

Protective Effect of Probiotics against Pseudomonas aeruginosa Infection of Human Corneal Epithelial Cells

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Strains and Growth Conditions
2.1.1. Aggregation and Coaggregation with P. aeruginosa
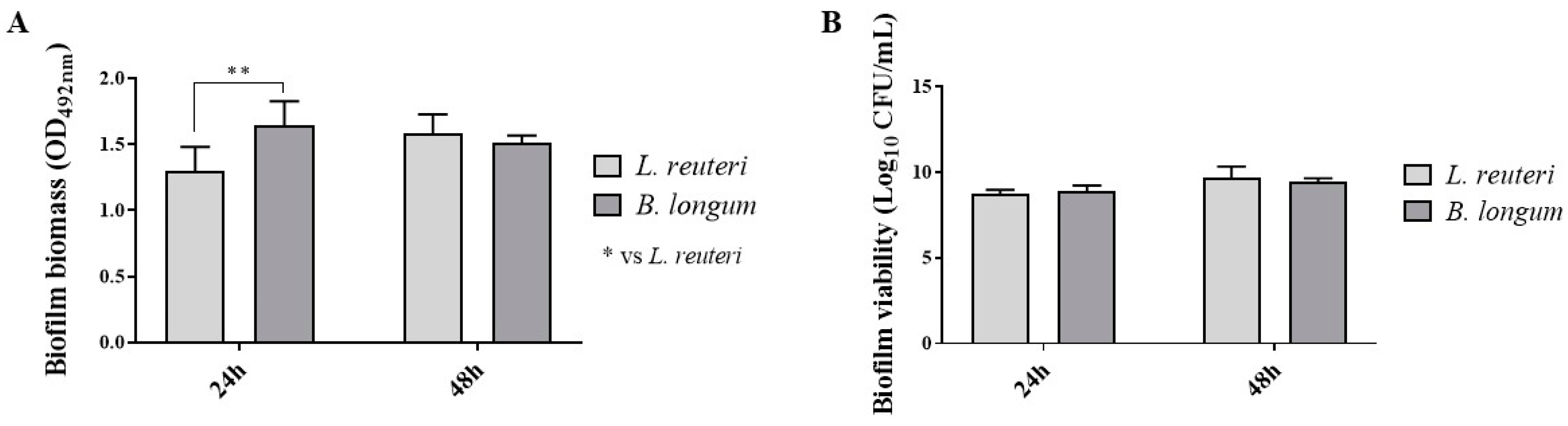
2.1.2. Biofilm Production
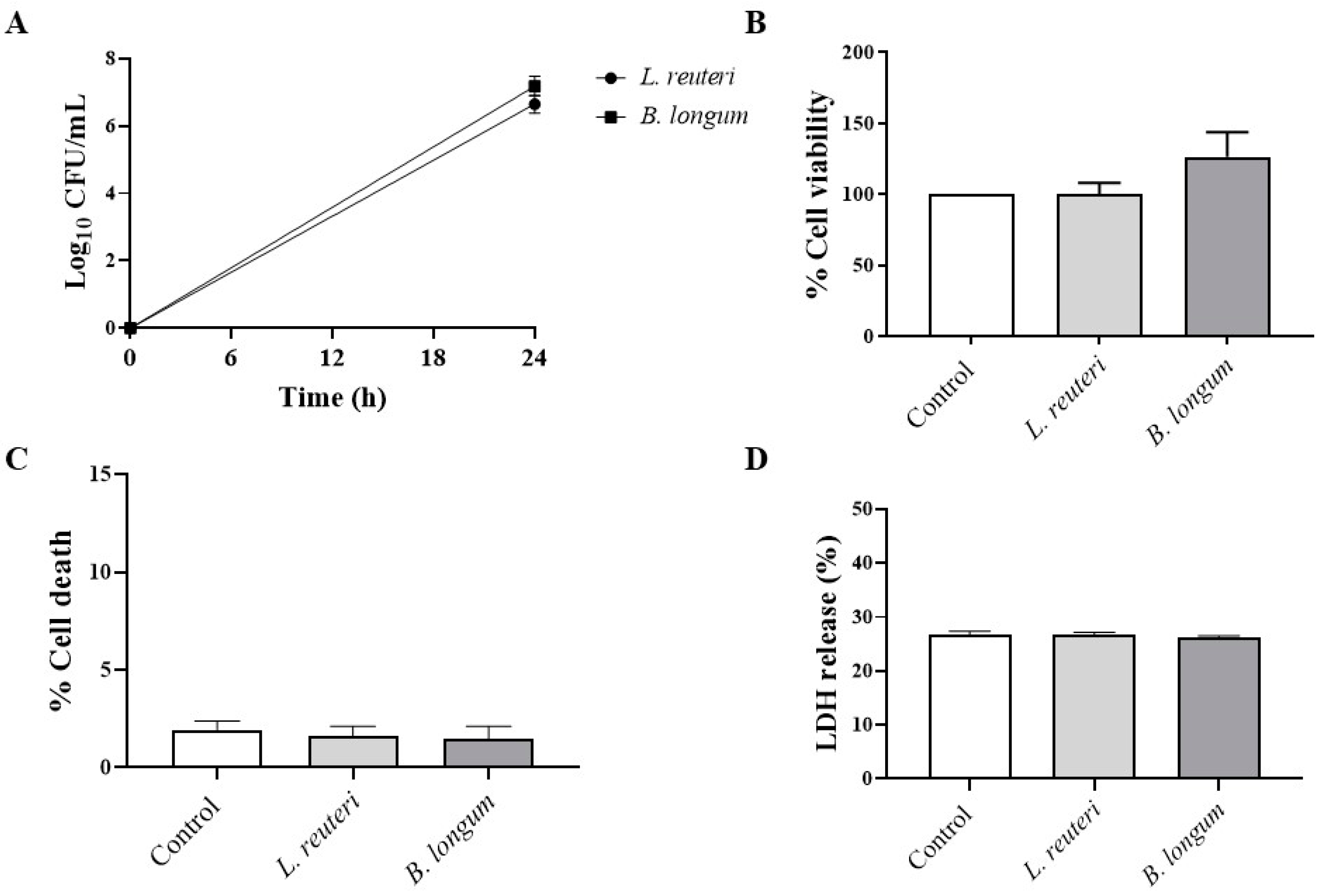
2.1.3. Effect on HCE Cell Viability
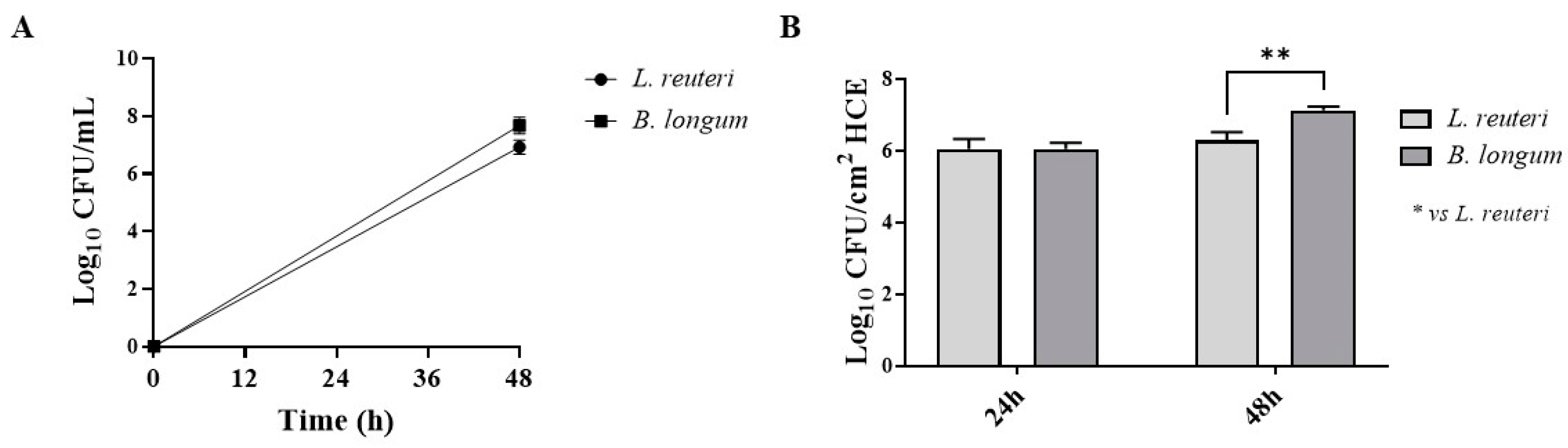
2.1.4. Adhesion to HCE Cells
2.1.5. Effect on Occludin and Mucin-1 in SkinEthicTM HCE Model
2.2. Prevention of P. aeruginosa Infection Damage via Pretreatment of HCE Cells with Probiotic Strains
2.2.1. Effect on HCE Cell Viability
2.2.2. Antagonistic Activity against P. aeruginosa Adhesion
2.2.3. Anti-Inflammatory Activity against P. aeruginosa
2.2.4. Antinitrosative Activity against P. aeruginosa
3. Discussion
4. Materials and Methods
4.1. Strains and Growth Conditions
4.2. Aggregation and Coaggregation with P. aeruginosa
4.3. Biofilm Production
4.4. Cell Culture
4.5. Viability Assays
4.6. Adhesion to HCE Cells
4.7. SkinEthicTM HCE Model
- SkinEthicTM HCE cells in DMEM (control group);
- SkinEthicTM HCE cells treated with each probiotic strain at 1 × 107 CFU/mL for 24 h at 37 °C, under 5% CO2 conditions.
