
A Single-Cell Atlas of Porcine Skeletal Muscle Reveals Mechanisms That Regulate Intramuscular Adipogenesis

 ,
,
Abstract
1. Introduction
2. Results
2.1. SnRNA-Seq Identified Distinct Cell Populations in Longissimus Dorsi Muscle at Two Developmental Stages
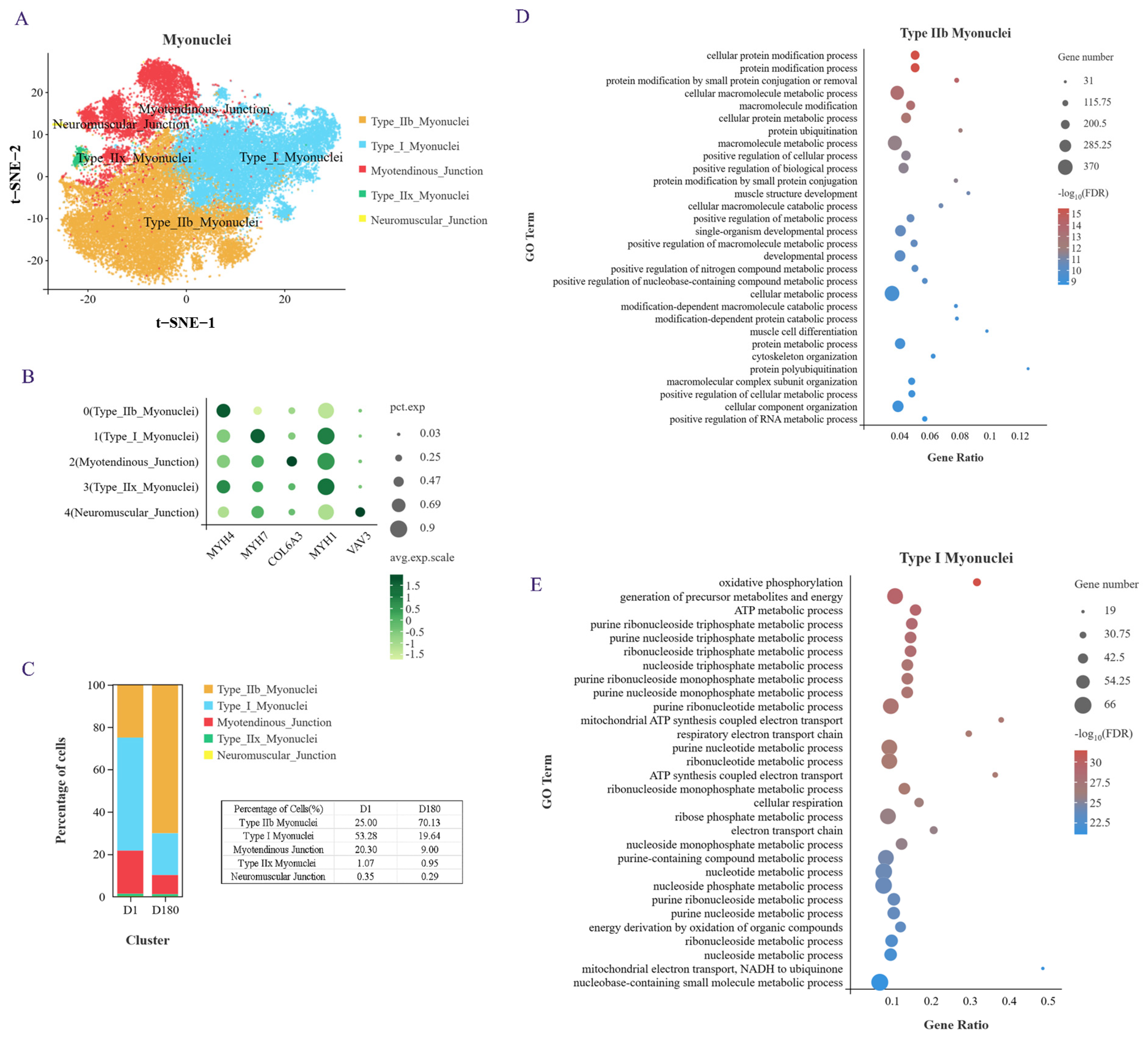
2.2. Clustering Analysis Identified Subgroups in the Subpopulation of Myonuclei Cells
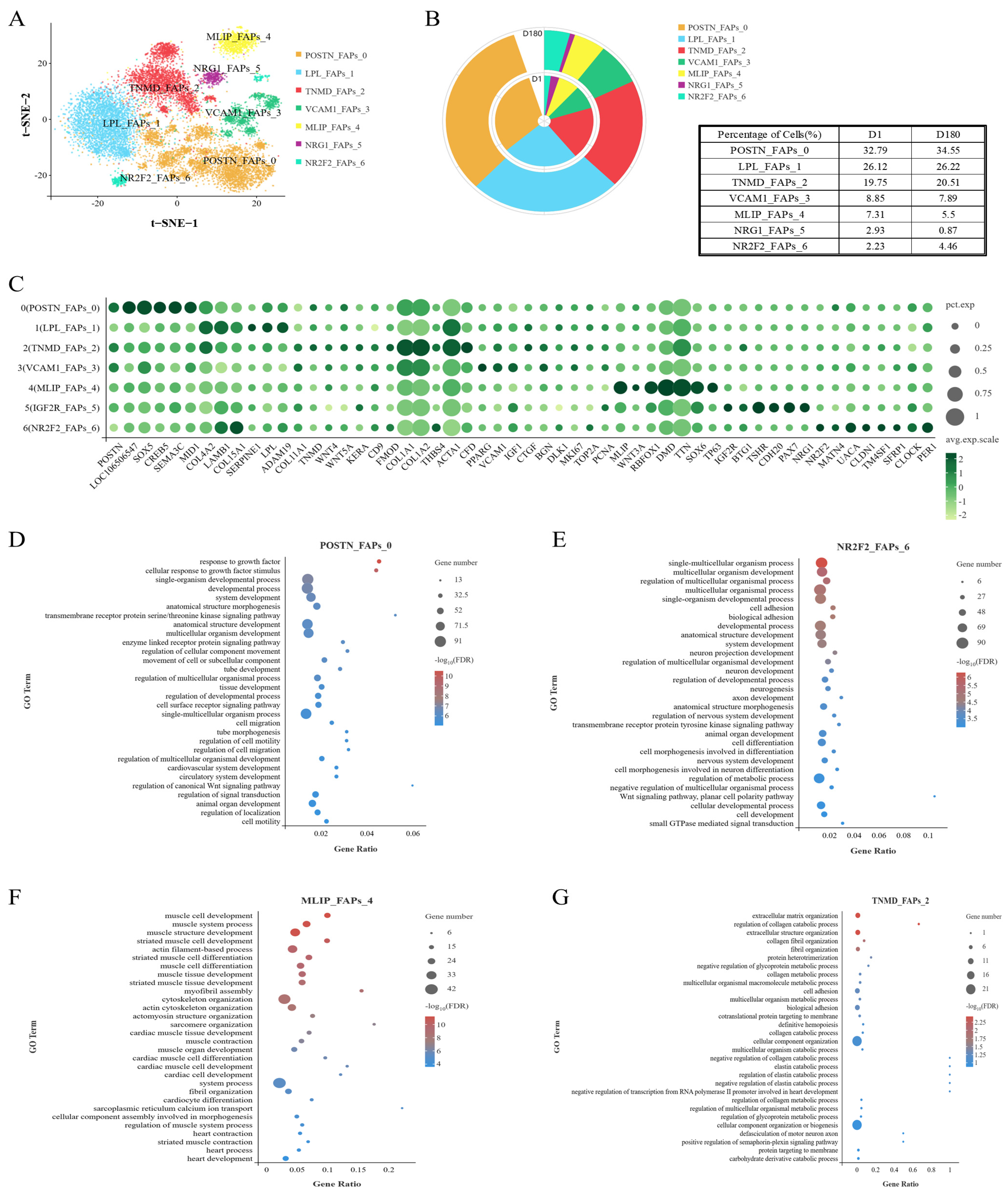
2.3. Analysis of the Heterogeneity of the FAP Subpopulation via Unsupervised Clustering
2.4. Genes Commonly Expressed During Two Stages of Adipocyte Development
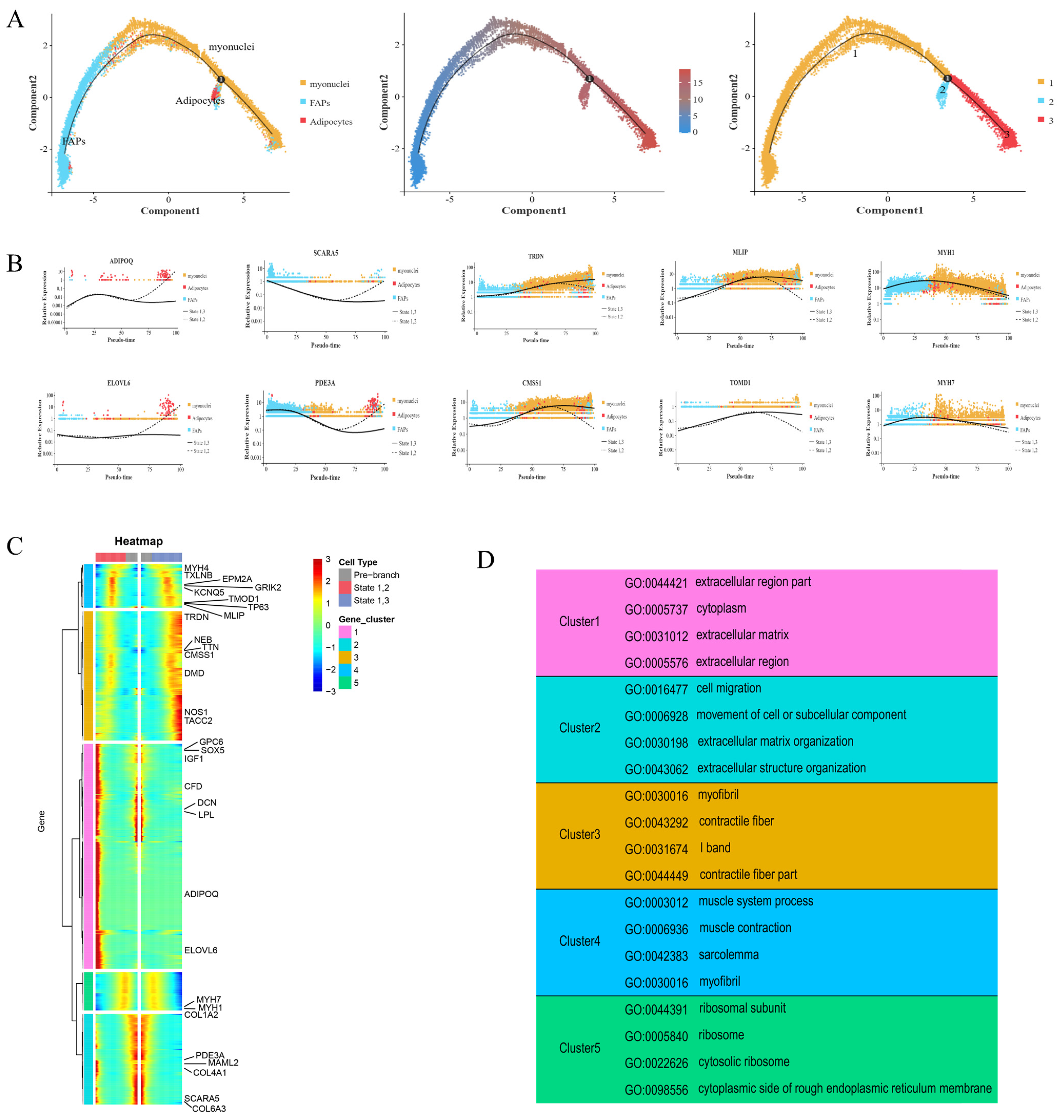
2.5. Pseudotime Analysis Revealed Key Progenitor Cell Characteristics and Gene Sets of FAPs
3. Discussion
4. Materials and Methods
4.1. Animals and Samples
4.2. Library Construction and Sequencing
4.3. SnRNA-Seq Bioinformatics Analysis
4.4. Single-Cell Trajectory Analysis
4.5. Pathway Enrichment Analysis
4.6. Cell Culture, Interference, and Induction of Differentiation
4.7. RT-qPCR and BODIPY, Oil-Red-O Staining
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Resare Sahlin, K.; Trewern, J. A Systematic Review of the Definitions and Interpretations in Scientific Literature of ‘Less but Better’ Meat in High-Income Settings. Nat. Food 2022, 3, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento-García, A.; Vieira-Aller, C. Improving Fatty Acid Profile in Native Breed Pigs Using Dietary Strategies: A Review. Animals 2023, 13, 1696. [Google Scholar] [CrossRef] [PubMed]
