
Decreased Expression of Placental Proteins in Recurrent Pregnancy Loss: Functional Relevance and Diagnostic Value

 , , , , , , ,
, , , , , , ,  , and
, and 
Abstract
1. Introduction
2. Results
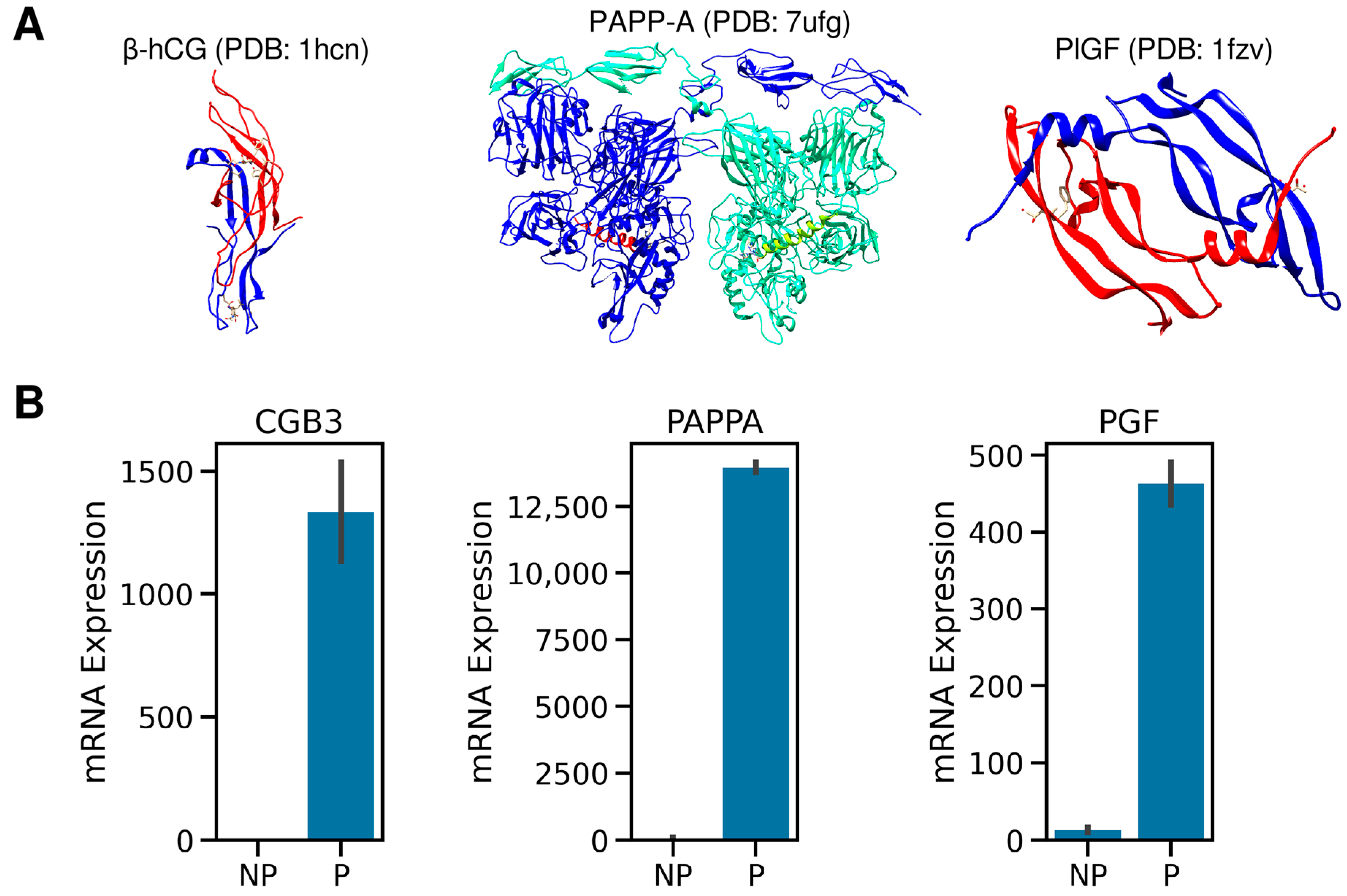
2.1. The Expression Patterns of hCG, PlGF, and PAPPA Proteins
2.2. Demographic and Clinical Data
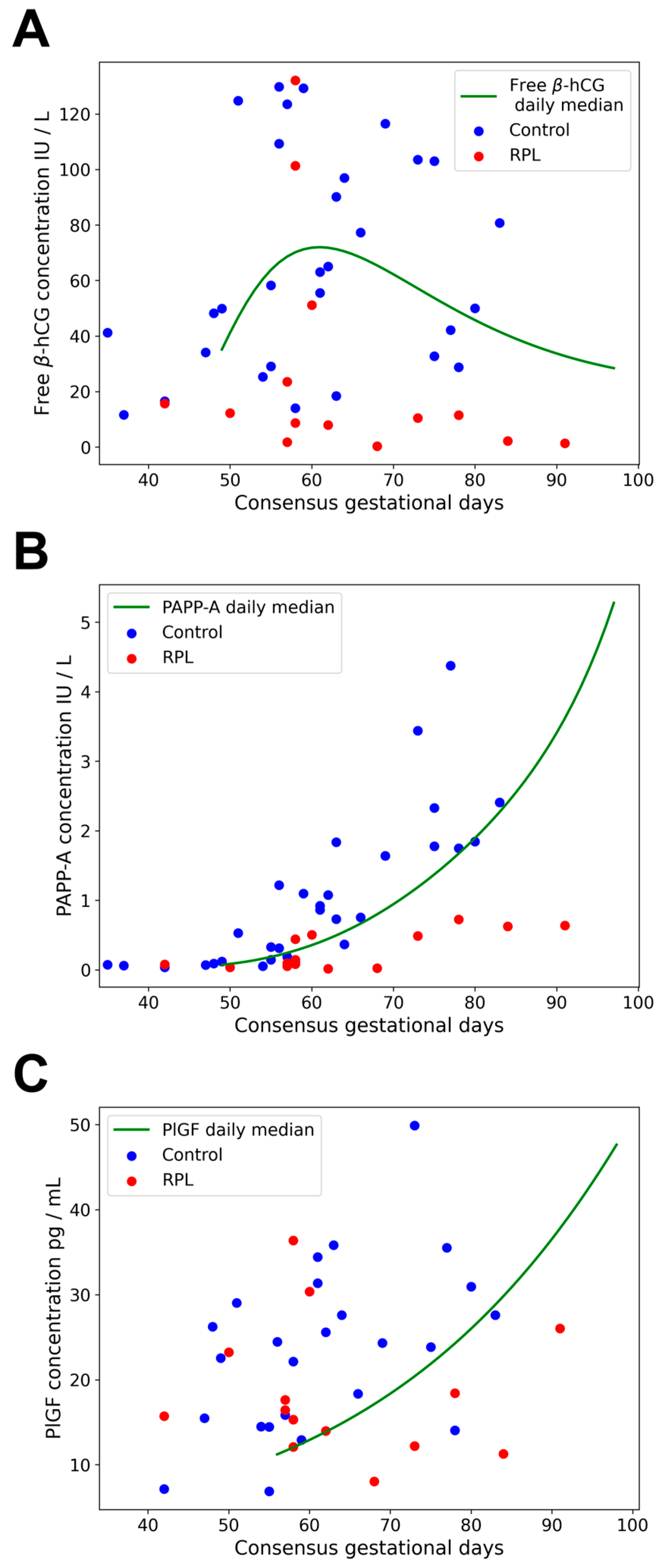
2.3. Gestational Age-Specific Distribution of Data
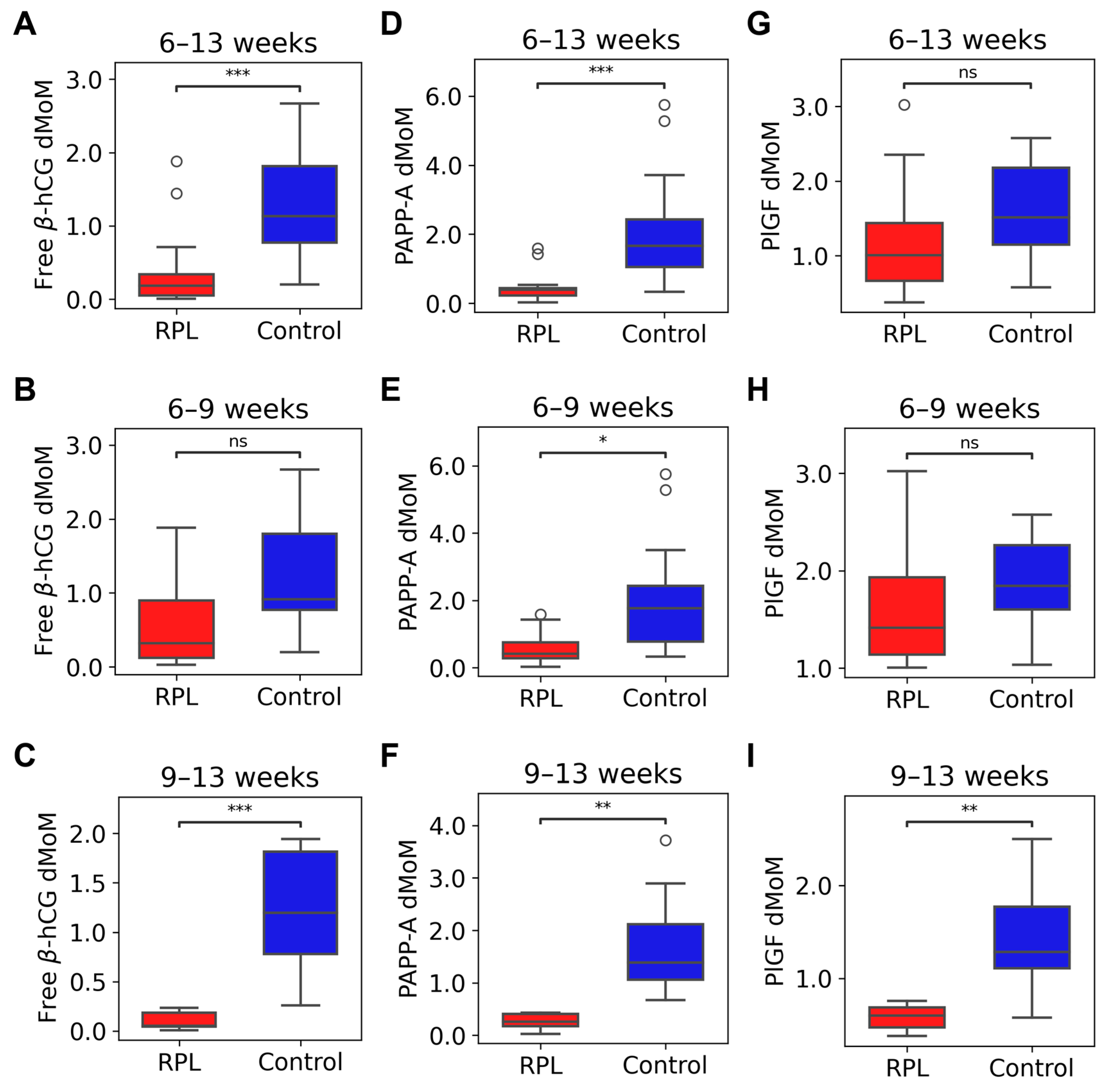
2.4. Maternal Serum Concentrations of Free β-hCG, PAPP-A, and PlGF in RPL and controls
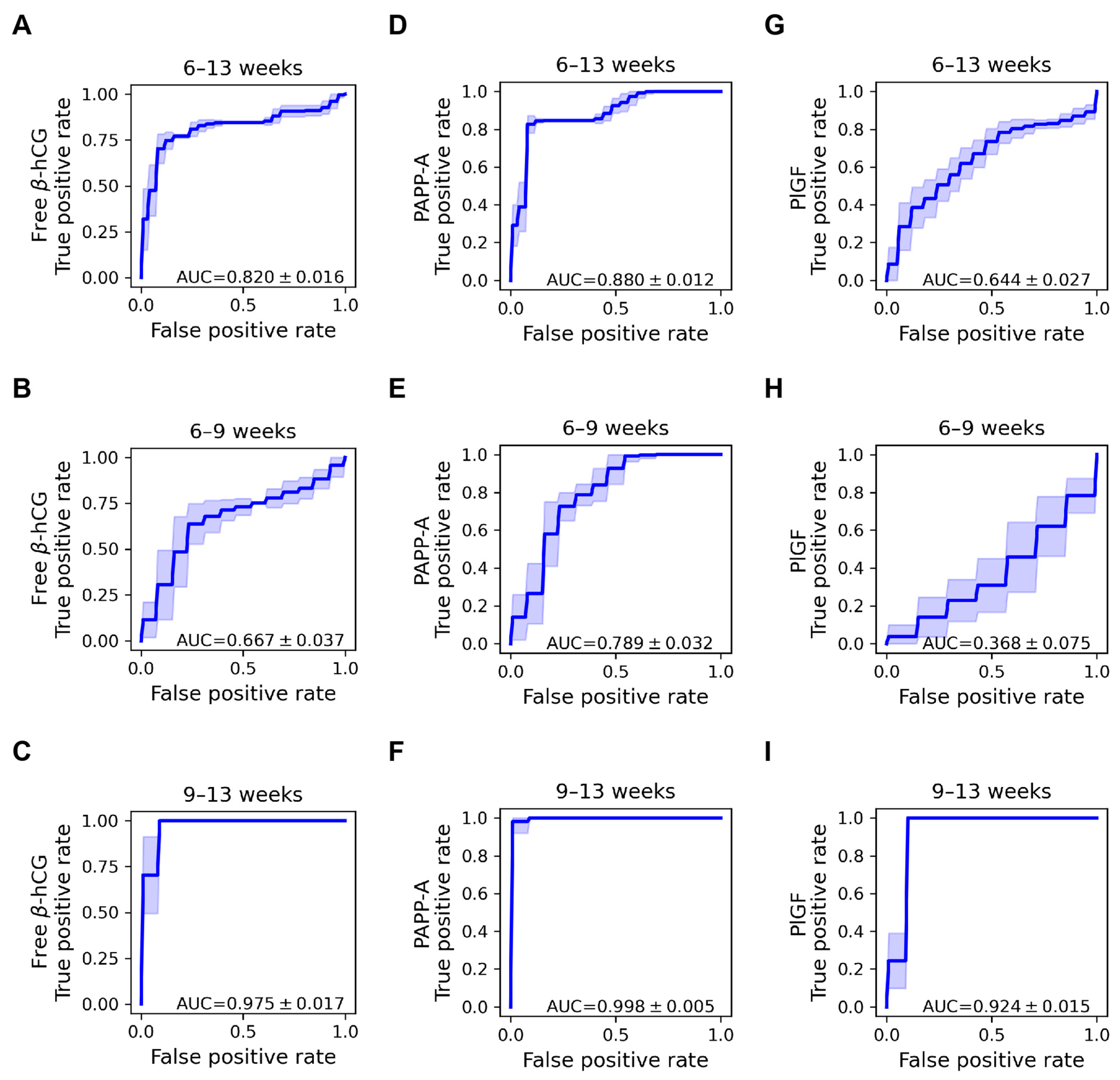
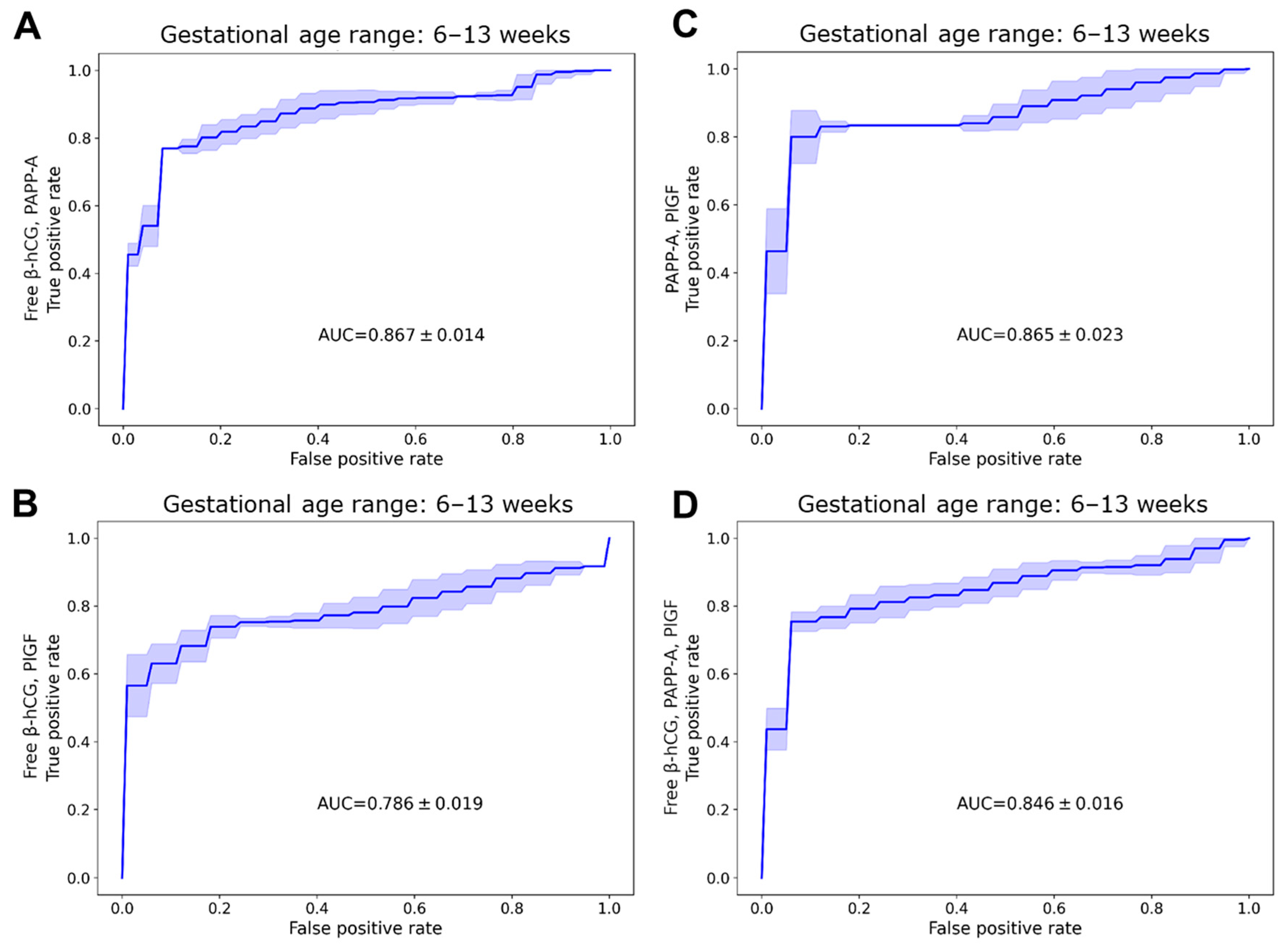
2.5. Discriminative Properties of Biomarker Proteins
3. Discussion
3.1. Principal Findings of the Study
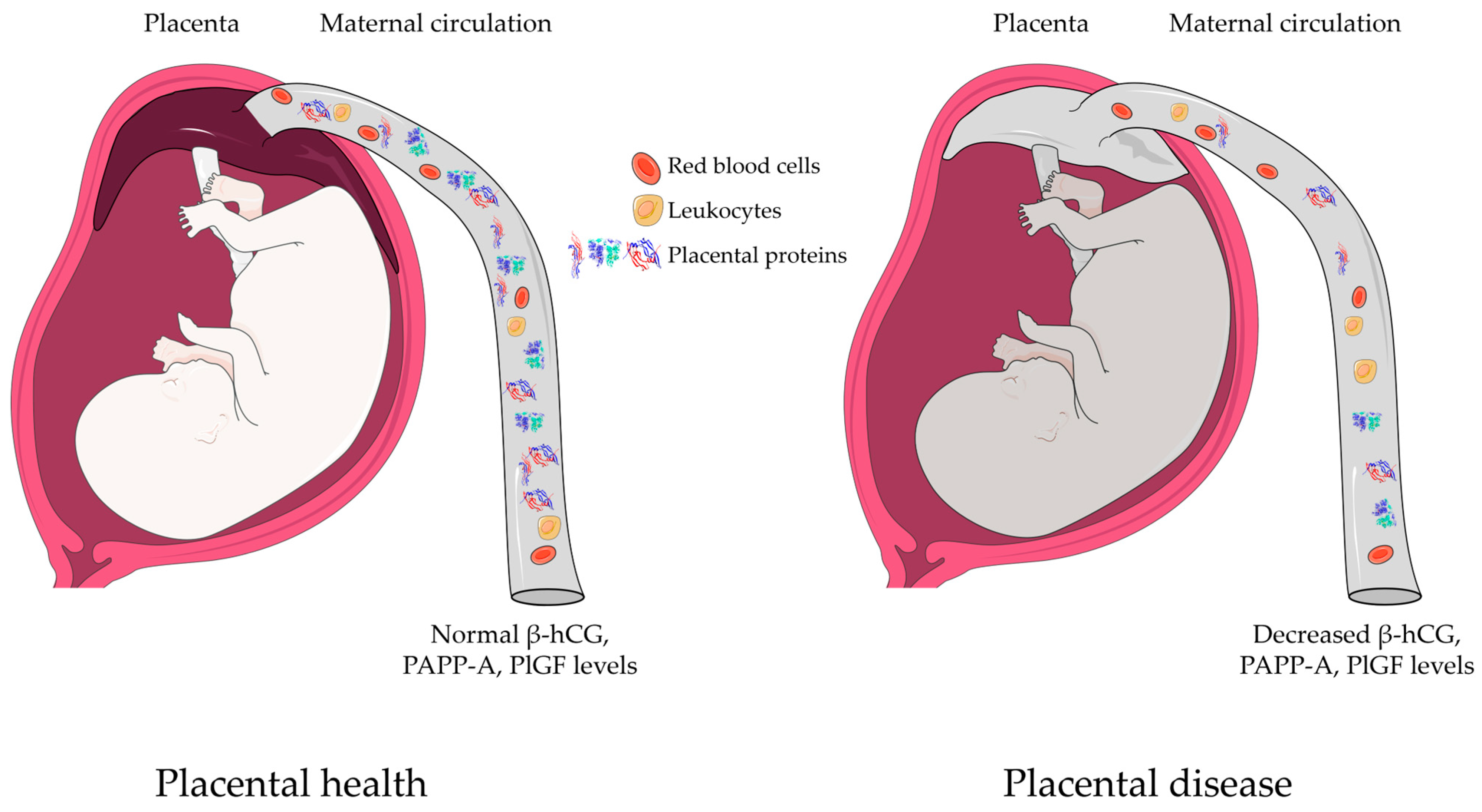
3.2. Placenta-Specific Proteins
3.3. Biomarker Proteins in Miscarriage and RPL
3.4. Concentration Changes of Biomarker Proteins in RPL
3.5. Strengths and Limitations of the Study
3.6. Implications and Future Directions
4. Materials and Methods
4.1. Study Groups, Clinical Definitions, and Sample Collection
4.2. Immunoassays
4.3. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dugas, C.; Slane, V.H. Miscarriage. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2020. [Google Scholar]
- WHO. Recommended definitions, terminology and format for statistical tables related to the perinatal period and use of a new certificate for cause of perinatal deaths. Modifications recommended by FIGO as amended 14 October 1976. Acta Obstet. Gynecol. Scand. 1977, 56, 247–253. [Google Scholar]
- Ammon Avalos, L.; Galindo, C.; Li, D.K. A systematic review to calculate background miscarriage rates using life table analysis. Birth Defects Res. Part A Clin. Mol. Teratol. 2012, 94, 417–423. [Google Scholar] [CrossRef]
- Larsen, E.C.; Christiansen, O.B.; Kolte, A.M.; Macklon, N. New insights into mechanisms behind miscarriage. BMC Med. 2013, 11, 154. [Google Scholar] [CrossRef]
- Savitz, D.A.; Hertz-Picciotto, I.; Poole, C.; Olshan, A.F. Epidemiologic measures of the course and outcome of pregnancy. Epidemiol. Rev. 2002, 24, 91–101. [Google Scholar] [CrossRef]
- Ford, H.B.; Schust, D.J. Recurrent pregnancy loss: Etiology, diagnosis, and therapy. Rev. Obstet. Gynecol. 2009, 2, 76–83. [Google Scholar]
- Zinaman, M.J.; Clegg, E.D.; Brown, C.C.; O’Connor, J.; Selevan, S.G. Estimates of human fertility and pregnancy loss. Fertil. Steril. 1996, 65, 503–509. [Google Scholar] [CrossRef]
- Ellish, N.J.; Saboda, K.; O’Connor, J.; Nasca, P.C.; Stanek, E.J.; Boyle, C. A prospective study of early pregnancy loss. Hum. Reprod. 1996, 11, 406–412. [Google Scholar] [CrossRef]
- Wilcox, A.J.; Weinberg, C.R.; O’Connor, J.F.; Baird, D.D.; Schlatterer, J.P.; Canfield, R.E.; Armstrong, E.G.; Nisula, B.C. Incidence of Early Loss of Pregnancy. N. Engl. J. Med. 1988, 319, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Rai, R.; Regan, L. Recurrent miscarriage. Lancet 2006, 368, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Chard, T. 11 Frequency of implantation and early pregnancy loss in natural cycles. Baillière’s Clin. Obstet. Gynaecol. 1991, 5, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Macklon, N.S.; Geraedts, J.P.; Fauser, B.C. Conception to ongoing pregnancy: The ‘black box’ of early pregnancy loss. Hum. Reprod. Update 2002, 8, 333–343. [Google Scholar] [CrossRef]
- Brezina, P.R.; Kutteh, W.H. Classic and cutting-edge strategies for the management of early pregnancy loss. Obstet. Gynecol. Clin. N. Am. 2014, 41, 1–18. [Google Scholar] [CrossRef]
- Regan, L.; Braude, P.R.; Trembath, P.L. Influence of past reproductive performance on risk of spontaneous abortion. Br. Med. J. 1989, 299, 541. [Google Scholar] [CrossRef]
- Knudsen, U.B.; Hansen, V.; Juul, S.; Secher, N.J. Prognosis of a new pregnancy following previous spontaneous abortions. Eur. J. Obstet. Gynecol. Reprod. Biol. 1991, 39, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Risch, H.A.; Weiss, N.S.; Aileen, C.E.; Miller, A.B. Risk factors for spontaneous abortion and its recurrence. Am. J. Epidemiol. 1988, 128, 420–430. [Google Scholar] [CrossRef] [PubMed]
- Clifford, K.; Rai, R.; Watson, H.; Regan, L. An informative protocol for the investigation of recurrent miscarriage: Preliminary experience of 500 consecutive cases. Hum. Reprod. 1994, 9, 1328–1332. [Google Scholar] [CrossRef] [PubMed]
- Stirrat, G.M. Recurrent miscarriage. Lancet 1990, 336, 673–675. [Google Scholar] [CrossRef] [PubMed]
- Li, T.C.; Makris, M.; Tomsu, M.; Tuckerman, E.; Laird, S. Recurrent miscarriage: Aetiology, management and prognosis. Hum. Reprod. Update 2002, 8, 463–481. [Google Scholar] [CrossRef]
- Vaiman, D. Genetic regulation of recurrent spontaneous abortion in humans. Biomed. J. 2015, 38, 11–24. [Google Scholar] [CrossRef]