4.8. Immunofluorescence (IF) Staining for Intercellular Junctions and Mucin Localization
4.9. Western Blot Analysis
4.10. Pretreatment of HCE Cells with Probiotic Strains Prevents P. aeruginosa Infection Damage
- HCE cells in DMEM (control group);
- HCE cells pretreated with each probiotic strain (pretreated cells);
- HCE cells infected with P. aeruginosa (infected cells);
- HCE cells pretreated with each probiotic strain and then infected with P. aeruginosa (pretreated and infected cells).
4.11. Viability Assays
4.12. Antagonistic Activity against P. aeruginosa Adhesion to HCE Cells
4.13. Enzyme-Linked Immunosorbent Assay for TNFα and IL-10 in HCE Cell Supernatant
4.14. Measurement of NOx Levels in HCE Cell Supernatant
4.15. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Mujaini, A.; Al-Kharusi, N.; Thakral, A.; Wali, U.K. Bacterial keratitis: Perspective on epidemiology, clinico-pathogenesis, diagnosis and treatment. Sultan Qaboos Univ. Med. J. 2009, 9, 184–195. [Google Scholar]
- Xue, W.; Li, J.J.; Zou, Y.; Zou, B.; Wei, L. Microbiota and Ocular Diseases. Front. Cell Infect. Microbiol. 2021, 11, 759333. [Google Scholar] [CrossRef]
- Sy, A.; Srinivasan, M.; Mascarenhas, J.; Lalitha, P.; Rajaraman, R.; Ravindran, M.; Oldenburg, C.E.; Ray, K.J.; Glidden, D.; Zegans, M.E.; et al. Pseudomonas aeruginosa keratitis: Outcomes and response to corticosteroid treatment. Investig. Ophthalmol. Vis. Sci. 2012, 53, 267–272. [Google Scholar] [CrossRef]
- Rossi, E.; Ghoul, M.; La Rosa, R. Editorial: Pseudomonas aeruginosa Pathogenesis: Virulence, Antibiotic Tolerance and Resistance, Stress Responses and Host-Pathogen Interactions. Front. Cell Infect. Microbiol. 2022, 12, 860314. [Google Scholar] [CrossRef]
- Fleiszig, S.M.; Evans, D.J. The pathogenesis of bacterial keratitis: Studies with Pseudomonas aeruginosa. Clin. Exp. Optom. 2002, 85, 271–278. [Google Scholar] [CrossRef]
- Hazlett, L.D.; Huang, X.; McClellan, S.A.; Barrett, R.P. Further studies on the role of IL-12 in Pseudomonas aeruginosa corneal infection. Eye 2003, 17, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, F.; Bakkar, M.; Carnt, N.; Chalmers, R.; Vijay, A.K.; Marasini, S.; Ng, A.; Tan, J.; Wagner, H.; Woods, C.; et al. CLEAR—Contact lens complications. Cont. Lens Anterior Eye 2021, 44, 330–367. [Google Scholar] [CrossRef] [PubMed]
- Chiang, M.C.; Chern, E. More than Antibiotics: Latest Therapeutics in the Treatment and Prevention of Ocular Surface Infections. J. Clin. Med. 2022, 11, 4195. [Google Scholar] [CrossRef] [PubMed]
- Aragona, P.; Baudouin, C.; Benitez Del Castillo, J.M.; Messmer, E.; Barabino, S.; Merayo-Lloves, J.; Brignole-Baudouin, F.; Inferrera, L.; Rolando, M.; Mencucci, R.; et al. The ocular microbiome and microbiota and their effects on ocular surface pathophysiology and disorders. Surv. Ophthalmol. 2021, 66, 907–925. [Google Scholar] [CrossRef] [PubMed]
- Petrillo, F.; Pignataro, D.; Lavano, M.A.; Santella, B.; Folliero, V.; Zannella, C.; Astarita, C.; Gagliano, C.; Franci, G.; Avitabile, T.; et al. Current Evidence on the Ocular Surface Microbiota and Related Diseases. Microorganisms 2020, 8, 1033. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.J.; Liu, J. Human Microbiota and Ophthalmic Disease. Yale J. Biol. Med. 2016, 89, 325–330. [Google Scholar]
- Hilliam, Y.; Kaye, S.; Winstanley, C. Pseudomonas aeruginosa and microbial keratitis. J. Med. Microbiol. 2020, 69, 3–13. [Google Scholar] [CrossRef]