- Rajic, S.; Simunovic, S.; Djordjevic, V.; Raseta, M.; Tomasevic, I.; Djekic, I. Quality Multiverse of Beef and Pork Meat in a Single Score. Foods 2022, 11, 1154. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Huang, Y.; Das, A.K.; Yang, Q.; Duarte, M.S.; Dodson, M.V.; Zhu, M.-J. Meat Science and Muscle Biology Symposium: Manipulating Mesenchymal Progenitor Cell Differentiation to Optimize Performance and Carcass Value of Beef Cattle1,2. J. Anim. Sci. 2013, 91, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xing, S.; Zhao, G.; Zheng, M.; Yang, X.; Sun, J.; Wen, J.; Liu, R. Identification of Diverse Cell Populations in Skeletal Muscles and Biomarkers for Intramuscular Fat of Chicken by Single-Cell RNA Sequencing. BMC Genom. 2020, 21, 752. [Google Scholar] [CrossRef]
- Chen, W.; Tu, Y.; Cai, P.; Wang, L.; Zhou, Y.; Liu, S.; Huang, Y.; Zhang, S.; Gu, X.; Yi, W.; et al. Melatonin Supplementation Promotes Muscle Fiber Hypertrophy and Regulates Lipid Metabolism of Skeletal Muscle in Weaned Piglets. J. Anim. Sci. 2023, 101, skad256. [Google Scholar] [CrossRef]
- Yang, Y.; Yan, J.; Fan, X.; Chen, J.; Wang, Z.; Liu, X.; Yi, G.; Liu, Y.; Niu, Y.; Zhang, L.; et al. The Genome Variation and Developmental Transcriptome Maps Reveal Genetic Differentiation of Skeletal Muscle in Pigs. PLOS Genet. 2021, 17, e1009910. [Google Scholar] [CrossRef]
- Chen, F.-F.; Wang, Y.-Q.; Tang, G.-R.; Liu, S.-G.; Cai, R.; Gao, Y.; Sun, Y.-M.; Yang, G.-S.; Pang, W.-J. Differences between Porcine Longissimus Thoracis and Semitendinosus Intramuscular Fat Content and the Regulation of Their Preadipocytes during Adipogenic Differentiation. Meat Sci. 2019, 147, 116–126. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, X.; Liu, S.; You, W.; Huang, Y.; Zhou, Y.; Chen, W.; Zhang, S.; Wang, J.; Zheng, Q.; et al. Single-Nucleus and Bulk RNA Sequencing Reveal Cellular and Transcriptional Mechanisms Underlying Lipid Dynamics in High Marbled Pork. NPJ Sci. Food 2023, 7, 23. [Google Scholar] [CrossRef]
