
Genome-Wide Identification of GRAS Gene Family in Cunninghamia lanceolata and Expression Pattern Analysis of ClDELLA Protein Under Abiotic Stresses


Abstract
1. Introduction
2. Results
2.1. Identification and Physicochemical Characterization of Chinese Fir GRAS Gene Family
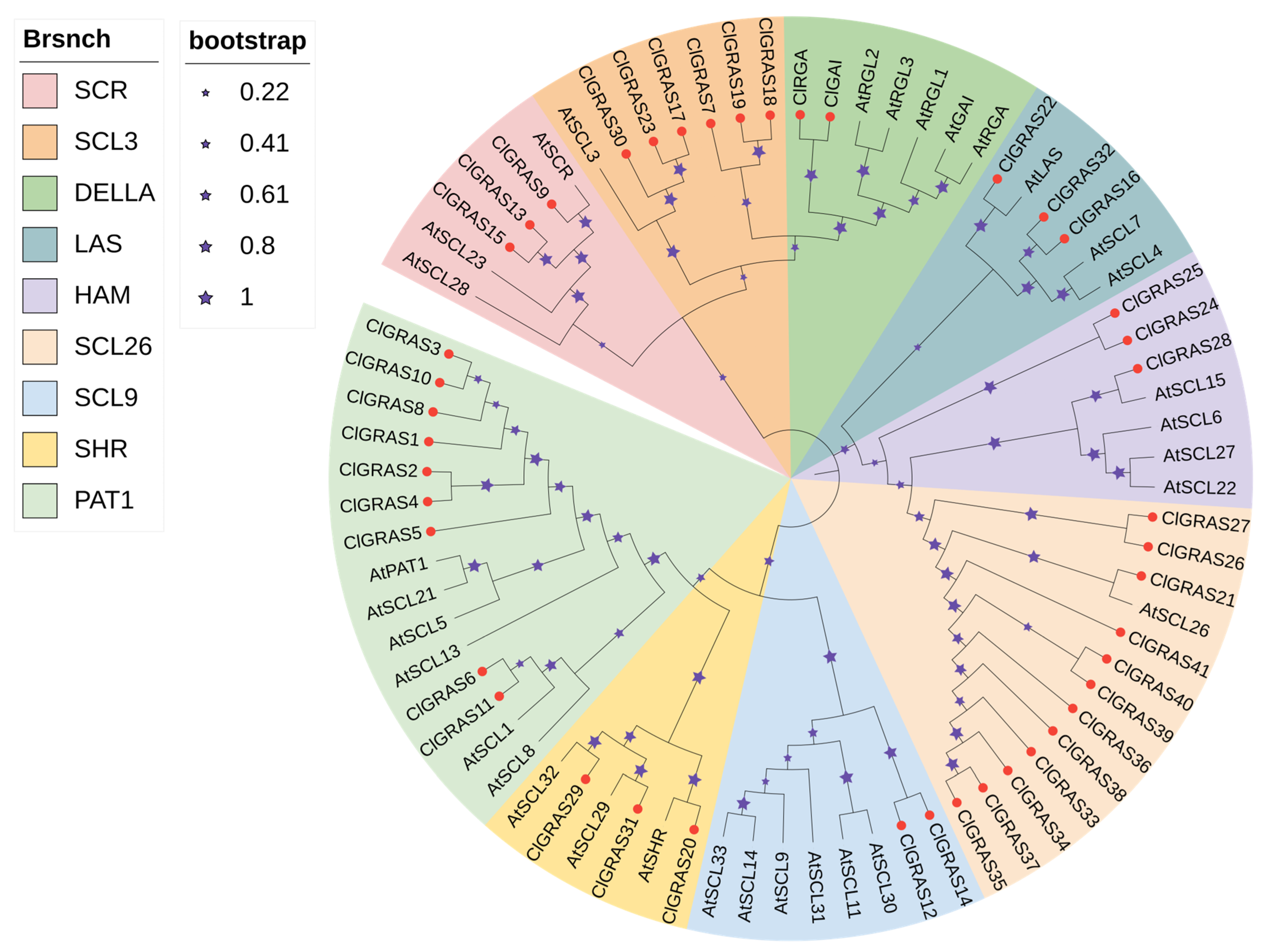
2.2. Phylogenetic Analysis of Chinese Fir GRAS Gene Family
2.3. Analysis of Gene Structure and Conserved Motifs in ClGRAS Family of Chinese Fir
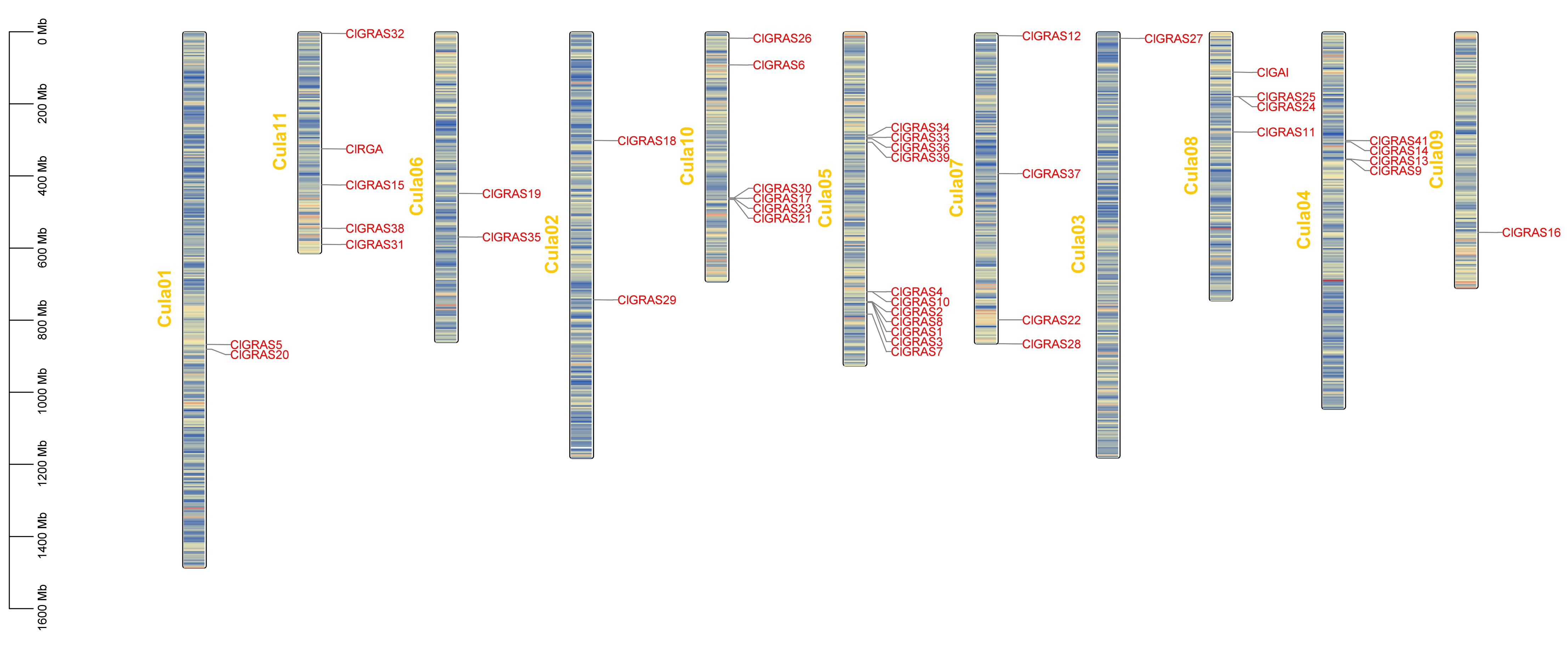
2.4. Analysis of Cis-Acting Elements and Chromosomal Localization in the ClGRAS Family of Chinese Fir
2.5. Collinearity Analysis in the ClGRAS Family of Chinese Fir
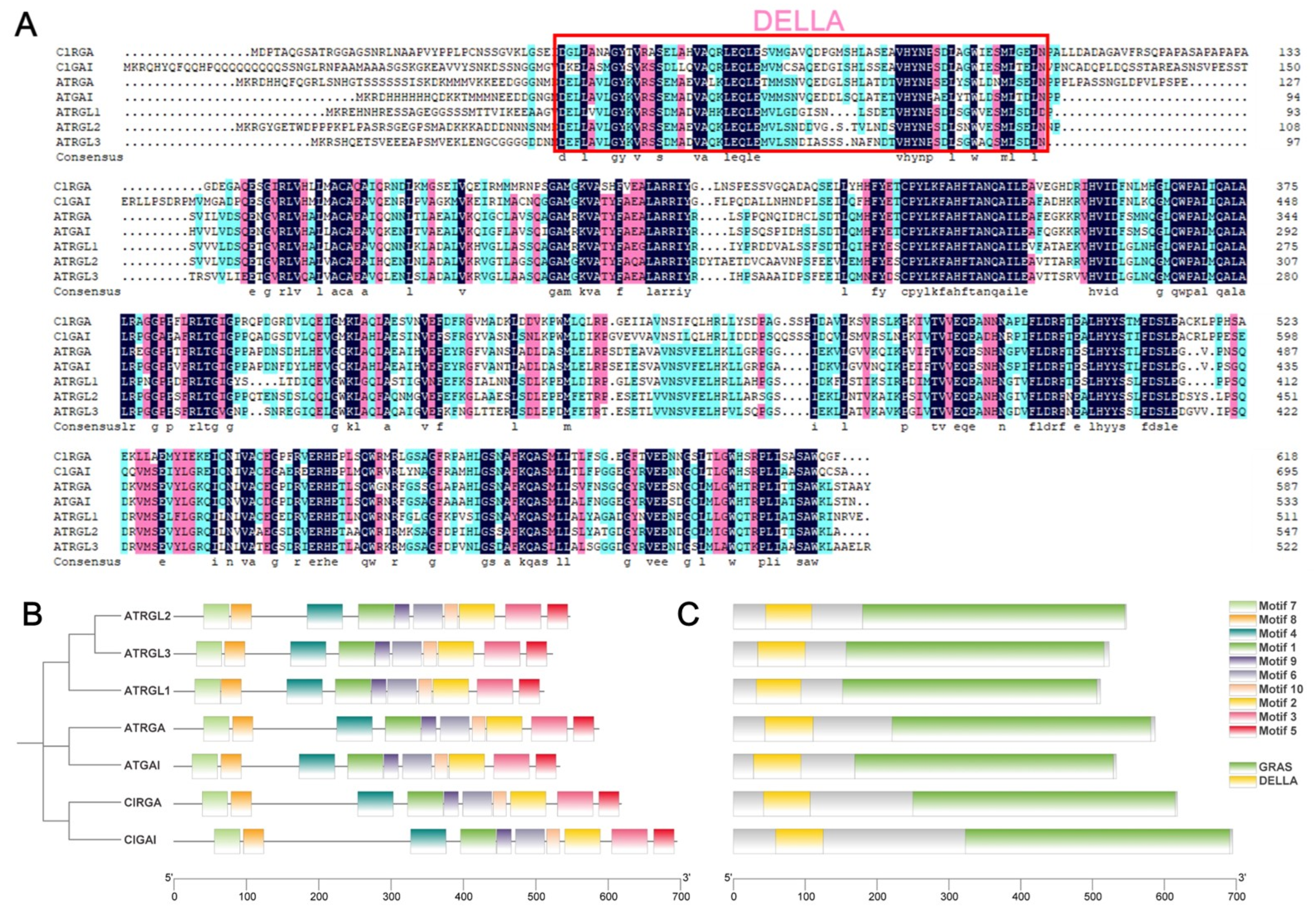
2.6. Sequence Comparison, Conserved Motifs, and Structural Domains Analysis of Chinese Fir DELLA Proteins
2.7. Subcellular Localization and Transcriptional Activation Activity of ClDELLA Protein
2.8. Expression Pattern of ClGAI and ClRGA in Different Tissues
2.9. Expression Pattern Analysis of ClDELLAs Under Different Hormonal and Abiotic Stresses
3. Discussion
4. Materials and Methods
4.1. Identification and Physicochemical Characterization of Chinese Fir GRAS Gene Family
4.2. Phylogenetic Analysis of the Chinese Fir GRAS Gene Family
4.3. Analysis of Gene Structure and Conserved Motifs in the ClGRAS Family of Chinese Fir
4.4. Analysis of Cis-Acting Elements and Chromosomal Localization in the ClGRAS Family of Chinese Fir
4.5. Expression Pattern Analysis of ClGAI and ClRGA
4.6. Subcellular Localization of the ClDELLA Protein
4.7. Transcriptional Activation Activity of the ClDELLA Protein
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hui, X.; Lin, Z.Z.; Su, S.D.; Jiang, X.L.; Chen, H.Q.; Wu, W.; Luo, S.J.; Pan, L.Y.; Zhen, R.H. Genetic variation analysis and ultra-early selection of nursery-determined traits in Chinese fir asexual lines. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2024, 48, 63–70. [Google Scholar]
- Lu, X.D.; Dong, Y.R.; Li, G.; Mao, L.F. Mechanisms of community construction in different stand development stages of subtropical Chinese fir plantation forests in China. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2024, 48, 67–73. [Google Scholar]
- Song, L.; Li, W.; Chen, X. Transcription factor is not just a transcription factor. Trends Plant. 2022, 27, 1087–1089. [Google Scholar] [CrossRef]
