Abstract
Breast cancer stands out as the most widespread form of cancer globally. In this study, the anticancer activities of Clerodendrum chinense (C. chinense) stem ethanolic extract were investigated. High-performance liquid chromatography (HPLC) analysis identified verbascoside and isoverbascoside as the major bioactive compounds in the C. chinense stem extract. Successfully developed nanoparticles exhibited favorable hydrodynamic diameter, polydispersity index, and surface charge, thus ensuring stability after four months of storage. The total phenolic content and total flavonoid contents in the nanoparticles were reported as 88.62% and 95.26%, respectively. The C. chinense stem extract demonstrated a dose-dependent inhibitory effect on MCF-7, HeLa, A549, and SKOV-3 cancer cell lines, with IC50 values of 109.2, 155.6, 206.9, and 423 µg/mL, respectively. C. chinense extract and NPs exhibited dose-dependent cytotoxicity and the highest selectivity index values against MCF-7 cells. A dose-dependent reduction in the colony formation of MCF-7 cells was observed following treatment with the extract and nanoparticles. The extract induced cytotoxicity in MCF-7 cells through apoptosis and necrosis. C. chinense stem extract and nanoparticles decreased mitochondrial membrane potential (MMP) and induced G0/G1 phase arrest in MCF-7 cells. In conclusion, use of C. chinense stem extract and nanoparticles may serve as a potential therapeutic approach for breast cancer, thus warranting further exploration.
1. Introduction
Cancer is a significant global health problem that affects millions of people worldwide, contributing to substantial morbidity, mortality, and economic burden. It is characterized by the uncontrolled growth and spread of abnormal cells in the body, leading to the formation of tumors and interference with normal bodily functions [1]. Breast cancer is the most common cancer in the world, and it continues to have a significant impact on the total number of cancer deaths. Global efforts are required to mitigate its growing burden, particularly in countries where its incidence is rapidly increasing, and mortality rates remain high. In 2020, there were over 2.3 million new cases of breast cancer and 685,000 deaths [2]. Despite increases in incidence over the last several decades, breast cancer mortality has decreased due to advancements in screening and targeted therapy [3]. Breast cancer is a genetic disease caused by mutations in neoplastic cells [4]. Mutations in tumor suppressor and oncogenic genes cause breast epithelial cells to develop a malignant phenotype [5]. These genetic changes influence breast cancer behavior, including responses to therapy and clinical outcomes.
The discovery of anticancer drugs from plants has played a pivotal role in the development of cancer treatments. Many successful anticancer drugs have originated from natural compounds found in plants. For example, vinca alkaloids, such as vinblastine and vincristine, derived from Catharanthus roseus have been used in the treatment of various cancers, including Hodgkin’s disease, choriocarcinoma, neuroblastoma, lymphosarcoma, and carcinomas of the breast and lungs [6]. While conventional anticancer drugs have revolutionized cancer treatment and improved patient outcomes, the rapid evolution of cancer cells and the development of resistance leads to ongoing challenges. As a result, the exploration of natural products, such as those found in plants, remains a promising approach for anticancer drug discovery.
Natural active compounds have garnered the interest of researchers due to their ability to deliver anticancer drugs with minimal toxicity [7]. Numerous herbs and their bioactive compounds have played an important role in the treatment of various types of cancer. Recent in vitro, in vivo, and clinical studies have revealed that a variety of bioactive compounds have the potential to be chemopreventive or chemotherapeutic agents against breast cancer. Various properties have been attributed to these natural bioactive compounds, including antiproliferative properties, induction of apoptosis, anti-inflammatory properties, antiangiogenic properties, anti-invasive and metastatic properties, cell cycle regulatory activity, tumor suppressor activities, and targeting of cancer stem cells against breast cancer [8].
Polyphenols exhibit cytotoxic effects on a variety of cancer cells [9]. Polyphenols exert their anticancer effects through diverse mechanisms, including the modification of signaling pathways, inhibition of cell cycle events, and induction of apoptosis, leading to the removal of cancer cells [10]. In addition, these compounds regulate the activities of enzymes pivotal in tumor cell proliferation [11]. Recent studies are increasingly implicating natural polyphenols in anticancer activities through a myriad of properties, including, but not limited to, antiangiogenic effects, antimetastatic properties, DNA interaction, and various other pathways [12,13,14].
Clerodendrum is a significant plant genus used as a traditional medicine to treat a variety of disorders. Several in vitro and in vivo tests have been conducted to validate these traditional claims. Various species of Clerodendrum genus have potent antimicrobial, anti-inflammatory, anti-malarial, anti-diabetic, anticancer, analgesic, and antioxidant properties [15]. Qi reported that the diterpene compound isolated from C. chinense root extract demonstrated anticancer activities against HL-60 and A-549 cell lines [16]. Barung et al. showed that ethanol extract, hexane fraction, ethyl acetate fraction, and water-soluble fraction of C. chinense had anticancer activity on A549 lung cancer cells [17]. The ethyl acetate fraction had the lowest IC50 value and was classified as moderately active against lung cancer. Previously, we reported the anticancer activities of C. chinense leaf and flower extracts against MCF-7, A549, and HeLa cells [18,19]. The leaf and flower extract of C. chinense have undergone extensive research in various cancer cell lines; however, there remains a gap in the existing knowledge, as the anticancer activities of the stem extract in relation to a breast cancer cell line have never been investigated.
Nanotechnology has emerged as an attractive research topic for drug delivery through the use of the designed nanomaterials in a variety of applications, such as pharmaceuticals and biomedicine. Nanoparticulate delivery systems aim to enhance the delivery of drugs or natural active compounds with poor water solubility. These features include facilitating site-specific targeting to minimize drug accumulation in healthy tissue, prolonging the presence of the drug in the body for effective treatment, extending drug bioactivity through protection from the biological environment, and enabling the transport of drugs across epithelial and endothelial barriers.
Recent advances in nanomaterials have paved the way for the development of novel cancer therapeutics. Due to their nanometer size, nanoparticles are considered an excellent option for passive and active targeted anticancer drug delivery [20]. In this investigation, we have used C. chinense stem extract for the first time to develop nanoparticles and determined its application for anticancer activity. C. chinense stem extract and NPs are expected to selectively target and affect cancer cells while preserving normal cells. The selectivity of these anticancer agents plays a pivotal role in mitigating the side effects associated with cancer treatment and has the potential to enhance therapeutic efficacy by concentrating on cancer cells. We hypothesized that C. chinense stem extract may exert anticancer activities through specific mechanisms and the nanoparticles of the extract can be used as a drug delivery system while preserving the anticancer activity of the extract.
2. Results
2.1. Yield of C. chinense Stem Extract
The extraction of C. chinense stem using 95% ethanol yielded 3.33 ± 0.58% w/w. The percentage yield indicated that approximately 3.33% of the soluble compounds in the C. chinense stem were successfully extracted using the maceration method. Several factors may have influenced the yield, including the temperature of extraction, extraction time, extraction solvent polarity, and the source of C. chinense stem [21]. The yield of C. chinense stem extraction was lower compared with C. chinense leaf (11.4%) and flower (11.2%) [18,19].
