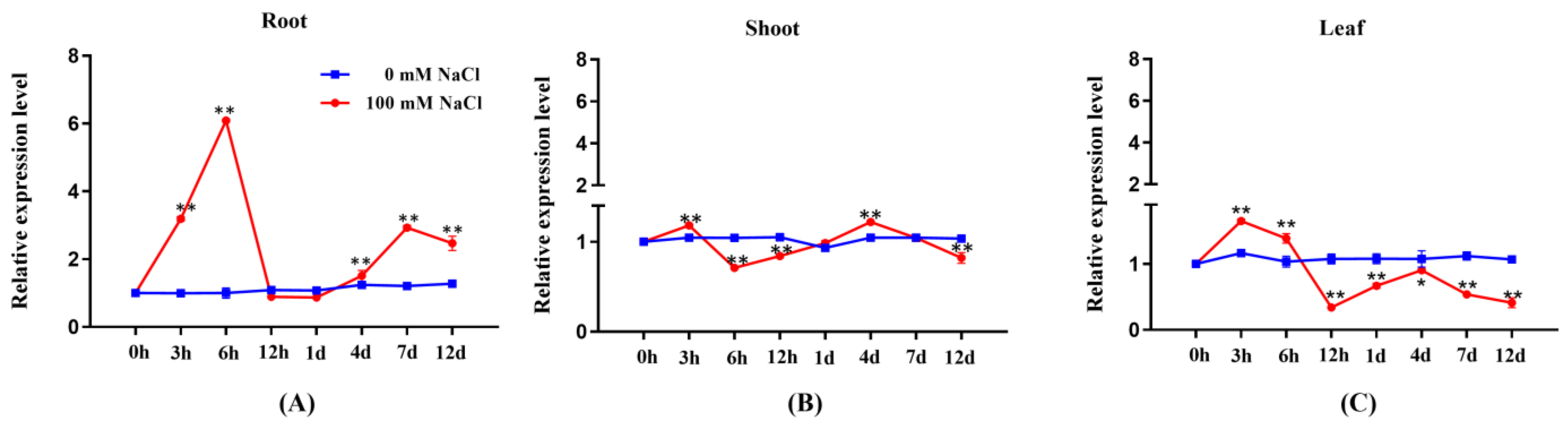
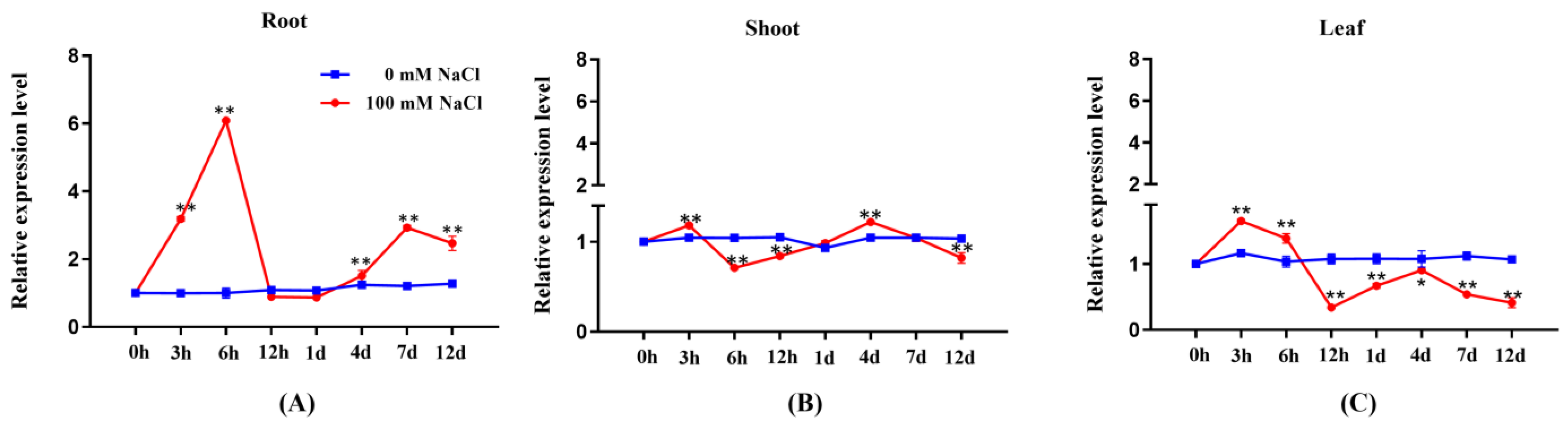
2.1. Expression Profile of PeGRP2 in Salt-Stressed P. euphratica
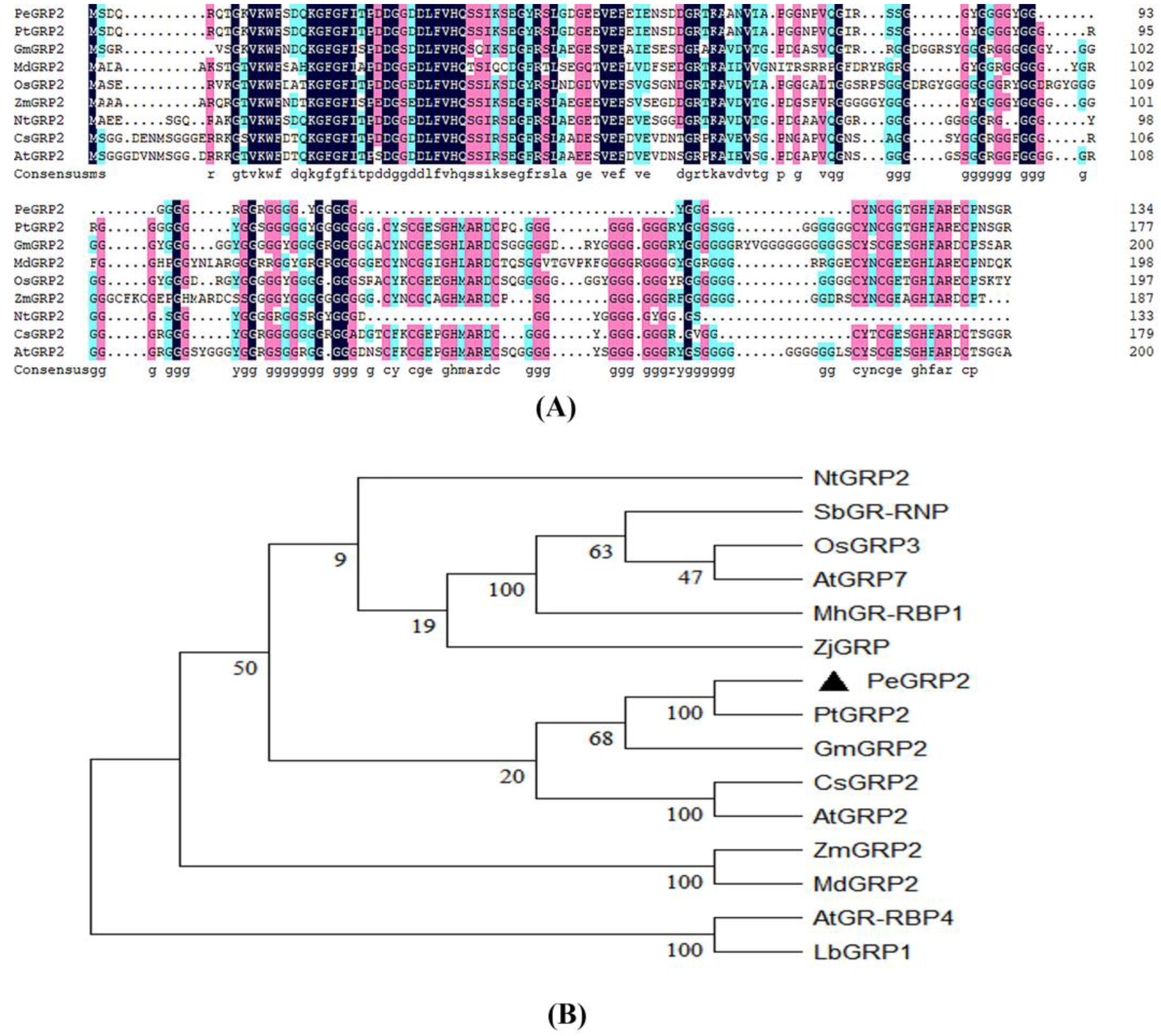
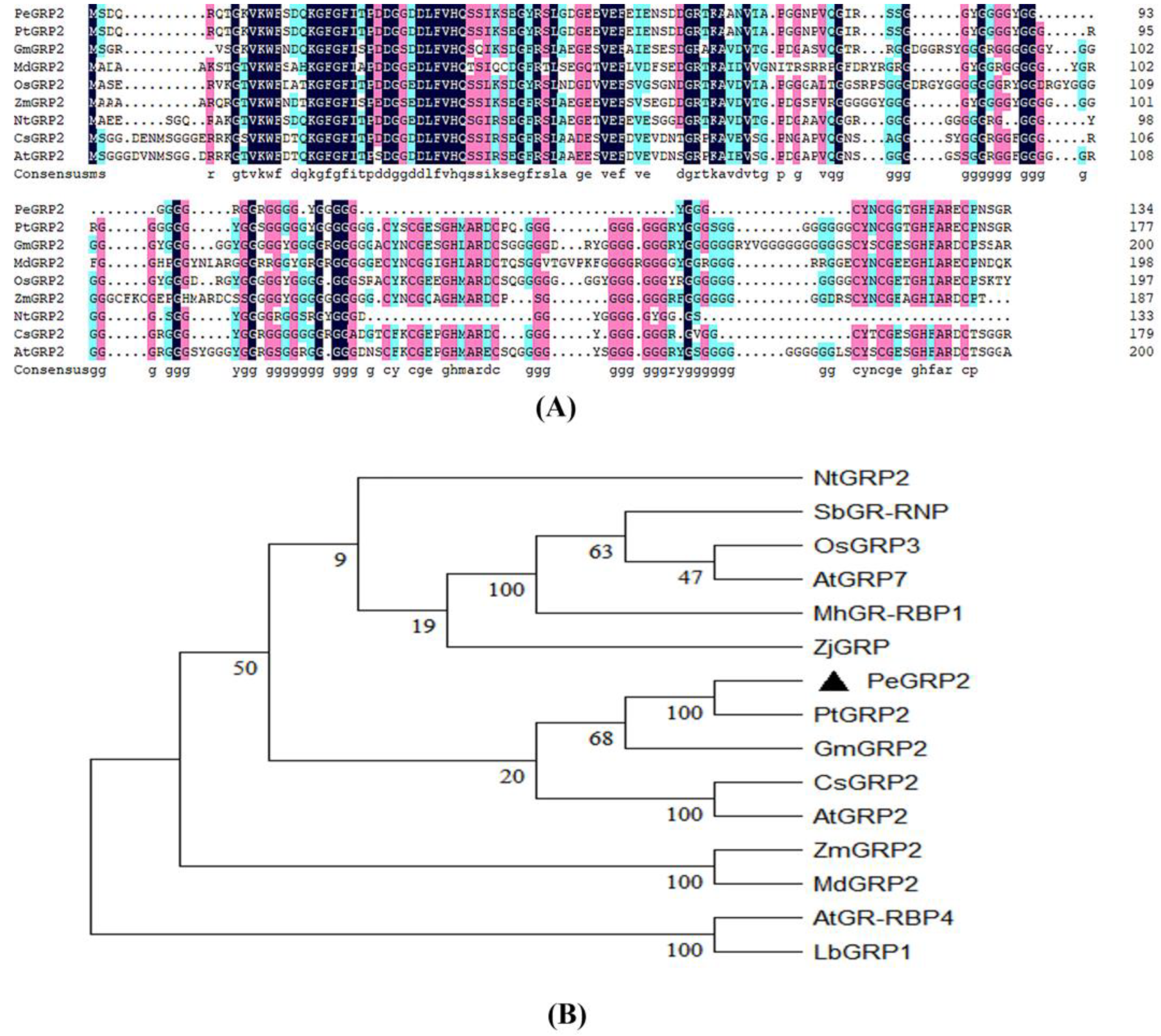
2.2. Sequence Analyses of PeGRP2
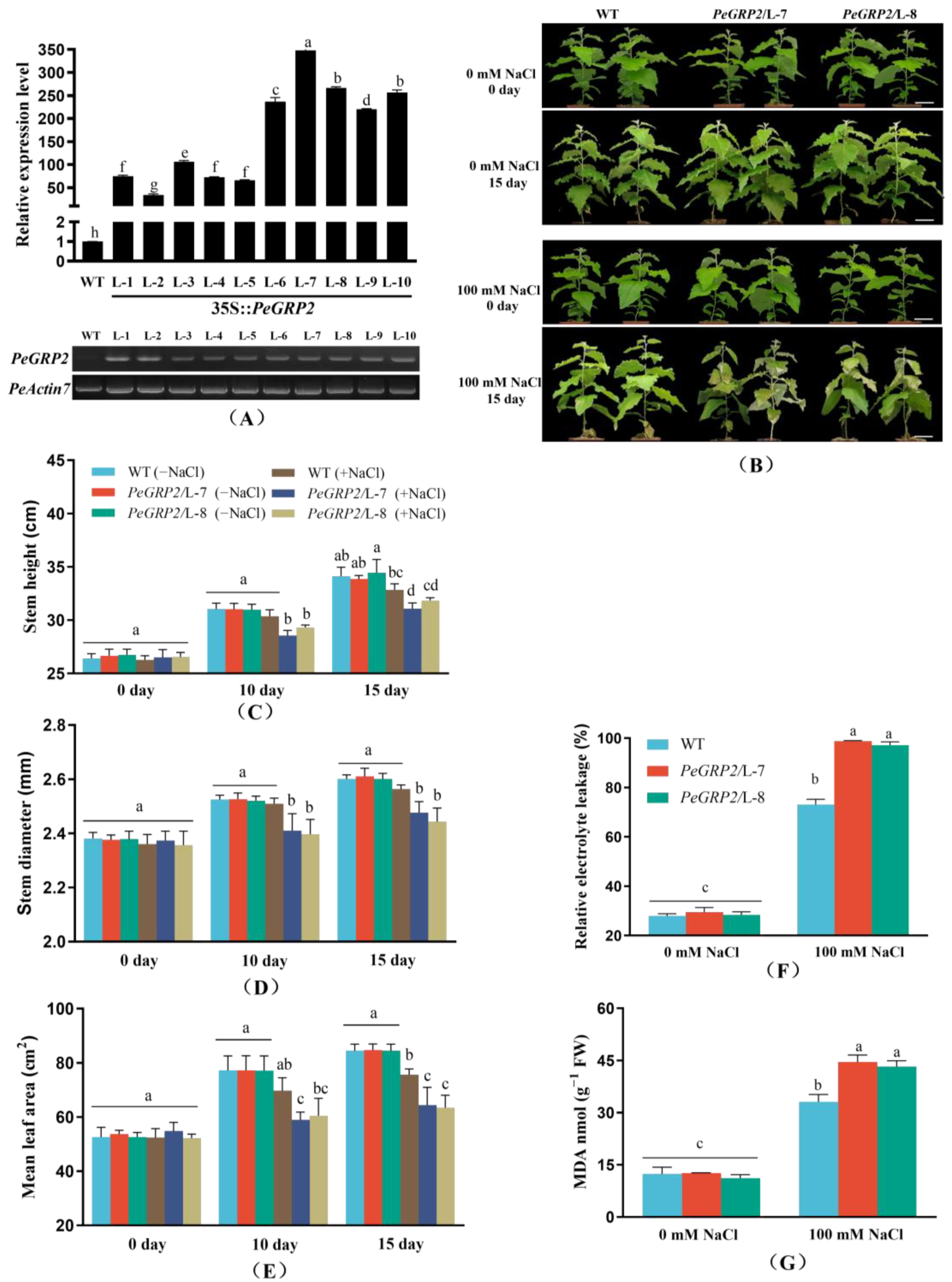
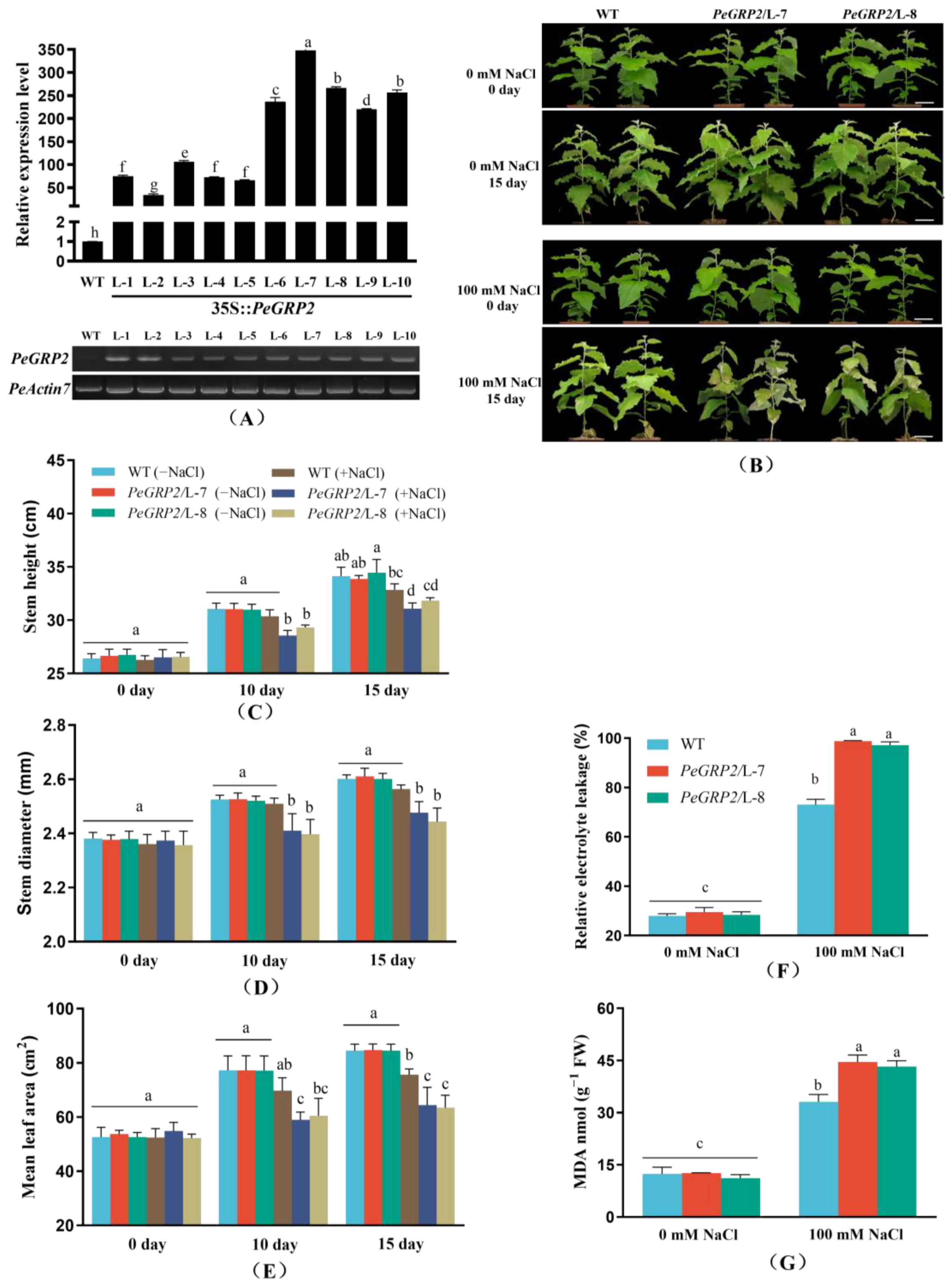