- Zhang, Z.; Cheng, J.; Wang, W.; Gao, Y.; Xian, X.; Li, C.; Wang, Y. Transcription factors dealing with Iron-deficiency stress in plants: Focus on the bHLH transcription factor family. Physiol. Plantarum. 2023, 175, e14091. [Google Scholar] [CrossRef]
- Bolle, C. The role of GRAS proteins in plant signal transduction and development. Planta 2004, 218, 683–692. [Google Scholar] [CrossRef]
- Zeng, X.; Ling, H.; Chen, X.; Guo, S. Genome-wide identification, phylogeny and function analysis of GRAS gene family in Dendrobium catenatum (Orchidaceae). Gene 2019, 705, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Cenci, A.; Rouard, M. Evolutionary Analyses of GRAS Transcription Factors in Angiosperms. Front. Plant Sci. 2017, 8, 273. [Google Scholar] [CrossRef]
- Lee, H.; Kim, B.; Song, S.; Heo, J.O.; Yu, N.L.; Lee, S.A.; Kim, M.; Sohn, S.O.; Lim, C.E.; Chang, K.; et al. Large-scale analysis of the GRAS gene family in Arabidopsis thaliana. Plant. Mol. Biol. 2008, 67, 659–670. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Tai, S.; Wang, D.W.; Ding, A.M.; Sun, T.; Wang, W.F.; Sun, Y.H. Homology-based analysis of the GRAS gene family in tobacco. Genet. Mol. Res. 2015, 14, 15188–15200. [Google Scholar] [CrossRef]
- Liu, X.Y.; Widmer, A. Genome-wide Comparative Analysis of the GRAS Gene Family in Populus, Arabidopsis and Rice. Plant Mol. Bio. Rep. 2014, 32, 1129–1145. [Google Scholar] [CrossRef]
- Tian, C.G.; Wan, P.; Sun, S.H.; Li, J.Y.; Chen, M.S. Genome-wide analysis of the GRAS gene family in rice and Arabidopsis. Plant Mol. Biol. 2004, 54, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Bhalothia, P. Evolutionary analysis of GRAS gene family for functional and structural insights into hexaploid bread wheat (Triticum aestivum). J. Biosci. 2021, 46, 15. [Google Scholar] [CrossRef]
- Zhu, L.; Yin, T.; Zhang, M.J.; Yang, X.Y.; Wu, J.X.; Cai, H.B.; Yang, N.; Li, X.L.; Wen, K.; Chen, D.M.; et al. Genome-wide identification and expression pattern analysis of the kiwifruit GRAS transcription factor family in response to salt stress. BMC Genom. 2024, 25, 12. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Xu, J.; Li, G.; Zhong, T.; Chen, D.; Lv, J. Genome-wide identification and expression analysis of GRAS gene family in Eucalyptus grandis. BMC Plant Biol. 2024, 24, 573. [Google Scholar] [CrossRef] [PubMed]
- Shan, Z.; Luo, X.; Wu, M.Y.; Wei, L.M.; Fan, Z.P.; Zhu, Y.M. Genome-wide identification and expression of GRAS gene family members in cassava. BMC Plant Biol. 2020, 20, 46. [Google Scholar] [CrossRef]
