

Long Noncoding RNA VLDLR-AS1 Levels in Serum Correlate with Combat-Related Chronic Mild Traumatic Brain Injury and Depression Symptoms in US Veterans

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Identifying lncRNAs Consistently Detected in Serum
2.2. LncRNAs Are Packaged in Exosomes Derived from the Brain
2.3. LncRNA VLDLR-AS1 Levels Are Lower in Participants with Repetitive mTBI
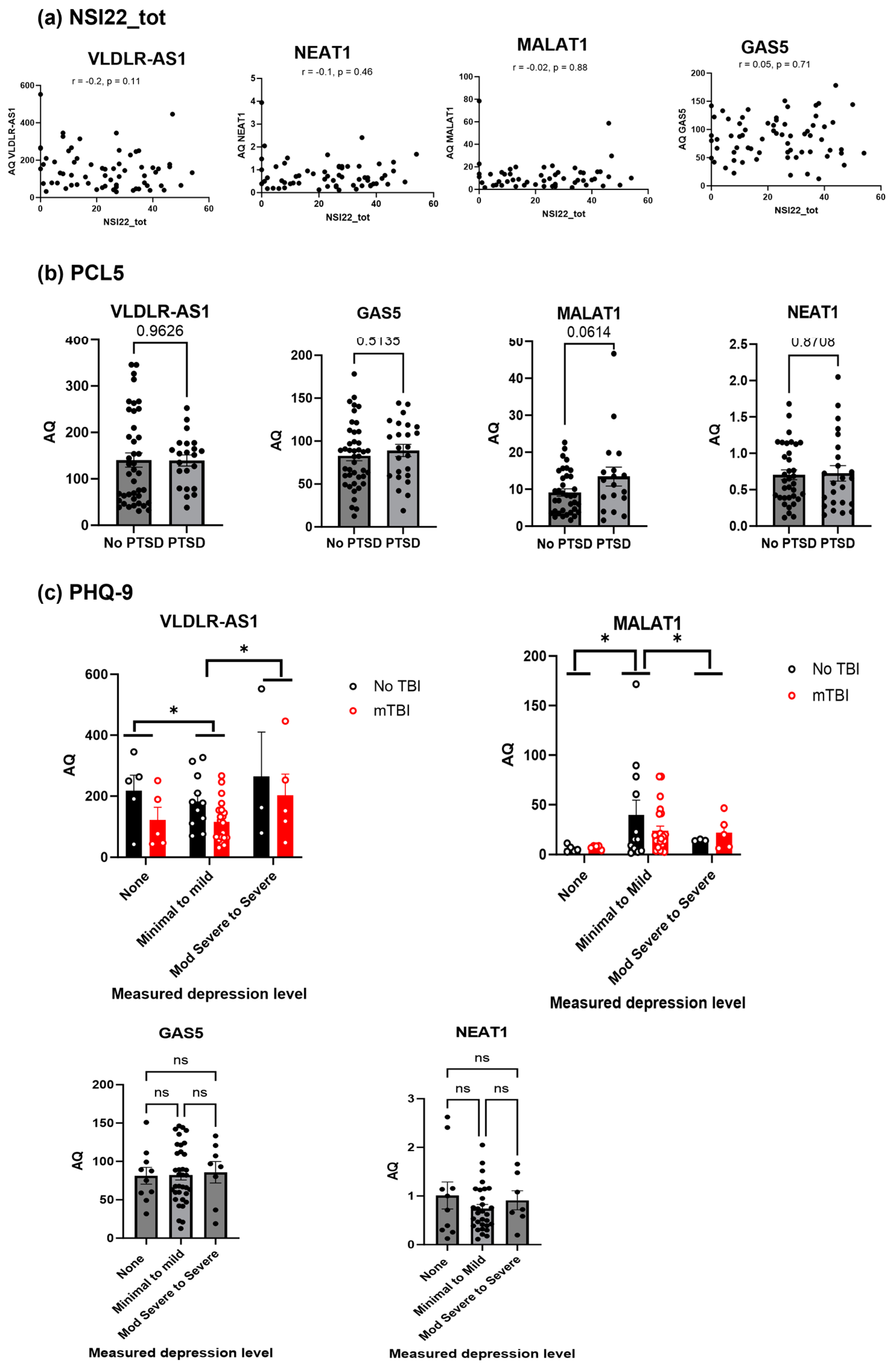
2.4. Secondary Analysis of the Clinical Data Shows VLDLR-AS1 and MALAT1 Levels Are Correlated with Depression
3. Discussion
Limitations
4. Materials and Methods
4.1. Study Samples
4.2. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
4.3. Droplet Digital Polymerase Chain Reaction (ddPCR)
4.4. CNS-Derived Exosome Purification
4.5. Neuropshycological Symptom Measures
4.6. Statistical Analyses Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dams-O’Connor, K.; Cuthbert, J.P.; Whyte, J.; Corrigan, J.D.; Faul, M.; Harrison-Felix, C. Traumatic brain injury among older adults at level I and II trauma centers. J. Neurotrauma 2013, 30, 2001–2013. [Google Scholar] [CrossRef]
- Leo, P.; McCrea, M. Epidemiology. In Translational Research in Traumatic Brain Injury; Laskowitz, D., Grant, G., Eds.; CRC Press/Taylor and Francis Group: Boca Raton, FL, USA, 2016. [Google Scholar]
- Vas, A.; Chapman, S.; Aslan, S.; Spence, J.; Keebler, M.; Rodriguez-Larrain, G.; Rodgers, B.; Jantz, T.; Martinez, D.; Rakic, J.; et al. Reasoning training in veteran and civilian traumatic brain injury with persistent mild impairment. Neuropsychol. Rehabil. 2016, 26, 502–531. [Google Scholar] [CrossRef]
- Kornblith, E.S.; Yaffe, K.; Langa, K.M.; Gardner, R.C. Prevalence of Lifetime History of Traumatic Brain Injury among Older Male Veterans Compared with Civilians: A Nationally Representative Study. J. Neurotrauma 2020, 37, 2680–2685. [Google Scholar] [CrossRef]
- Lindberg, M.A.; Martin, E.M.M.; Marion, D.W. Military Traumatic Brain Injury: The History, Impact, and Future. J. Neurotrauma 2022, 39, 1133–1145. [Google Scholar] [CrossRef]
- Eibner, C.; Schell, T.L.; Jaycox, L.H. Care of war veterans with mild traumatic brain injury. N. Engl. J. Med. 2009, 361, 537. [Google Scholar]
- Dismuke-Greer, C.; Hirsch, S.; Carlson, K.; Pogoda, T.; Nakase-Richardson, R.; Bhatnagar, S.; Eapen, B.; Troyanskaya, M.; Miles, S.; Nolen, T.; et al. Health Services Utilization, Health Care Costs, and Diagnoses by Mild Traumatic Brain Injury Exposure: A Chronic Effects of Neurotrauma Consortium Study. Arch. Phys. Med. Rehabil. 2020, 101, 1720–1730. [Google Scholar] [CrossRef]
- The Management of Concussion-mild Traumatic Brain Injury Working Group. VA/DoD Clinical Practice Guideline for Management of Concussion/Mild Traumatic Brain Injury. J. Rehabil. Res. Dev. 2009, 46, CP1–CP68. [Google Scholar] [CrossRef]
- O’Neil, M.E.; Carlson, K.; Storzbach, D.; Brenner, L.; Freeman, M.; Quinones, A.; Motu’apuaka, M.; Ensley, M.; Kansagara, D. Complications of Mild Traumatic Brain Injury in Veterans and Military Personnel: A Systematic Review; Department of Veterans Affairs Health Services Research & Development Service: Washington, DC, USA, 2013. [Google Scholar]
- Corrigan, F.; Wee, I.C.; Collins-Praino, L.E. Chronic motor performance following different traumatic brain injury severity—A systematic review. Front. Neurol. 2023, 14, 1180353. [Google Scholar] [CrossRef]
- Taylor, C.A.; Bell, J.M.; Breiding, M.J.; Xu, L. Traumatic Brain Injury-Related Emergency Department Visits, Hospitalizations, and Deaths—United States, 2007 and 2013. MMWR Surveill. Summ. 2017, 66, 1–16. [Google Scholar] [CrossRef]