- Urwin, L.; Okurowska, K.; Crowther, G.; Roy, S.; Garg, P.; Karunakaran, E.; MacNeil, S.; Partridge, L.J.; Green, L.R.; Monk, P.N. Corneal Infection Models: Tools to Investigate the Role of Biofilms in Bacterial Keratitis. Cells 2020, 9, 2450. [Google Scholar] [CrossRef]
- Mohamed, S.; Elmohamady, M.N.; Abdelrahman, S.; Amer, M.M.; Abdelhamid, A.G. Antibacterial effects of antibiotics and cell-free preparations of probiotics against Staphylococcus aureus and Staphylococcus epidermidis associated with conjunctivitis. Saudi Pharm. J. 2020, 28, 1558–1565. [Google Scholar] [CrossRef] [PubMed]
- Chiang, M.C.; Chern, E. Ocular surface microbiota: Ophthalmic infectious disease and probiotics. Front. Microbiol. 2022, 13, 952473. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Langella, P. Emerging Health Concepts in the Probiotics Field: Streamlining the Definitions. Front. Microbiol. 2019, 10, 1047. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Vishakha, K.; Banerjee, S.; Bera, T.; Mondal, S.; Ganguli, A. A novel probiotic strain of Lactobacillus fermentum TIU19 isolated from Haria beer showing both in vitro antibacterial and antibiofilm properties upon two multi resistant uro-pathogen strains. Curr. Res. Microb. Sci. 2022, 3, 100150. [Google Scholar] [CrossRef] [PubMed]
- Tegegne, B.A.; Kebede, B. Probiotics, their prophylactic and therapeutic applications in human health development: A review of the literature. Heliyon 2022, 8, e09725. [Google Scholar] [CrossRef] [PubMed]
- Isenring, J.; Geirnaert, A.; Lacroix, C.; Stevens, M.J.A. Bistable auto-aggregation phenotype in Lactiplantibacillus plantarum emerges after cultivation in in vitro colonic microbiota. BMC Microbiol. 2021, 21, 268. [Google Scholar] [CrossRef]
- Nwoko, E.Q.A.; Okeke, I.N. Bacteria autoaggregation: How and why bacteria stick together. Biochem. Soc. Trans. 2021, 49, 1147–1157. [Google Scholar] [CrossRef]
- Tuo, Y.; Yu, H.; Ai, L.; Wu, Z.; Guo, B.; Chen, W. Aggregation and adhesion properties of 22 Lactobacillus strains. J. Dairy Sci. 2013, 96, 4252–4257. [Google Scholar] [CrossRef]
- Salas-Jara, M.J.; Ilabaca, A.; Vega, M.; Garcia, A. Biofilm Forming Lactobacillus: New Challenges for the Development of Probiotics. Microorganisms 2016, 4, 35. [Google Scholar] [CrossRef]
- Carvalho, F.M.; Teixeira-Santos, R.; Mergulhao, F.J.M.; Gomes, L.C. The Use of Probiotics to Fight Biofilms in Medical Devices: A Systematic Review and Meta-Analysis. Microorganisms 2020, 9, 27. [Google Scholar] [CrossRef]
- Bubnov, R.V.; Babenko, L.P.; Lazarenko, L.M.; Mokrozub, V.V.; Spivak, M.Y. Specific properties of probiotic strains: Relevance and benefits for the host. EPMA J. 2018, 9, 205–223. [Google Scholar] [CrossRef] [PubMed]
- Chisari, G.; Chisari, E.M.; Francaviglia, A.; Chisari, C.G. The mixture of bifidobacterium associated with fructo-oligosaccharides reduces the damage of the ocular surface. Clin. Ter. 2017, 168, e181–e185. [Google Scholar] [CrossRef] [PubMed]
- Iovieno, A.; Lambiase, A.; Sacchetti, M.; Stampachiacchiere, B.; Micera, A.; Bonini, S. Preliminary evidence of the efficacy of probiotic eye-drop treatment in patients with vernal keratoconjunctivitis. Graefes Arch. Clin. Exp. Ophthalmol. 2008, 246, 435–441. [Google Scholar] [CrossRef]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [PubMed]
- Mantelli, F.; Argueso, P. Functions of ocular surface mucins in health and disease. Curr. Opin. Allergy Clin. Immunol. 2008, 8, 477–483. [Google Scholar] [CrossRef]