- Cai, H.; Xuan, L.; Xu, L.A.; Huang, M.R.; Xu, M. Identification and characterization of nine PAT1 branch genes in poplar. Plant Growth Regul. 2016, 81, 355–364. [Google Scholar] [CrossRef]
- Bisht, A.; Eekhout, T.; Canher, B.; Lu, R.; Vercauteren, I.; De Jaeger, G.; Heyman, J.; De Veylder, L. PAT1-type GRAS-domain proteins control regeneration by activating DOF3.4 to drive cell proliferation in Arabidopsis roots. Plant Cell 2023, 35, 1513–1531. [Google Scholar] [CrossRef]
- Wang, Z.; Wong, D.; Wang, Y.; Xu, G.Z.; Ren, C.; Liu, Y.F.; Kuang, Y.F.; Fan, P.G.; Li, S.H.; Xin, H.P.; et al. GRAS-domain transcription factor PAT1 regulates jasmonic acid biosynthesis in grape cold stress response. Plant Physiol. 2021, 186, 1660–1678. [Google Scholar] [CrossRef]
- Chen, C.; Lu, L.L.; Ma, S.Y.; Zhao, Y.P.; Wu, N.; Li, W.J.; Ma, L.; Kong, X.H.; Xie, Z.M.; Hou, Y.X. Analysis of PAT1 subfamily members in the GRAS family of upland cotton and functional characterization of GhSCL13-2A in Verticillium dahliae resistance. Plant Cell Rep. 2023, 42, 487–504. [Google Scholar] [CrossRef]
- Clark, N.M.; Hinde, E.; Winter, C.M.; Fisher, A.P.; Crosti, G.; Blilou, I.; Gratton, E.; Benfey, P.N.; Sozzani, R. Tracking transcription factor mobility and interaction in Arabidopsis roots with fluorescence correlation spectroscopy. eLife 2016, 5, 25. [Google Scholar] [CrossRef]
- Heo, J.O.; Chang, K.S.; Kim, I.A.; Lee, M.H.; Lee, S.A.; Song, S.K.; Lee, M.; Lim, J. Funneling of gibberellin signaling by the GRAS transcription regulator SCARECROW-LIKE 3 in the Arabidopsis root. Proc. Natl. Acad. Sci. USA 2011, 108, 2166–2171. [Google Scholar] [CrossRef]
- Blanco-Tourin, N.; Serrano-Mislata, A.; Alabad, D. Regulation of DELLA Proteins by Post-translational Modifications. Plant Cell Physiol. 2020, 61, 1891–1901. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.F.; Zhang, Q.Q.; Chen, Y.C.; Zhao, Y.X.; Ren, F.S.; Shi, H.M.; Wu, X.Y. Comprehensive identification and analysis of DELLA genes throughout the plant kingdom. BMC Plant Biol. 2020, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Hirano, K.; Sato, T.; Mistuda, N.; Nomoto, M.; Maeo, K.; Koketsu, E.; Mitani, R.; Kawamura, M.; Ishiguro, S.; et al. DELLA protein functions as a transcriptional activator through the DNA binding of the INDETERMINATE DOMAIN family proteins. Proc. Natl. Acad. Sci. USA 2014, 111, 7861–7866. [Google Scholar] [CrossRef] [PubMed]