- Seabury, S.A.; Gaudette, E.; Goldman, D.P.; Markowitz, A.J.; Brooks, J.; McCrea, M.A.; Okonkwo, D.O.; Manley, G.T.; The TRACK-TBI Investigators. Assessment of Follow-up Care After Emergency Department Presentation for Mild Traumatic Brain Injury and Concussion: Results from the TRACK-TBI Study. JAMA Netw. Open 2018, 1, e180210. [Google Scholar] [CrossRef]
- de Souza, N.L.; Esopenko, C.; Jia, Y.; Parrott, J.S.; Merkley, T.L.; Dennis, E.L.; Hillary, F.G.; Velez, C.; Cooper, D.B.; Kennedy, J.E.; et al. Discriminating Mild Traumatic Brain Injury and Posttraumatic Stress Disorder Using Latent Neuroimaging and Neuropsychological Profiles in Active-Duty Military Service Members. J. Head Trauma Rehabil. 2023, 38, E254–E266. [Google Scholar] [CrossRef]
- Nelson, L.D.; Temkin, N.R.; Barber, J.; Brett, B.L.; Okonkwo, D.O.; McCrea, M.A.; Giacino, J.T.; Bodien, Y.G.; Robertson, C.; Corrigan, J.D.; et al. Functional Recovery, Symptoms, and Quality of Life 1 to 5 Years After Traumatic Brain Injury. JAMA Netw. Open 2023, 6, e233660. [Google Scholar] [CrossRef]
- Agtarap, S.; Hungerford, L.D.; Ettenhofer, M.L. Identifying Unique Symptom Groups Following Mild Traumatic Brain Injury Using the Neurobehavioral Symptom Inventory and PTSD Checklist-5 in Military Personnel: A Bifactor Analysis. J. Head Trauma Rehabil. 2023, 38, E371–E383. [Google Scholar] [CrossRef]
- Maas, A.I.R.; Menon, D.K.; Manley, G.T.; Abrams, M.; Akerlund, C.; Andelic, N.; Aries, M.; Bashford, T.; Bell, M.J.; Bodien, Y.G.; et al. Traumatic brain injury: Progress and challenges in prevention, clinical care, and research. Lancet Neurol. 2022, 21, 1004–1060. [Google Scholar] [CrossRef]
- Feinberg, C.; Mayes, K.D.; Portman, E.; Carr, C.; Mannix, R. Non-invasive fluid biomarkers in the diagnosis of mild traumatic brain injury (mTBI): A systematic review. J. Neurol. Neurosurg. Psychiatry 2023, 95, 184–192. [Google Scholar] [CrossRef]
- Newcombe, V.; Richter, S.; Whitehouse, D.P.; Bloom, B.M.; Lecky, F. Fluid biomarkers and neuroimaging in mild traumatic brain injury: Current uses and potential future directions for clinical use in emergency medicine. Emerg. Med. J. 2023, 40, 671–677. [Google Scholar] [CrossRef]
- Vedaei, F.; Mashhadi, N.; Zabrecky, G.; Monti, D.; Navarreto, E.; Hriso, C.; Wintering, N.; Newberg, A.B.; Mohamed, F.B. Identification of chronic mild traumatic brain injury using resting state functional MRI and machine learning techniques. Front. Neurosci. 2022, 16, 1099560. [Google Scholar] [CrossRef]
- Agoston, D.V.; Helmy, A. Fluid-Based Protein Biomarkers in Traumatic Brain Injury: The View from the Bedside. Int. J. Mol. Sci. 2023, 24, 16267. [Google Scholar] [CrossRef]
- Wang, K.K.; Munoz Pareja, J.C.; Mondello, S.; Diaz-Arrastia, R.; Wellington, C.; Kenney, K.; Puccio, A.M.; Hutchison, J.; McKinnon, N.; Okonkwo, D.O.; et al. Blood-based traumatic brain injury biomarkers—Clinical utilities and regulatory pathways in the United States, Europe and Canada. Expert. Rev. Mol. Diagn. 2021, 21, 1303–1321. [Google Scholar] [CrossRef]
- Biberthaler, P.; Musaelyan, K.; Krieg, S.; Meyer, B.; Stimmer, H.; Zapf, J.; von Matthey, F.; Chandran, R.; Marino, J.A.; Beligere, G.; et al. Evaluation of Acute Glial Fibrillary Acidic Protein and Ubiquitin C-Terminal Hydrolase-L1 Plasma Levels in Traumatic Brain Injury Patients with and without Intracranial Lesions. Neurotrauma Rep. 2021, 2, 617–625. [Google Scholar] [CrossRef]
- Boucher, V.; Frenette, J.; Neveu, X.; Tardif, P.A.; Mercier, E.; Chauny, J.M.; Berthelot, S.; Archambault, P.; Lee, J.; Perry, J.J.; et al. Lack of association between four biomarkers and persistent post-concussion symptoms after a mild traumatic brain injury. J. Clin. Neurosci. 2023, 118, 34–43. [Google Scholar] [CrossRef]