2.2. High-Performance Liquid Chromatography (HPLC) Profiles of Phytochemicals in C. chinense Stem Extract
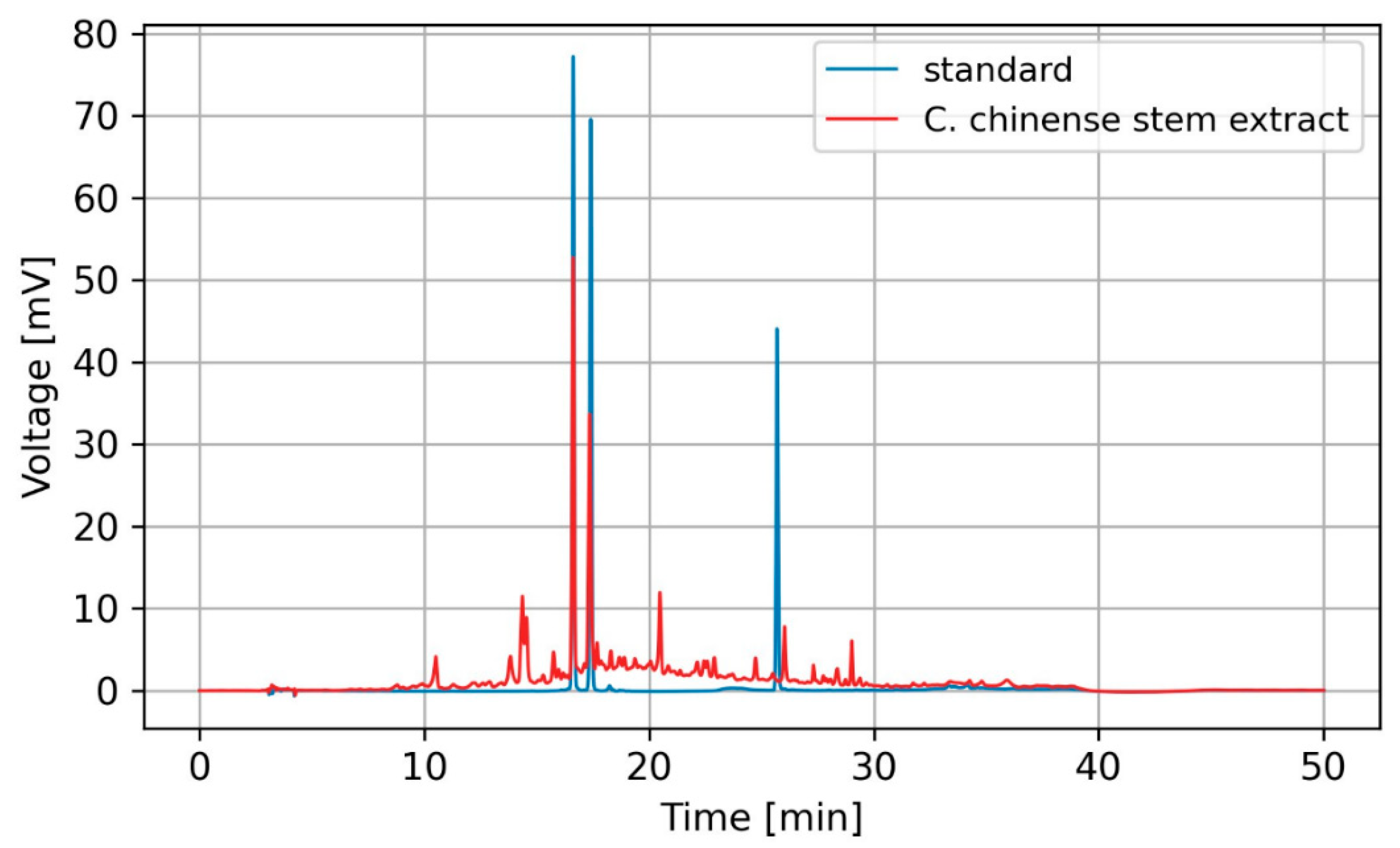
The HPLC analysis results revealed the presence of verbascoside and isoverbascoside as major bioactive compounds in C. chinense stem extract. Figure 1 shows representative HPLC chromatograms generated for the detection of two compounds. The results showed that C. chinense stem extract contained verbascoside and isoverbascoside. Hispidulin was not observed in this extract. Verbascoside was more abundant than isoverbascoside. Stem extract of C. chinense contained 9.42 ± 0.15 µg verbascoside per mg of extract and 8.32 ± 0.15 µg isoverbascoside per mg of extract. In our earlier investigations, we identified verbascoside, isoverbascoside, and hispidulin in the extracts from the leaves and flowers of C. chinense. Verbascoside exhibited higher abundance in the flower extract, whereas isoverbascoside was identified as a predominant compound in the leaf extract. The amount of verbascoside detected in the stem of C. chinense was lower compared with the flower (11.27 ± 0.08 µg/mg extract) [18]. The amounts of other compounds in the extract were too low to be identified or quantified through HPLC analysis and were neglected.

Figure 1.
HPLC chromatogram of verbascoside, isoverbascoside, and hispidulin standards (blue) and C. chinense stem extract (red).
2.3. Characterization and Physical Stability Study of C. chinense Stem Extract Nanoparticles (NPs): Particle Size, Polydispersity Index (PDI), and Zeta Potential Value
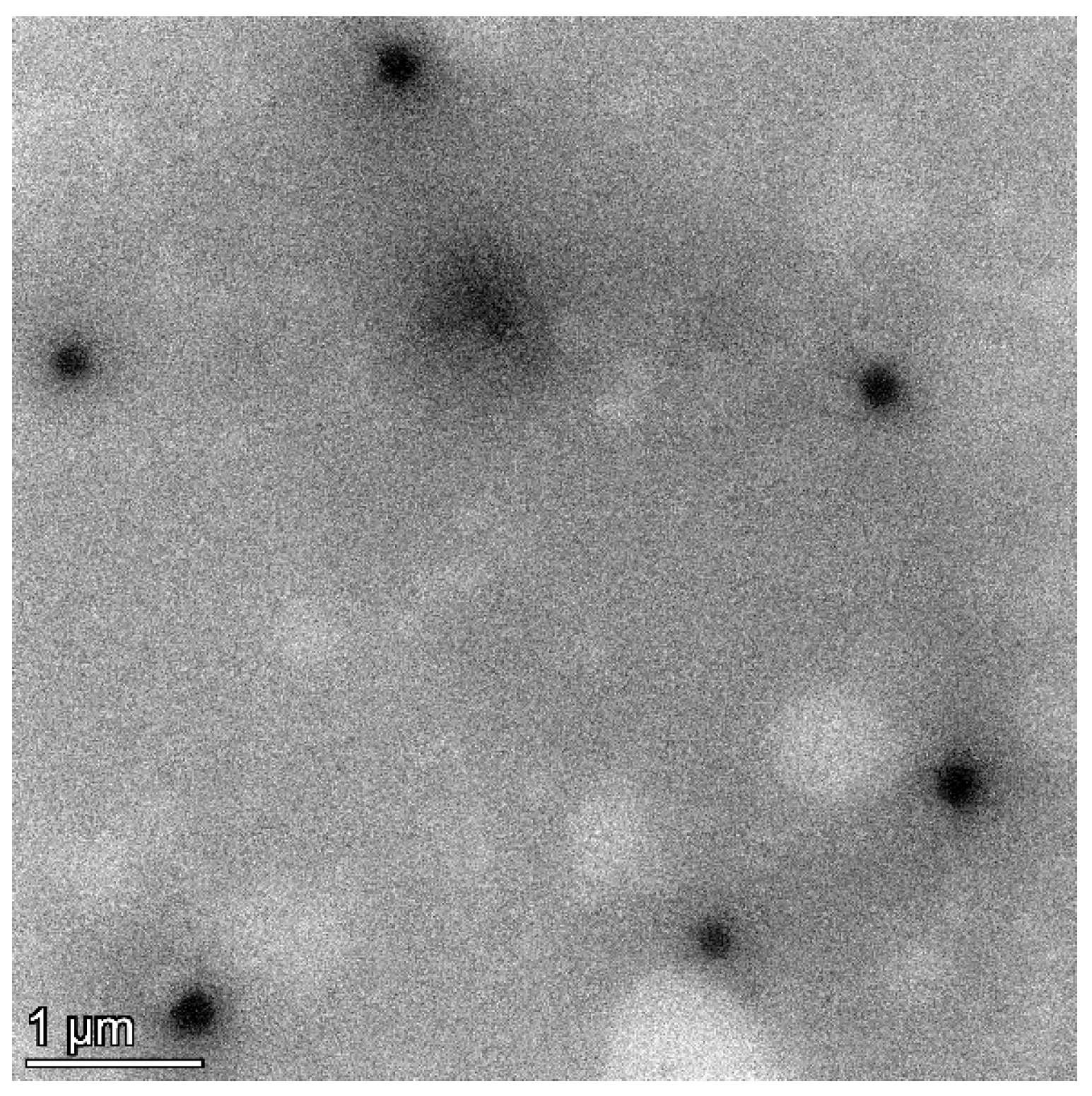
To determine the particle size, polydispersity index (PDI), and zeta potential, the C. chinense stem extract NPs were diluted tenfold with deionized water and subjected to analysis using a dynamic light-scattering-based particle size analyzer (Zetasizer Nano ZS, Malvern Instruments, Malvern, UK). Figure 2 and Supplementary Figure S1 revealed that the hydrodynamic diameter of C. chinense stem extract NPs measured 164.73 ± 2.37 nm. This size aligns closely with the findings from transmission electron microscopy (TEM) analysis, thereby confirming the accuracy of the results. The C. chinense stem extract NPs appear as spherical structures with a uniform size distribution (Figure 3). The NPs exhibited a relatively narrow size distribution, as evidenced by the low PDI value of 0.098 ± 0.008. The zeta potential of C. chinense stem extract NPs was determined to be −29.17 ± 0.32 mV (Supplementary Figure S2). These findings collectively demonstrate the successful application of the solvent displacement method in the preparation of C. chinense stem extract NPs.