- ACOG. ACOG practice bulletin. Management of recurrent pregnancy loss. Number 24, February 2001. (Replaces Technical Bulletin Number 212, September 1995). American College of Obstetricians and Gynecologists. Int. J. Gynecol. Obstet. 2002, 78, 179–190. [Google Scholar] [CrossRef]
- American Society for Reproductive Medicine (ASRM). Evaluation and treatment of recurrent pregnancy loss: A committee opinion. Fertil. Steril. 2012, 98, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Van den Akker, O.B. The psychological and social consequences of miscarriage. Expert Rev. Obstet. Gynecol. 2011, 6, 295–304. [Google Scholar] [CrossRef]
- Murphy, S.; Cacciatore, J. The psychological, social, and economic impact of stillbirth on families. Semin. Fetal Neonatal Med. 2017, 22, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Giorlandino, C.; Calugi, G.; Iaconianni, L.; Santoro, M.L.; Lippa, A. Spermatozoa with chromosomal abnormalities may result in a higher rate of recurrent abortion. Fertil. Steril. 1998, 70, 576–577. [Google Scholar] [CrossRef] [PubMed]
- Rubio, C.; Simón, C.; Vidal, F.; Rodrigo, L.; Pehlivan, T.; Remohí, J.; Pellicer, A. Chromosomal abnormalities and embryo development in recurrent miscarriage couples. Hum. Reprod. 2003, 18, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Stirrat, G.M. Recurrent miscarriage II: Clinical associations, causes, and management. Lancet 1990, 336, 728–733. [Google Scholar] [CrossRef] [PubMed]
- Tulppala, M.; Palosuo, T.; Ramsay, T.; Miettinen, A.; Salonen, R.; Ylikorkala, O. A prospective study of 63 couples with a history of recurrent spontaneous abortion: Contributing factors and outcome of subsequent pregnancies. Hum. Reprod. 1993, 8, 764–770. [Google Scholar] [CrossRef]
- Uehara, S.; Hashiyada, M.; Sato, K.; Sato, Y.; Fujimori, K.; Okamura, K. Preferential X-chromosome inactivation in women with idiopathic recurrent pregnancy loss. Fertil. Steril. 2001, 76, 908–914. [Google Scholar] [CrossRef]
- Aldrich, C.L.; Stephenson, M.D.; Karrison, T.; Odem, R.R.; Branch, D.W.; Scott, J.R.; Schreiber, J.R.; Ober, C. HLA-G genotypes and pregnancy outcome in couples with unexplained recurrent miscarriage. Mol. Hum. Reprod. 2001, 7, 1167–1172. [Google Scholar] [CrossRef][Green Version]
- Hirahara, F.; Andoh, N.; Sawai, K.; Hirabuki, T.; Uemura, T.; Minaguchi, H. Hyperprolactinemic recurrent miscarriage and results of randomized bromocriptine treatment trials. Fertil. Steril. 1998, 70, 246–252. [Google Scholar] [CrossRef]
- Garzia, E.; Borgato, S.; Cozzi, V.; Doi, P.; Bulfamante, G.; Persani, L.; Cetin, I. Lack of expression of endometrial prolactin in early implantation failure: A pilot study. Hum. Reprod. 2004, 19, 1911–1916. [Google Scholar] [CrossRef][Green Version]
- Craig, L.B.; Ke, R.W.; Kutteh, W.H. Increased prevalence of insulin resistance in women with a history of recurrent pregnancy loss. Fertil. Steril. 2002, 78, 487–490. [Google Scholar] [CrossRef] [PubMed]
- Li, T.C.; Spuijbroek, M.D.E.H.; Tuckerman, E.; Anstie, B.; Loxley, M.; Laird, S. Endocrinological and endometrial factors in recurrent miscarriage. Br. J. Obstet. Gynaecol. 2000, 107, 1471–1479. [Google Scholar] [CrossRef] [PubMed]
- Bussen, S.; Sütterlin, M.; Steck, T. Endocrine abnormalities during the follicular phase in women with recurrent spontaneous abortion. Hum. Reprod. 1999, 14, 18–20. [Google Scholar] [CrossRef]
- Grimbizis, G.F.; Camus, M.; Tarlatzis, B.C.; Bontis, J.N.; Devroey, P. Clinical implications of uterine malformations and hysteroscopic treatment results. Hum. Reprod. Update 2001, 7, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Salim, R.; Regan, L.; Woelfer, B.; Backos, M.; Jurkovic, D. A comparative study of the morphology of congenital uterine anomalies in women with and without a history of recurrent first trimester miscarriage. Hum. Reprod. 2003, 18, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Homer, H.A.; Li, T.-C.; Cooke, I.D. The septate uterus: A review of management and reproductive outcome. Fertil. Steril. 2000, 73, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Clifford, K.; Flanagan, A.M.; Regan, L. Endometrial CD56+ natural killer cells in women with recurrent miscarriage: A histomorphometric study. Hum. Reprod. 1999, 14, 2727–2730. [Google Scholar] [CrossRef] [PubMed]
- Quenby, S.; Bates, M.; Doig, T.; Brewster, J.; Lewis-Jones, D.I.; Johnson, P.M.; Vince, G. Pre-implantation endometrial leukocytes in women with recurrent miscarriage. Hum. Reprod. 1999, 14, 2386–2391. [Google Scholar] [CrossRef]
- Lachapelle, M.H.; Miron, P.; Hemmings, R.; Roy, D.C. Endometrial T, B, and NK cells in patients with recurrent spontaneous abortion. Altered profile and pregnancy outcome. J. Immunol. 1996, 156, 4027. [Google Scholar] [CrossRef]