2.3. Phenotypic Tests of PeGRP2-Overexpressing Poplars under Salt Stress
2.4. Membrane Permeability and Lipid Peroxidation
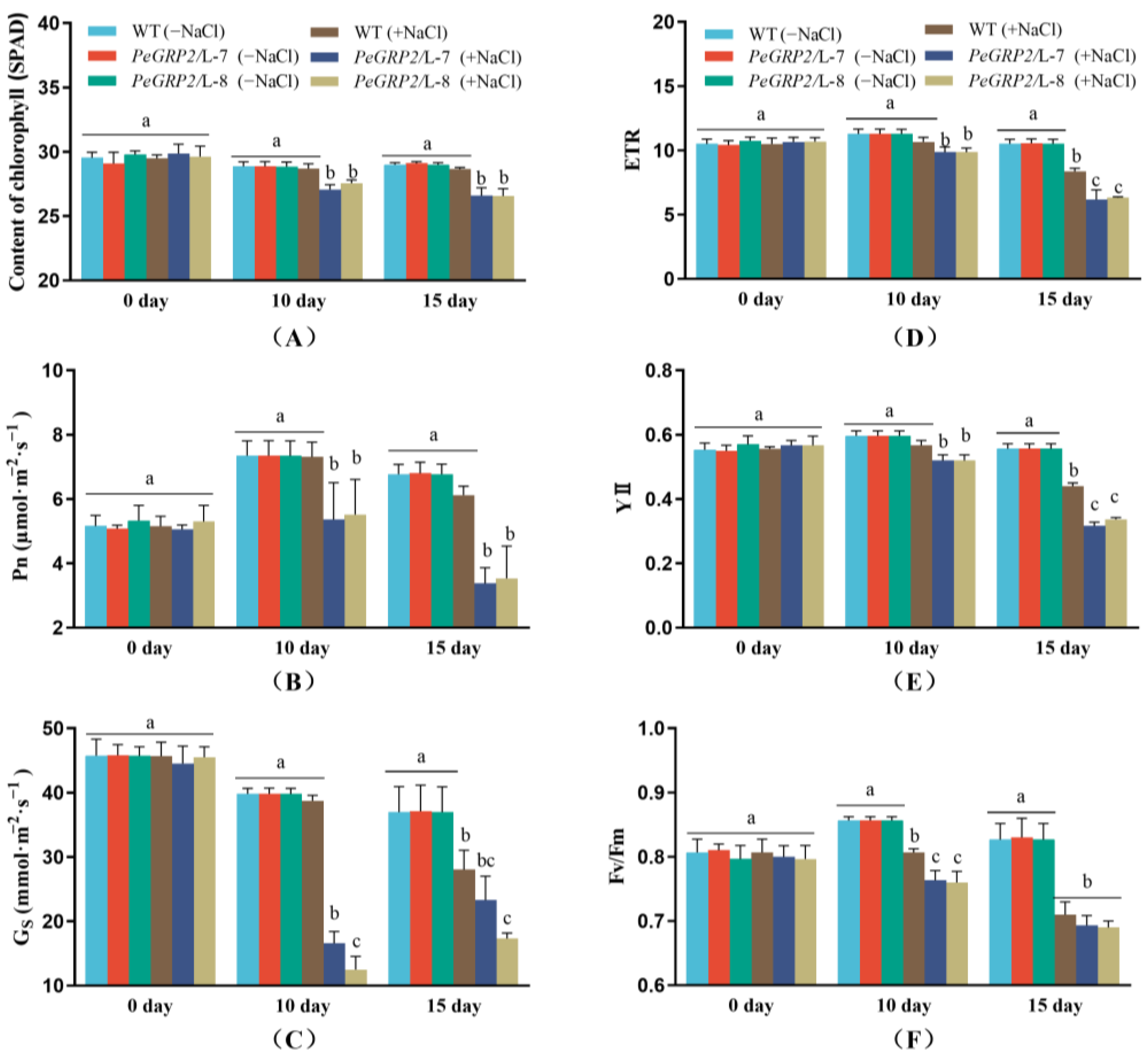
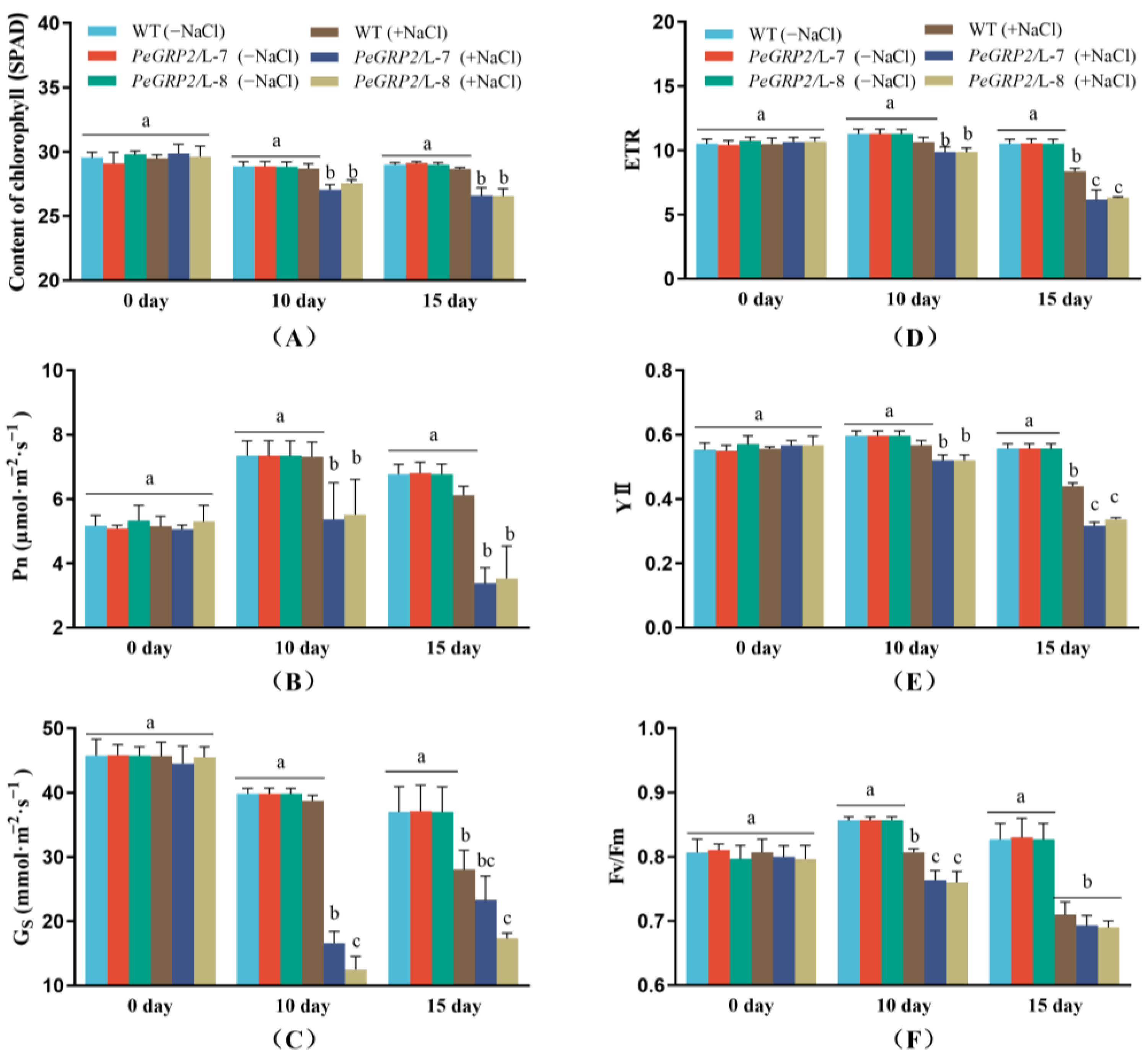
2.5. Photosynthetic Capacity of the Salinized Poplars
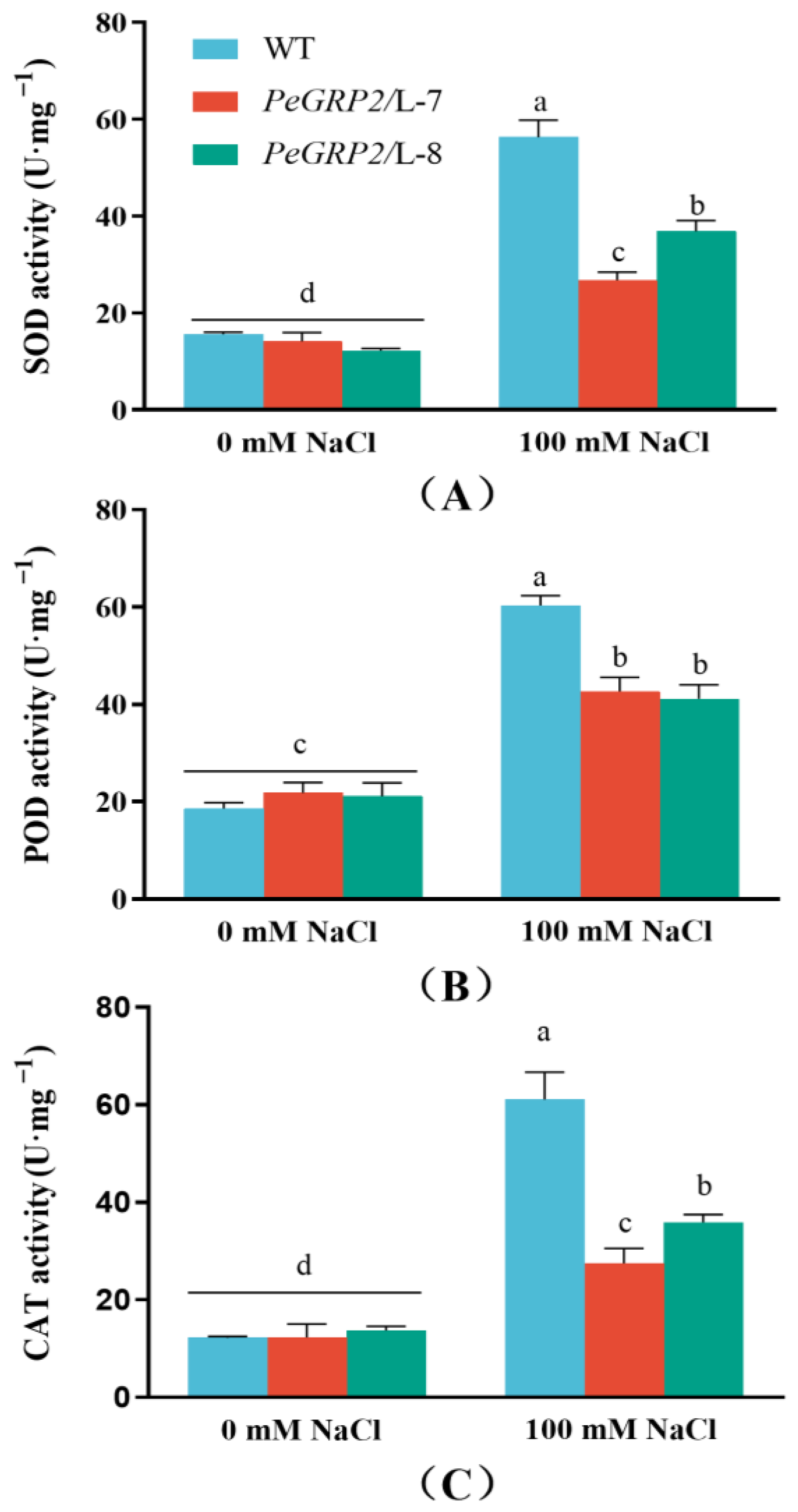
2.6. Activity of Antioxidant Enzymes
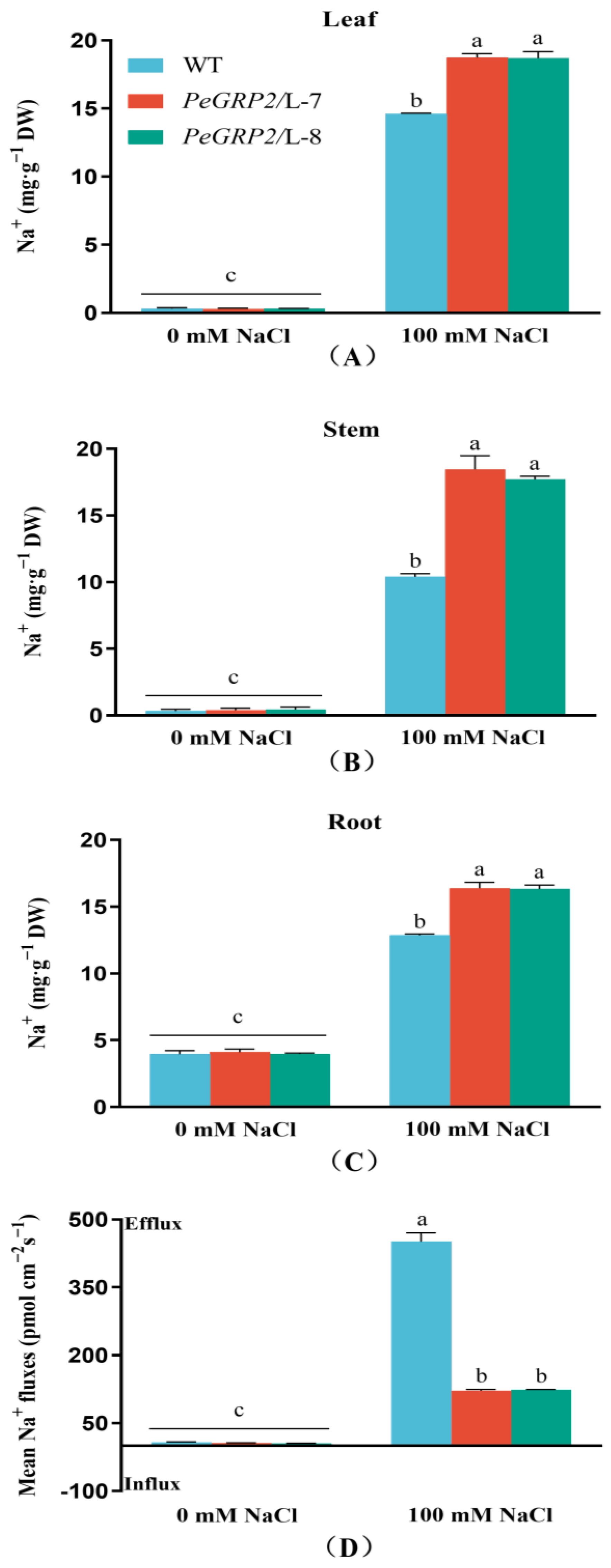