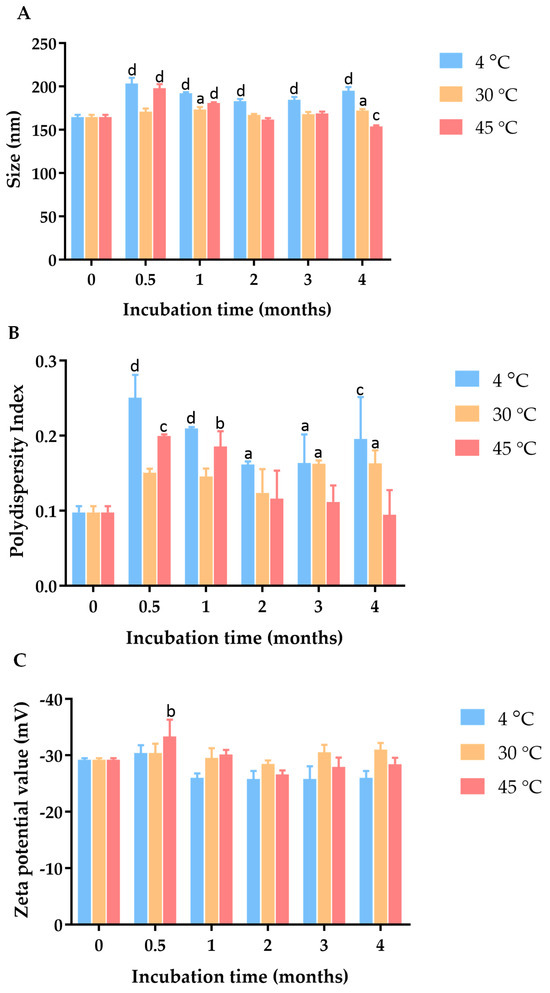
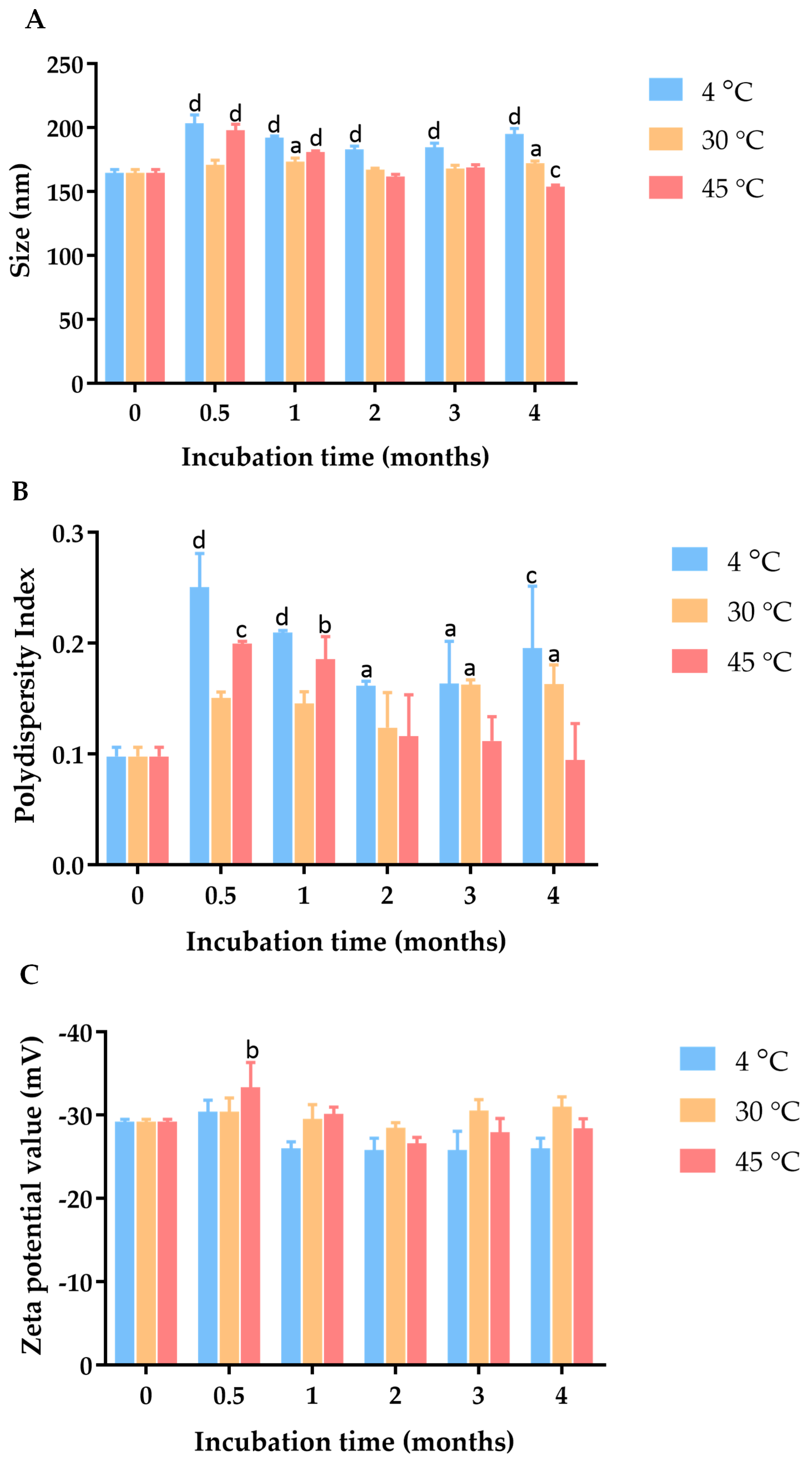
Figure 2.
Effect of incubation time and temperature on (A) particle size, (B) PDI, and (C) zeta potential of C. chinense stem extract nanoparticles. Data are presented as the mean ± SD. The letters a, b, c, and d indicate p < 0.05, 0.01, 0.001, and 0.001, respectively. Tukey’s multiple comparisons test for size, polydispersity index, and zeta potential statistical analysis are shown in Supplementary Tables S1–S3.
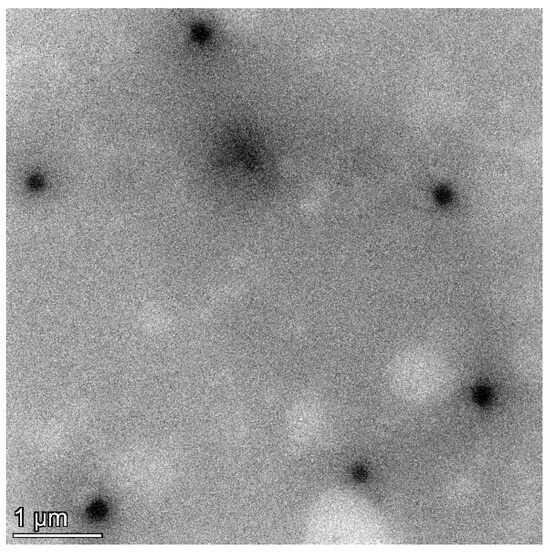
Figure 3.
TEM image of C. chinense stem extract nanoparticles.
To investigate the physical stability of these C. chinense stem extract NPs, samples were stored in a sealed container at three different temperature conditions, including 4 °C, 30 °C, and 45 °C, for a duration of 4 months. At specific intervals, particle size, PDI, and zeta potential measurements were conducted to monitor the influence of time and temperature on the NPs’ physical properties. The hydrodynamic diameter and polydispersity index of the C. chinense extract NPs were assessed at various time points during the stability study (Supplementary Figure S3). After four weeks of storage at 4 °C, 30 °C, and 45 °C, the maximum average particle sizes of NPs were 203.47 ± 6.39 nm, 173.30 ± 2.77 nm, and 197.80 ± 4.87 nm, respectively. The PDI values of NPs were 0.25 ± 0.03, 0.16 ± 0.02, and 0.20 ± 0.00 when stored at 4 °C, 30 °C, and 45 °C, respectively. These results suggested that there was minimal and acceptable change in particle size and size distribution during this period. Zeta potential measurements were conducted to assess the surface charge of the nanoparticles, which can influence their stability. The results showed that the zeta potential ranged between −29.2 mV and −33.33 mV at 4 °C, 30 °C, and 45 °C, suggesting that the surface charge was relatively maintained, thus contributing to the stability of the nanoparticles.
2.4. Total Phenolic Content and Total Flavonoid Content in C. chinense Stem Extract and NPs
The total phenolic contents in C. chinense stem extract and NPs were found to be 553.20 ± 68.74 and 490.26 ± 24.12 milligrams of gallic acid equivalent (GAE) per gram of extract, respectively. The total flavonoid compounds in C. chinense stem extract and NPs were expressed in 185.44 ± 37.39 and 176.65 ± 13.07 milligrams of quercetin equivalent (QE) per gram of extract. These results revealed that the total phenolic content and total flavonoid contents in the C. chinense stem extract NPs were 88.62% and 95.26%, respectively. The strong correlations between NP concentrations and TPC and TFC are shown in Supplementary Figures S4 and S5.
Among different parts of the plants, ethanolic extract of C. chinense leaf offered the highest TPC (1.03 g GAE/g extract), followed by stem extract (0.553 g GAE/g extract) and flower extract (0.487 g GAE/g extract) [18,19]. The results of total flavonoid content analysis showed that the stem of C. chinense exhibited the lowest concentration of total flavonoid content, followed by the leaf and flower [18,19]. These results indicated that each part of C. chinense contained different amounts of phenolic compounds and flavonoids.