- Eblen, A.C.; Gercel-Taylor, C.; Shields, L.B.E.; Sanfilippo, J.S.; Nakajima, S.T.; Taylor, D.D. Alterations in humoral immune responses associated with recurrent pregnancy loss. Fertil. Steril. 2000, 73, 305–313. [Google Scholar] [CrossRef]
- Wilson, R.; Maclean, M.A.; Jenkins, C.; Kinnane, D.; Mooney, J.; Walker, J.J. Abnormal immunoglobulin subclass patterns in women with a history of recurrent miscarriage. Fertil. Steril. 2001, 76, 915–917. [Google Scholar] [CrossRef] [PubMed]
- Makhseed, M.; Raghupathy, R.; Azizieh, F.; Omu, A.; Al-Shamali, E.; Ashkanani, L. Th1 and Th2 cytokine profiles in recurrent aborters with successful pregnancy and with subsequent abortions. Hum. Reprod. 2001, 16, 2219–2226. [Google Scholar] [CrossRef] [PubMed]
- Quack, K.C.; Vassiliadou, N.; Pudney, J.; Anderson, D.J.; Hill, J.A. Leukocyte activation in the decidua of chromosomally normal and abnormal fetuses from women with recurrent abortion. Hum. Reprod. 2001, 16, 949–955. [Google Scholar] [CrossRef]
- Kovalevsky, G.; Gracia, C.R.; Berlin, J.A.; Sammel, M.D.; Barnhart, K.T. Evaluation of the Association Between Hereditary Thrombophilias and Recurrent Pregnancy Loss: A Meta-analysis. Arch. Intern. Med. 2004, 164, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Rey, E.; Kahn, S.R.; David, M.; Shrier, I. Thrombophilic disorders and fetal loss: A meta-analysis. Lancet 2003, 361, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Rai, R.; Backos, M.; Elgaddal, S.; Shlebak, A.; Regan, L. Factor V Leiden and recurrent miscarriage—Prospective outcome of untreated pregnancies. Hum. Reprod. 2002, 17, 442–445. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rai, R.; Shlebak, A.; Cohen, H.; Backos, M.; Holmes, Z.; Marriott, K.; Regan, L. Factor V Leiden and acquired activated protein C resistance among 1000 women with recurrent miscarriage. Hum. Reprod. 2001, 16, 961–965. [Google Scholar] [CrossRef]
- Rai, R.S.; Clifford, K.; Cohen, H.; Regan, L. High prospective fetal loss rate in untreated pregnancies of women with recurrent miscarriage and antiphospholipid antibodies. Hum. Reprod. 1995, 10, 3301–3304. [Google Scholar] [CrossRef]
- Rai, R.S.; Regan, L.; Clifford, K.; Pickering, W.; Dave, M.; Mackie, I.; McNally, T.; Cohen, H. Immunology: Antiphospholipid antibodies and β2-glycoprotein-I in 500 women with recurrent miscarriage: Results of a comprehensive screening approach. Hum. Reprod. 1995, 10, 2001–2005. [Google Scholar] [CrossRef]
- Greaves, M.; Cohen, H.; Machin, S.J.; Mackie, I. Guidelines on the Investigation and Management of the Antiphospholipid Syndrome. Br. J. Haematol. 2000, 109, 704–715. [Google Scholar] [CrossRef]
- Christiansen, O.B.; Steffensen, R.; Nielsen, H.S.; Varming, K. Multifactorial Etiology of Recurrent Miscarriage and Its Scientific and Clinical Implications. Gynecol. Obstet. Investig. 2008, 66, 257–267. [Google Scholar] [CrossRef]
- Trout, S.W.; Seifer, D.B. Do women with unexplained recurrent pregnancy loss have higher day 3 serum FSH and estradiol values? Fertil. Steril. 2000, 74, 335–337. [Google Scholar] [CrossRef]
- Gürbüz, B.; Yalti, S.; Ozden, S.; Ficicioglu, C. High basal estradiol level and FSH/LH ratio in unexplained recurrent pregnancy loss. Arch. Gynecol. Obstet. 2004, 270, 37–39. [Google Scholar] [CrossRef]
- Pils, S.; Promberger, R.; Springer, S.; Joura, E.; Ott, J. Decreased Ovarian Reserve Predicts Inexplicability of Recurrent Miscarriage? A Retrospective Analysis. PLoS ONE 2016, 11, e0161606. [Google Scholar] [CrossRef] [PubMed]
- Okon, M.A.; Laird, S.M.; Tuckerman, E.M.; Li, T.-C. Serum Androgen Levels in Women who Have Recurrent Miscarriages and their Correlation with Markers of Endometrial Function. Fertil. Steril. 1998, 69, 682–690. [Google Scholar] [CrossRef]
- Kim, M.-S.; Gu, B.-H.; Song, S.; Choi, B.-C.; Cha, D.-H.; Baek, K.-H. ITI-H4, as a biomarker in the serum of recurrent pregnancy loss (RPL) patients. Mol. BioSystems 2011, 7, 1430–1440. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, M.; Kajiura, S.; Katano, K.; Aoyama, T.; Aoki, K. Are serum progesterone levels predictive of recurrent miscarriage in future pregnancies? Fertil. Steril. 1997, 68, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; He, J.; Guo, C.; Zhang, Y.; Yang, W.; Xin, M.; Liang, X.; Yin, X.; Wang, J.; Liu, Y. Serum biomarker analysis in patients with recurrent spontaneous abortion. Mol. Med. Rep. 2017, 16, 2367–2378. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; He, L.; Yang, C.-Y.; Ye, Q. iTRAQ and PRM-based quantitative proteomics in early recurrent spontaneous abortion: Biomarkers discovery. Clin. Proteom. 2019, 16, 36. [Google Scholar] [CrossRef] [PubMed]
- Al-Azemi, M.; Ledger, W.L.; Diejomaoh, M.; Mousa, M.; Makhseed, M.; Omu, A. Measurement of inhibin A and inhibin pro-alphaC in early human pregnancy and their role in the prediction of pregnancy outcome in patients with recurrent pregnancy loss. Fertil. Steril. 2003, 80, 1473–1479. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Li, X.; Jiao, X.; Zhang, R.; Zhang, J. Predictive value of serum β-hCG for early pregnancy outcomes among women with recurrent spontaneous abortion. Int. J. Gynecol. Obstet. 2016, 135, 16–21. [Google Scholar] [CrossRef]