- Beylerli, O.; Tamrazov, R.; Gareev, I.; Ilyasova, T.; Shumadalova, A.; Bai, Y.; Yang, B. Role of exosomal ncRNAs in traumatic brain injury. Noncoding RNA Res. 2023, 8, 686–692. [Google Scholar] [CrossRef]
- Wapinski, O.; Chang, H.Y. Long noncoding RNAs and human disease. Trends Cell Biol. 2011, 21, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Dieter, C.; Lourenco, E.D.; Lemos, N.E. Association of long non-coding RNA and leukemia: A systematic review. Gene 2020, 735, 144405. [Google Scholar] [CrossRef]
- He, Z.; Yang, D.; Fan, X.; Zhang, M.; Li, Y.; Gu, X.; Yang, M. The Roles and Mechanisms of lncRNAs in Liver Fibrosis. Int. J. Mol. Sci. 2020, 21, 1482. [Google Scholar] [CrossRef]
- Moore, J.B., 4th; Uchida, S. Functional characterization of long noncoding RNAs. Curr. Opin. Cardiol. 2020, 35, 199–206. [Google Scholar] [CrossRef]
- Gao, S.; Fan, C.; Wang, Y.; Yang, W.; Jiang, H. LncRNA ENST00000440246.1 Promotes Alzheimer’s Disease Progression by Targeting PP2A. Biochem. Genet. 2023. [Google Scholar] [CrossRef]
- Ghamari, M.; Mohseni, M.M.; Taheri, M.; Neishabouri, S.M.; Shirvani-Farsani, Z. Abnormal expression of long non-coding RNAs RMRP, CTC-487M23.5, and DGCR5 in the peripheral blood of patients with Bipolar disorder. Metab. Brain Dis. 2023. [Google Scholar] [CrossRef]
- Greco, S.; Made, A.; Mutoli, M.; Zhang, L.; Piella, S.N.; Vausort, M.; Lumley, A.I.; Beltrami, A.P.; Srivastava, P.K.; Milani, V.; et al. HCG18, LEF1AS1 and lncCEACAM21 as biomarkers of disease severity in the peripheral blood mononuclear cells of COVID-19 patients. J. Transl. Med. 2023, 21, 758. [Google Scholar] [CrossRef]
- Qiu, T.; Xue, M.; Li, X.; Li, F.; Liu, S.; Yao, C.; Chen, W. Comparative evaluation of long non-coding RNA-based biomarkers in the urinary sediment and urinary exosomes for non-invasive diagnosis of bladder cancer. Mol. Omics 2022, 18, 938–947. [Google Scholar] [CrossRef]
- Sun, Z.; Wu, J.; Bi, Q.; Wang, W. Exosomal lncRNA TUG1 derived from human urine-derived stem cells attenuates renal ischemia/reperfusion injury by interacting with SRSF1 to regulate ASCL4-mediated ferroptosis. Stem Cell Res. Ther. 2022, 13, 297. [Google Scholar] [CrossRef]
- Lopes, C.; Chaves, J.; Ortigao, R.; Dinis-Ribeiro, M.; Pereira, C. Gastric cancer detection by non-blood-based liquid biopsies: A systematic review looking into the last decade of research. United Eur. Gastroenterol. J. 2023, 11, 114–130. [Google Scholar] [CrossRef]
- Jin, H.; Du, W.; Huang, W.; Yan, J.; Tang, Q.; Chen, Y.; Zou, Z. lncRNA and breast cancer: Progress from identifying mechanisms to challenges and opportunities of clinical treatment. Mol. Ther. Nucleic Acids 2021, 25, 613–637. [Google Scholar] [CrossRef]
- Li, Z.; Cai, S.; Li, H.; Gu, J.; Tian, Y.; Cao, J.; Yu, D.; Tang, Z. Developing a lncRNA Signature to Predict the Radiotherapy Response of Lower-Grade Gliomas Using Co-expression and ceRNA Network Analysis. Front. Oncol. 2021, 11, 622880. [Google Scholar] [CrossRef] [PubMed]
- Kucukakcali, Z.; Colak, C.; Gozukara Bag, H.G.; Balikci Cicek, I.; Ozhan, O.; Yildiz, A.; Danis, N.; Koc, A.; Parlakpinar, H.; Akbulut, S. Modeling Based on Ensemble Learning Methods for Detection of Diagnostic Biomarkers from LncRNA Data in Rats Treated with Cis-Platinum-Induced Hepatotoxicity. Diagnostics 2023, 13, 1583. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Zhang, X.; Cai, Z.; Xi, Z.; Wang, F.; Wang, X.; Li, W.; Dai, P. Identification of novel lncRNA prognostic biomarkers and their associated ceRNAs in bladder urothelial carcinoma. J. Biochem. Mol. Toxicol. 2023, 37, e23441. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, J.; Yu, S.; Wang, Z.; He, X.; Su, Y.; Guo, T.; Sheng, H.; Chen, J.; Zheng, Q.; et al. Extracellular Vesicles Long RNA Sequencing Reveals Abundant mRNA, circRNA, and lncRNA in Human Blood as Potential Biomarkers for Cancer Diagnosis. Clin. Chem. 2019, 65, 798–808. [Google Scholar] [CrossRef] [PubMed]