2.5. Antioxidant Activities of C. chinense Stem Extract Nanoparticles and NPs
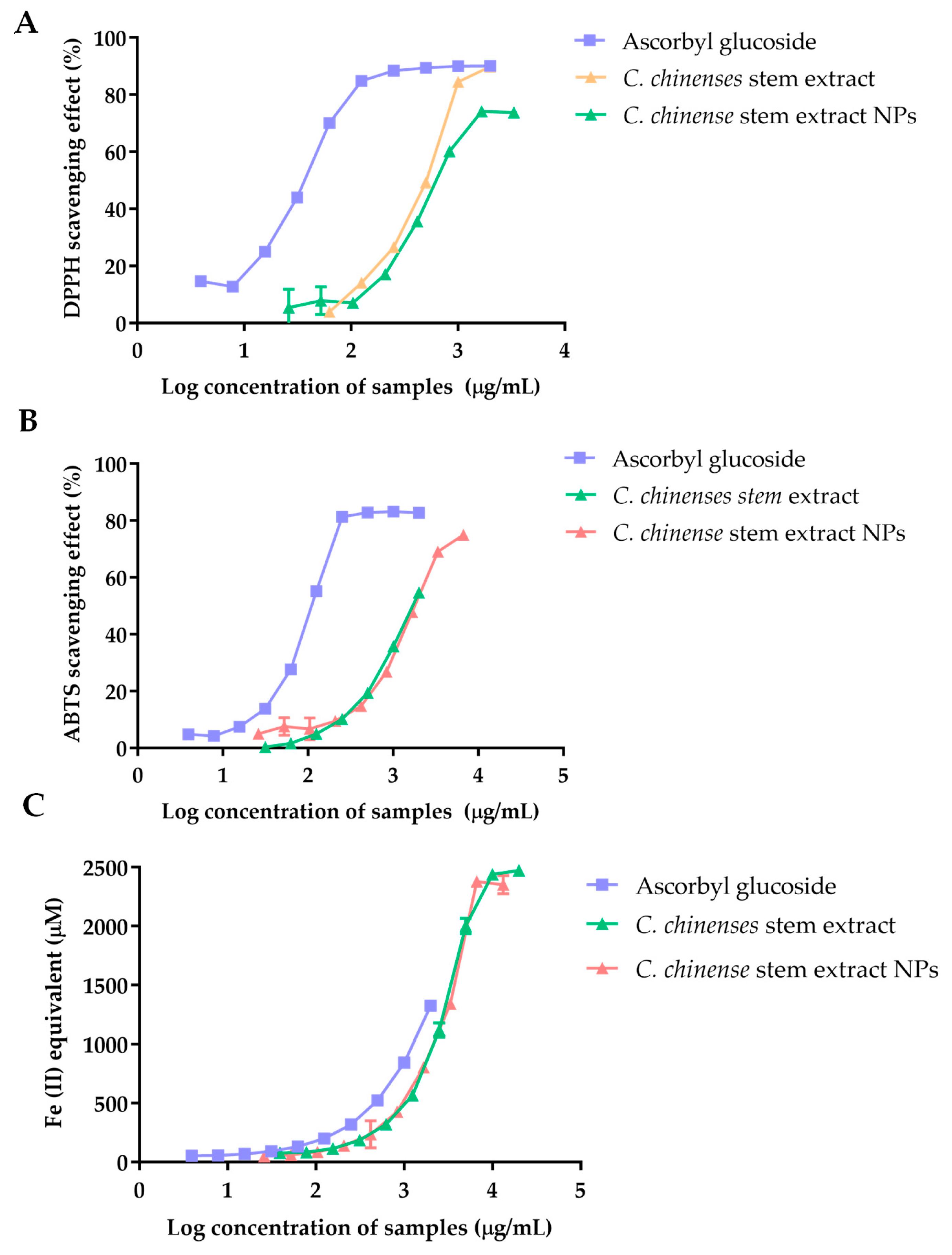
Figure 4 illustrates the 2,2-diphenyl-1-picrylhydrazyl (DPPH) scavenging activity of ascorbyl glucoside, the extract from C. chinense stems, and NPs. The scavenging ability against DPPH radicals was expressed as a percentage (%), and the results demonstrated a concentration-dependent relationship. A plateau effect was observed at higher concentrations, indicating the saturation of the free radical scavenging. The IC50 values were calculated as 35.71 µg/mL for ascorbyl glucoside, 457.1 µg/mL for the C. chinense stem extract, and 724.9 µg/mL for C. chinense stem extract NPs. The results suggested that the extract exhibited greater DPPH radical scavenging activity when compared to the NPs.

Figure 4.
The antioxidant activity of ascorbyl glucoside, C. chinense ethanolic stem extract, and NPs determined using (A) a DPPH free radical scavenging assay, (B) an ABTS free radical scavenging assay, and (C) a FRAP assay.
2,2′-Azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) radical scavenging activity of ascorbyl glucoside, C. chinense stem extract, and C. chinense stem extract NPs was correlated with the concentration of the samples. The higher ABTS radical scavenging activity was observed in C. chinense stem extract compared to its NPs. The IC50 values were reported at 117.2 µg/mL, 1688 µg/mL, and 1892 µg/mL for ascorbyl glucoside, extract, and NPs, respectively.
The reducing activity of ascorbyl glucoside, C. chinense stem extract, and NPs was measured using Ferric Reducing Antioxidant Power (FRAP) assay. Ascorbyl glucoside at a concentration range of 3.9–2000 µg/mL had a FRAP value of 52.52 ± 1.25 to 1324.48 ± 8.84 µM. The FRAP values of C. chinense stem extract and NPs were observed at 76.96 ± 2.93 to 2471.94 ± 15.16 µM and 48.04 ± 3.95 to 2350.52 ± 77.74 µM, respectively. It was found that the ferric reducing power of all samples depended on the concentration of the samples. A previous report showed that FRAP values of C. chinense leaf extract were in the range of 88.73 ± 4.59 to 2480.81 ± 0.00 µM. Therefore, C. chinense stem extract showed lower FRAP values compared with leaf extract. The lower antioxidant activities of C. chinense stem extract NPs might be due to the undissolved extract encapsulated in the NPs.
Deb and Sarkar have shown that the total polyphenol and verbascoside contents were directly proportional to the antioxidant potential of Clerodendrum glandulosum Lindl. leaves [22]. Funes et al. also reported the correlation between verbascoside in Aloysia triphylla and the significant ferric-reducing ability of plasma of this compound [23]. Therefore, it might be concluded that the ferric-reducing potential of C. chinense stem extract might be partly due to verbascoside.
The correlations between the total phenolic content and the total flavonoid content and the antioxidant activities of C. chinense stem extract and the NPs were calculated using the Pearson correlation coefficient. The results of the correlation are shown in Table 1.

Table 1.
Pearson correlation coefficients of total phenolic and flavonoid contents and antioxidant activities of C. chinense stem ethanolic extract and NPs measured using DPPH, ABTS, and FRAP assays.