- Muttukrishna, S.; Jauniaux, E.; Greenwold, N.; McGarrigle, H.; Jivraj, S.; Carter, S.; Elgaddal, S.; Groome, N.; Regan, L. Circulating levels of inhibin A, activin A and follistatin in missed and recurrent miscarriages. Hum. Reprod. 2002, 17, 3072–3078. [Google Scholar] [CrossRef]
- Rull, K.; Laan, M. Expression of β-subunit of HCG genes during normal and failed pregnancy. Hum. Reprod. 2005, 20, 3360–3368. [Google Scholar] [CrossRef] [PubMed]
- Darwish, A.; Ghorab, N.; El-Ashmawy, M.H.; Kamal, M.M.; Soliman, M.S.A. Biochemical Markers for Prediction of Pregnancy Outcome in Cases of Recurrent Pregnancy Loss; Middle East Fertility Society: Beirut, Lebanon, 2005. [Google Scholar]
- Heinig, J.; Steinhard, J.; Schmitz, R.; Nofer, J.R.; Kiesel, L.; Klockenbusch, W. Maternal serum free ß-hCG and PAPP-A in patients with habitual abortion-influence on first-trimester screening for chromosomal abnormalities. Prenat. Diagn. 2007, 27, 814–816. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Mostafa, M.H.; Mann, K.; Schindler, A.E.; Hoermann, R. Human chorionic gonadotropin exhibits normal biological activity in patients with recurrent pregnancy loss. Gynecol. Endocrinol. 2002, 16, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Prakash, A.; Laird, S.; Tuckerman, E.; Li, T.C.; Ledger, W.L. Inhibin A and activin A may be used to predict pregnancy outcome in women with recurrent miscarriage. Fertil. Steril. 2005, 83, 1758–1763. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.H.; Shuai, W.; Tong, J.; Wang, L.; Chen, P.; Duan, T. Increased Dickkopf-1 expression in patients with unexplained recurrent spontaneous miscarriage. Clin. Exp. Immunol. 2013, 172, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Rull, K.; Tomberg, K.; Kõks, S.; Männik, J.; Möls, M.; Sirotkina, M.; Värv, S.; Laan, M. Increased placental expression and maternal serum levels of apoptosis-inducing TRAIL in recurrent miscarriage. Placenta 2013, 34, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Pang, L.; Wei, Z.; Li, O.; Huang, R.; Qin, J.; Chen, H.; Fan, X.; Chen, Z.J. An increase in vascular endothelial growth factor (VEGF) and VEGF soluble receptor-1 (sFlt-1) are associated with early recurrent spontaneous abortion. PLoS ONE 2013, 8, e75759. [Google Scholar] [CrossRef]
- Laird, S.M.; Quinton, N.D.; Anstie, B.; Li, T.C.; Blakemore, A.I.F. Leptin and leptin-binding activity in women with recurrent miscarriage: Correlation with pregnancy outcome. Hum. Reprod. 2001, 16, 2008–2013. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.F. Soluble fms-like tyrosine kinase-1 and vascular endothelial growth factor: Novel markers for unexplained early recurrent pregnancy loss. Asian Pac. J. Reprod. 2014, 3, 30–34. [Google Scholar] [CrossRef]
- Christiansen, O.B.; Nybo Andersen, A.M.; Bosch, E.; Daya, S.; Delves, P.J.; Hviid, T.V.; Kutteh, W.H.; Laird, S.M.; Li, T.C.; van der Ven, K. Evidence-based investigations and treatments of recurrent pregnancy loss. Fertil. Steril. 2005, 83, 821–839. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, O.B. Evidence-based investigations and treatments of recurrent pregnancy loss. Curr. Opin. Obstet. Gynecol. 2006, 18, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Placental Oxidative Stress: From Miscarriage to Preeclampsia. J. Soc. Gynecol. Investig. 2004, 11, 342–352. [Google Scholar] [CrossRef]
- Lyu, S.W.; Song, H.; Yoon, J.A.; Chin, M.-U.; Sung, S.R.; Kim, Y.S.; Lee, W.S.; Yoon, T.K.; Cha, D.H.; Shim, S.H. Transcriptional profiling with a pathway-oriented analysis in the placental villi of unexplained miscarriage. Placenta 2013, 34, 133–140. [Google Scholar] [CrossRef]
- Liu, A.X.; Jin, F.; Zhang, W.W.; Zhou, T.H.; Zhou, C.Y.; Yao, W.M.; Qian, Y.L.; Huang, H.F. Proteomic analysis on the alteration of protein expression in the placental villous tissue of early pregnancy loss. Biol. Reprod. 2006, 75, 414–420. [Google Scholar] [CrossRef]
- Ni, X.; Li, X.; Guo, Y.; Zhou, T.; Guo, X.; Zhao, C.; Lin, M.; Zhou, Z.; Shen, R.; Guo, X.; et al. Quantitative proteomics analysis of altered protein expression in the placental villous tissue of early pregnancy loss using isobaric tandem mass tags. BioMed Res. Int. 2014, 2014, 647143. [Google Scholar] [CrossRef]
- Jauniaux, E.; Burton, G.J. Pathophysiology of histological changes in early pregnancy loss. Placenta 2005, 26, 114–123. [Google Scholar] [CrossRef]
- Jeschke, U.; Toth, B.; Scholz, C.; Friese, K.; Makrigiannakis, A. Glycoprotein and carbohydrate binding protein expression in the placenta in early pregnancy loss. J. Reprod. Immunol. 2010, 85, 99–105. [Google Scholar] [CrossRef]