- Hwang, B.; Lee, J.H.; Bang, D. Single-Cell RNA Sequencing Technologies and Bioinformatics Pipelines. Exp. Mol. Med. 2018, 50, 96. [Google Scholar] [CrossRef]
- Qiu, K.; Xu, D.; Wang, L.; Zhang, X.; Jiao, N.; Gong, L.; Yin, J. Association Analysis of Single-Cell RNA Sequencing and Proteomics Reveals a Vital Role of Ca2+ Signaling in the Determination of Skeletal Muscle Development Potential. Cells 2020, 9, 1045. [Google Scholar] [CrossRef] [PubMed]
- Chemello, F.; Wang, Z.; Li, H.; McAnally, J.R.; Liu, N.; Bassel-Duby, R.; Olson, E.N. Degenerative and Regenerative Pathways Underlying Duchenne Muscular Dystrophy Revealed by Single-Nucleus RNA Sequencing. Proc. Natl. Acad. Sci. USA 2020, 117, 29691. [Google Scholar] [CrossRef]
- Xu, Z.; Wu, J.; Zhou, J.; Zhang, Y.; Qiao, M.; Sun, H.; Li, Z.; Li, L.; Chen, N.; Oyelami, F.O.; et al. Integration of ATAC-Seq and RNA-Seq Analysis Identifies Key Genes Affecting Intramuscular Fat Content in Pigs. Front. Nutr. 2022, 9, 1016956. [Google Scholar] [CrossRef]
- Ganassi, M.; Zammit, P.S. Involvement of Muscle Satellite Cell Dysfunction in Neuromuscular Disorders: Expanding the Portfolio of Satellite Cell-Opathies. Eur. J. Transl. Myol. 2022, 32, 10064. [Google Scholar] [CrossRef]
- Tong, S.; Sun, Y.; Kuang, B.; Wang, M.; Chen, Z.; Zhang, W.; Chen, J. A Comprehensive Review of Muscle–Tendon Junction: Structure, Function, Injury and Repair. Biomedicines 2024, 12, 423. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, D.; Liu, Y. Research Progress on the Regulating Factors of Muscle Fiber Heterogeneity in Livestock: A Review. Animals 2024, 14, 2225. [Google Scholar] [CrossRef]