- Vanderploeg, R.D.; Cooper, D.B.; Belanger, H.G.; Donnell, A.J.; Kennedy, J.E.; Hopewell, C.A.; Scott, S.G. Screening for postdeployment conditions: Development and cross-validation of an embedded validity scale in the neurobehavioral symptom inventory. J. Head Trauma Rehabil. 2014, 29, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Belanger, H.G.; Silva, M.A.; Donnell, A.J.; McKenzie-Hartman, T.; Lamberty, G.J.; Vanderploeg, R.D. Utility of the Neurobehavioral Symptom Inventory As an Outcome Measure: A VA TBI Model Systems Study. J. Head Trauma Rehabil. 2017, 32, 46–54. [Google Scholar] [CrossRef]
- Silva, M.A. Review of the Neurobehavioral Symptom Inventory. Rehabil. Psychol. 2021, 66, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Soble, J.R.; Silva, M.A.; Vanderploeg, R.D.; Curtiss, G.; Belanger, H.G.; Donnell, A.J.; Scott, S.G. Normative Data for the Neurobehavioral Symptom Inventory (NSI) and post-concussion symptom profiles among TBI, PTSD, and nonclinical samples. Clin. Neuropsychol. 2014, 28, 614–632. [Google Scholar] [CrossRef] [PubMed]
- Kroenke, K.; Spitzer, R.L.; Williams, J.B. The PHQ-9: Validity of a brief depression severity measure. J. Gen. Intern. Med. 2001, 16, 606–613. [Google Scholar] [CrossRef]
- You, Y.; Wang, W.; Zhu, W.; Xu, J. Identification of functional lncRNAs in atrial fibrillation based on RNA sequencing. BMC Cardiovasc. Disord. 2023, 23, 539. [Google Scholar] [CrossRef]
- Xiong, L.; Tang, M.; Xing, S.; Yang, X. The role of noncoding RNA and its diagnostic potential in intrahepatic cholestasis of pregnancy: A research update. Front. Genet. 2023, 14, 1239693. [Google Scholar] [CrossRef]
- Xie, Y.; Ye, J.; Luo, H. HOXC Cluster Antisense RNA 3, a Novel Long Non-Coding RNA as an Oncological Biomarker and Therapeutic Target in Human Malignancies. Onco Targets Ther. 2023, 16, 849–865. [Google Scholar] [CrossRef]
- Wang, L.; Xu, D. Regulation of long noncoding RNAs in the pathogenesis and clinical implications of pituitary adenomas. Immun. Inflamm. Dis. 2023, 11, e1047. [Google Scholar] [CrossRef]
- Cheng, C.; Liu, Z.; Liu, D.; Chen, H.; Wang, Y.; Sun, B. LncRNA CCAT1 participates in pancreatic ductal adenocarcinoma progression by forming a positive feedback loop with c-Myc. Carcinogenesis 2023. [Google Scholar] [CrossRef]
- Carter, G.; Miladinovic, B.; Patel, A.A.; Deland, L.; Mastorides, S.; Patel, N.A. Circulating long noncoding RNA GAS5 levels are correlated to prevalence of type 2 diabetes mellitus. BBA Clin. 2015, 4, 102–107. [Google Scholar] [CrossRef]
- Beylerli, O.; Gareev, I.; Sufianov, A.; Ilyasova, T.; Guang, Y. Long noncoding RNAs as promising biomarkers in cancer. Noncoding RNA Res. 2022, 7, 66–70. [Google Scholar] [CrossRef]
- Ransohoff, J.D.; Wei, Y.; Khavari, P.A. The functions and unique features of long intergenic non-coding RNA. Nat. Rev. Mol. Cell Biol. 2018, 19, 143–157. [Google Scholar] [CrossRef]
- Ang, C.E.; Trevino, A.E.; Chang, H.Y. Diverse lncRNA mechanisms in brain development and disease. Curr. Opin. Genet. Dev. 2020, 65, 42–46. [Google Scholar] [CrossRef]
- Samaddar, S.; Banerjee, S. Far from the nuclear crowd: Cytoplasmic lncRNA and their implications in synaptic plasticity and memory. Neurobiol. Learn. Mem. 2021, 185, 107522. [Google Scholar] [CrossRef]
- Srinivas, T.; Siqueira, E.; Guil, S. Techniques for investigating lncRNA transcript functions in neurodevelopment. Mol. Psychiatry 2023. [Google Scholar] [CrossRef] [PubMed]