2.6. Cytotoxicity of C. chinense Stem Extract and NPs against Cancer Cell Lines
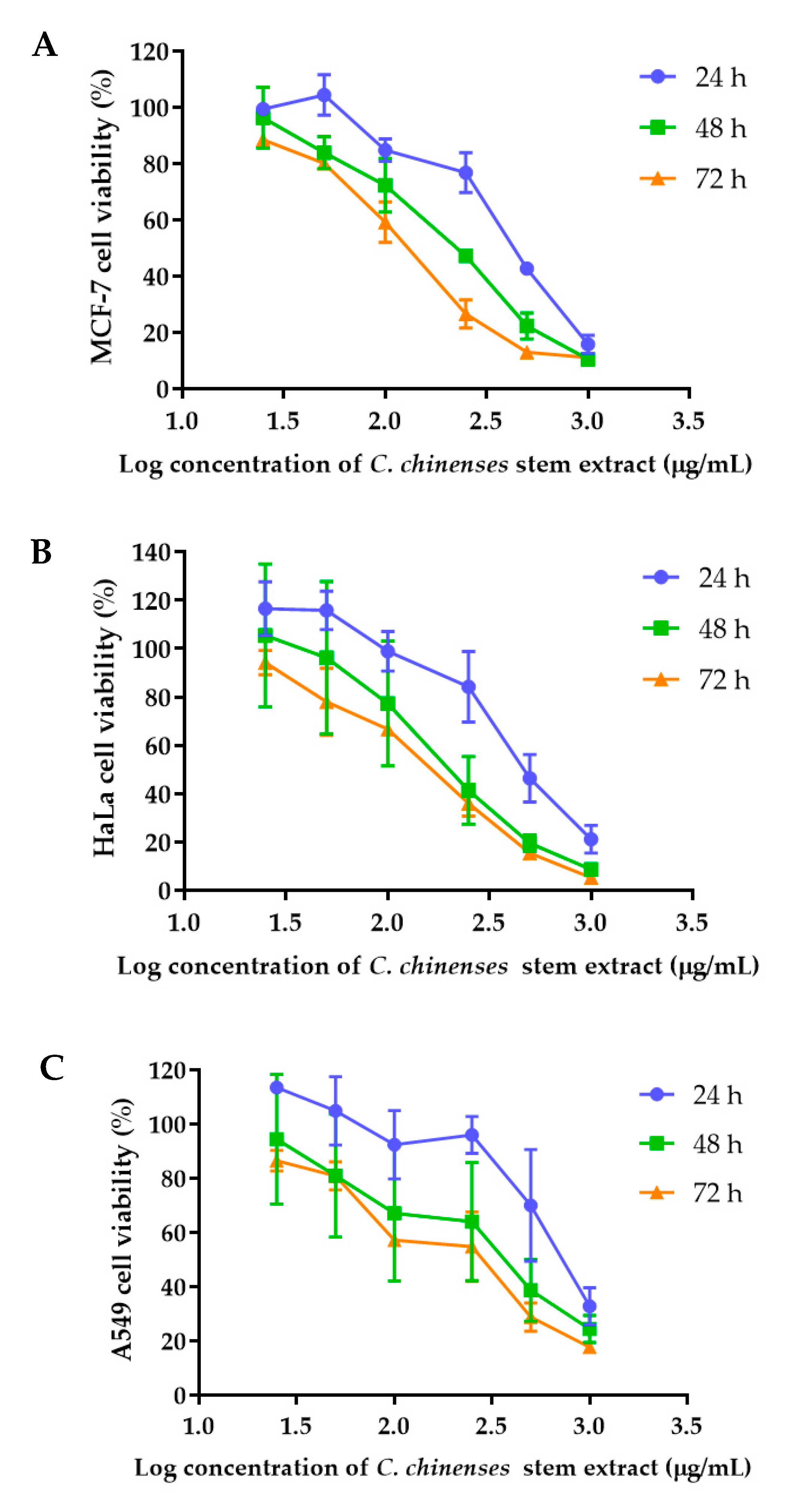
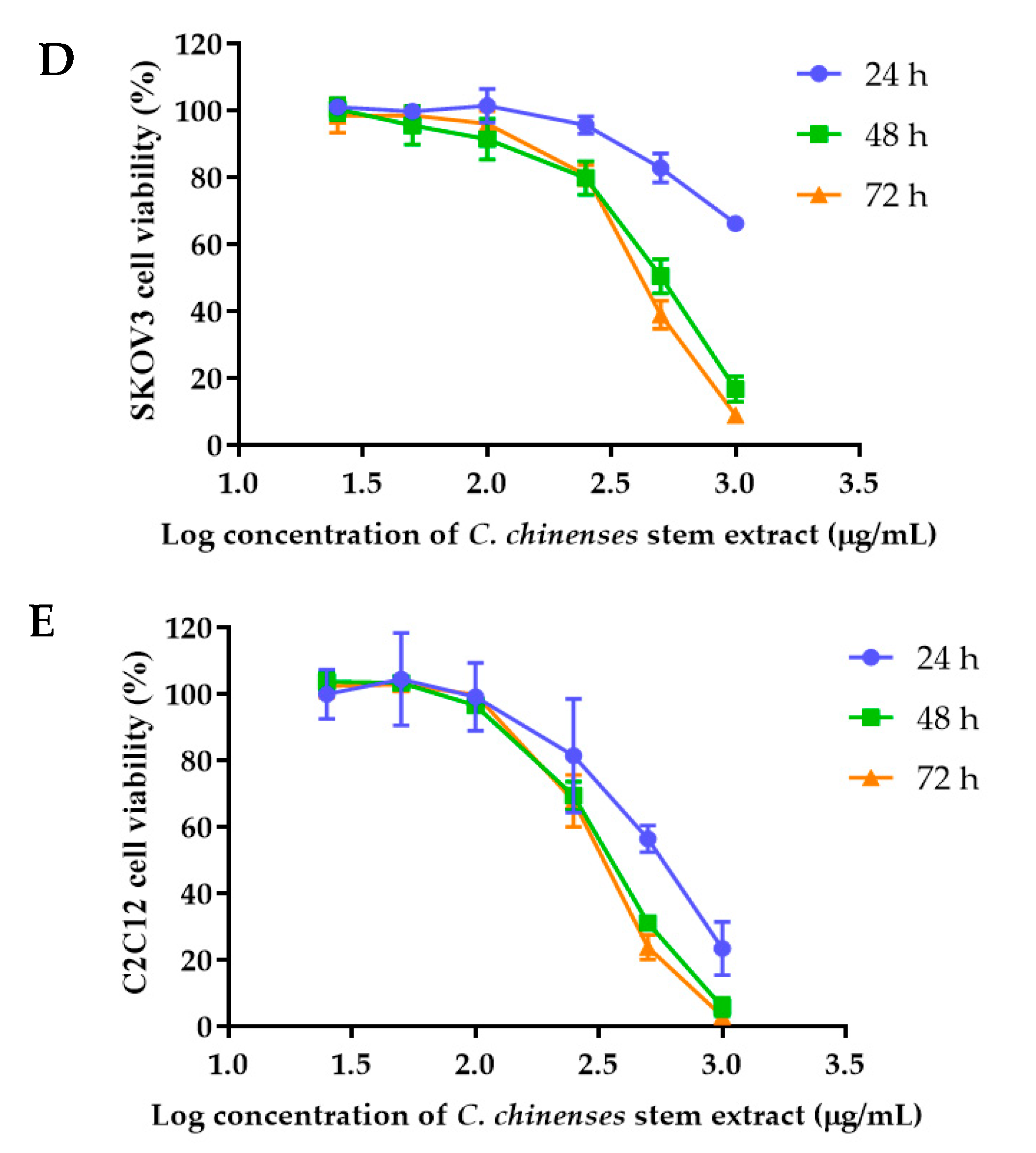
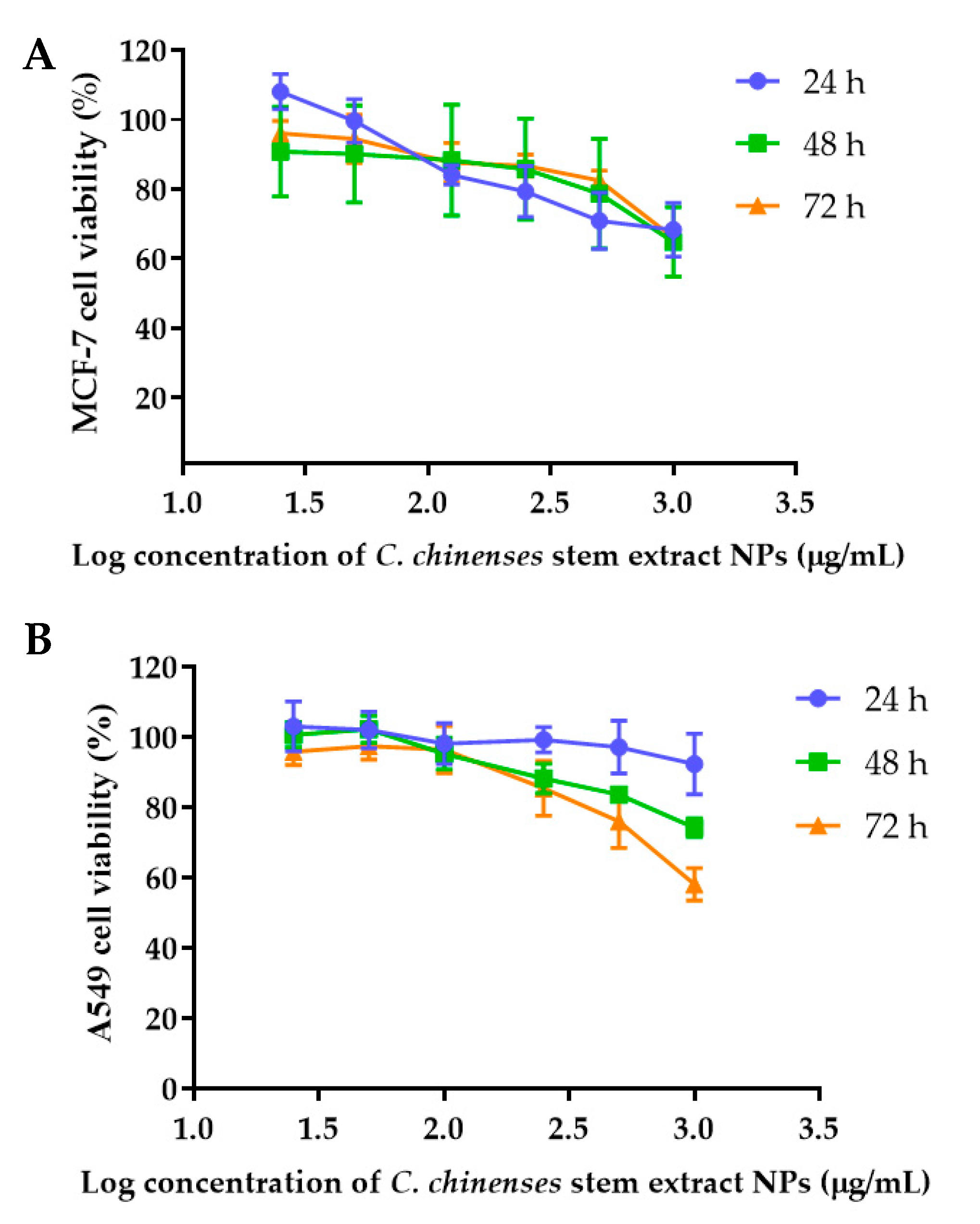
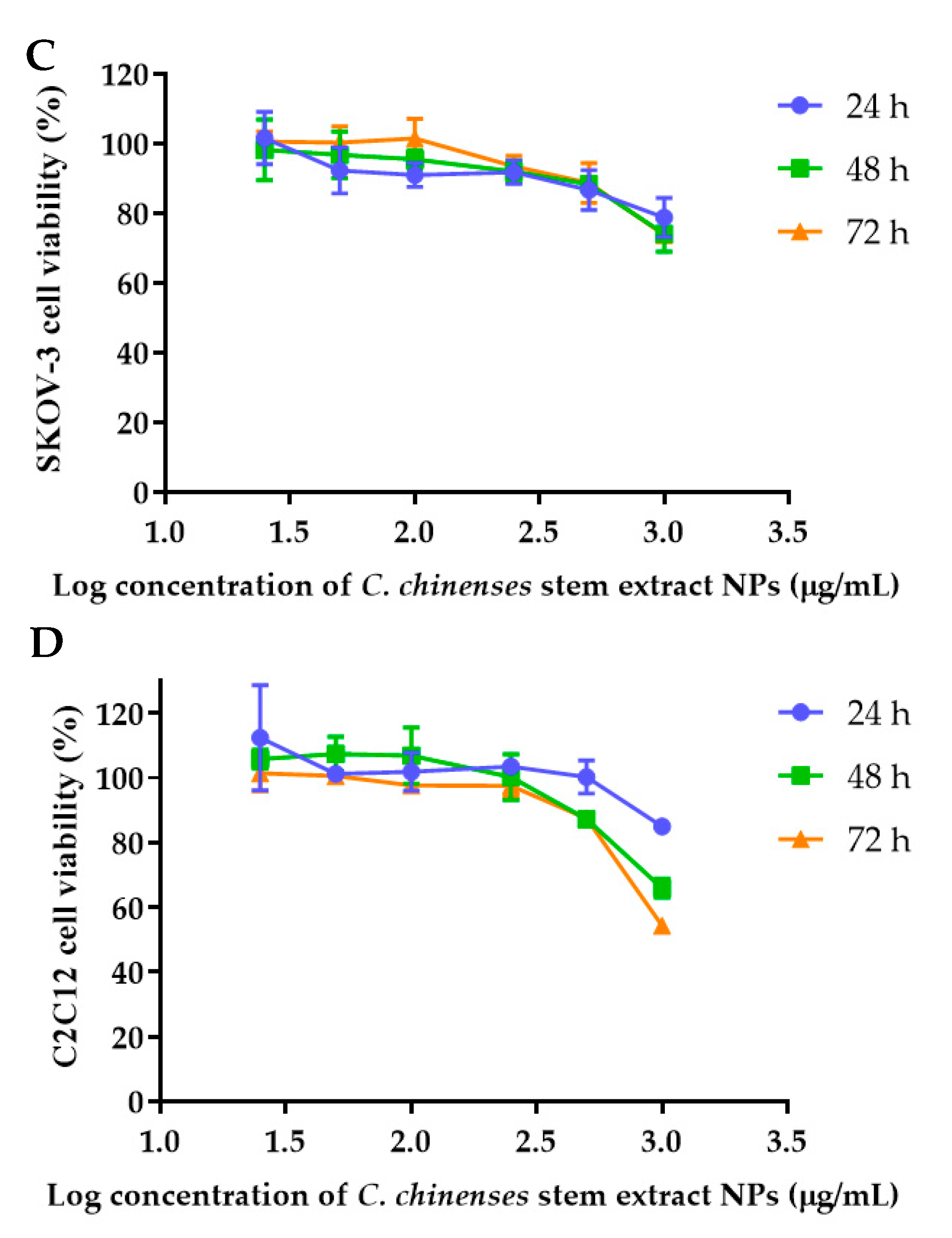
The anticancer activities of the C. chinense stem extract were tested for the first time in our study. To confirm the anticancer activity of the C. chinense stem extract, the cytotoxicity of the extract was tested in several cancer cell lines, including HeLa, A549, MCF-7, and SKOV-3 cells. The most sensitive cell line to this extract was further investigated for the anticancer activity mechanisms. C. chinense stem extracts exhibited a dose-dependent growth-inhibitory effect on the MCF-7 breast cancer cell line, the HeLa cervical cancer cell line, the A549 adenocarcinomic human alveolar basal epithelial cells, and the SKOV-3 ovarian cancer cells (Figure 5). C. chinense stem extract showed the lowest IC50 values against the MCF-7 breast cancer cell line compared with HeLa cells, A549, and SKOV-3 cells. The ethanol extract of C. chinense stem showed the most potent inhibitory effect with IC50 values of 109.2, 155.6, 206.9, and 2044 µg/mL on the MCF-7, HeLa, A549, and SKOV-3 cell lines, respectively. The MCF-7 growth-inhibitory effects were also detected in the ethanolic extract of C. chinense flower, with an IC50 