- Murase, K.; Hirano, Y.; Sun, T.P.; Hakoshima, T. Gibberellin-induced DELLA recognition by the gibberellin receptor GID1. Nature 2008, 456, 459–463. [Google Scholar] [CrossRef]
- Huang, H.; Gong, Y.L.; Liu, B.; Wu, D.W.; Zhamg, M.; Xie, D.X.; Song, S.S. The DELLA proteins interact with MYB21 and MYB24 to regulate filament elongation in Arabidopsis. BMC Plant Biol. 2020, 20, 64. [Google Scholar] [CrossRef]
- Li, M.; An, F.Y.; Li, W.Y.; Ma, M.D.; Feng, Y.; Zhang, X.; Guo, H.W. DELLA proteins interact with FLC to repress flowering transition. J. Integr. Plant Biol. 2016, 58, 642–655. [Google Scholar] [CrossRef]
- Fukazawa, J.; Mori, K.; Ando, H.; Mori, R.; Kanno, Y.; Seo, M.; Takahashi, Y. Jasmonate inhibits plant growth and reduces gibberellin levels via microRNA5998 and transcription factor MYC2. Plant Physiol. 2023, 193, 2197–2214. [Google Scholar] [CrossRef]
- Hui, C. Protein Interactions Between DELLA, ARK2 and WOX4 Mediate the Molecular Mechanism of Gibberellin Regulation of Poplar Formative Layer Activity. Master’s Thesis, Southwest University, Chongqing, China, 2022. [Google Scholar]
- Tai, C.H.; Sam, V.; Gibrat, J.F.; Garnier, J.; Munson, P.; Lee, B. Protein domain assignment from the recurrence of locally similar structures. Proteins 2011, 79, 853–866. [Google Scholar] [CrossRef]
- Ben-Targem, M.; Ripper, D.; Bayer, M.; Ragni, L. Auxin and gibberellin signaling cross-talk promotes hypocotyl xylem expansion and cambium homeostasis. J. Exp. Bot. 2021, 72, 3647–3660. [Google Scholar] [CrossRef]
- Huang, M.X.; Zhang, G.F.; Gan, H.Y.; Liu, C.; Li, M.M.; Shu, Y.J. Genome-wide analysis of the GRAS gene family in white clover Trifolium repens L. provides insight into its critical role in response to cold stress. Biotechnol. Biotec. Eq. 2024, 38, 14. [Google Scholar] [CrossRef]
- He, Z.H.; Tian, Z.Z.; Zhang, Q.; Wang, Z.B.; Huang, R.K.; Xu, X.; Wang, Y.C.; Ji, X.Y. Genome-wide identification, expression and salt stress tolerance analysis of the GRAS transcription factor family in Betula platyphylla. Front. Plant Sci. 2022, 13, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yang, Y.B.; Yu, Y.H.; Mei, D.; Mao, X.; Fu, X.X. Genome-Wide Characterization of the GRAS Gene Family in Cyclocarya paliurus and Its Involvement in Heterodichogamy. Agronomy 2024, 14, 2397. [Google Scholar] [CrossRef]