- Li, Y.; Ma, J.; Yao, K.; Su, W.; Tan, B.; Wu, X.; Huang, X.; Li, T.; Yin, Y.; Tosini, G.; et al. Circadian Rhythms and Obesity: Timekeeping Governs Lipid Metabolism. J. Pineal Res. 2020, 69, e12682. [Google Scholar] [CrossRef]
- Allada, R.; Bass, J. Circadian Mechanisms in Medicine. N. Engl. J. Med. 2021, 384, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Wauman, J. Leptin Receptor Signaling: Pathways to Leptin Resistance. Front. Biosci. 2011, 16, 2771. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Wan, B.; Qiu, K.; Wang, Y.; Zhang, X.; Jiao, N.; Yan, E.; Wu, J.; Yu, R.; Gao, S.; et al. Single-Cell RNA-Sequencing Provides Insight into Skeletal Muscle Evolution during the Selection of Muscle Characteristics. Adv. Sci. 2023, 10, 2305080. [Google Scholar] [CrossRef]
- Hu, H.; Wang, J.; Zhu, R.; Guo, J.; Wu, Y. Effect of Myosin Heavy Chain Composition of Muscles on Meat Quality in Laiwu Pigs and Duroc. Sci. China Ser. C-Life Sci. 2008, 51, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Zhu, Q.; Duo, T.; Liang, Z.; Zhang, C.; Cai, S.; Wang, X.; Liu, Y.; Li, Y.; Liu, X.; et al. The Impact of FBN1-A5β1 Axis in Fibro/Adipogenic Progenitor Cells (FAPCD9−) on Intramuscular Fat Content in Pigs. J. Agr. Food Chem. 2024, 72, 9507–9521. [Google Scholar] [CrossRef] [PubMed]
- Ding, R.; Yang, M.; Quan, J.; Li, S.; Zhuang, Z.; Zhou, S.; Zheng, E.; Hong, L.; Li, Z.; Cai, G.; et al. Single-Locus and Multi-Locus Genome-Wide Association Studies for Intramuscular Fat in Duroc Pigs. Front. Genet. 2019, 10, 619. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Huang, M.; Tang, J.; Yang, L.; Yu, Z.; Li, D.; Li, G.; Jiang, Y.; Sun, Y.; et al. Whole-genome SNP Markers Reveal Conservation Status, Signatures of Selection, and Introgression in Chinese Laiwu Pigs. Evol. Appl. 2021, 14, 383–398. [Google Scholar] [CrossRef] [PubMed]