- Mantelli, F.; Mauris, J.; Argueso, P. The ocular surface epithelial barrier and other mechanisms of mucosal protection: From allergy to infectious diseases. Curr. Opin. Allergy Clin. Immunol. 2013, 13, 563–568. [Google Scholar] [CrossRef]
- Hazlett, L.D. Corneal response to Pseudomonas aeruginosa infection. Prog. Retin. Eye Res. 2004, 23, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Bourges, J.L.; Torriglia, A.; Valamanesh, F.; Benezra, D.; Renard, G.; Behar-Cohen, F.F. Nitrosative stress and corneal transplant endothelial cell death during acute graft rejection. Transplantation 2007, 84, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Canto, A.; Olivar, T.; Romero, F.J.; Miranda, M. Nitrosative Stress in Retinal Pathologies: Review. Antioxidants 2019, 8, 543. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yuan, Q.; Chen, F.; Pang, J.; Pan, C.; Xu, F.; Chen, Y. Fundamental Mechanisms of the Cell Death Caused by Nitrosative Stress. Front. Cell Dev. Biol. 2021, 9, 742483. [Google Scholar] [CrossRef] [PubMed]
- Shan, Q.; Dwyer, M.; Rahman, S.; Gadjeva, M. Distinct susceptibilities of corneal Pseudomonas aeruginosa clinical isolates to neutrophil extracellular trap-mediated immunity. Infect. Immun. 2014, 82, 4135–4143. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Miller, D.; Iovieno, A. The role of microbial flora on the ocular surface. Curr. Opin. Allergy Clin. Immunol. 2009, 9, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Mshangila, B.; Paddy, M.; Kajumbula, H.; Ateenyi-Agaba, C.; Kahwa, B.; Seni, J. External ocular surface bacterial isolates and their antimicrobial susceptibility patterns among pre-operative cataract patients at Mulago National Hospital in Kampala, Uganda. BMC Ophthalmol. 2013, 13, 71. [Google Scholar] [CrossRef]
- Huang, F.C.; Lu, Y.T.; Liao, Y.H. Beneficial effect of probiotics on Pseudomonas aeruginosa-infected intestinal epithelial cells through inflammatory IL-8 and antimicrobial peptide human beta-defensin-2 modulation. Innate Immun. 2020, 26, 592–600. [Google Scholar] [CrossRef]
- Forestier, C.; Guelon, D.; Cluytens, V.; Gillart, T.; Sirot, J.; De Champs, C. Oral probiotic and prevention of Pseudomonas aeruginosa infections: A randomized, double-blind, placebo-controlled pilot study in intensive care unit patients. Crit. Care 2008, 12, R69. [Google Scholar] [CrossRef]
- Valdez, J.C.; Peral, M.C.; Rachid, M.; Santana, M.; Perdigon, G. Interference of Lactobacillus plantarum with Pseudomonas aeruginosa in vitro and in infected burns: The potential use of probiotics in wound treatment. Clin. Microbiol. Infect. 2005, 11, 472–479. [Google Scholar] [CrossRef]
- Milner, E.; Stevens, B.; An, M.; Lam, V.; Ainsworth, M.; Dihle, P.; Stearns, J.; Dombrowski, A.; Rego, D.; Segars, K. Utilizing Probiotics for the Prevention and Treatment of Gastrointestinal Diseases. Front. Microbiol. 2021, 12, 689958. [Google Scholar] [CrossRef] [PubMed]
- Castiblanco, G.A.; Yucel-Lindberg, T.; Roos, S.; Twetman, S. Effect of Lactobacillus reuteri on Cell Viability and PGE(2) Production in Human Gingival Fibroblasts. Probiotics Antimicrob. Proteins 2017, 9, 278–283. [Google Scholar] [CrossRef]
- Maldonado Galdeano, C.; Cazorla, S.I.; Lemme Dumit, J.M.; Velez, E.; Perdigon, G. Beneficial Effects of Probiotic Consumption on the Immune System. Ann. Nutr. Metab. 2019, 74, 115–124. [Google Scholar] [CrossRef]