- Glasgow, A.M.A.; De Santi, C.; Greene, C.M. Non-coding RNA in cystic fibrosis. Biochem. Soc. Trans. 2018, 46, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Magee, P.; Fassan, M.; Sahoo, S.; Leong, H.S.; Lee, D.; Sellers, R.; Brulle-Soumare, L.; Cairo, S.; Monteverde, T.; et al. A KRAS-responsive long non-coding RNA controls microRNA processing. Nat. Commun. 2021, 12, 2038. [Google Scholar] [CrossRef] [PubMed]
- Aliperti, V.; Skonieczna, J.; Cerase, A. Long Non-Coding RNA (lncRNA) Roles in Cell Biology, Neurodevelopment and Neurological Disorders. Noncoding RNA 2021, 7, 36. [Google Scholar] [CrossRef]
- Takahashi, K.; Yan, I.K.; Wood, J.; Haga, H.; Patel, T. Involvement of extracellular vesicle long noncoding RNA (linc-VLDLR) in tumor cell responses to chemotherapy. Mol. Cancer Res. 2014, 12, 1377–1387. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Guo, J.; Zhang, H.; Cao, B.; Xu, G.; Zhang, Z.; Tong, J. Four Prognosis-Associated lncRNAs Serve as Biomarkers in Ovarian Cancer. Front. Genet. 2021, 12, 672674. [Google Scholar] [CrossRef]
- Liu, H.; Zhou, T.; Wang, B.; Li, L.; Ye, D.; Yu, S. Identification and functional analysis of a potential key lncRNA involved in fat loss of cancer cachexia. J. Cell Biochem. 2018, 119, 1679–1688. [Google Scholar] [CrossRef]
- Rouillard, A.D.; Hurle, M.R.; Agarwal, P. Systematic interrogation of diverse Omic data reveals interpretable, robust, and generalizable transcriptomic features of clinically successful therapeutic targets. PLoS Comput. Biol. 2018, 14, e1006142. [Google Scholar] [CrossRef]
- Rouillard, A.D.; Gundersen, G.W.; Fernandez, N.F.; Wang, Z.; Monteiro, C.D.; McDermott, M.G.; Ma’ayan, A. The harmonizome: A collection of processed datasets gathered to serve and mine knowledge about genes and proteins. Database 2016, 2016, baw100. [Google Scholar] [CrossRef]
- Ros, G.; Pegoraro, S.; De Angelis, P.; Sgarra, R.; Zucchelli, S.; Gustincich, S.; Manfioletti, G. HMGA2 Antisense Long Non-coding RNAs as New Players in the Regulation of HMGA2 Expression and Pancreatic Cancer Promotion. Front. Oncol. 2019, 9, 1526. [Google Scholar] [CrossRef]
- Shi, Y.; Parag, S.; Patel, R.; Lui, A.; Murr, M.; Cai, J.; Patel, N.A. Stabilization of lncRNA GAS5 by a Small Molecule and Its Implications in Diabetic Adipocytes. Cell Chem. Biol. 2019, 26, 319–330.e6. [Google Scholar] [CrossRef]
- Powell, J.M.; Ferraro, J.V.; Dikmen, S.S.; Temkin, N.R.; Bell, K.R. Accuracy of mild traumatic brain injury diagnosis. Arch. Phys. Med. Rehabil. 2008, 89, 1550–1555. [Google Scholar] [CrossRef] [PubMed]
- Pape, T.L.B.; Smith, B.; Babcock-Parziale, J.; Evans, C.T.; Herrold, A.A.; Maieritsch, K.P.; High, W.M., Jr. Diagnostic Accuracy of the Veteran Affairs’ Traumatic Brain Injury Screen. Arch. Phys. Med. Rehabil. 2018, 99, 1370–1382. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.H.; Powell-Cope, G.; Belanger, H.G. The Veterans Health Administration’s Traumatic Brain Injury Screen and Evaluation: Service Delivery Insights. Mil. Med. 2018, 183, e494–e501. [Google Scholar] [CrossRef]
- Kohler, M.J.; Hendrickx, M.D.; Powell-Jones, A.; Bryan-Hancock, C. A Systematic Review of Cognitive Functioning After Traumatic Brain Injury in Individuals Aged 10–30 Years. Cogn. Behav. Neurol. 2020, 33, 233–252. [Google Scholar] [CrossRef]
- Kenney, K.; Qu, B.X.; Lai, C.; Devoto, C.; Motamedi, V.; Walker, W.C.; Levin, H.S.; Nolen, T.; Wilde, E.A.; Diaz-Arrastia, R.; et al. Higher exosomal phosphorylated tau and total tau among veterans with combat-related repetitive chronic mild traumatic brain injury. Brain Inj. 2018, 32, 1276–1284. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.S.; Carter, G.; El Bassit, G.; Patel, A.A.; Cooper, D.R.; Murr, M.; Patel, N.A. Adipose-derived stem cells from lean and obese humans show depot specific differences in their stem cell markers, exosome contents and senescence: Role of protein kinase C delta (PKCdelta) in adipose stem cell niche. Stem Cell Investig. 2016, 3, 2. [Google Scholar]