- Do, D.N.; Schenkel, F.S.; Miglior, F.; Zhao, X.; Ibeagha-Awemu, E.M. Genome Wide Association Study Identifies Novel Potential Candidate Genes for Bovine Milk Cholesterol Content. Sci. Rep. 2018, 8, 13239. [Google Scholar] [CrossRef]
- Löbbert, R.W.; Winterpacht, A.; Seipel, B.; Zabel, B.U. Zabel Molecular Cloning and Chromosomal Assignment of the Human Homologue of the Rat cGMP-Inhibited Phosphodiesterase 1 (PDE3A)—A Gene Involved in Fat Metabolism Located at 11p15.1. Genomics 1996, 37, 211–218. [Google Scholar] [CrossRef]
- Chopra, N.; Knollmann, B.C. Triadin Regulates Cardiac Muscle Couplon Structure and Microdomain Ca(2+) Signalling: A Path towards Ventricular Arrhythmias. Cardiovasc. Res. 2013, 98, 187–191. [Google Scholar] [CrossRef]
- Gong, P.; Jing, Y.; Liu, Y.; Wang, L.; Wu, C.; Du, Z.; Li, H. Whole-Genome Bisulfite Sequencing of Abdominal Adipose Reveals DNA Methylation Pattern Variations in Broiler Lines Divergently Selected for Fatness. J. Anim. Sci. 2021, 99, skaa408. [Google Scholar] [CrossRef] [PubMed]
- Schering, L.; Albrecht, E.; Komolka, K.; Kühn, C.; Maak, S. Increased Expression of Thyroid Hormone Responsive Protein (THRSP) Is the Result but Not the Cause of Higher Intramuscular Fat Content in Cattle. Int. J. Biol. Sci. 2017, 13, 532–544. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Estimated Number of Nuclei | Mean Reads Per Nuclei | Median Genes Per Nuclei | Total Genes Detected | Median UMI Counts Per Nuclei |
|---|---|---|---|---|---|