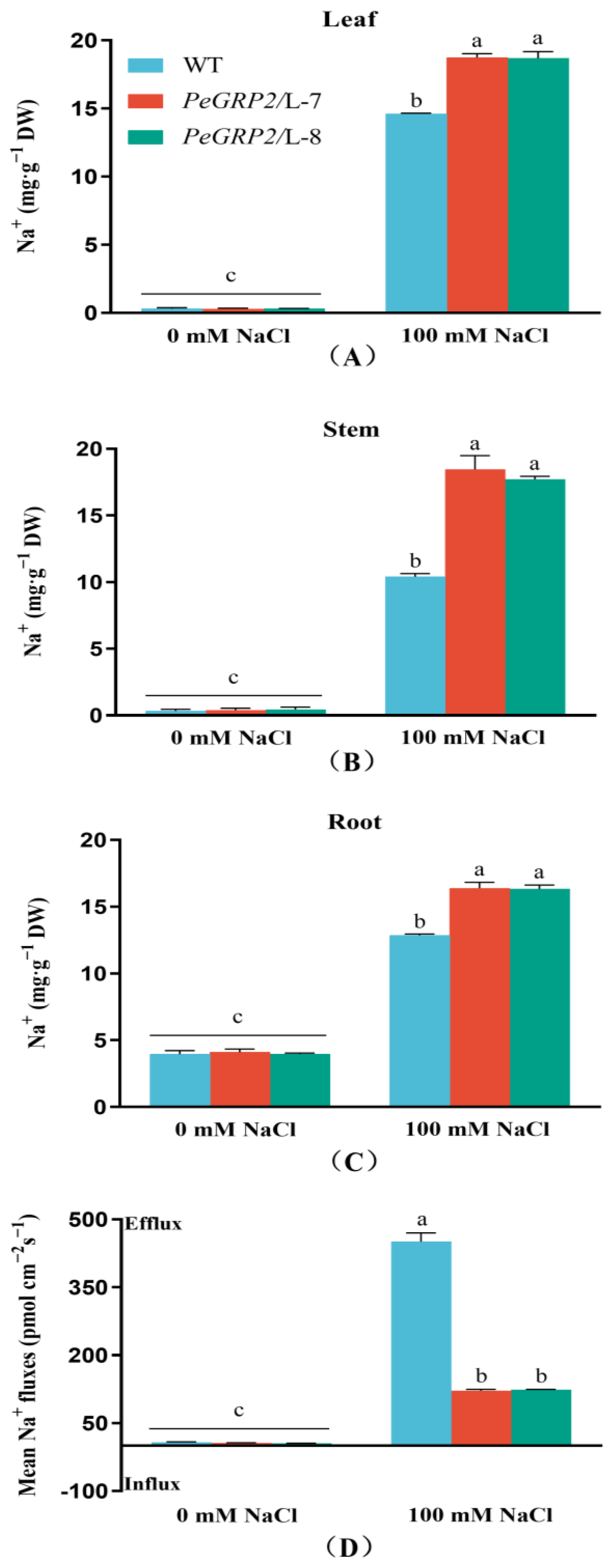
2.7. Na+ Content and Flux in Roots
2.8. PeGRP2-Interacting Target mRNAs in P. × Canescens
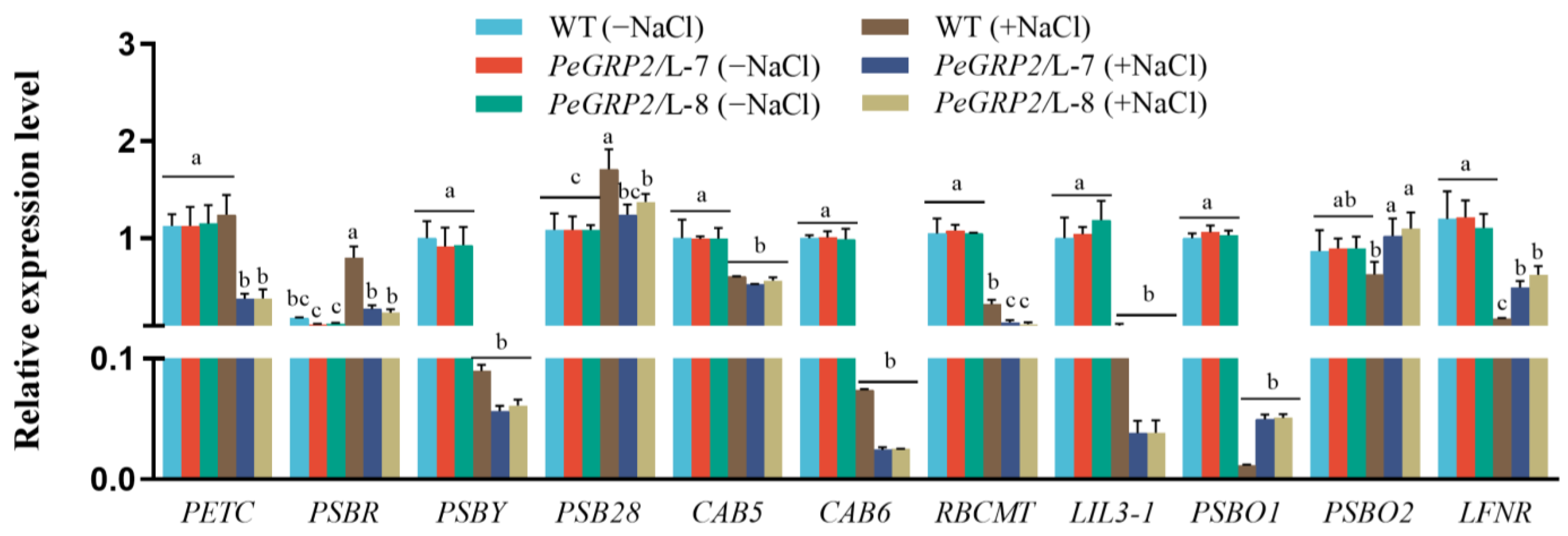
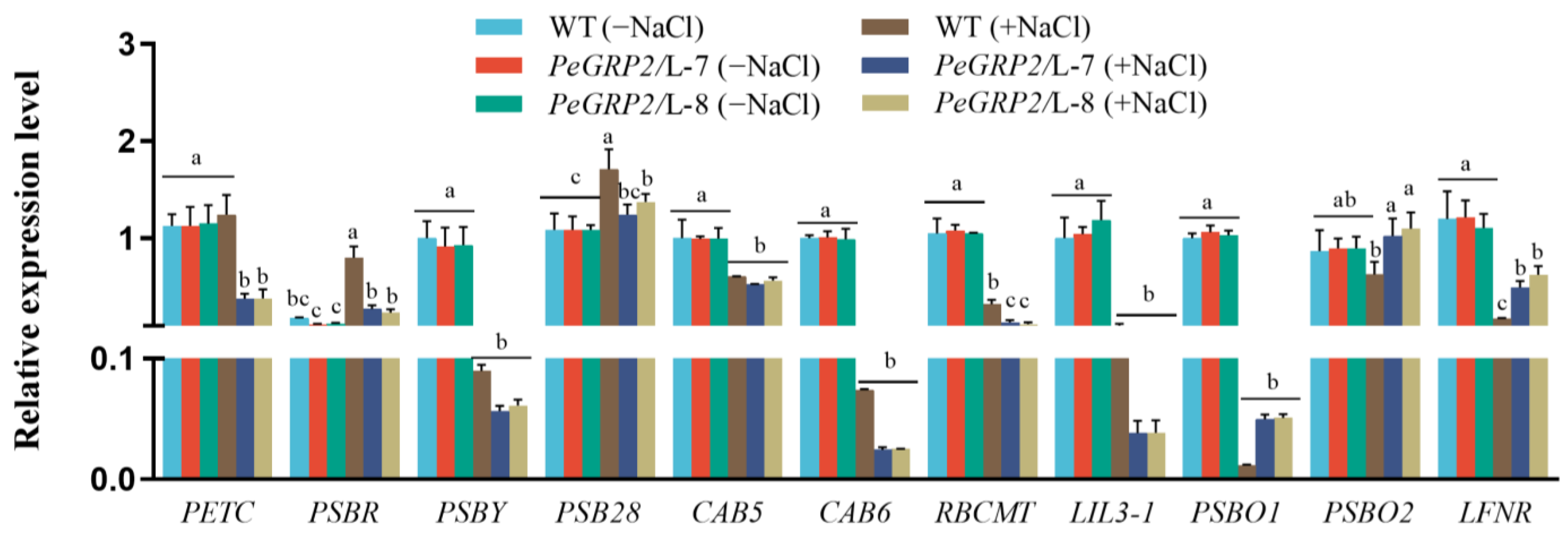
- Chloroplastic photosynthetic proteins: cytochrome b6-f complex iron-sulfur subunit (PETC), photosystem II 10 kDa polypeptide (PSBR), photosystem II core complex protein psbY (PSBY), photosystem II reaction center PSB28 protein (PSB28), chlorophyll a/b-binding protein 5 (CAB5), CAB6, ribulose-1, 5 bisphosphate carboxylase/oxygenase large subunit N-methyltransferase (RBCMT), light-harvesting complex-like protein 3 isotype 1 (LIL3-1), oxygen-evolving enhancer protein 1 (PSBO1), PSBO2, and ferredoxin-NADP reductase (LFNR).