- Yang, N.; Li, S.; Li, G.; Zhang, S.; Tang, X.; Ni, S.; Jian, X.; Xu, C.; Zhu, J.; Lu, M. The role of extracellular vesicles in mediating progression, metastasis and potential treatment of hepatocellular carcinoma. Oncotarget 2017, 8, 3683–3695. [Google Scholar] [CrossRef]
- Ren, D.; Chen, W.; Cao, K.; Wang, Z.; Zheng, P. Expression Profiles of Long Non-coding RNA and Messenger RNA in Human Traumatic Brain Injury. Mol. Ther. Nucleic Acids 2020, 22, 99–113. [Google Scholar] [CrossRef]
- Meng, J.; Ding, T.; Chen, Y.; Long, T.; Xu, Q.; Lian, W.; Liu, W. LncRNA-Meg3 promotes Nlrp3-mediated microglial inflammation by targeting miR-7a-5p. Int. Immunopharmacol. 2021, 90, 107141. [Google Scholar] [CrossRef]
- Li, Z.; Han, K.; Zhang, D.; Chen, J.; Xu, Z.; Hou, L. The role of long noncoding RNA in traumatic brain injury. Neuropsychiatr. Dis. Treat. 2019, 15, 1671–1677. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Gou, Y.; Jiang, X.; Wang, S.; Wang, R.; Liang, C.; Yang, G.; Wang, T.; Yu, A.; Zhu, G. Long Non-coding RNAs in Traumatic Brain Injury Accelerated Fracture Healing. Front. Surg. 2021, 8, 663377. [Google Scholar] [CrossRef]
- Cheng, S.; Zhang, Y.; Chen, S.; Zhou, Y. LncRNA HOTAIR Participates in Microglia Activation and Inflammatory Factor Release by Regulating the Ubiquitination of MYD88 in Traumatic Brain Injury. J. Mol. Neurosci. 2021, 71, 169–177. [Google Scholar] [CrossRef]
- Li, S.; Qiu, N.; Ni, A.; Hamblin, M.H.; Yin, K.J. Role of regulatory non-coding RNAs in traumatic brain injury. Neurochem. Int. 2023, 172, 105643. [Google Scholar] [CrossRef]
- Smith, C.M.; Steitz, J.A. Classification of gas5 as a multi-small-nucleolar-RNA (snoRNA) host gene and a member of the 5′-terminal oligopyrimidine gene family reveals common features of snoRNA host genes. Mol. Cell. Biochem. 1998, 18, 6897–6909. [Google Scholar] [CrossRef]
- Coccia, E.M.; Cicala, C.; Charlesworth, A.; Ciccarelli, C.; Rossi, G.B.; Philipson, L.; Sorrentino, V. Regulation and expression of a growth arrest-specific gene (gas5) during growth, differentiation, and development. Mol. Cell. Biochem. 1992, 12, 3514–3521. [Google Scholar]
- Ferguson, S.; McCartan, R.; Browning, M.; Hahn-Townsend, C.; Gratkowski, A.; Morin, A.; Abdullah, L.; Ait-Ghezala, G.; Ojo, J.; Sullivan, K.; et al. Impact of gulf war toxic exposures after mild traumatic brain injury. Acta Neuropathol. Commun. 2022, 10, 147. [Google Scholar] [CrossRef]
- Kryza-Lacombe, M.; Santiago, R.; Hwang, A.; Raptentsetsang, S.; Maruyama, B.A.; Chen, J.; Cassar, M.; Abrams, G.; Novakovic-Agopian, T.; Mukherjee, P. Resting-State Connectivity Changes After Goal-Oriented Attentional Self-Regulation Training in Veterans With Mild Traumatic Brain Injury: Preliminary Findings from a Randomized Controlled Trial. Neurotrauma Rep. 2023, 4, 420–432. [Google Scholar] [CrossRef]
- Martindale, S.L.; Vujanovic, A.A.; Ord, A.S.; Cary, A.; Rowland, J.A. Distress tolerance mitigates effects of posttraumatic stress, traumatic brain injury, and blast exposure on psychiatric and health outcomes. Rehabil. Psychol. 2023, 68, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Pickett, T.C.; Walker, W.C.; Lippa, S.M.; Lange, R.T.; Brickell, T.A.; Dittmer, T.A.; Smith, J.M.; Cifu, D.X.; French, L.M. Cross-Walk Comparison of the DVBIC-TBICoE and LIMBIC-CENC Combat-Related Concussion Prospective Longitudinal Study Datasets. Arch. Phys. Med. Rehabil. 2023, 104, 1072–1080.e1. [Google Scholar] [CrossRef] [PubMed]