- Maudsdotter, L.; Jonsson, H.; Roos, S.; Jonsson, A.B. Lactobacilli reduce cell cytotoxicity caused by Streptococcus pyogenes by producing lactic acid that degrades the toxic component lipoteichoic acid. Antimicrob. Agents Chemother. 2011, 55, 1622–1628. [Google Scholar] [CrossRef]
- Zhao, X.; Zhou, J.; Liang, W.; Sheng, Q.; Lu, L.; Chen, T.; Chen, J.; Tan, K.; Lv, Z. Probiotics mixture reinforces barrier function to ameliorate necrotizing enterocolitis by regulating PXR-JNK pathway. Cell Biosci. 2021, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- Sultana, R.; McBain, A.J.; O’Neill, C.A. Strain-dependent augmentation of tight-junction barrier function in human primary epidermal keratinocytes by Lactobacillus and Bifidobacterium lysates. Appl. Environ. Microbiol. 2013, 79, 4887–4894. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, W.; Wang, S.; Wang, Y.; Chu, X.; Ji, H. Lactobacillus plantarum Exhibits Antioxidant and Cytoprotective Activities in Porcine Intestinal Epithelial Cells Exposed to Hydrogen Peroxide. Oxid. Med. Cell Longev. 2021, 2021, 8936907. [Google Scholar] [CrossRef] [PubMed]
- Iorio, R.; Petricca, S.; Luzi, C.; Bellio, P.; Cristiano, L.; Festuccia, C.; Amicosante, G.; Celenza, G. Lactobacillus sakei Pro-Bio65 Reduces TNF-alpha Expression and Upregulates GSH Content and Antioxidant Enzymatic Activities in Human Conjunctival Cells. Transl. Vis. Sci. Technol. 2021, 10, 8. [Google Scholar] [CrossRef]
- Tavakoli, A.; Markoulli, M.; Papas, E.; Flanagan, J. The Impact of Probiotics and Prebiotics on Dry Eye Disease Signs and Symptoms. J. Clin. Med. 2022, 11, 4889. [Google Scholar] [CrossRef]
- Pena, J.A.; Rogers, A.B.; Ge, Z.; Ng, V.; Li, S.Y.; Fox, J.G.; Versalovic, J. Probiotic Lactobacillus spp. diminish Helicobacter hepaticus-induced inflammatory bowel disease in interleukin-10-deficient mice. Infect. Immun. 2005, 73, 912–920. [Google Scholar] [CrossRef]
- Oh, P.L.; Benson, A.K.; Peterson, D.A.; Patil, P.B.; Moriyama, E.N.; Roos, S.; Walter, J. Diversification of the gut symbiont Lactobacillus reuteri as a result of host-driven evolution. ISME J. 2010, 4, 377–387. [Google Scholar] [CrossRef]
- Hemarajata, P.; Gao, C.; Pflughoeft, K.J.; Thomas, C.M.; Saulnier, D.M.; Spinler, J.K.; Versalovic, J. Lactobacillus reuteri-specific immunoregulatory gene rsiR modulates histamine production and immunomodulation by Lactobacillus reuteri. J. Bacteriol. 2013, 195, 5567–5576. [Google Scholar] [CrossRef]
- Spinler, J.K.; Sontakke, A.; Hollister, E.B.; Venable, S.F.; Oh, P.L.; Balderas, M.A.; Saulnier, D.M.; Mistretta, T.A.; Devaraj, S.; Walter, J.; et al. From prediction to function using evolutionary genomics: Human-specific ecotypes of Lactobacillus reuteri have diverse probiotic functions. Genome Biol. Evol. 2014, 6, 1772–1789. [Google Scholar] [CrossRef]
- Mu, Q.; Tavella, V.J.; Luo, X.M. Role of Lactobacillus reuteri in Human Health and Diseases. Front. Microbiol. 2018, 9, 757. [Google Scholar] [CrossRef]
- Hou, C.; Zeng, X.; Yang, F.; Liu, H.; Qiao, S. Study and use of the probiotic Lactobacillus reuteri in pigs: A review. J. Anim. Sci. Biotechnol. 2015, 6, 14. [Google Scholar] [CrossRef] [PubMed]
- Li, X.J.; Yue, L.Y.; Guan, X.F.; Qiao, S.Y. The adhesion of putative probiotic lactobacilli to cultured epithelial cells and porcine intestinal mucus. J. Appl. Microbiol. 2008, 104, 1082–1091. [Google Scholar] [CrossRef] [PubMed]