- Yu, L.; Hui, C.; Huang, R.; Wang, D.; Fei, C.; Guo, C.; Zhang, J. Genome-wide identification, evolution and transcriptome analysis of GRAS gene family in Chinese chestnut (Castanea mollissima). Front. Genet. 2023, 13, 1080759. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Zhang, D.; Gao, C.; Zhao, M.; Wu, H.; Li, Y.; Shen, Y.; Han, M. Identification, classification, and expression analysis of GRAS gene family in Malus domestica. Front. Physiol. 2017, 8, 253. [Google Scholar] [CrossRef]
- Wang, L.; Ding, X.L.; Gao, Y.Q.; Yang, S.P. Genome-wide identification and characterization of GRAS genes in soybean (Glycine max). BMC Plant Biol. 2020, 20, 415. [Google Scholar] [CrossRef]
- Wang, S.; Duan, Z.; Yan, Q.; Wu, F.; Zhou, P.; Zhang, J.Y. Genome–Wide Identification of the GRAS Family Genes in Melilotus albus and Expression Analysis under Various Tissues and Abiotic Stresses. Int. J. Mol. Sci. 2022, 23, 7403. [Google Scholar] [CrossRef]
- Huang, Y.; Zheng, Q.Y.; Zhang, M.M.; He, X.; Zhao, X.W.; Wang, L.Y.; Lan, S.R.; Liu, Z.J. Genome-Wide Identification and Expression Analysis of the GRAS Gene Family and Their Responses to Heat Stress in Cymbidium goeringii. Int. J. Mol. Sci. 2024, 25, 6363. [Google Scholar] [CrossRef]
- Yuan, W.; Yu, J.; Li, Z.C. Rapid functional activation of horizontally transferred eukaryotic intron-containing genes in the bacterial recipient. Nucleic Acids Res. 2024, 52, 8344–8355. [Google Scholar] [CrossRef]
- Rushton, P.J. What Have We Learned About Synthetic Promoter Construction? Methods Mol. Biol. 2016, 1482, 1–13. [Google Scholar]
- Ma, M.; Li, L.; Wang, X.H.; Zhang, C.Y.; Pak, S.; Li, C.H. Comprehensive Analysis of GRAS Gene Family and Their Expression under GA3, Drought Stress and ABA Treatment in Larix kaempferi. Forests 2022, 13, 1424. [Google Scholar] [CrossRef]
- Li, M.; Zhou, Y.; Yang, Y. Inventory of gymnosperm species in China based on the latest gymnosperm taxonomic system. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2024, 48, 49–56. [Google Scholar]
- Aoyanagi, T.; Ilwya, S.; Kobayashi, A.; Kozaki, A. Gene Regulation via the Combination of Transcription Factors in the INDETERMINATE DOMAIN and GRAS Families. Genes 2020, 11, 18. [Google Scholar] [CrossRef]
- Briones-Moreno, A.; Hwrnandez-Garcia, J.; Vargas-Chavea, C.; Blanco-Tourinan, N.; Phokas, A.; Urbez, C.; Cerdan, P.D.; Coates, J.C.; Alabadi, D.; Blazquez, M.A. DELLA functions evolved by rewiring of associated transcriptional networks. Nat. Plants 2023, 9, 535–543. [Google Scholar] [CrossRef]