| D1_1 | 27,560 | 14,121 | 1150 | 22,556 | 2059 |
| D1_2 | 27,677 | 13,028 | 1033 | 22,446 | 1748 |
| D180_1 | 19,842 | 22,662 | 1194 | 22,547 | 2399 |
| D180_2 | 3223 | 128,122 | 1391 | 21,808 | 2910 |
| Groups | Nuclei Number after Filter | Median UMI Counts Per Nuclei After Filter | Median Genes Per Nuclei After Filter |
|---|---|---|---|
| D1 | 43,586 | 1716.5 | 994 |
| D180 | 19,685 | 2163 | 1115 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Z.; Wu, J.; Li, Y.; Zhou, J.; Zhang, Y.; Qiao, M.; Feng, Y.; Sun, H.; Li, Z.; Li, L.; et al. A Single-Cell Atlas of Porcine Skeletal Muscle Reveals Mechanisms That Regulate Intramuscular Adipogenesis. Int. J. Mol. Sci. 2024, 25, 12935. https://doi.org/10.3390/ijms252312935
Xu Z, Wu J, Li Y, Zhou J, Zhang Y, Qiao M, Feng Y, Sun H, Li Z, Li L, et al. A Single-Cell Atlas of Porcine Skeletal Muscle Reveals Mechanisms That Regulate Intramuscular Adipogenesis. International Journal of Molecular Sciences. 2024; 25(23):12935. https://doi.org/10.3390/ijms252312935
Chicago/Turabian StyleXu, Zhong, Junjing Wu, Yujie Li, Jiawei Zhou, Yu Zhang, Mu Qiao, Yue Feng, Hua Sun, Zipeng Li, Lianghua Li, and et al. 2024. "A Single-Cell Atlas of Porcine Skeletal Muscle Reveals Mechanisms That Regulate Intramuscular Adipogenesis" International Journal of Molecular Sciences 25, no. 23: 12935. https://doi.org/10.3390/ijms252312935
APA StyleXu, Z., Wu, J., Li, Y., Zhou, J., Zhang, Y., Qiao, M., Feng, Y., Sun, H., Li, Z., Li, L., Oyelami, F. O., Peng, X., & Mei, S. (2024). A Single-Cell Atlas of Porcine Skeletal Muscle Reveals Mechanisms That Regulate Intramuscular Adipogenesis. International Journal of Molecular Sciences, 25(23), 12935. https://doi.org/10.3390/ijms252312935