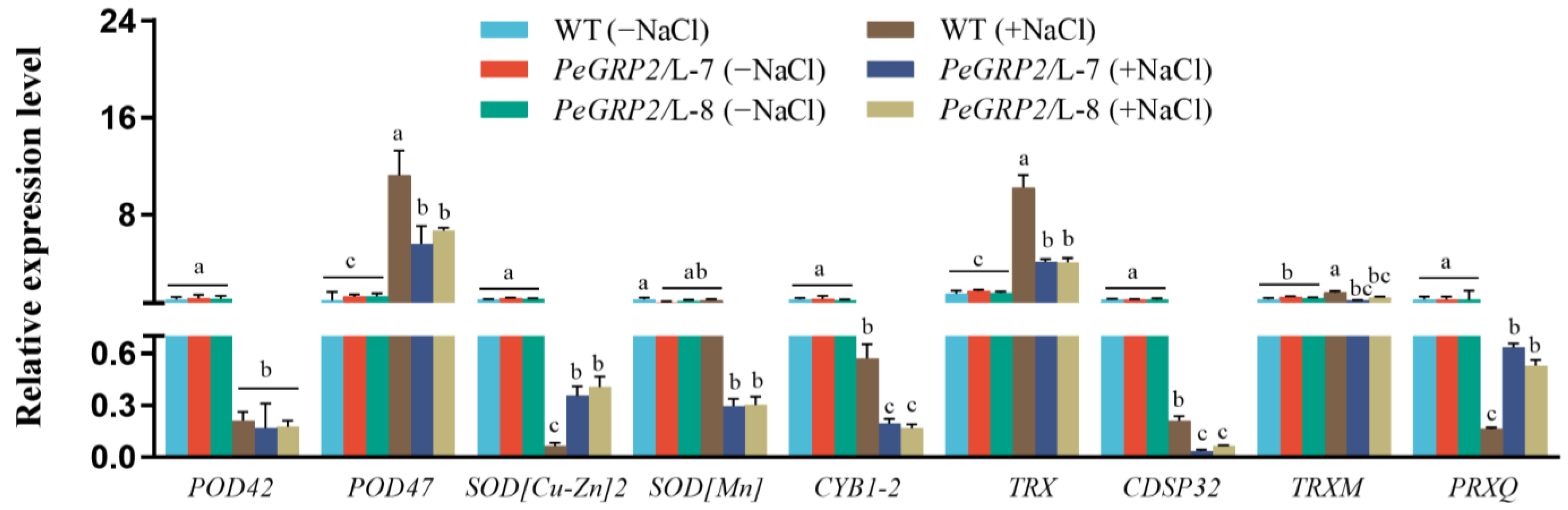
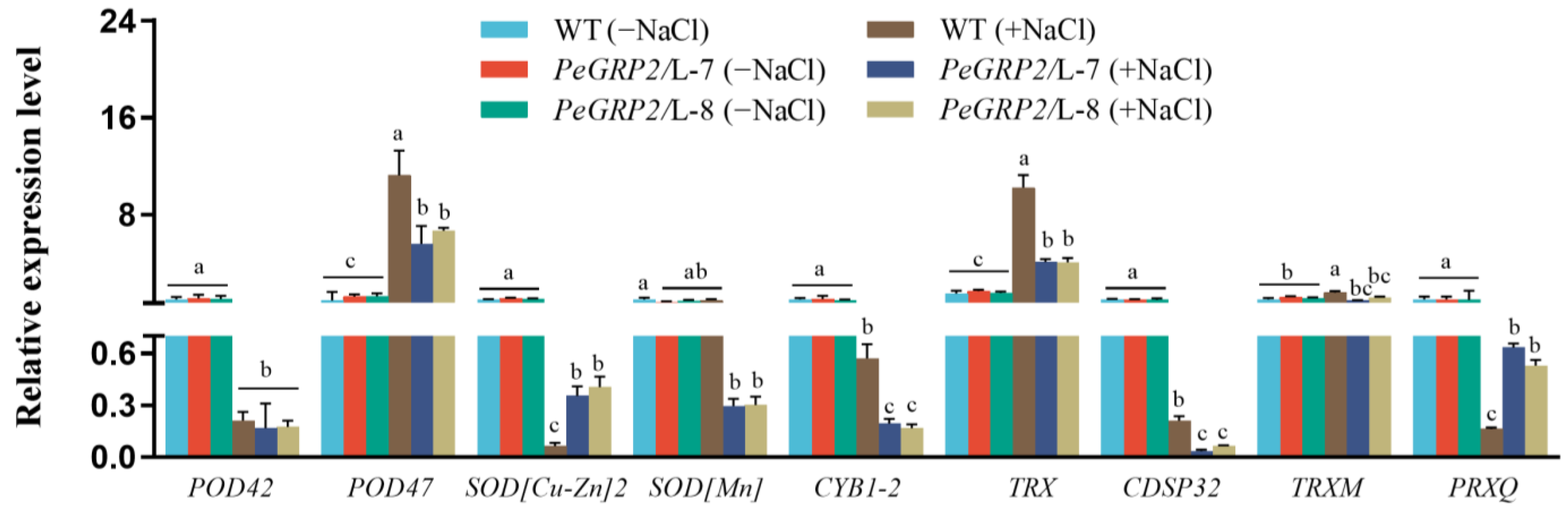
- Antioxidant enzymes: peroxidase 42 (POD42), POD47, superoxide dismutase (SOD [Cu-Zn]2), mitochondrial SOD[Mn], transmembrane ascorbate ferrireductase 1 isoform X2 (CYB1-2), chloroplastic thioredoxin X (TRX), chloroplastic thioredoxin-like protein CDSP32 (CDSP32), chloroplastic thioredoxin M-type (TRXM), and chloroplastic peroxiredoxin Q (PRXQ).
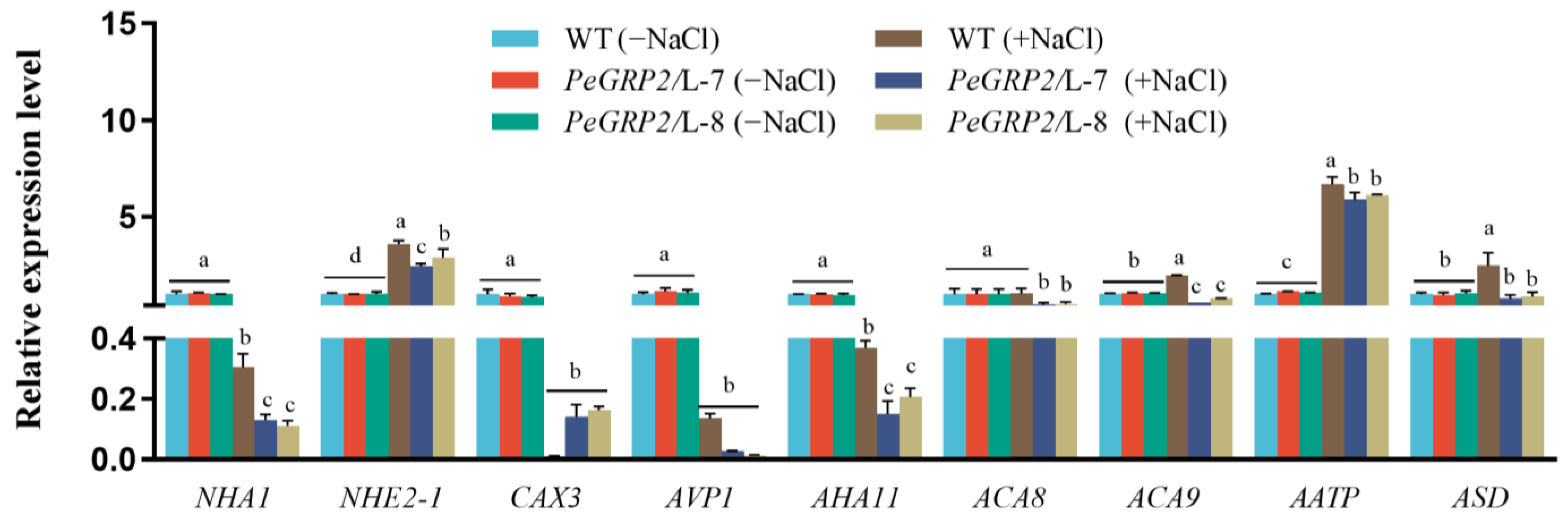
- Cation/H+ exchangers and ATPases: sodium/proton antiporter 1 (NHA1), sodium/hydrogen exchanger 2 isoform X1 (NHE2-1), vacuolar cation/proton exchanger 3 (CAX3), pyrophosphate-energized vacuolar membrane proton pump 1 (AVP1), plasma membrane (PM)-type ATPase 11, (AHA11), PM-type calcium-transporting ATPase 8 (ACA8), ACA9, AAA-ATPase At1g43910 (AATP), and mitochondrial AAA-ATPase ASD (ASD).