- Walker, W.C.; Clark, S.W.; Eppich, K.; Wilde, E.A.; Martin, A.M.; Allen, C.M.; Cortez, M.M.; Pugh, M.J.; Walton, S.R.; Kenney, K. Headache among combat-exposed veterans and service members and its relation to mild traumatic brain injury history and other factors: A LIMBIC-CENC study. Front. Neurol. 2023, 14, 1242871. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, K.A.; Lurie, J.K. Principal components analysis of the Neurobehavioral Symptom Inventory in a nonclinical civilian sample. Appl. Neuropsychol. Adult 2017, 24, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Esterov, D.; Lennon, R.J.; Bergquist, T.; Brown, A. Predictors of neurobehavioral symptom reporting in a community based sample with mild traumatic brain injury. NeuroRehabilitation 2020, 47, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Cole, W.R.; Cecchini, A.S.; Remigio-Baker, R.A.; Gregory, E.; Bailie, J.M.; Ettenhofer, M.L.; McCulloch, K.L. “Return to duty” as an outcome metric in military concussion research: Problems, pitfalls, and potential solutions. Clin. Neuropsychol. 2020, 34, 1156–1174. [Google Scholar] [CrossRef] [PubMed]
- Lind, P.A.; Zhu, G.; Montgomery, G.W.; Madden, P.A.; Heath, A.C.; Martin, N.G.; Slutske, W.S. Genome-wide association study of a quantitative disordered gambling trait. Addict. Biol. 2013, 18, 511–522. [Google Scholar] [CrossRef]
- Wang, L.; Li, S.; Stone, S.S.; Liu, N.; Gong, K.; Ren, C.; Sun, K.; Zhang, C.; Shao, G. The Role of the lncRNA MALAT1 in Neuroprotection against Hypoxic/Ischemic Injury. Biomolecules 2022, 12, 146. [Google Scholar] [CrossRef]
- Patel, N.A.; Moss, L.D.; Lee, J.Y.; Tajiri, N.; Acosta, S.; Hudson, C.; Parag, S.; Cooper, D.R.; Borlongan, C.V.; Bickford, P.C. Long noncoding RNA MALAT1 in exosomes drives regenerative function and modulates inflammation-linked networks following traumatic brain injury. J. Neuroinflamm. 2018, 15, 204. [Google Scholar] [CrossRef]
- Qiu, J.J.; Lin, X.J.; Tang, X.Y.; Zheng, T.T.; Lin, Y.Y.; Hua, K.Q. Exosomal Metastasis-Associated Lung Adenocarcinoma Transcript 1 Promotes Angiogenesis and Predicts Poor Prognosis in Epithelial Ovarian Cancer. Int. J. Biol. Sci. 2018, 14, 1960–1973. [Google Scholar] [CrossRef]
- Pan, Y.; Tong, S.; Cui, R.; Fan, J.; Liu, C.; Lin, Y.; Tang, J.; Xie, H.; Lin, P.; Zheng, T.; et al. Long Non-Coding MALAT1 Functions as a Competing Endogenous RNA to Regulate Vimentin Expression by Sponging miR-30a-5p in Hepatocellular Carcinoma. Cell Physiol. Biochem. 2018, 50, 108–120. [Google Scholar] [CrossRef]
- Huang, J.; Zheng, Y.; Xiao, X.; Liu, C.; Lin, J.; Zheng, S.; Yang, B.; Ou, Q. A Circulating Long Noncoding RNA Panel Serves as a Diagnostic Marker for Hepatocellular Carcinoma. Dis. Markers 2020, 2020, 5417598. [Google Scholar] [CrossRef]
- Hu, Y.; Zheng, L.; Zhang, J.; Shen, Y.; Zhang, X.; Lin, L. LncRNA-MALAT1 is a promising biomarker for prognostic evaluation of tongue squamous cell carcinoma. Eur. Arch. Otorhinolaryngol. 2020, 277, 3155–3160. [Google Scholar] [CrossRef]
- de Aguiar Neto, F.S.; Rosa, J.L.G. Depression biomarkers using non-invasive EEG: A review. Neurosci. Biobehav. Rev. 2019, 105, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Harsanyi, S.; Kupcova, I.; Danisovic, L.; Klein, M. Selected Biomarkers of Depression: What Are the Effects of Cytokines and Inflammation? Int. J. Mol. Sci. 2022, 24, 578. [Google Scholar] [CrossRef] [PubMed]
- Arriaga-Canon, C.; Contreras-Espinosa, L.; Aguilar-Villanueva, S.; Bargallo-Rocha, E.; Garcia-Gordillo, J.A.; Cabrera-Galeana, P.; Castro-Hernandez, C.; Jimenez-Trejo, F.; Herrera, L.A. The Clinical Utility of lncRNAs and Their Application as Molecular Biomarkers in Breast Cancer. Int. J. Mol. Sci. 2023, 24, 7426. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.; Apostolatos, A.; Patel, R.; Mathur, A.; Cooper, D.; Murr, M.; Patel, N.A. Dysregulated Alternative Splicing Pattern of PKC during Differentiation of Human Preadipocytes Represents Distinct Differences between Lean and Obese Adipocytes. ISRN Obes. 2013, 2013, 9. [Google Scholar]