value of 67.05 µg/mL. The lower inhibitory effect of most of the stem extracts compared with the flower extract might be due to the absence of hispidulin, which is considered one of the bioactive compounds in the C. chinense extract. NPs of C. chinense stem extract have demonstrated cytotoxicity against MCF-7, A549, and SKOV-3 cells (Figure 6). However, the cell-growth-inhibitory effect of the NPs encapsulating the extract was lower compared with the extract itself. The cytotoxicity of C. chinense stem extract and NPs was also investigated in normal myoblast cells, namely C2C12, to investigate the selectivity of the extract and the NPs. The cytotoxicity of both the extract and the NPs against C2C12 was found to be dose dependent. Both the extract and the NPs were less toxic to C2C12 cells compared to MCF-7 and A549 cells. Micrographs of A549, SKOV-3, and C2C12 cells treated with C. chinense stem extract or NPs at 1 mg/mL are shown in Supplementary Figure S6.
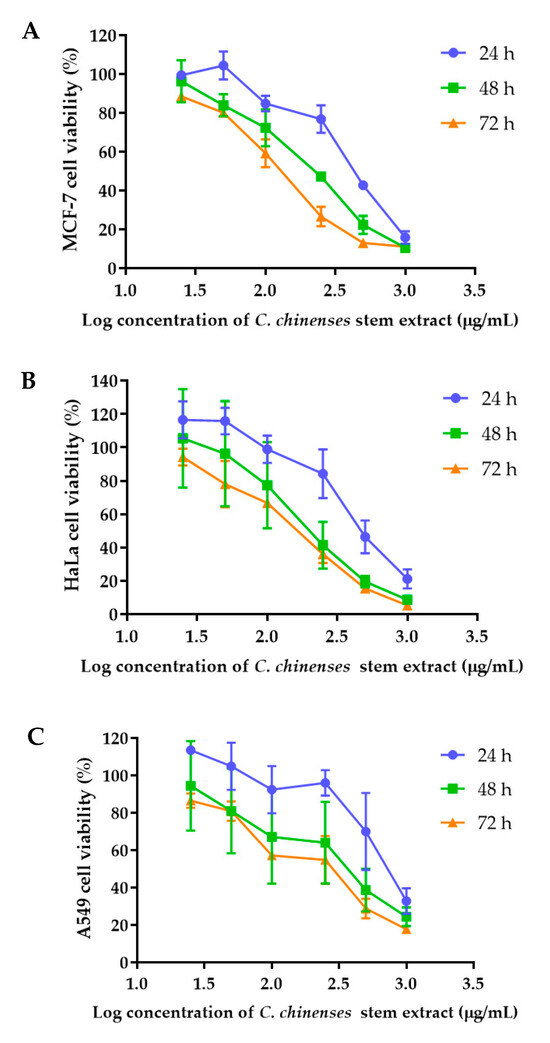
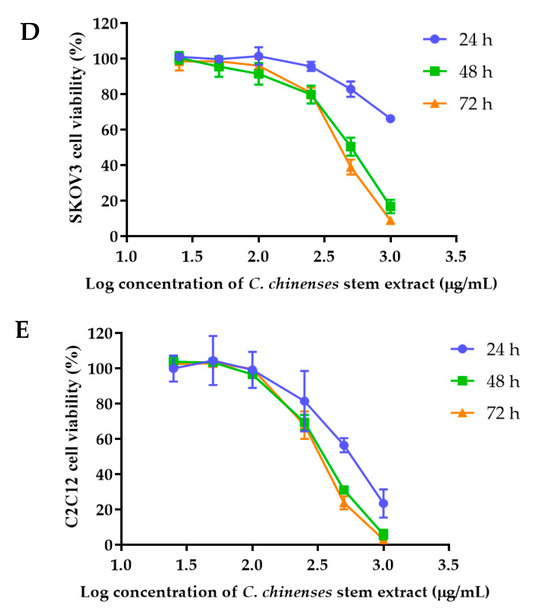
Figure 5.
Effect of C. chinense stem extract on viability and growth of (A) MCF-7 breast cancer cells, (B) HeLa cervical cancer cells, (C) A549 lung cancer cells, (D) SKVO-3 ovarian cancer cells, and (E) C2LC12 myoblast cells. Data are presented as the mean ± SD of the three independent experiments.


Figure 6.