2.9. Transcriptional Profiling of the PeGRP2-Interacting mRNAs under Salt Stress
2.9.1. Transcripts of the PeGRP2 Target mRNAs Encoding Photosynthetic Proteins
2.9.2. Transcripts of the PeGRP2 Target mRNAs Encoding Antioxidant Enzymes
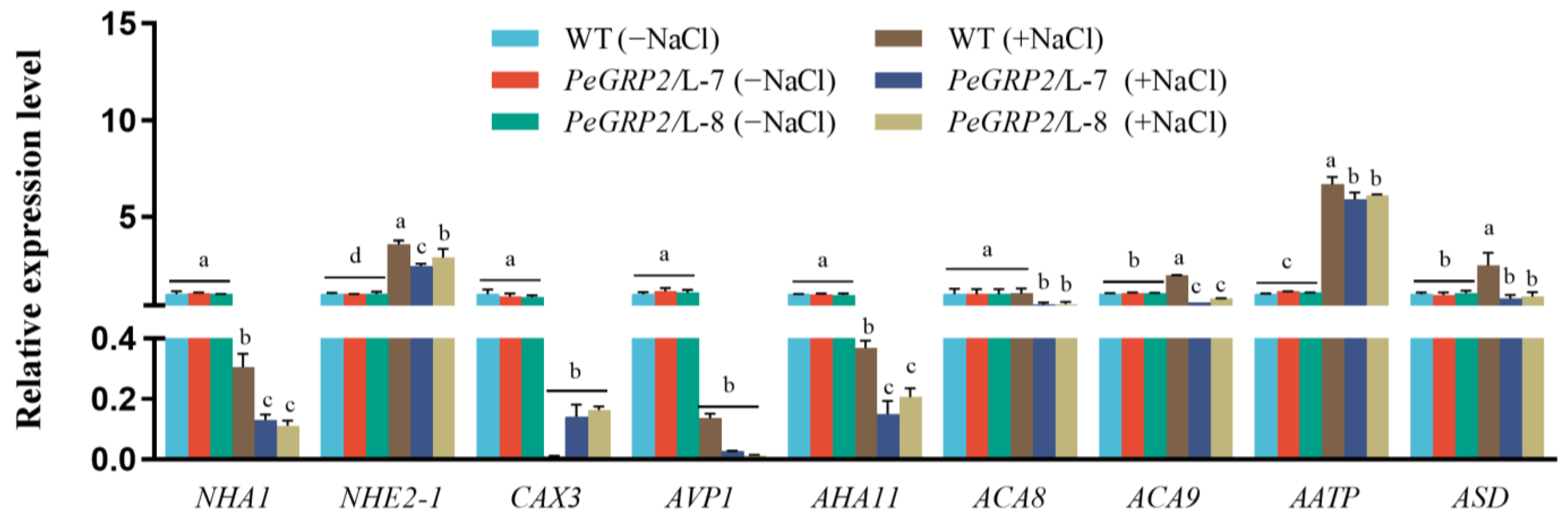
2.9.3. Transcripts of the PeGRP2 Target mRNAs Encoding Cation/H+ Exchangers and ATPases
3. Discussion
3.1. PeGRP2 Increases the Salt Sensitivity of Transgenic P. × canescens
3.2. PeGRP2 Interacts with Target mRNAs Encoding Photosynthetic Proteins and Affects Photosynthesis under Salt Stress
3.3. PeGRP2 Interacts with Target mRNAs Encoding Antioxidant Enzymes and Affects the ROS Scavenging Capacity under Salt Stress
3.4. PeGRP2 Interacts with Target mRNAs Encoding ATPases and Na+/H+ Transporters and Affects Na+ Homeostasis under Salt Stress
4. Materials and Methods
4.1. NaCl Treatment of P. euphratica
4.2. PeGRP2 Cloning and Sequence Analysis
4.3. PeGRP2 Transformation into P. × canescens
4.4. Phenotype Testing of Salt Tolerance
4.4.1. Growth Measurement
4.4.2. Relative Electrolyte Leakage (REL) and Malondialdehyde (MDA) Content
4.4.3. Measurement of the Chlorophyll Fluorescence and Gas Exchange
4.5. Determination of the Antioxidative Enzyme Activity
4.6. Na+ Concentration in the Roots, Leaves, and Stems
4.7. Flux Records of Na+ in the Roots
4.8. RNA Affinity Purification Sequencing
4.9. Semiquantitative RT-PCR and RT-qPCR Analysis
4.10. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.M.; Zhu, J.H.; Gong, Z.Z.; Zhu, J.K. Abiotic stress responses in plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.B.; Polle, A. Wood composition and energy content in a poplar short rotation plantation on fertilized agricultural land in a future CO2 atmosphere. Glob. Chang. Biol. 2009, 15, 38–47. [Google Scholar] [CrossRef]
- Polle, A.; Douglas, C. The molecular physiology of poplars: Paving the way for knowledge-based biomass production. Plant Biol. 2010, 12, 239–241. [Google Scholar] [CrossRef] [PubMed]
- Polle, A.; Chen, S.L. On the salty side of life: Molecular, physiological and anatomical adaptation and acclimation of trees to extreme habitats. Plant Cell Environ. 2015, 38, 1794–1816. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Shen, Z.D.; Zhang, Y.L.; Wu, X.; Wang, J.H.; Sang, G.; Zhang, Y.H.; Zhang, H.L.; Deng, C.; Liu, J.; et al. Populus euphratica WRKY1 binds the promoter of H+-ATPase gene to enhance gene expression and salt tolerance. J. Exp. Bot. 2020, 71, 1527–1539. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yin, K.X.; Yao, J.; Zhao, Z.Y.; Liu, Z.; Yan, C.X.; Zhang, Y.L.; Liu, J.; Li, J.; Zhao, N.; et al. Populus euphratica GLABRA3 binds PLDδ promoters to enhance salt tolerance. Int. J. Mol. Sci. 2023, 24, 8208. [Google Scholar] [CrossRef]
- Li, D.D.; Song, S.Y.; Xia, X.L.; Yin, W.L. Two CBL genes from Populus euphratica confer multiple stress tolerance in transgenic triploid white poplar. Plant Cell Tissue Organ. Cult. 2012, 109, 477–489. [Google Scholar] [CrossRef]
- Qiu, Q.; Ma, T.; Hu, Q.J.; Liu, B.B.; Wu, Y.X.; Zhou, H.H.; Wang, Q.; Wang, J.; Liu, J.Q. Genome-scale transcriptome analysis of the desert poplar, Populus euphratica. Tree Physiol. 2011, 31, 452–461. [Google Scholar] [CrossRef]
- Zhang, H.C.; Yin, W.L.; Xia, X.L. Calcineurin B-like family in Populus: Comparative genome analysis and expression pattern under cold, drought and salt stress treatment. Plant Growth Regul. 2008, 56, 129–140. [Google Scholar] [CrossRef]
- Zhang, H.C.; Lv, F.L.; Han, X.; Xia, X.L.; Yin, W.L. The calcium sensor PeCBL1, interacting with PeCIPK24/25 and PeCIPK26, regulates Na+/K+ homeostasis in Populus euphratica. Plant Cell Rep. 2013, 32, 611–621. [Google Scholar] [CrossRef]
- Zhang, J.; Xie, P.H.; Lascoux, M.; Meagher, T.R.; Liu, J.Q. Rapidly evolving genes and stress adaptation of two desert poplars, Populus euphratica and P. pruinosa. PLoS ONE 2013, 8, e66370. [Google Scholar] [CrossRef]
- Qu, M.Q.; Sun, Q.; Chen, N.N.; Chen, Z.Y.; Zhang, H.C.; Lv, F.L.; An, Y. Functional characterization of a new salt stress response gene, PeCBL4, in Populus euphratica Oliv. Forests 2023, 14, 1504. [Google Scholar] [CrossRef]
- Meng, H.J.; Zhao, J.N.; Yang, Y.F.; Diao, K.H.; Zheng, G.S.; Li, T.; Dai, X.R.; Li, J.B. PeGSTU58, a glutathione S-transferase from Populus euphratica, enhances salt and drought stress tolerance in transgenic Arabidopsis. Int. J. Mol. Sci. 2023, 24, 9354. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.L.; Du, J.J.; Zhang, L.; Qu, G.Z.; Hu, J.J. PeCLH2 gene positively regulate salt tolerance in transgenic Populus alba × Populus glandulosa. Genes 2023, 14, 538. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.L.; Zhang, L.; Du, J.J.; Wen, S.S.; Qu, G.Z.; Hu, J.J. Transcriptome analysis of Populus euphratica under salt treatment and PeERF1 gene enhances salt tolerance in transgenic Populus alba × Populus glandulosa. Int. J. Mol. Sci. 2022, 23, 3727. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.D.; Shalapy, M.; Zhao, S.F.; Liu, J.H.; Wang, J.Y. A stress-responsive transcription factor PeNAC1 regulating beta-D-glucan biosynthetic genes enhances salt tolerance in oat. Planta 2021, 254, 6. [Google Scholar] [CrossRef] [PubMed]
- Lorkovic, Z.J.; Barta, A. Genome analysis: RNA recognition motif (RRM) and K homology (KH) domain RNA-binding proteins from the flowering plant Arabidopsis thaliana. Nucleic Acids Res. 2002, 30, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, S.J.; Jang, B.S.; Jung, C.H.; Ahn, S.J.; Goh, C.H.; Cho, K.; Han, O.; Kang, H.S. Functional characterization of a glycine-rich RNA-binding protein 2 in Arabidopsis thaliana under abiotic stress conditions. Plant J. 2007, 50, 439–451. [Google Scholar] [CrossRef]
- Shim, J.S.; Park, S.H.; Lee, D.K.; Kim, Y.S.; Park, S.C.; Redillas, M.; Seo, J.S.; Kim, J.K. The rice glycine-rich protein 3 confers drought tolerance by regulating mRNA stability of ROS scavenging-related genes. Rice 2021, 14, 31. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, W.Y.; Kwak, K.J.; Oh, S.H.; Han, Y.S.; Kang, H. Glycine-rich RNA-binding proteins are functionally conserved in Arabidopsis thaliana and Oryza sativa during cold adaptation process. J. Exp. Bot. 2010, 61, 2317–2325. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.Q.; Jiang, L.; Song, S.Y.; Jing, R.; Xu, G.S. AtGRP7 is involved in the regulation of abscisic acid and stress responses in Arabidopsis. Cell. Mol. Biol. Lett. 2006, 11, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Kwak, K.J.; Kang, H.; Han, K.H.; Ahn, S.J. Molecular cloning, characterization, and stress-responsive expression of genes encoding glycine-rich RNA-binding proteins in Camelina sativa L. Plant Physiol. Biochem. 2013, 68, 44–51. [Google Scholar] [CrossRef]
- Wang, S.C.; Wang, R.C.; Liang, D.; Ma, F.W.; Shu, H.R. Molecular characterization and expression analysis of a glycine-rich RNA-binding protein gene from Malus hupehensis Rehd. Mol. Biol. Rep. 2012, 39, 4145–4153. [Google Scholar] [CrossRef] [PubMed]
- Tripet, B.P.; Mason, K.E.; Eilers, B.J.; Burns, J.; Powell, P.; Fischer, A.M.; Copie, V. Structural and biochemical analysis of the Hordeum vulgare L. HvGR-RBP1 protein, a glycine-rich RNA-binding protein involved in the regulation of barley plant development and stress response. Biochemistry 2014, 53, 7945–7960. [Google Scholar]