- Blanco, G.; Ruiz, L.; Tames, H.; Ruas-Madiedo, P.; Fdez-Riverola, F.; Sanchez, B.; Lourenco, A.; Margolles, A. Revisiting the Metabolic Capabilities of Bifidobacterium longum susbp. longum and Bifidobacterium longum subsp. infantis from a Glycoside Hydrolase Perspective. Microorganisms 2020, 8, 723. [Google Scholar] [CrossRef] [PubMed]
- Chichlowski, M.; Shah, N.; Wampler, J.L.; Wu, S.S.; Vanderhoof, J.A. Bifidobacterium longum Subspecies infantis (B. infantis) in Pediatric Nutrition: Current State of Knowledge. Nutrients 2020, 12, 1581. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Han, S.Y.; Lee, J.; Kim, N.R.; Lee, B.R.; Kim, H.; Kwon, M.; Ahn, K.; Noh, Y.; Kim, S.J.; et al. Bifidobacterium longum and Galactooligosaccharide Improve Skin Barrier Dysfunction and Atopic Dermatitis-like Skin. Allergy Asthma Immunol. Res. 2022, 14, 549–564. [Google Scholar] [CrossRef] [PubMed]
- Quinn, E.M.; Slattery, H.; Walsh, D.; Joshi, L.; Hickey, R.M. Bifidobacterium longum subsp. infantis ATCC 15697 and Goat Milk Oligosaccharides Show Synergism In Vitro as Anti-Infectives against Campylobacter jejuni. Foods 2020, 9, 348. [Google Scholar] [CrossRef] [PubMed]
- Greppi, A.; Asare, P.T.; Schwab, C.; Zemp, N.; Stephan, R.; Lacroix, C. Isolation and Comparative Genomic Analysis of Reuterin-Producing Lactobacillus reuteri From the Chicken Gastrointestinal Tract. Front. Microbiol. 2020, 11, 1166. [Google Scholar] [CrossRef]
- Golowczyc, M.A.; Mobili, P.; Garrote, G.L.; Abraham, A.G.; De Antoni, G.L. Protective action of Lactobacillus kefir carrying S-layer protein against Salmonella enterica serovar Enteritidis. Int. J. Food Microbiol. 2007, 118, 264–273. [Google Scholar] [CrossRef]
- Speranza, B.; Liso, A.; Russo, V.; Corbo, M.R. Evaluation of the Potential of Biofilm Formation of Bifidobacterium longum subsp. infantis and Lactobacillus reuteri as Competitive Biocontrol Agents Against Pathogenic and Food Spoilage Bacteria. Microorganisms 2020, 8, 177. [Google Scholar] [CrossRef]
- Szewczyk, A.; Marino, A.; Taviano, M.F.; Cambria, L.; Davi, F.; Trepa, M.; Grabowski, M.; Miceli, N. Studies on the Accumulation of Secondary Metabolites and Evaluation of Biological Activity of In Vitro Cultures of Ruta montana L. in Temporary Immersion Bioreactors. Int. J. Mol. Sci. 2023, 24, 7045. [Google Scholar] [CrossRef]
- Cristaldi, M.; Olivieri, M.; Spampinato, G.; Anfuso, C.D.; Scalia, M.; Lupo, G.; Rusciano, D. Isolation and Characterization of a New Human Corneal Epithelial Cell Line: HCE-F. Cornea 2020, 39, 1419–1425. [Google Scholar] [CrossRef]
- Tuo, Y.F.; Zhang, L.W.; Yi, H.X.; Zhang, Y.C.; Zhang, W.Q.; Han, X.; Du, M.; Jiao, Y.H.; Wang, S.M. Short communication: Antiproliferative effect of wild Lactobacillus strains isolated from fermented foods on HT-29 cells. J. Dairy Sci. 2010, 93, 2362–2366. [Google Scholar] [CrossRef]
- Polak-Berecka, M.; Wasko, A.; Paduch, R.; Skrzypek, T.; Sroka-Bartnicka, A. The effect of cell surface components on adhesion ability of Lactobacillus rhamnosus. Antonie Van Leeuwenhoek 2014, 106, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Paterniti, I.; Cordaro, M.; Campolo, M.; Siracusa, R.; Cornelius, C.; Navarra, M.; Cuzzocrea, S.; Esposito, E. Neuroprotection by association of palmitoylethanolamide with luteolin in experimental Alzheimer’s disease models: The control of neuroinflammation. CNS Neurol. Disord. Drug Targets 2014, 13, 1530–1541. [Google Scholar] [CrossRef] [PubMed]
- Jung, I.G.; Jeong, J.Y.; Yum, S.H.; Hwang, Y.J. Inhibitory Effects of Selected Medicinal Plants on Bacterial Growth of Methicillin-Resistant Staphylococcus aureus. Molecules 2022, 27, 7780. [Google Scholar] [CrossRef] [PubMed]