- Coulter, S.J. Mitigation of the effect of variability in digital PCR assays through use of duplexed reference assays for normalization. Biotechniques 2018, 65, 86–91. [Google Scholar] [CrossRef]
- King, P.R.; Donnelly, K.T.; Donnelly, J.P.; Dunnam, M.; Warner, G.; Kittleson, C.J.; Bradshaw, C.B.; Alt, M.; Meier, S.T. Psychometric study of the Neurobehavioral Symptom Inventory. J. Rehabil. Res. Dev. 2012, 49, 879–888. [Google Scholar] [CrossRef]
- Blevins, C.A.; Weathers, F.W.; Davis, M.T.; Witte, T.K.; Domino, J.L. The Posttraumatic Stress Disorder Checklist for DSM-5 (PCL-5): Development and Initial Psychometric Evaluation. J. Trauma Stress 2015, 28, 489–498. [Google Scholar] [CrossRef]
- Budczies, J.; Klauschen, F.; Sinn, B.V.; Gyorffy, B.; Schmitt, W.D.; Darb-Esfahani, S.; Denkert, C. Cutoff Finder: A comprehensive and straightforward Web application enabling rapid biomarker cutoff optimization. PLoS ONE 2012, 7, e51862. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Study Group | |
|---|---|---|
| No TBI (No. = 24) | mTBI (No. = 43) | |
| Age, mean (SD) (year) | 41.0 (12.7) | 41.0 (12.7) |
| Male, No. (%) | 19 (79.2) | 34 (79.1) |
| Racial background, No. (%) | ||
| White | 16 (66.6) | 33 (76.7) |
| Black | 8 (33.3) | 10 (23.3) |
| Education, No. (%) | ||
| High school graduate or GED | 3 (12.5) | 2 (4.7) |
| Some college or technical training | 9 (37.5) | 22 (51.2) |
| College graduate or higher | 12 (50.0) | 19 (44.2) |
| Number of TBI, mean (SD) | 0 | 2.8 (2.1) |
| Number of blast TBI, mean (SD) | 0 | 0.8 (1.0) |
| Number of general TBI, mean (SD) | 0 | 2.0 (1.6) |
| Years since first TBI, mean (SD) | 20.2 (12.2) | |
| Years since last TBI, mean (SD) | 9.6 (8.5) | |
| PHQ-9 total, mean (SD) | 4.0 (4.6) | 8.0 (5.6) |
| PCL-5 total, mean (SD) | 14.2 (15.4) | 26.5 (16.5) |
| NSI, mean (SD) | ||
| NSI total | 13.3 (13.1) | 28.1 (13.3) |
| Somatic | 3.0 (4.2) | 7.4 (4.1) |
| Affective | 5.8 (5.0) | 9.9 (5.1) |
| Cognitive | 2.8 (2.8) | 6.1 (3.7) |
| Vestibular | 1.0 (1.6) | 2.9 (2.2) |
| Number obese (BMI over 30), No. (%) | 11 (46.0) | 19 (44.0) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patel, R.S.; Krause-Hauch, M.; Kenney, K.; Miles, S.; Nakase-Richardson, R.; Patel, N.A. Long Noncoding RNA VLDLR-AS1 Levels in Serum Correlate with Combat-Related Chronic Mild Traumatic Brain Injury and Depression Symptoms in US Veterans. Int. J. Mol. Sci. 2024, 25, 1473. https://doi.org/10.3390/ijms25031473
Patel RS, Krause-Hauch M, Kenney K, Miles S, Nakase-Richardson R, Patel NA. Long Noncoding RNA VLDLR-AS1 Levels in Serum Correlate with Combat-Related Chronic Mild Traumatic Brain Injury and Depression Symptoms in US Veterans. International Journal of Molecular Sciences. 2024; 25(3):1473. https://doi.org/10.3390/ijms25031473
Chicago/Turabian StylePatel, Rekha S., Meredith Krause-Hauch, Kimbra Kenney, Shannon Miles, Risa Nakase-Richardson, and Niketa A. Patel. 2024. "Long Noncoding RNA VLDLR-AS1 Levels in Serum Correlate with Combat-Related Chronic Mild Traumatic Brain Injury and Depression Symptoms in US Veterans" International Journal of Molecular Sciences 25, no. 3: 1473. https://doi.org/10.3390/ijms25031473
APA StylePatel, R. S., Krause-Hauch, M., Kenney, K., Miles, S., Nakase-Richardson, R., & Patel, N. A. (2024). Long Noncoding RNA VLDLR-AS1 Levels in Serum Correlate with Combat-Related Chronic Mild Traumatic Brain Injury and Depression Symptoms in US Veterans. International Journal of Molecular Sciences, 25(3), 1473. https://doi.org/10.3390/ijms25031473