- Kim, Y.O.; Pan, S.; Jung, C.H.; Kang, H. A zinc finger-containing glycine-rich RNA-binding protein, atRZ-1a, has a negative impact on seed germination and seedling growth of Arabidopsis thaliana under salt or drought stress conditions. Plant Cell Physiol. 2007, 48, 1170–1181. [Google Scholar] [CrossRef]
- Kwak, K.J.; Park, S.J.; Han, J.H.; Kim, M.K.; Oh, S.H.; Han, Y.S.; Kang, H. Structural determinants crucial to the RNA chaperone activity of glycine-rich RNA-binding proteins 4 and 7 in Arabidopsis thaliana during the cold adaptation process. J. Exp. Bot. 2011, 62, 4003–4011. [Google Scholar] [CrossRef]
- Choi, M.J.; Park, Y.R.; Park, S.J.; Kang, H. Stress-responsive expression patterns and functional characterization of cold shock domain proteins in cabbage (Brassica rapa) under abiotic stress conditions. Plant Physiol. Biochem. 2015, 96, 132–140. [Google Scholar] [CrossRef]
- Kim, Y.O.; Kang, H. The role of a zinc finger-containing glycine-rich RNA-binding protein during the cold adaptation process in Arabidopsis thaliana. Plant Cell Rep. 2006, 47, 793–798. [Google Scholar] [CrossRef]
- Schmidt, F.; Marnef, A.; Cheung, M.-K.; Wilson, I.; Hancock, J.; Staiger, D.; Ladomery, M. A proteomic analysis of oligo(dT)-bound mRNP containing oxidative stress-induced Arabidopsis thaliana RNA-binding proteins ATGRP7 and ATGRP8. Mol. Biol. Rep. 2010, 37, 839–845. [Google Scholar] [CrossRef]
- Yeap, W.C.; Ooi, T.E.K.; Namasivayam, P.; Kulaveerasingam, H.; Ho, C.L. EgRBP42 encoding an hnRNP-like RNA-binding protein from Elaeis guineensis Jacq. is responsive to abiotic stresses. Plant Cell Rep. 2012, 31, 1829–1843. [Google Scholar] [CrossRef]
- Wang, B.; Wang, G.; Shen, F.; Zhu, S. A glycine-rich RNA-binding protein, CsGR-RBP3, is involved in defense responses against cold stress in Harvested Cucumber (Cucumis sativus L.) fruit. Front. Plant Sci. 2018, 9, 540. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Jung, H.J.; Lee, H.J.; Kim, K.A.; Goh, C.H.; Woo, Y.; Oh, S.H.; Han, Y.S.; Kang, H. Glycine-rich RNA-binding protein 7 affects abiotic stress responses by regulating stomata opening and closing in Arabidopsis thaliana. Plant J. 2008, 55, 455–466. [Google Scholar] [CrossRef]
- Lu, Y.; Sun, J.; Yang, Z.; Zhao, C.; Zhu, M.; Ma, D.; Dong, T.; Zhou, Z.; Liu, M.; Yang, D.; et al. Genome-wide identification and expression analysis of glycine-rich RNA-binding protein family in sweet potato wild relative Ipomoea trifida. Gene 2019, 686, 177–186. [Google Scholar] [CrossRef]
- Shinozuka, H.; Hisano, H.; Yoneyama, S.; Shimamoto, Y.; Jones, E.S.; Forster, J.W.; Yamada, T.; Kanazawa, A. Gene expression and genetic mapping analyses of a perennial ryegrass glycine-rich RNA-binding protein gene suggest a role in cold adaptation. Mol. Genet. Genom. 2006, 275, 399–408. [Google Scholar] [CrossRef]
- Xu, T.; Gu, L.; Choi, M.J.; Kim, R.J.; Suh, M.C.; Kang, H. Comparative functional analysis of wheat (Triticum aestivum) zinc finger-containing glycine-rich RNA-binding proteins in response to abiotic stresses. PLoS ONE 2014, 9, e96877. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.Y.; Kim, J.Y.; Jung, H.J.; Oh, S.H.; Han, Y.S.; Kang, H. Comparative analysis of Arabidopsis zinc finger-containing glycine-rich RNA-binding proteins during cold adaptation. Plant Physiol. Biochem. 2010, 48, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zeng, Q.C.; Lu, X.P.; Yu, D.Q.; Li, W.Z. Characterization and expression analysis of four glycine-rich RNA-binding proteins involved in osmotic response in tobacco (Nicotiana tabacum cv. Xanthi). Agric. Sci. China 2010, 9, 1577–1587. [Google Scholar] [CrossRef]
- Kim, M.K.; Jung, H.J.; Kim, D.H.; Kang, H. Characterization of glycine-rich RNA-binding proteins in Brassica napus under stress conditions. Physiol. Plant. 2012, 146, 297–307. [Google Scholar] [CrossRef]
- Ma, L.Q.; Cheng, K.; Li, J.Y.; Deng, Z.Q.; Zhang, C.J.; Zhu, H.L. Roles of plant glycine-rich RNA-binding proteins in development and stress responses. Int. J. Mol. Sci. 2021, 22, 5849. [Google Scholar] [CrossRef]
- Kim, J.S.; Park, S.J.; Kwak, K.J.; Kim, Y.O.; Kim, J.Y.; Song, J.; Jang, B.; Jung, C.H.; Kang, H. Cold shock domain proteins and glycine-rich RNA-binding proteins from Arabidopsis thaliana can promote the cold adaptation process in Escherichia coli. Nucleic Acids Res. 2007, 35, 506–516. [Google Scholar] [CrossRef]
- Kim, Y.O.; Kim, J.S.; Kang, H. Cold-inducible zinc finger-containing glycine-rich RNA-binding protein contributes to the enhancement of freezing tolerance in Arabidopsis thaliana. Plant J. 2005, 42, 890–900. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, W.Y.; Kwak, K.J.; Oh, S.H.; Han, Y.S.; Kang, H. Zinc finger-containing glycine-rich RNA-binding protein in Oryza sativa has an RNA chaperone activity under cold stress conditions. Plant Cell Environ. 2010, 33, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Sahi, C.; Agarwal, M.; Singh, A.; Grover, A. Molecular characterization of a novel isoform of rice (Oryza sativa L.) glycine rich-RNA binding protein and evidence for its involvement in high temperature stress response. Plant Sci. 2007, 173, 144–155. [Google Scholar] [CrossRef]
- Yang, D.H.; Kwak, K.J.; Kim, M.K.; Park, S.J.; Yang, K.Y.; Kang, H. Expression of Arabidopsis glycine-rich RNA-binding protein AtGRP2 or AtGRP7 improves grain yield of rice (Oryza sativa) under drought stress conditions. Plant Sci. 2014, 214, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Aneeta; Sanan-Mishra, N.; Tuteja, N.; Kumar Sopory, S. Salinity- and ABA-induced up-regulation and light-mediated modulation of mRNA encoding glycine-rich RNA-binding protein from Sorghum bicolor. Biochem. Bioph. Res. Commun. 2002, 296, 1063–1068. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Amaro, M.A.; Rodriguez-Hernandez, A.A.; Rodriguez-Kessler, M.; Hernandez-Lucero, E.; Rosales-Mendoza, S.; Ibanez-Salazar, A.; Delgado-Sanchez, P.; Jimenez-Bremont, J.F. Overexpression of AtGRDP2, a novel glycine-rich domain protein, accelerates plant growth and improves stress tolerance. Front. Plant Sci. 2014, 5, 782. [Google Scholar] [CrossRef] [PubMed]
- Long, R.C.; Yang, Q.C.; Kang, J.M.; Zhang, T.J.; Wang, H.M.; Li, M.N.; Zhang, Z. Overexpression of a novel salt stress-induced glycine-rich protein gene from alfalfa causes salt and ABA sensitivity in Arabidopsis. Plant Cell Rep. 2013, 32, 1289–1298. [Google Scholar] [CrossRef] [PubMed]
- Teng, K.; Tan, P.H.; Xiao, G.Z.; Han, L.B.; Chang, Z.H.; Chao, Y.H. Heterologous expression of a novel Zoysia japonica salt-induced glycine-rich RNA-binding protein gene, ZjGRP, caused salt sensitivity in Arabidopsis. Plant Cell Rep. 2017, 36, 179–191. [Google Scholar] [CrossRef]
- Kwak, K.J.; Kim, Y.O.; Kang, H. Characterization of transgenic Arabidopsis plants overexpressing GR-RBP4 under high salinity, dehydration, or cold stress. J. Exp. Bot. 2005, 56, 3007–3016. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, D.W.; Wang, Y.C.; Zheng, L.; Yang, C.P. A glycine-rich RNA-binding protein can mediate physiological responses in transgenic plants under salt stress. Mol. Biol. Rep. 2012, 39, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.X.; Qin, Y.; Li, Y.L.; Li, M.J.; Ma, F.W. Overexpression of MpGR-RBP1, a glycine-rich RNA-binding protein gene from Malus prunifolia (Willd.) Borkh., confers salt stress tolerance and protects against oxidative stress in Arabidopsis. Plant Cell Tissue Organ. Cult. 2014, 119, 635–646. [Google Scholar] [CrossRef]