- Gopal, P.K.; Prasad, J.; Smart, J.; Gill, H.S. In vitro adherence properties of Lactobacillus rhamnosus DR20 and Bifidobacterium lactis DR10 strains and their antagonistic activity against an enterotoxigenic Escherichia coli. Int. J. Food Microbiol. 2001, 67, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Alepee, N.; Grandidier, M.H.; Teluob, S.; Amaral, F.; Caviola, E.; De Servi, B.; Martin, S.; Meloni, M.; Nardelli, L.; Pasdelou, C.; et al. Validation of the SkinEthic HCE Time-to-Toxicity test method for eye hazard classification of chemicals according to UN GHS. Toxicol. Vitr. 2022, 80, 105319. [Google Scholar] [CrossRef]
- Mannino, D.; Scuderi, S.A.; Casili, G.; Bova, V.; Cucinotta, L.; Lanza, M.; Filippone, A.; Esposito, E.; Paterniti, I. Neuroprotective effects of GSK-343 in an in vivo model of MPTP-induced nigrostriatal degeneration. J. Neuroinflamm. 2023, 20, 155. [Google Scholar] [CrossRef] [PubMed]
- Khoh-Reiter, S.; Jessen, B.A. Evaluation of the cytotoxic effects of ophthalmic solutions containing benzalkonium chloride on corneal epithelium using an organotypic 3-D model. BMC Ophthalmol. 2009, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Schaefer, L.; Bian, F.; Yu, Z.; Pflugfelder, S.C.; Britton, R.A.; de Paiva, C.S. Dysbiosis Modulates Ocular Surface Inflammatory Response to Liposaccharide. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4224–4233. [Google Scholar] [CrossRef] [PubMed]
- Mohammedsaeed, W.; McBain, A.J.; Cruickshank, S.M.; O’Neill, C.A. Lactobacillus rhamnosus GG inhibits the toxic effects of Staphylococcus aureus on epidermal keratinocytes. Appl. Environ. Microbiol. 2014, 80, 5773–5781. [Google Scholar] [CrossRef]
- Paterniti, I.; Impellizzeri, D.; Cordaro, M.; Siracusa, R.; Bisignano, C.; Gugliandolo, E.; Carughi, A.; Esposito, E.; Mandalari, G.; Cuzzocrea, S. The Anti-Inflammatory and Antioxidant Potential of Pistachios (Pistacia vera L.) In Vitro and In Vivo. Nutrients 2017, 9, 915. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Contact Time | P.a. | P.a. + L.r. | P.a. + B.l |
|---|---|---|---|
| Short | 7.1 ± 0.2 | 6.4 ± 0.6 | 6.5 ± 0.1 |
| Long | 8.8 ± 0.4 | 8.7 ± 0.3 | 8.4 ± 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paterniti, I.; Scuderi, S.A.; Cambria, L.; Nostro, A.; Esposito, E.; Marino, A. Protective Effect of Probiotics against Pseudomonas aeruginosa Infection of Human Corneal Epithelial Cells. Int. J. Mol. Sci. 2024, 25, 1770. https://doi.org/10.3390/ijms25031770
Paterniti I, Scuderi SA, Cambria L, Nostro A, Esposito E, Marino A. Protective Effect of Probiotics against Pseudomonas aeruginosa Infection of Human Corneal Epithelial Cells. International Journal of Molecular Sciences. 2024; 25(3):1770. https://doi.org/10.3390/ijms25031770
Chicago/Turabian StylePaterniti, Irene, Sarah Adriana Scuderi, Lucia Cambria, Antonia Nostro, Emanuela Esposito, and Andreana Marino. 2024. "Protective Effect of Probiotics against Pseudomonas aeruginosa Infection of Human Corneal Epithelial Cells" International Journal of Molecular Sciences 25, no. 3: 1770. https://doi.org/10.3390/ijms25031770
APA StylePaterniti, I., Scuderi, S. A., Cambria, L., Nostro, A., Esposito, E., & Marino, A. (2024). Protective Effect of Probiotics against Pseudomonas aeruginosa Infection of Human Corneal Epithelial Cells. International Journal of Molecular Sciences, 25(3), 1770. https://doi.org/10.3390/ijms25031770