- Luo, W.R.; Zhao, Z.X.; Chen, H.Z.; Ao, W.H.; Lu, L.; Liu, J.J.; Li, X.Z.; Sun, Y.D. Genome-wide characterization and expression of DELLA genes in Cucurbita moschata reveal their potential roles under development and abiotic stress. Front. Plant Sci. 2023, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Wang, T.; Yang, K.; Li, L.B. Characterization of cireRNAs and its expression in germinating seeds of Phyllostachys edulis under PEG and NaCl stress. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2023, 47, 17–24. [Google Scholar]
- Zhou, H.; Wang, Y.W.; Wang, X.Y.; Cheng, R.; Zhang, H.X.; Yang, L. Genome-wide characterization of DELLA gene family in blueberry (Vaccinium darrowii) and their expression profiles in development and response to abiotic stress. BMC Genom. 2024, 25, 815. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.X.; Cao, Y.B.; Fan, Y.J.; Fan, G.Q. Comprehensive Analysis of the GRAS Gene Family in Paulownia fortunei and the Response of DELLA Proteins to Paulownia Witches’ Broom. Int. J. Mol. Sci. 2024, 25, 15. [Google Scholar] [CrossRef]
- Mäkilä, R.; Wybouw, B.; Smetana, O.; Vainio, L.; Sole-Gil, A.; Lyu, M.; Ye, L.L.; Wang, X.; Siligato, R.; Jeness, M.K.; et al. Gibberellins promote polar auxin transport to regulate stem cell fate decisions in cambium. Nat. Plants 2023, 9, 631–644. [Google Scholar] [CrossRef]
- Wang, D.B.; Qiu, Z.M.; Xu, T.; Yao, S.; Zhang, M.Y.; Cheng, X.; Zhao, Y.L.; Ji, K.S. Identification and Expression Patterns of WOX Transcription Factors under Abiotic Stresses in Pinus massoniana. Int. J. Mol. Sci. 2024, 25, 1627. [Google Scholar] [CrossRef]
- Zhu, L.J.; Yang, J.J.; Zhang, Y.T.; Hu, H.L.; Cui, J.B.; Xue, J.Y.; Xu, J. Overexpression of CfICE1 from Cryptomeria fortunei Enhances Cold, Drought and Salt Stress in Poplar. Int. J. Mol. Sci. 2022, 23, 15214. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.T.; Xue, J.Y.; Zhu, L.J.; Hu, H.L.; Yang, J.J.; Cui, J.B.; Xu, J. Selection and Optimization of Reference Genes for MicroRNA Expression Normalization by qRT-PCR in Chinese Cedar (Cryptomeria fortunei) under Multiple Stresses. Int. J. Mol. Sci. 2021, 22, 7246. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Chromosome Localization | Prediction of Subcellular Localization | Length (aa) | Molecular Weight (kDa) | pI | Aliphatic Index | Instability Index | GRAVY |