- Farmer, E.E.; Mueller, M.J. ROS-mediated lipid peroxidation and RES-activated signaling. Annu. Rev. Plant Biol. 2013, 64, 429–450. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Kwak, K.J.; Oh, T.R.; Kim, Y.O.; Kang, H. Cold shock domain proteins affect seed germination and growth of Arabidopsis thaliana under abiotic stress conditions. Plant Cell Physiol. 2009, 50, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.G.; Chen, S.L.; Zhou, X.Y.; Shen, X.; Deng, L.; Zhu, H.J.; Shao, J.; Shi, Y.; Dai, S.X.; Fritz, E.; et al. Ionic homeostasis and reactive oxygen species control in leaves and xylem sap of two poplars subjected to NaCl stress. Tree Physiol. 2008, 28, 947–957. [Google Scholar] [CrossRef]
- Wang, R.G.; Chen, S.L.; Deng, L.; Fritz, E.; Hüttermann, A.; Polle, A. Leaf photosynthesis, fluorescence response to salinity and the relevance to chloroplast salt compartmentation and anti-oxidative stress in two poplars. Trees 2007, 21, 581–591. [Google Scholar] [CrossRef]
- Sun, J.; Dai, S.X.; Wang, R.G.; Chen, S.L.; Li, N.Y.; Zhou, X.Y.; Lu, C.F.; Shen, X.; Zheng, X.J.; Hu, Z.M.; et al. Calcium mediates root K+/Na+ homeostasis in poplar species differing in salt tolerance. Tree Physiol. 2009, 29, 1175–1186. [Google Scholar] [CrossRef]
- Ma, X.Y.; Deng, L.; Li, J.K.; Zhou, X.Y.; Li, N.Y.; Zhang, D.C.; Lu, Y.J.; Wang, R.G.; Sun, J.A.; Lu, C.F.; et al. Effect of NaCl on leaf H+-ATPase and the relevance to salt tolerance in two contrasting poplar species. Trees-Struct. Funct. 2010, 24, 597–607. [Google Scholar] [CrossRef]
- Li, N.Y.; Chen, S.L.; Zhou, X.Y.; Li, C.Y.; Shao, J.; Wang, R.G.; Fritz, E.; Hüttermann, A.; Polle, A. Effect of NaCl on photosynthesis, salt accumulation and ion compartmentation in two mangrove species, Kandelia candel and Bruguiera gymnorhiza. Aquat. Bot. 2008, 88, 303–310. [Google Scholar] [CrossRef]
- Li, N.Y.; Zhou, X.Y.; Wang, R.G.; Li, J.K.; Lu, C.F.; Chen, S.L. Salt compartmentation and antioxidant defense in roots and leaves of two non-salt secretor mangroves under salt stress. In Mangrove Ecosystem Ecology and Function; Sharma, S., Ed.; InTechOpen: London, UK, 2018; pp. 81–104. [Google Scholar]
- Leple, J.C.; Brasileiro, A.C.; Michel, M.F.; Delmotte, F.; Jouanin, L. Transgenic poplars: Expression of chimeric genes using four different constructs. Plant Cell Rep. 1992, 11, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Gafur, A.; Schützendübel, A.; Langenfeld-Heyser, R.; Fritz, E.; Polle, A. Compatible and incompetent Paxillus involutus isolates for ectomycorrhiza formation in vitro with poplar (Populus × canescens) differ in H2O2 production. Plant Biol. 2004, 6, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.F.; Zhou, Y.Q.; Chen, Y.R.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Shen, Z.D.; Sun, J.; Yao, J.; Wang, S.J.; Ding, M.Q.; Zhang, H.L.; Qian, Z.Y.; Zhao, N.; Sa, G.; Zhao, R.; et al. High rates of virus-induced gene silencing by tobacco rattle virus in Populus. Tree Physiol. 2015, 35, 1016–1029. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number | Description | Abbr. | Gene ID | Log2 (Fold_Change) |
|---|---|---|---|---|
| Photosynthetic proteins | ||||
| XP_002319934.1 | cytochrome b6-f complex iron-sulfur subunit, chloroplastic | PETC | Potri.013G148900 | 4.65 |
| XP_002317015.1 | photosystem II 10 kDa polypeptide, chloroplastic | PSBR | Potri.011G142200 | 4.96 |
| XP_002315645.1 | photosystem II core complex protein psbY, chloroplastic | PSBY | Potri.010G052000 | 1.92 |
| XP_002303109.2 | photosystem II reaction center PSB28 protein, chloroplastic | PSB28 | Potri.002G256400 | 2.32 |
| XP_002301582.1 | chlorophyll a/b-binding protein 5, chloroplastic | CAB5 | Potri.002G221400 | 3.45 |
| XP_002315298.1 | chlorophyll a/b-binding protein 6, chloroplastic | CAB6 | Potri.010G221100 | 3.48 |
| XP_024441106.1 | ribulose-1, 5 bisphosphate carboxylase/oxygenase large subunit N-methyltransferase, chloroplastic | RBCMT | Potri.014G169300 | 1.57 |
| XP_006368947.2 | light-harvesting complex-like protein 3 isotype 1, chloroplastic | LIL3-1 | Potri.001G151300 | 1.24 |
| XP_002310188.1 | oxygen-evolving enhancer protein 1, chloroplastic | PSBO1 | Potri.007G033700 | 2.91 |
| XP_002300858.1 | oxygen-evolving enhancer protein 2, chloroplastic | PSBO2 | Potri.002G055700 | 2.69 |
| XP_006383096.1 | ferredoxin-NADP reductase, leaf isozyme, chloroplastic | LFNR | Potri.005G112900 | 2.70 |
| Antioxidant enzymes | ||||
| XP_002304909.1 | peroxidase 42 | POD42 | Potri.004G015300 | 2.25 |
| XP_024448088.1 | peroxidase 47 | POD47 | Potri.018G136900 | 2.77 |
| XP_002325843.1 | superoxide dismutase [Cu-Zn] 2 | SOD[Cu-Zn]2 | Potri.019G035800 | 2.04 |
| XP_002319332.2 | superoxide dismutase [Mn], mitochondrial | SOD[Mn] | Potri.013G092600 | 1.00 |
| XP_024438027.1 | transmembrane ascorbate ferrireductase 1 isoform X2 | CYB1-2 | Potri.012G141000 | 5.76 |
| XP_002310066.2 | thioredoxin X, chloroplastic | TRX | Potri.007G074000 | 1.02 |
| XP_002307752.2 | thioredoxin-like protein CDSP32, chloroplastic | CDSP32 | Potri.005G245700 | 2.85 |
| XP_002306676.1 | thioredoxin M-type, chloroplastic | TRXM | Potri.005G186800 | 4.54 |
| XP_002308370.2 | peroxiredoxin Q, chloroplastic | PRXQ | Potri.006G137500 | 2.66 |
| Cation/H+ exchangers and ATPases | ||||
| XP_002298746.1 | sodium/proton antiporter 1 | NHA1 | Potri.001G301000 | 2.49 |
| XP_002307194.2 | sodium/hydrogen exchanger 2 isoform X1 | NHE2-1 | Potri.005G045100 | 2.95 |
| XP_002323578.2 | vacuolar cation/proton exchanger 3 | CAX3 | Potri.016G115500 | 2.73 |
| XP_006382405.2 | pyrophosphate-energized vacuolar membrane proton pump 1 | AVP1 | Potri.005G018700 | 1.43 |
| XP_024438330.1 | ATPase 11, plasma membrane-type | AHA11 | Potri.012G071600 | 1.01 |
| XP_024459503.1 | calcium-transporting ATPase 8, plasma membrane-type | ACA8 | Potri.018G139800 | 2.02 |
| XP_024466795.1 | calcium-transporting ATPase 9, plasma membrane-type | ACA9 | Potri.010G250800 | 1.66 |
| XP_024461157.1 | AAA-ATPase At1g43910 | AATP | Potri.007G019600 | 1.10 |
| XP_024455649.1 | AAA-ATPase ASD, mitochondrial | ASD | Potri.004G012500 | 1.94 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Zhao, R.; Liu, J.; Yao, J.; Ma, S.; Yin, K.; Zhang, Y.; Liu, Z.; Yan, C.; Zhao, N.; et al. Populus euphratica GRP2 Interacts with Target mRNAs to Negatively Regulate Salt Tolerance by Interfering with Photosynthesis, Na+, and ROS Homeostasis. Int. J. Mol. Sci. 2024, 25, 2046. https://doi.org/10.3390/ijms25042046
Li J, Zhao R, Liu J, Yao J, Ma S, Yin K, Zhang Y, Liu Z, Yan C, Zhao N, et al. Populus euphratica GRP2 Interacts with Target mRNAs to Negatively Regulate Salt Tolerance by Interfering with Photosynthesis, Na+, and ROS Homeostasis. International Journal of Molecular Sciences. 2024; 25(4):2046. https://doi.org/10.3390/ijms25042046
Chicago/Turabian StyleLi, Jing, Rui Zhao, Jian Liu, Jun Yao, Siyuan Ma, Kexin Yin, Ying Zhang, Zhe Liu, Caixia Yan, Nan Zhao, and et al. 2024. "Populus euphratica GRP2 Interacts with Target mRNAs to Negatively Regulate Salt Tolerance by Interfering with Photosynthesis, Na+, and ROS Homeostasis" International Journal of Molecular Sciences 25, no. 4: 2046. https://doi.org/10.3390/ijms25042046
APA StyleLi, J., Zhao, R., Liu, J., Yao, J., Ma, S., Yin, K., Zhang, Y., Liu, Z., Yan, C., Zhao, N., Zhou, X., & Chen, S. (2024). Populus euphratica GRP2 Interacts with Target mRNAs to Negatively Regulate Salt Tolerance by Interfering with Photosynthesis, Na+, and ROS Homeostasis. International Journal of Molecular Sciences, 25(4), 2046. https://doi.org/10.3390/ijms25042046