- Gupta, S.; Agarwal, A.; Banerjee, J.; Alvarez, J.G. The Role of Oxidative Stress in Spontaneous Abortion and Recurrent Pregnancy Loss: A Systematic Review. Obstet. Gynecol. Surv. 2007, 62, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Su, A.I.; Wiltshire, T.; Batalov, S.; Lapp, H.; Ching, K.A.; Block, D.; Zhang, J.; Soden, R.; Hayakawa, M.; Kreiman, G.; et al. A gene atlas of the mouse and human protein-encoding transcriptomes. Proc. Natl. Acad. Sci. USA 2004, 101, 6062–6067. [Google Scholar] [CrossRef] [PubMed]
- Szilagyi, A.; Gelencser, Z.; Romero, R.; Xu, Y.; Kiraly, P.; Demeter, A.; Palhalmi, J.; Gyorffy, B.A.; Juhasz, K.; Hupuczi, P.; et al. Placenta-Specific Genes, Their Regulation During Villous Trophoblast Differentiation and Dysregulation in Preterm Preeclampsia. Int. J. Mol. Sci. 2020, 21, 628. [Google Scholar] [CrossRef]
- Than, N.G.; Romero, R.; Tarca, A.L.; Kekesi, K.A.; Xu, Y.; Xu, Z.; Juhasz, K.; Bhatti, G.; Leavitt, R.J.; Gelencser, Z.; et al. Integrated Systems Biology Approach Identifies Novel Maternal and Placental Pathways of Preeclampsia. Front. Immunol. 2018, 9, 1661. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.; Spencer, K.; Kagan, K.K.; Torring, N.; Petersen, O.B.; Christou, A.; Kallikas, J.; Nicolaides, K.H. First-trimester combined screening for trisomy 21 at 7-14 weeks’ gestation. Ultrasound Obstet. Gynecol. 2010, 36, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Tsiakkas, A.; Duvdevani, N.; Wright, A.; Wright, D.; Nicolaides, K.H. Serum placental growth factor in the three trimesters of pregnancy: Effects of maternal characteristics and medical history. Ultrasound Obstet. Gynecol. 2015, 45, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Erez, O.; Maymon, E.; Chaemsaithong, P.; Xu, Z.; Pacora, P.; Chaiworapongsa, T.; Done, B.; Hassan, S.S.; Tarca, A.L. The maternal plasma proteome changes as a function of gestational age in normal pregnancy: A longitudinal study. Am. J. Obstet. Gynecol. 2017, 217, 67.e1–67.e21. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. What is the placenta? Am. J. Obstet. Gynecol. 2015, 213, S6.E1–S6.E4. [Google Scholar] [CrossRef]
- Fulop, V.; Lakatos, K.; Demeter, J.; Vegh, G.; Pallinger, E. Clinical aspects of decidualization. Orvosi Hetil. 2022, 163, 1823–1833. [Google Scholar] [CrossRef]
- Lakatos, K.; Elias, K.M.; Berkowitz, R.S.; Hasselblatt, K.; Vegh, G.; Fulop, V. The role of natural killer cells in the immune homeostasis of the maternal fetal interface. Orvosi Hetil. 2022, 163, 734–742. [Google Scholar] [CrossRef]
- Nagy, B.; Sulyok, E.; Varnagy, A.; Barabas, A.; Kovacs, K.; Bodis, J. The role of platelets in reproduction. Orvosi Hetil. 2022, 163, 1254–1260. [Google Scholar] [CrossRef]
- Huppertz, B. Placental Origins of Preeclampsia. Hypertension 2008, 51, 970–975. [Google Scholar] [CrossRef] [PubMed]
- Myatt, L. Role of placenta in preeclampsia. Endocrine 2002, 19, 103–111. [Google Scholar] [CrossRef]
- Roberts, J.M.; Escudero, C. The placenta in preeclampsia. Pregnancy Hypertens. 2012, 2, 72–83. [Google Scholar] [CrossRef]
- Redman, C.W.; Sargent, I.L. Latest Advances in Understanding Preeclampsia. Science 2005, 308, 1592. [Google Scholar] [CrossRef] [PubMed]
- Sitras, V.; Paulssen, R.H.; Grønaas, H.; Leirvik, J.; Hanssen, T.A.; Vårtun, Å.; Acharya, G. Differential Placental Gene Expression in Severe Preeclampsia. Placenta 2009, 30, 424–433. [Google Scholar] [CrossRef]
- Hromadnikova, I. Extracellular nucleic acids in maternal circulation as potential biomarkers for placental insufficiency. DNA Cell Biol. 2012, 31, 1221–1232. [Google Scholar] [CrossRef]
- Ruano, C.S.M.; Miralles, F.; Mehats, C.; Vaiman, D. The Impact of Oxidative Stress of Environmental Origin on the Onset of Placental Diseases. Antioxidants 2022, 11, 106. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.T.; Ding, H.G.; Fang, M.; Yu, B.; Cheng, Y.; Tan, Y.J.; Fu, Q.Q.; Lu, B.; Cai, H.G.; Jin, X.; et al. Proteomics and bioinformatics analysis of altered protein expression in the placental villous tissue from early recurrent miscarriage patients. Placenta 2018, 61, 1–10. [Google Scholar] [CrossRef]
- Redline, R.W. Placental Pathology: A Systematic Approach with Clinical Correlations. Placenta 2008, 29, 86–91. [Google Scholar] [CrossRef]
- Lob, S.; Ochmann, B.; Ma, Z.; Vilsmaier, T.; Kuhn, C.; Schmoeckel, E.; Herbert, S.L.; Kolben, T.; Wockel, A.; Mahner, S.; et al. The role of Interleukin-18 in recurrent early pregnancy loss. J. Reprod. Immunol. 2021, 148, 103432. [Google Scholar] [CrossRef] [PubMed]
- Than, G.N.; Bohn, H.; Szabo, D.G. Advances in Pregnancy-Related Protein Research; CRC Press: Boca Raton, FL, USA, 1993. [Google Scholar]
- Human Protein Atlas. Available online: http://www.proteinatlas.org (accessed on 24 January 2024).