Effect of C. chinense stem extract NPs on viability and growth of (A) MCF-7 breast cancer cells, (B) A549 lung cancer cells, (C) SKVO-3 ovarian cancer cells, and (D) C2CL12 myoblast cells. Data are presented as the mean ± SD of the three independent experiments.
The selectivity index of C. chinense stem extract and NPs, calculated by dividing the IC50 value of samples against noncancerous cells (C2C12) with that of samples against cancerous cells (MCF-7, HeLa, A549, and SKOV-3 cells), is shown in Table 2. It was found that C. chinense stem extract had the highest selectivity index values against MCF-7 cells compared with HeLa, A549, and SKOV-3 cells. Therefore, C. chinense stem extract not only showed the highest cytotoxicity against the MCF-7 cell line but also the highest selectivity to this cell line. For this reason, we further developed the NPs of C. chinense stem extract and tested their toxicity and selectivity index on MCF-7 cells. The results were confirmed by using A549 and SKOV-3 cells. The NPs of C. chinense stem extract have been shown to be preferably selectively toxic to MCF-7 and A549 cells as opposed to C2C12 cell lines. The %cell viability of MCF-7 and C2C12 after exposure to the extract and the NPs were compared at each time point. The results are shown in Supplementary Figure S7. NPs had lower IC50 values against C2C12 cells compared with the SKOV-3 cancer cell line, suggesting that C. chinense stem extract NPs are not selective to this type of cancer cell line. The selectivity index was greatest when cells were incubated with NPs for 24 h and decreased after incubation for 48 and 72 h. These results suggested that as the incubation time increased, the NPs may exhibit more toxicity towards healthy cells or tissues.
Table 2.
IC50 values and selectivity index of C. chinense stem extract and NPs against cancerous and non-cancerous cells.
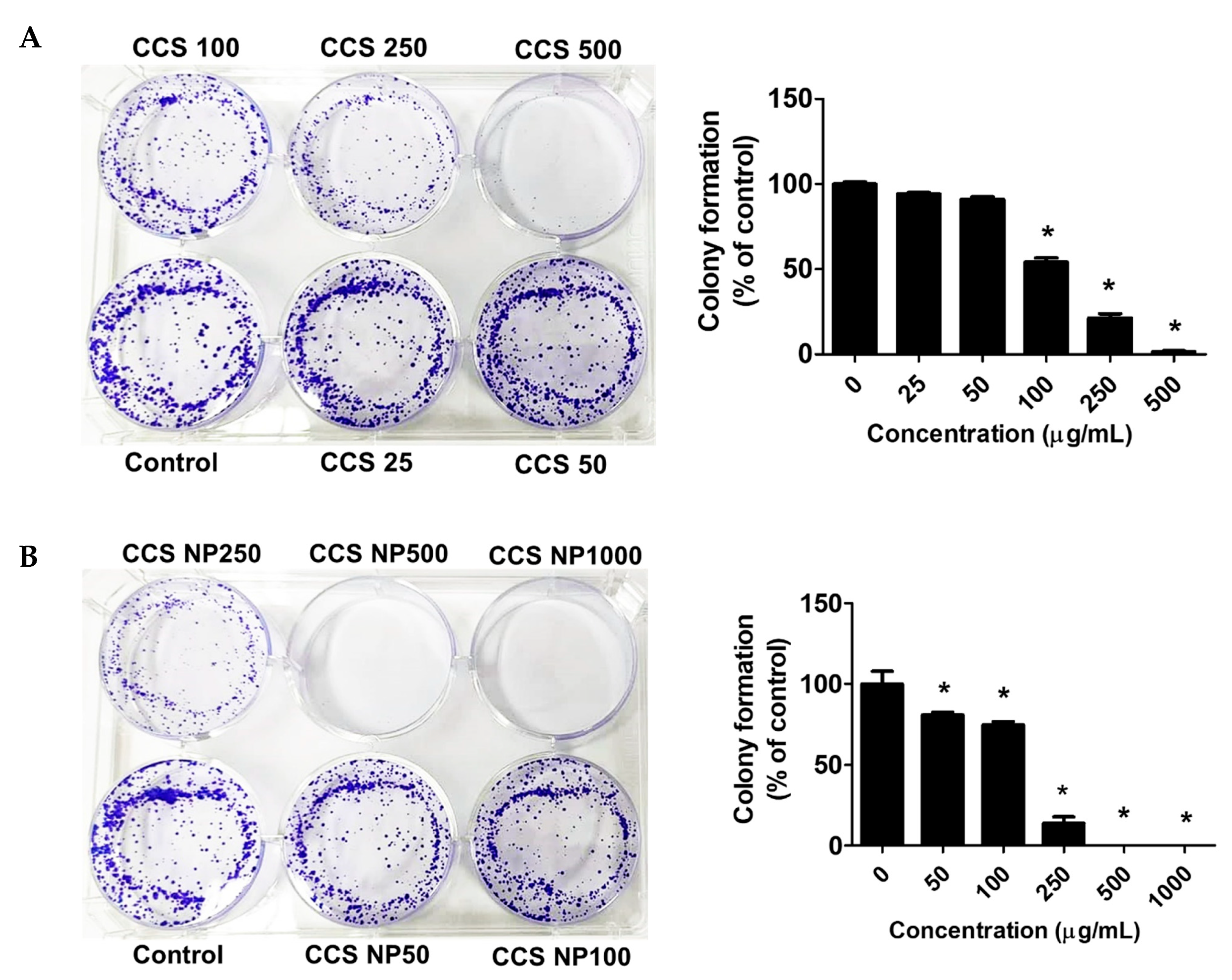
2.7. C. chinense Stem Extract and NPs Inhibited Colony Formation of MCF-7 Cells
The antiproliferative activity of the extract was further evaluated using the clonogenic survival assay. Colony formation assays suggested that the colony-forming ability of the MCF-7 cells was remarkably reduced in a dose-dependent manner after treatment with C. chinense stem extract and NPs for 24 h (Figure 7). MCF-7 cells formed 100.00 ± 1.99% without treatment. MCF-7 cells treated with 25, 50, 100, 250, and 500 µg/mL C. chinense stem extract showed 94.05 ± 1.60%, 90.91 ± 2.69%, 54.02 ± 4.48%, 21.33 ± 4.21%, and 1.40 ± 1.09% colony formation, respectively, compared with the control. MCF-7 cells treated with 50, 100, and 250 µg/mL NPs showed 80.81 ± 3.08%, 74.81 ± 3.69%, and 13.76 ± 6.94% colony formation, respectively, compared with the control. When treated with 500 and 1000 μg/mL of NPs, all of the cells were unable to form visible colonies. The ability of C. chinense stem extract and NPs to affect long-term colony formation may be in part through both suppression and/or induction of cell death [24]. C. chinense stem extract at 500 µg/mL reduced MCF-7 colony formation to 1.40 ± 1.09%, while NPs at the same concentration completely inhibited cancer cell colony formation.
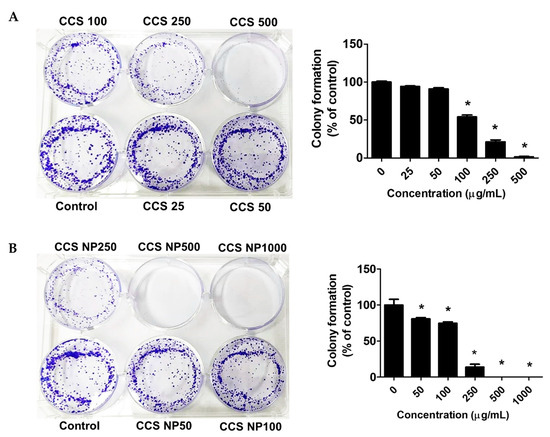
Figure 7.
Effects of (A) C. chinense stem extract and (B) NPs on colony formation of the MCF-7 cancer cell lines. Data are presented as the mean ± SD of the three independent experiments. * indicates p < 0.05. Tukey’s multiple comparisons test for colony formation percentage statistical analysis are shown in Supplementary Table S4.