|---|---|---|---|---|---|---|---|---|
| ClGRAS1 | Cula05 | Nucleus | 1316 | 146.94 | 5.19 | 74.98 | 55.63 | −0.432 |
| ClGRAS2 | Cula05 | Nucleus | 692 | 77.33 | 4.98 | 71.69 | 56.07 | −0.510 |
| ClGRAS3 | Cula05 | Nucleus | 646 | 72.55 | 5.15 | 74.85 | 52.66 | −0.443 |
| ClGRAS4 | Cula05 | Nucleus | 692 | 77.36 | 4.98 | 71.69 | 55.92 | −0.495 |
| ClGRAS5 | Cula01 | Nucleus | 548 | 61.25 | 4.87 | 83.67 | 39.28 | −0.328 |
| ClGRAS6 | Cula10 | Nucleus | 693 | 77.13 | 5.02 | 83.16 | 54.46 | −0.336 |
| ClGRAS7 | Cula05 | Nucleus | 652 | 71.70 | 5.84 | 79.19 | 56.74 | −0.327 |
| ClGAI | Cula08 | Nucleus | 695 | 77.30 | 5.29 | 78.62 | 60.04 | −0.480 |
| ClRGA | Cula11 | Nucleus | 618 | 66.72 | 5.55 | 85.66 | 50.30 | −0.103 |
| ClGRAS8 | Cula05 | Nucleus | 688 | 76.81 | 4.89 | 74.69 | 56.44 | −0.463 |
| ClGRAS9 | Cula04 | Nucleus | 866 | 94.44 | 5.79 | 76.09 | 48.58 | −0.476 |
| ClGRAS10 | Cula05 | Nucleus | 639 | 72.06 | 5.57 | 74.90 | 53.17 | −0.470 |
| ClGRAS11 | Cula08 | Nucleus | 707 | 78.21 | 6.13 | 86.35 | 48.55 | −0.304 |
| ClGRAS12 | Cula07 | Nucleus | 826 | 92.98 | 5.76 | 75.04 | 49.17 | −0.535 |
| ClGRAS13 | Cula04 | Nucleus | 691 | 75.96 | 5.36 | 74.43 | 48.26 | −0.364 |
| ClGRAS14 | Cula04 | Nucleus | 549 | 62.21 | 6.06 | 84.70 | 45.36 | −0.335 |
| ClGRAS15 | Cula11 | Nucleus | 609 | 67.17 | 5.69 | 76.77 | 56.08 | −0.263 |
| ClGRAS16 | Cula09 | Nucleus | 746 | 82.27 | 5.45 | 72.12 | 55.39 | −0.353 |
| ClGRAS17 | Cula10 | Nucleus | 491 | 54.87 | 6.33 | 86.05 | 60.54 | −0.245 |
| ClGRAS18 | Cula02 | Nucleus | 599 | 67.13 | 6.45 | 80.18 | 59.07 | −0.393 |
| ClGRAS19 | Cula06 | Nucleus | 552 | 61.35 | 5.31 | 82.07 | 45.73 | −0.298 |
| ClGRAS20 | Cula01 | Nucleus | 485 | 55.56 | 5.58 | 70.43 | 39.75 | −0.353 |
| ClGRAS21 | Cula10 | Nucleus | 605 | 67.13 | 5.69 | 79.85 | 49.92 | −0.266 |
| ClGRAS22 | Cula07 | Nucleus | 433 | 48.88 | 5.41 | 78.48 | 56.54 | −0.470 |
| ClGRAS23 | Cula10 | Nucleus | 478 | 53.39 | 6.33 | 86.34 | 60.06 | −0.267 |
| ClGRAS24 | Cula08 | Nucleus | 509 | 57.83 | 8.39 | 88.53 | 48.61 | −0.302 |
| ClGRAS25 | Cula08 | Nucleus | 492 | 55.79 | 8.88 | 89.21 | 62.62 | −0.375 |
| ClGRAS26 | Cula10 | Nucleus | 580 | 65.18 | 5.29 | 87.95 | 62.29 | −0.206 |
| ClGRAS27 | Cula03 | Nucleus | 601 | 67.67 | 5.61 | 90.55 | 62.04 | −0.169 |
| ClGRAS28 | Cula07 | Nucleus | 704 | 79.47 | 5.22 | 81.99 | 54.76 | −0.314 |