- Simpson, E.R.; MacDonald, P.C. Endocrine physiology of the placenta. Annu. Rev. Physiol. 1981, 43, 163–188. [Google Scholar] [CrossRef] [PubMed]
- Bohn, H.; Winckler, W. Isolation and characterization of membrane-associated placental proteins. Arch. Gynecol. Obstet. 1991, 248, 191–198. [Google Scholar] [CrossRef]
- Bohn, H.; Winckler, W.; Grundmann, U. Immunochemically detected placental proteins and their biological functions. Arch. Gynecol. Obstet. 1991, 249, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Malek, A.; Bersinger, N.A.; Di Santo, S.; Mueller, M.D.; Sager, R.; Schneider, H.; Ghezzi, F.; Karousou, E.; Passi, A.; De Luca, G.; et al. C-Reactive Protein Production in Term Human Placental Tissue. Placenta 2006, 27, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Chamley, L.W.; Allen, J.L.; Johnson, P.M. Synthesis of β2 glycoprotein 1 by the human placenta. Placenta 1997, 18, 403–410. [Google Scholar] [CrossRef]
- Ethier-Chiasson, M.; Duchesne, A.; Forest, J.C.; Giguère, Y.; Masse, A.; Mounier, C.; Lafond, J. Influence of maternal lipid profile on placental protein expression of LDLr and SR-BI. Biochem. Biophys. Res. Commun. 2007, 359, 8–14. [Google Scholar] [CrossRef]
- Jansson, T.; Wennergren, M.; Illsley, N.P. Glucose transporter protein expression in human placenta throughout gestation and in intrauterine growth retardation. J. Clin. Endocrinol. Metab. 1993, 77, 1554–1562. [Google Scholar] [CrossRef]
- Crowley, E.; Di Nicolantonio, F.; Loupakis, F.; Bardelli, A. Liquid biopsy: Monitoring cancer-genetics in the blood. Nat. Rev. Clin. Oncol. 2013, 10, 472–484. [Google Scholar] [CrossRef]
- Alix-Panabières, C.; Pantel, K. Circulating Tumor Cells: Liquid Biopsy of Cancer. Clin. Chem. 2013, 59, 110–118. [Google Scholar] [CrossRef]
- Heitzer, E.; Ulz, P.; Geigl, J.B. Circulating Tumor DNA as a Liquid Biopsy for Cancer. Clin. Chem. 2015, 61, 112–123. [Google Scholar] [CrossRef]
- Alix-Panabières, C.; Pantel, K. Clinical Applications of Circulating Tumor Cells and Circulating Tumor DNA as Liquid Biopsy. Cancer Discov. 2016, 6, 479. [Google Scholar] [CrossRef]
- Karaszi, K.; Szabo, S.; Juhasz, K.; Kiraly, P.; Kocsis-Deak, B.; Hargitai, B.; Krenacs, T.; Hupuczi, P.; Erez, O.; Papp, Z.; et al. Increased placental expression of Placental Protein 5 (PP5)/Tissue Factor Pathway Inhibitor-2 (TFPI-2) in women with preeclampsia and HELLP syndrome: Relevance to impaired trophoblast invasion? Placenta 2019, 76, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Than, N.G.; Balogh, A.; Romero, R.; Kárpáti, É.; Erez, O.; Szilágyi, A.; Kovalszky, I.; Sammar, M.; Gizurarson, S.; Matkó, J.; et al. Placental Protein 13 (PP13)—A Placental Immunoregulatory Galectin Protecting Pregnancy. Front. Immunol. 2014, 5, 348. [Google Scholar] [CrossRef] [PubMed]
- Balogh, A.; Pozsgay, J.; Matkó, J.; Dong, Z.; Kim, C.J.; Várkonyi, T.; Sammar, M.; Rigó, J.; Meiri, H.; Romero, R.; et al. Placental protein 13 (PP13/galectin-13) undergoes lipid raft-associated subcellular redistribution in the syncytiotrophoblast in preterm preeclampsia and HELLP syndrome. Am. J. Obstet. Gynecol. 2011, 205, 156.e1–156.e14. [Google Scholar] [CrossRef]
- Than, N.G.; Abdul Rahman, O.; Magenheim, R.; Nagy, B.; Fule, T.; Hargitai, B.; Sammar, M.; Hupuczi, P.; Tarca, A.L.; Szabo, G.; et al. Placental protein 13 (galectin-13) has decreased placental expression but increased shedding and maternal serum concentrations in patients presenting with preterm pre-eclampsia and HELLP syndrome. Virchows Arch. 2008, 453, 387–400. [Google Scholar] [CrossRef]
- Than, N.G.; Romero, R.; Balogh, A.; Karpati, E.; Mastrolia, S.A.; Staretz-Chacham, O.; Hahn, S.; Erez, O.; Papp, Z.; Kim, C.J. Galectins: Double-edged Swords in the Cross-roads of Pregnancy Complications and Female Reproductive Tract Inflammation and Neoplasia. J. Pathol. Transl. Med. 2015, 49, 181–208. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Kusanovic, J.P.; Than, N.G.; Erez, O.; Gotsch, F.; Espinoza, J.; Edwin, S.; Chefetz, I.; Gomez, R.; Nien, J.K.; et al. First-trimester maternal serum PP13 in the risk assessment for preeclampsia. Am. J. Obstet. Gynecol. 2008, 199, 122.e1–122.e11. [Google Scholar] [CrossRef]
- Than, N.G.; Romero, R.; Meiri, H.; Erez, O.; Xu, Y.; Tarquini, F.; Barna, L.; Szilagyi, A.; Ackerman, R.; Sammar, M.; et al. PP13, maternal ABO blood groups and the risk assessment of pregnancy complications. PLoS ONE 2011, 6, e21564. [Google Scholar] [CrossRef]
- Balogh, A.; Toth, E.; Romero, R.; Parej, K.; Csala, D.; Szenasi, N.L.; Hajdu, I.; Juhasz, K.; Kovacs, A.F.; Meiri, H.; et al. Placental Galectins Are Key Players in Regulating the Maternal Adaptive Immune Response. Front. Immunol. 2019, 10, 1240. [Google Scholar] [CrossRef]
- Vokalova, L.; Balogh, A.; Toth, E.; Van Breda, S.V.; Schäfer, G.; Hoesli, I.; Lapaire, O.; Hahn, S.; Than, N.G.; Rossi, S.W. Placental Protein 13 (Galectin-13) Polarizes Neutrophils Toward an Immune Regulatory Phenotype. Front. Immunol. 2020, 11, 145. [Google Scholar] [CrossRef] [PubMed]
- Spencer, K.; Cowans, N.J.; Nicolaides, K.H. Low levels of maternal serum PAPP-A in the first trimester and the risk of pre-eclampsia. Prenat. Diagn. 2008, 28, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Goetzinger, K.R.; Singla, A.; Gerkowicz, S.; Dicke, J.M.; Gray, D.L.; Odibo, A.O. Predicting the risk of pre-eclampsia between 11 and 13 weeks’ gestation by combining maternal characteristics and serum analytes, PAPP-A and free β-hCG. Prenat. Diagn. 2010, 30, 1138–1142. [Google Scholar] [CrossRef] [PubMed]
- Mikat, B.; Zeller, A.; Scherag, A.; Drommelschmidt, K.; Kimmig, R.; Schmidt, M. βhCG and PAPP-A in First Trimester: Predictive Factors for Preeclampsia? Hypertens. Pregnancy 2012, 31, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Spencer, K.; Yu, C.K.H.; Cowans, N.J.; Otigbah, C.; Nicolaides, K.H. Prediction of pregnancy complications by first-trimester maternal serum PAPP-A and free β-hCG and with second-trimester uterine artery Doppler. Prenat. Diagn. 2005, 25, 949–953. [Google Scholar] [CrossRef] [PubMed]
- Basirat, Z.; Barat, S.; Hajiahmadi, M. Serum beta human chorionic gonadotropin levels and preeclampsia. Saudi Med. J. 2006, 27, 1001–1004. [Google Scholar] [PubMed]
- Cicero, S.; Bindra, R.; Rembouskos, G.; Spencer, K.; Nicolaides, K.H. Integrated ultrasound and biochemical screening for trisomy 21 using fetal nuchal translucency, absent fetal nasal bone, free β-hCG and PAPP-A at 11 to 14 weeks. Prenat. Diagn. 2003, 23, 306–310. [Google Scholar] [CrossRef]
- Spencer, K.; Ong, C.; Skentou, H.; Liao, A.W.; Nicolaides, K.H. Screening for trisomy 13 by fetal nuchal translucency and maternal serum free β-hCG and PAPP-A at 10–14 weeks of gestation. Prenat. Diagn. 2000, 20, 411–416. [Google Scholar] [CrossRef]
- Tul, N.; Spencer, K.; Noble, P.; Chan, C.; Nicolaides, K. Screening for trisomy 18 by fetal nuchal translucency and maternal serum free β-hCG and PAPP-A at 10–14 weeks of gestation. Prenat. Diagn. 1999, 19, 1035–1042. [Google Scholar] [CrossRef]
- Spencer, K.; Nicolaides, K.H. A first trimester trisomy 13/trisomy 18 risk algorithm combining fetal nuchal translucency thickness, maternal serum free β-hCG and PAPP-A. Prenat. Diagn. 2002, 22, 877–879. [Google Scholar] [CrossRef]
- Brizot, M.L.; Snijders, R.J.; Bersinger, N.A.; Kuhn, P.; Nicolaides, K.H. Maternal serum pregnancy-associated plasma protein A and fetal nuchal translucency thickness for the prediction of fetal trisomies in early pregnancy. Obstet. Gynecol. 1994, 84, 918–922. [Google Scholar] [PubMed]
- Zeisler, H.; Llurba, E.; Chantraine, F.; Vatish, M.; Staff, A.C.; Sennström, M.; Olovsson, M.; Brennecke, S.P.; Stepan, H.; Allegranza, D.; et al. Predictive Value of the sFlt-1:PlGF Ratio in Women with Suspected Preeclampsia. N. Engl. J. Med. 2016, 374, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Sunderji, S.; Gaziano, E.; Wothe, D.; Rogers, L.C.; Sibai, B.; Karumanchi, S.A.; Hodges-Savola, C. Automated assays for sVEGF R1 and PlGF as an aid in the diagnosis of preterm preeclampsia: A prospective clinical study. Am. J. Obstet. Gynecol. 2010, 202, 40.e1–40.e7. [Google Scholar] [CrossRef] [PubMed]
- Cowans, N.J.; Stamatopoulou, A.; Spencer, K. First trimester maternal serum placental growth factor in trisomy 21 pregnancies. Prenat. Diagn. 2010, 30, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Stenman, U.H.; Tiitinen, A.; Alfthan, H.; Valmu, L. The classification, functions and clinical use of different isoforms of HCG. Hum. Reprod. Update 2006, 12, 769–784. [Google Scholar] [CrossRef] [PubMed]
- Carmona, F.; Balasch, J.; Creus, M.; Fábregues, F.; Casamitjana, R.; Cívico, S.; Vidal, E.; Calafell, J.M.; Moreno, V.; Vanrell, J.A. Early hormonal markers of pregnancy outcome after in vitro fertilization and embryo transfer. J. Assist. Reprod. Genet. 2003, 20, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Pillai, R.N.; Konje, J.C.; Tincello, D.G.; Potdar, N. Role of serum biomarkers in the prediction of outcome in women with threatened miscarriage: A systematic review and diagnostic accuracy meta-analysis. Hum. Reprod. Update 2016, 22, 228–239. [Google Scholar] [CrossRef] [PubMed]
- The UniProt Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2018, 47, D506–D515. [Google Scholar] [CrossRef]
- Lawrence, J.B.; Oxvig, C.; Overgaard, M.T.; Sottrup-Jensen, L.; Gleich, G.J.; Hays, L.G.; Yates, J.R., 3rd; Conover, C.A. The insulin-like growth factor (IGF)-dependent IGF binding protein-4 protease secreted by human fibroblasts is pregnancy-associated plasma protein-A. Proc. Natl. Acad. Sci. USA 1999, 96, 3149–3153. [Google Scholar] [CrossRef]
- Laursen, L.S.; Overgaard, M.T.; Soe, R.; Boldt, H.B.; Sottrup-Jensen, L.; Giudice, L.C.; Conover, C.A.; Oxvig, C. Pregnancy-associated plasma protein-A (PAPP-A) cleaves insulin-like growth factor binding protein (IGFBP)-5 independent of IGF: Implications for the mechanism of IGFBP-4 proteolysis by PAPP-A. FEBS Lett. 2001, 504, 36–40. [Google Scholar] [CrossRef]
- Conover, C.A.; Oxvig, C.; Overgaard, M.T.; Christiansen, M.; Giudice, L.C. Evidence that the insulin-like growth factor binding protein-4 protease in human ovarian follicular fluid is pregnancy associated plasma protein-A. J. Clin. Endocrinol. Metab. 1999, 84, 4742–4745. [Google Scholar] [CrossRef] [PubMed]
- Prasad, P.; Romero, R.; Chaiworapongsa, T.; Gomez-Lopez, N.; Lo, A.; Galaz, J.; Taran, A.B.; Jung, E.; Gotsch, F.; Than, N.G.; et al. Further Evidence that an Episode of Premature Labor Is a Pathologic State: Involvement of the Insulin-Like Growth Factor System. Fetal Diagn. Ther. 2023, 50, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, Q.; Xie, Y.; Su, K.; Yang, J.; Yang, L. A correlation analysis between the expression of pregnancy-associated plasma protein A in basal decidual cells and recurrent spontaneous abortion. Exp. Ther. Med. 2013, 6, 485–488. [Google Scholar] [CrossRef] [PubMed]
- Rolny, C.; Mazzone, M.; Tugues, S.; Laoui, D.; Johansson, I.; Coulon, C.; Squadrito, M.L.; Segura, I.; Li, X.; Knevels, E.; et al. HRG inhibits tumor growth and metastasis by inducing macrophage polarization and vessel normalization through downregulation of PlGF. Cancer Cell 2011, 19, 31–44. [Google Scholar] [CrossRef]
- Lizio, M.; Harshbarger, J.; Shimoji, H.; Severin, J.; Kasukawa, T.; Sahin, S.; Abugessaisa, I.; Fukuda, S.; Hori, F.; Ishikawa-Kato, S.; et al. Gateways to the FANTOM5 promoter level mammalian expression atlas. Genome Biol. 2015, 16, 22. [Google Scholar] [CrossRef] [PubMed]
- Genotype-Tissue Expression (GTEx) Project. Available online: https://www.gtexportal.org/home/ (accessed on 20 December 2023).
- Plaisier, M.; Rodrigues, S.; Willems, F.; Koolwijk, P.; van Hinsbergh, V.W.; Helmerhorst, F.M. Different degrees of vascularization and their relationship to the expression of vascular endothelial growth factor, placental growth factor, angiopoietins, and their receptors in first-trimester decidual tissues. Fertil. Steril. 2007, 88, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Vailhe, B.; Dietl, J.; Kapp, M.; Toth, B.; Arck, P. Increased blood vessel density in decidua parietalis is associated with spontaneous human first trimester abortion. Hum. Reprod. 1999, 14, 1628–1634. [Google Scholar] [CrossRef]
- Plaisier, M.; Dennert, I.; Rost, E.; Koolwijk, P.; van Hinsbergh, V.W.; Helmerhorst, F.M. Decidual vascularization and the expression of angiogenic growth factors and proteases in first trimester spontaneous abortions. Hum. Reprod. 2009, 24, 185–197. [Google Scholar] [CrossRef]
- Dev, S.; Singh, A.; Banerjee, B.; Radhakrishnan, G.; Agarwal, R. Assessment of Maternal Serum Levels of Vascular Endothelial Growth Factor and Placental Growth Factor in Threatened Abortion: A Case Control Study. J. Clin. Diagn. Res. 2020, 14, 16–18. [Google Scholar] [CrossRef]
- Muttukrishna, S.; Swer, M.; Suri, S.; Jamil, A.; Calleja-Agius, J.; Gangooly, S.; Ludlow, H.; Jurkovic, D.; Jauniaux, E. Soluble Flt-1 and PlGF: New markers of early pregnancy loss? PLoS ONE 2011, 6, e18041. [Google Scholar] [CrossRef]
- Andersen, L.B.; Dechend, R.; Karumanchi, S.A.; Nielsen, J.; Joergensen, J.S.; Jensen, T.K.; Christesen, H.T. Early pregnancy angiogenic markers and spontaneous abortion: An Odense Child Cohort study. Am. J. Obstet. Gynecol. 2016, 215, 594.E1–594.E11. [Google Scholar] [CrossRef]
- Horne, A.W.; Shaw, J.L.; Murdoch, A.; McDonald, S.E.; Williams, A.R.; Jabbour, H.N.; Duncan, W.C.; Critchley, H.O. Placental growth factor: A promising diagnostic biomarker for tubal ectopic pregnancy. J. Clin. Endocrinol. Metab. 2011, 96, E104–E108. [Google Scholar] [CrossRef] [PubMed]
- Abu-Ghazaleh, N.; Brennecke, S.; Murthi, P.; Karanam, V. Association of Vascular Endothelial Growth Factors (VEGFs) with Recurrent Miscarriage: A Systematic Review of the Literature. Int. J. Mol. Sci. 2023, 24, 9449. [Google Scholar] [CrossRef]
- Vranken, G.; Reynolds, T.; Van Nueten, J. Medians for second-trimester maternal serum markers: Geographical differences and variation caused by median multiples-of-median equations. J. Clin. Pathol. 2006, 59, 639. [Google Scholar] [CrossRef]
- Levy, K.J. Pairwise Comparisons Associated with the K Independent Sample Median Test. Am. Stat. 1979, 33, 138–139. [Google Scholar] [CrossRef]
- Nemenyi, P. Distribution-Free Multiple Comparisons; Princeton University: Princeton, NJ, USA, 1963. [Google Scholar]
- Tukey, J.W. Comparing individual means in the analysis of variance. Biometrics 1949, 5, 99–114. [Google Scholar] [CrossRef]
- Kramer, C.Y. Extension of multiple range tests to group means with unequal numbers of replications. Biometrics 1956, 12, 309–310. [Google Scholar] [CrossRef]
- Richter, S.J.; McCann, M.H. Multiple Comparison of Medians Using Permutation Tests. J. Mod. Appl. Stat. Methods 2007, 6, 399–412. [Google Scholar] [CrossRef]
- Cole, L.A. Biological functions of hCG and hCG-related molecules. Reprod. Biol. Endocrinol. 2010, 8, 102. [Google Scholar] [CrossRef]
- Cole, L.A. hCG physiology. Placenta 2013, 34, 1257. [Google Scholar] [CrossRef]
- Cole, L.A. Hyperglycosylated hCG and pregnancy failures. J. Reprod. Immunol. 2012, 93, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Development of the Human Placenta and Fetal Heart: Synergic or Independent? Front. Physiol. 2018, 9, 373. [Google Scholar] [CrossRef] [PubMed]
- Huppertz, B. Traditional and New Routes of Trophoblast Invasion and Their Implications for Pregnancy Diseases. Int. J. Mol. Sci. 2019, 21, 289. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E.; Watson, A.L. Maternal arterial connections to the placental intervillous space during the first trimester of human pregnancy: The Boyd collection revisited. Am. J. Obstet. Gynecol. 1999, 181, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Chaiworapongsa, T.; Romero, R.; Korzeniewski, S.J.; Kusanovic, J.P.; Soto, E.; Lam, J.; Dong, Z.; Than, N.G.; Yeo, L.; Hernandez-Andrade, E.; et al. Maternal plasma concentrations of angiogenic/antiangiogenic factors in the third trimester of pregnancy to identify the patient at risk for stillbirth at or near term and severe late preeclampsia. Am. J. Obstet. Gynecol. 2013, 208, 287.E1–287.E15. [Google Scholar] [CrossRef]