| ClGRAS29 | Cula02 | Nucleus | 390 | 43.72 | 5.77 | 85.05 | 54.84 | −0.196 |
| ClGRAS30 | Cula10 | Nucleus | 498 | 56.01 | 5.68 | 86.08 | 61.90 | −0.251 |
| ClGRAS31 | Cula11 | Nucleus | 590 | 65.03 | 5.42 | 77.10 | 35.08 | −0.455 |
| ClGRAS32 | Cula11 | Nucleus | 468 | 52.05 | 4.91 | 83.55 | 55.18 | −0.163 |
| ClGRAS33 | Cula05 | Nucleus | 570 | 64.49 | 4.62 | 84.68 | 46.32 | −0.312 |
| ClGRAS34 | Cula05 | Nucleus | 497 | 55.60 | 4.76 | 86.36 | 45.98 | −0.239 |
| ClGRAS35 | Cula06 | Nucleus | 551 | 62.12 | 4.46 | 86.93 | 47.96 | −0.244 |
| ClGRAS36 | Cula05 | Nucleus | 476 | 53.72 | 4.53 | 86.85 | 46.55 | −0.157 |
| ClGRAS37 | Cula07 | Nucleus | 491 | 54.95 | 4.70 | 90.81 | 38.73 | −0.192 |
| ClGRAS38 | Cula11 | Nucleus | 534 | 59.21 | 4.75 | 89.87 | 48.72 | −0.213 |
| ClGRAS39 | Cula05 | Nucleus | 559 | 62.57 | 4.61 | 84.96 | 36.75 | −0.177 |
| ClGRAS40 | contig09198 | Nucleus | 488 | 55.25 | 4.73 | 93.93 | 57.21 | −0.039 |
| ClGRAS41 | Cula04 | Nucleus | 487 | 55.12 | 5.16 | 82.20 | 50.01 | −0.179 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Y.; Jin, M.; Yang, J.; Yang, Y.; Guo, R.; Luo, H.; Guo, T.; Xu, J. Genome-Wide Identification of GRAS Gene Family in Cunninghamia lanceolata and Expression Pattern Analysis of ClDELLA Protein Under Abiotic Stresses. Int. J. Mol. Sci. 2024, 25, 12262. https://doi.org/10.3390/ijms252212262
Luo Y, Jin M, Yang J, Yang Y, Guo R, Luo H, Guo T, Xu J. Genome-Wide Identification of GRAS Gene Family in Cunninghamia lanceolata and Expression Pattern Analysis of ClDELLA Protein Under Abiotic Stresses. International Journal of Molecular Sciences. 2024; 25(22):12262. https://doi.org/10.3390/ijms252212262
Chicago/Turabian StyleLuo, Yi, Mengshuang Jin, Junjie Yang, Ye Yang, Runxin Guo, Huan Luo, Tianhao Guo, and Jin Xu. 2024. "Genome-Wide Identification of GRAS Gene Family in Cunninghamia lanceolata and Expression Pattern Analysis of ClDELLA Protein Under Abiotic Stresses" International Journal of Molecular Sciences 25, no. 22: 12262. https://doi.org/10.3390/ijms252212262
APA StyleLuo, Y., Jin, M., Yang, J., Yang, Y., Guo, R., Luo, H., Guo, T., & Xu, J. (2024). Genome-Wide Identification of GRAS Gene Family in Cunninghamia lanceolata and Expression Pattern Analysis of ClDELLA Protein Under Abiotic Stresses. International Journal of Molecular Sciences, 25(22), 12262. https://doi.org/10.3390/ijms252212262