- Chaiworapongsa, T.; Romero, R.; Erez, O.; Tarca, A.L.; Conde-Agudelo, A.; Chaemsaithong, P.; Kim, C.J.; Kim, Y.M.; Kim, J.S.; Yoon, B.H.; et al. The prediction of fetal death with a simple maternal blood test at 20–24 weeks: A role for angiogenic index-1 (PlGF/sVEGFR-1 ratio). Am. J. Obstet. Gynecol. 2017, 217, 682.E1–682.E13. [Google Scholar] [CrossRef]
- Whitten, A.E.; Romero, R.; Korzeniewski, S.J.; Tarca, A.L.; Schwartz, A.G.; Yeo, L.; Dong, Z.; Hassan, S.S.; Chaiworapongsa, T. Evidence of an imbalance of angiogenic/antiangiogenic factors in massive perivillous fibrin deposition (maternal floor infarction): A placental lesion associated with recurrent miscarriage and fetal death. Am. J. Obstet. Gynecol. 2013, 208, 310.E1–310.E11. [Google Scholar] [CrossRef]
- Romero, R.; Kusanovic, J.P.; Chaiworapongsa, T.; Hassan, S.S. Placental bed disorders in preterm labor, preterm PROM, spontaneous abortion and abruptio placentae. Best Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 313–327. [Google Scholar] [CrossRef]
- Brosens, I.; Pijnenborg, R.; Vercruysse, L.; Romero, R. The “Great Obstetrical Syndromes” are associated with disorders of deep placentation. Am. J. Obstet. Gynecol. 2011, 204, 193–201. [Google Scholar] [CrossRef]
- Boyd, T.K. The placenta in intrauterine demise. J. Pathol. Microbiol. Immunol. 2018, 126, 621–625. [Google Scholar] [CrossRef]
- King, J.R.; Wilson, M.L.; Hetey, S.; Kiraly, P.; Matsuo, K.; Castaneda, A.V.; Toth, E.; Krenacs, T.; Hupuczi, P.; Mhawech-Fauceglia, P.; et al. Dysregulation of Placental Functions and Immune Pathways in Complete Hydatidiform Moles. Int. J. Mol. Sci. 2019, 20, 4999. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Sayre, J.; Castaneda, A.V.; Roman, L.D.; Matsuo, K. Diagnosis and Management of Gestational Trophoblastic Disease. In Handbook of Gynecology; Springer: New York, NY, USA, 2017; pp. 1–15. [Google Scholar]
- Reus, A.D.; El-Harbachi, H.; Rousian, M.; Willemsen, S.P.; Steegers-Theunissen, R.P.; Steegers, E.A.; Exalto, N. Early first-trimester trophoblast volume in pregnancies that result in live birth or miscarriage. Ultrasound Obstet. Gynecol. 2013, 42, 577–584. [Google Scholar] [CrossRef] [PubMed]
- ACOG. Practice Bulletin No. 200: Early Pregnancy Loss. Obstet. Gynecol. 2018, 132, e197–e207. [Google Scholar] [CrossRef] [PubMed]
- Vomstein, K.; Feil, K.; Strobel, L.; Aulitzky, A.; Hofer-Tollinger, S.; Kuon, R.J.; Toth, B. Immunological Risk Factors in Recurrent Pregnancy Loss: Guidelines Versus Current State of the Art. J. Clin. Med. 2021, 10, 869. [Google Scholar] [CrossRef]
- Tabacco, S.; Giannini, A.; Garufi, C.; Botta, A.; Salvi, S.; Del Sordo, G.; Benedetti Panici, P.; Lanzone, A.; De Carolis, S. Complementemia in pregnancies with antiphospholipid syndrome. Lupus 2019, 28, 1503–1509. [Google Scholar] [CrossRef]
- ACOG. The American College of Obstetricians and Gynecologists Practice Bulletin no. 150. Early pregnancy loss. Obstet. Gynecol. 2015, 125, 1258–1267. [Google Scholar] [CrossRef]
- Poon, L.C.; Shennan, A.; Hyett, J.A.; Kapur, A.; Hadar, E.; Divakar, H.; McAuliffe, F.; da Silva Costa, F.; von Dadelszen, P.; McIntyre, H.D.; et al. The International Federation of Gynecology and Obstetrics (FIGO) initiative on pre-eclampsia: A pragmatic guide for first-trimester screening and prevention. Int. J. Gynecol. Obstet. 2019, 145 (Suppl. 1), 1–33. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | RPL | Control |
|---|---|---|
| Number of cases a | 14 | 30 |
| Maternal age (years) a | 37.2 ± 4.5 ** | 30.1 ± 7.0 |
| Gestational age at surgery (weeks) a | 9.1 ± 1.9 | 8.6 ± 1.8 |
| Gravidity b | 3 (2–4) * | 2 (1–3) |
| Parity b | 1 (0–1) | 0 (0–1) |
| Number of previous miscarriages b,c | 1 (1–1.8) *** | 0 (0–0) |
| Concentration | dMoM | |||||
|---|---|---|---|---|---|---|
| free β-hCG | PAPP-A | PlGF | free β-hCG | PAPP-A | PlGF | |
| AUC | 0.788 | 0.632 | 0.626 | 0.820 | 0.880 | 0.644 |
| TPR % (5% FPR) | 42.86 | 7.57 | 1.00 | 47.54 | 38.92 | 8.67 |
| LR+ (5% FPR) | 8.57 | 1.51 | 0.20 | 9.51 | 7.78 | 1.73 |
| LR− (5% FPR) | 0.60 | 0.97 | 1.04 | 0.55 | 0.64 | 0.96 |
| TPR % (10% FPR) | 64.14 | 13.57 | 14.71 | 70.31 | 82.62 | 28.50 |
| LR+ (10% FPR) | 6.41 | 1.36 | 1.47 | 7.03 | 8.26 | 2.85 |
| LR− (10% FPR) | 0.40 | 0.96 | 0.95 | 0.33 | 0.19 | 0.79 |
| Concentration | dMoM | |||||
|---|---|---|---|---|---|---|
| free β-hCG | PAPP-A | PlGF | free β-hCG | PAPP-A | PlGF | |
| AUC | 0.619 | 0.463 | 0.279 | 0.667 | 0.789 | 0.368 |
| TPR % (5% FPR) | 10.00 | 4.89 | 0.00 | 11.50 | 14.00 | 3.71 |
| LR+ (5% FPR) | 2.00 | 0.98 | 0.00 | 2.30 | 2.80 | 0.74 |
| LR− (5% FPR) | 0.95 | 1.00 | 1.05 | 0.93 | 0.91 | 1.01 |
| TPR % (10% FPR) | 17.11 | 8.44 | 0.22 | 30.50 | 26.50 | 3.71 |
| LR+ (10% FPR) | 1.71 | 0.84 | 0.02 | 3.05 | 2.65 | 0.37 |
| LR− (10% FPR) | 0.92 | 1.02 | 1.11 | 0.78 | 0.82 | 1.07 |
| Concentration | dMoM | |||||
|---|---|---|---|---|---|---|
| free β-hCG | PAPP-A | PlGF | free β-hCG | PAPP-A | PlGF | |
| AUC | 0.999 | 0.635 | 0.778 | 0.975 | 0.998 | 0.924 |
| TPR % (5% FPR) | 99.60 | 14.40 | 26.00 | 70.40 | 98.00 | 24.40 |
| LR+ (5% FPR) | 19.92 | 2.88 | 5.20 | 14.08 | 19.60 | 4.88 |
| LR− (5% FPR) | 0.00 | 0.90 | 0.78 | 0.31 | 0.02 | 0.80 |
| TPR % (10% FPR) | 100.00 | 32.40 | 59.20 | 100.00 | 100.00 | 100.00 |
| LR+ (10% FPR) | 10.00 | 3.24 | 5.92 | 10.00 | 10.00 | 10.00 |
| LR− (10% FPR) | 0.00 | 0.75 | 0.45 | 0.00 | 0.00 | 0.00 |
| free β-hCG, PAPP-A | free β-hCG, PlGF | PAPP-A, PlGF | free β-hCG, PAPP-A, PlGF | ||
|---|---|---|---|---|---|
| concentration | AUC | 0.806 | 0.784 | 0.695 | 0.793 |
| TPR % (5% FPR) | 44.14 | 56.71 | 14.71 | 45.86 | |
| LR+ (5% FPR) | 8.83 | 11.34 | 2.94 | 9.17 | |
| LR− (5% FPR) | 0.59 | 0.46 | 0.90 | 0.57 | |
| TPR % (10% FPR) | 61.29 | 65.00 | 21.43 | 55.86 | |
| LR+ (10% FPR) | 6.13 | 6.50 | 2.14 | 5.59 | |
| LR− (10% FPR) | 0.43 | 0.39 | 0.87 | 0.49 | |
| dMoM | AUC | 0.867 | 0.786 | 0.865 | 0.846 |
| TPR % (5% FPR) | 54.00 | 56.50 | 46.33 | 43.67 | |
| LR+ (5% FPR) | 10.80 | 11.30 | 9.27 | 8.73 | |
| LR− (5% FPR) | 0.48 | 0.46 | 0.56 | 0.59 | |
| TPR % (10% FPR) | 76.92 | 63.00 | 80.00 | 75.33 | |
| LR+ (10% FPR) | 7.69 | 6.30 | 8.00 | 7.53 | |
| LR− (10% FPR) | 0.26 | 0.41 | 0.22 | 0.27 |
| Independent Variables | |
|---|---|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tóth, E.; Györffy, D.; Posta, M.; Hupuczi, P.; Balogh, A.; Szalai, G.; Orosz, G.; Orosz, L.; Szilágyi, A.; Oravecz, O.; et al. Decreased Expression of Placental Proteins in Recurrent Pregnancy Loss: Functional Relevance and Diagnostic Value. Int. J. Mol. Sci. 2024, 25, 1865. https://doi.org/10.3390/ijms25031865
Tóth E, Györffy D, Posta M, Hupuczi P, Balogh A, Szalai G, Orosz G, Orosz L, Szilágyi A, Oravecz O, et al. Decreased Expression of Placental Proteins in Recurrent Pregnancy Loss: Functional Relevance and Diagnostic Value. International Journal of Molecular Sciences. 2024; 25(3):1865. https://doi.org/10.3390/ijms25031865
Chicago/Turabian StyleTóth, Eszter, Dániel Györffy, Máté Posta, Petronella Hupuczi, Andrea Balogh, Gábor Szalai, Gergő Orosz, László Orosz, András Szilágyi, Orsolya Oravecz, and et al. 2024. "Decreased Expression of Placental Proteins in Recurrent Pregnancy Loss: Functional Relevance and Diagnostic Value" International Journal of Molecular Sciences 25, no. 3: 1865. https://doi.org/10.3390/ijms25031865
APA StyleTóth, E., Györffy, D., Posta, M., Hupuczi, P., Balogh, A., Szalai, G., Orosz, G., Orosz, L., Szilágyi, A., Oravecz, O., Veress, L., Nagy, S., Török, O., Murthi, P., Erez, O., Papp, Z., Ács, N., & Than, N. G. (2024). Decreased Expression of Placental Proteins in Recurrent Pregnancy Loss: Functional Relevance and Diagnostic Value. International Journal of Molecular Sciences, 25(3), 1865. https://doi.org/10.3390/ijms25031865