2.8. C. chinense Stem Extract and NPs Promoted Apoptosis in MCF-7 Cells
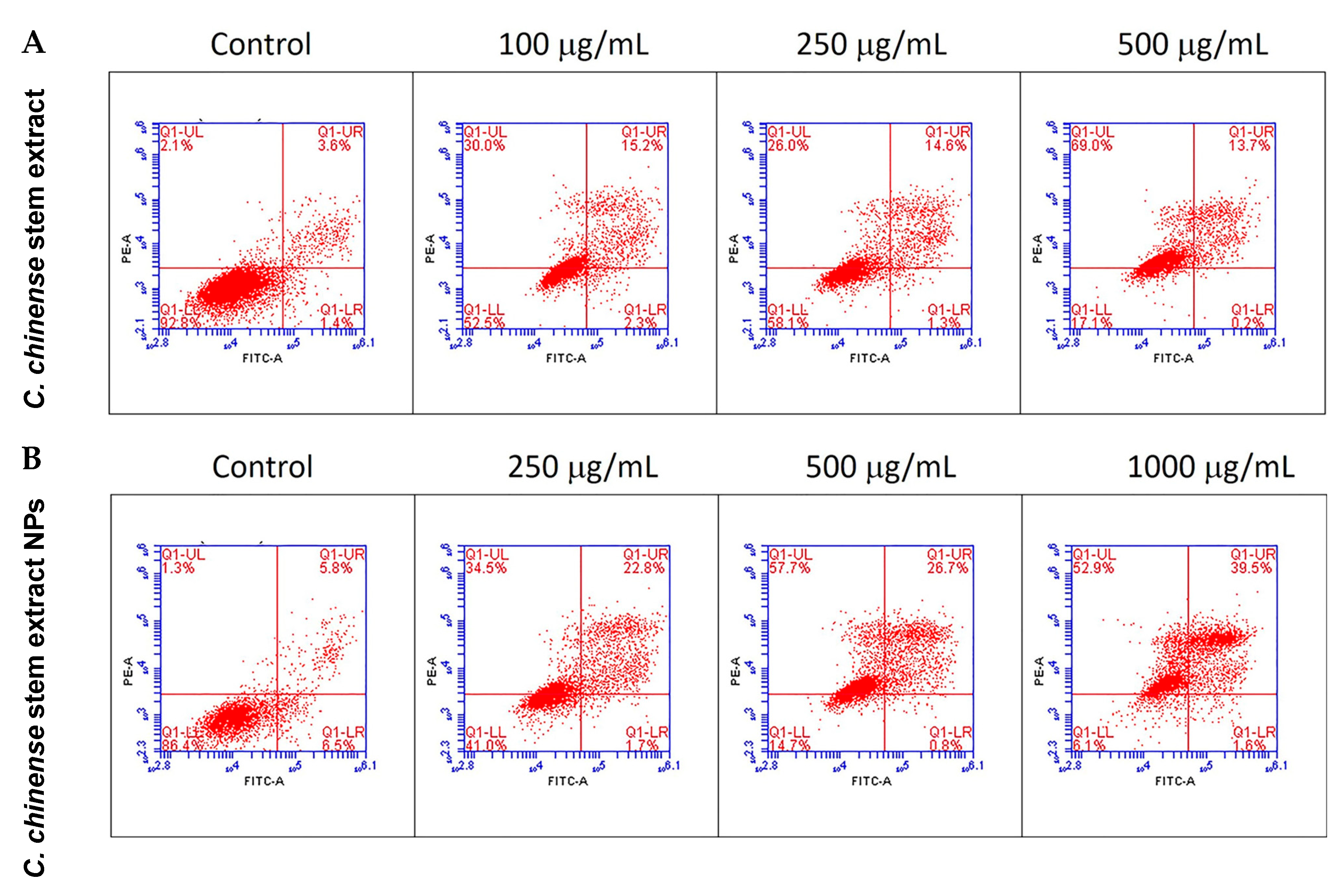
The apoptotic effect of the C. chinense stem ethanolic extract was investigated by staining the cells with the Annexin V-FITC apoptosis detection kit and evaluation through flow cytometry. Viable cells were identified as those negative for both propidium iodide (PI) and annexin V. Early apoptotic cells were characterized as PI negative with annexin V staining and reduced cell size. Necrotic cells were identified as PI-positive cells with either normal or enlarged cell size. Late apoptotic cells were defined as PI-positive cells exhibiting annexin V staining and diminished cell size [25]. The results showed that C. chinense stem extract and NPs induced MCF-7 cytotoxicity through the mechanisms associated with apoptosis and necrosis in a concentration-dependent manner (Table 3, Figure 8). The late apoptosis and necrosis of the negative control group were 3.3–5.7% and 1.27–2.17%, respectively. About 14.07%, 14.77%, and 12.13% of the MCF-7 cells experienced late apoptosis at 100, 250, and 500 µg/mL of the C. chinense stem extract, while the NPs induced 26.73%, 25.57%, and 40.13% of late-apoptotic MCF-7 cells at 250, 500, and 1000 µg/mL, respectively.
Table 3.
Analysis of cell apoptosis induced by C. chinense stem extract and NPs.
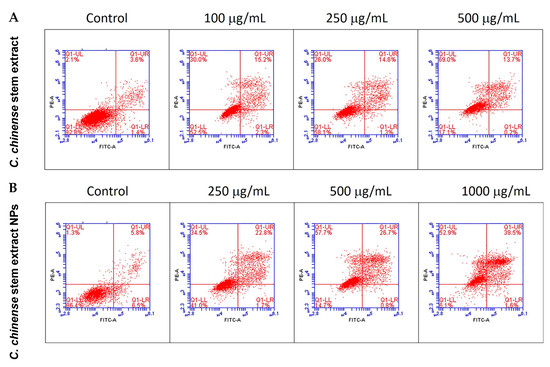
Figure 8.
Flow cytometry analysis with Annexin V-PI staining was performed to evaluate the percentage of apoptotic cells in (A) C. chinense stem extract and (B) NP-induced MCF-7 cells.
2.9. C. chinense Stem Extract and NPs Decreased Mitochondrial Membrane Potential (MMP)
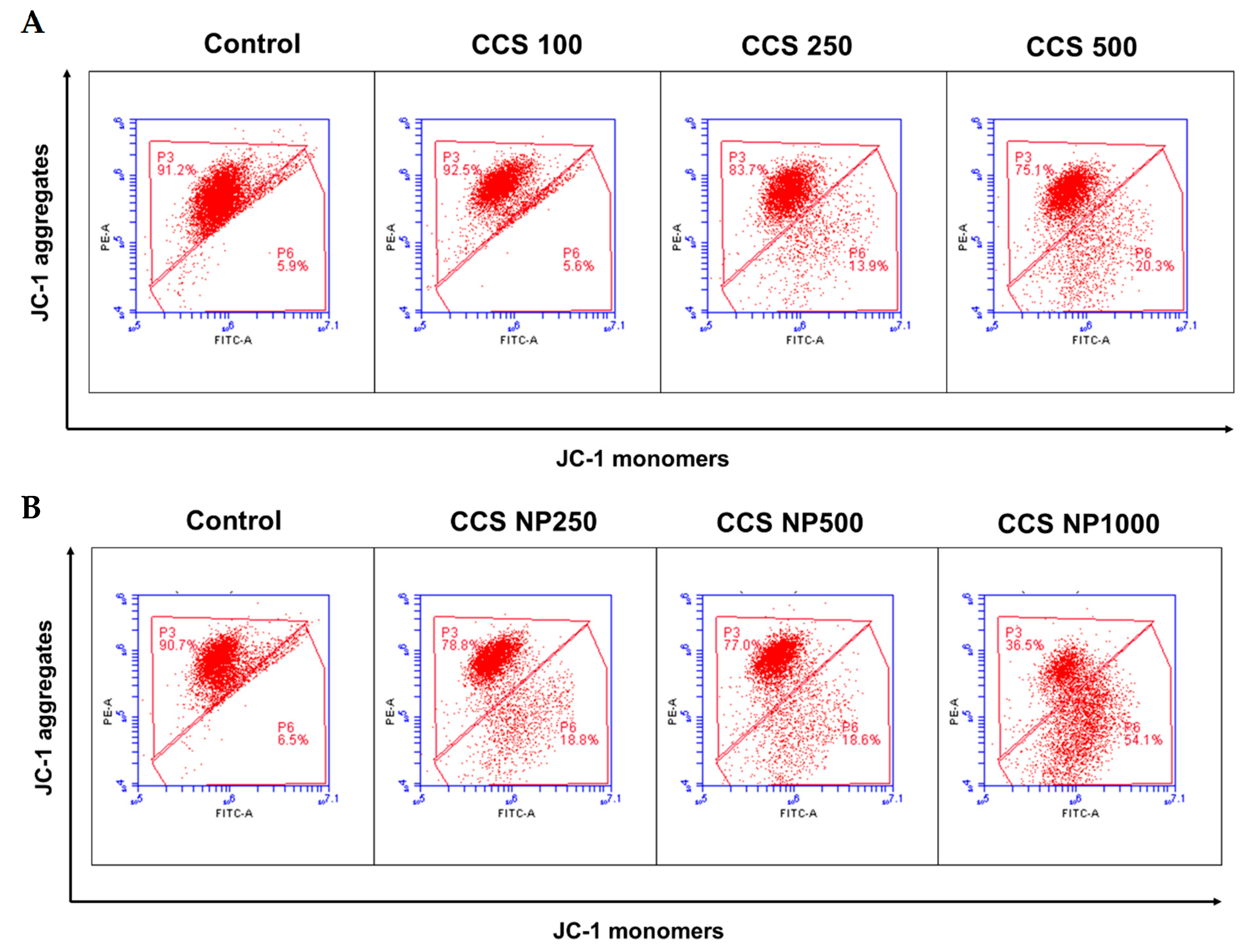
Flow cytometry analysis of 5,5,6,6-tetrachloro-1,1,3,3-tetraethylbenzimidazolylcarbocyanine iodide (JC-1) staining demonstrated a significantly decreased fluorescence intensity of aggregates and an increased fluorescence intensity of monomers (Figure 9). MCF-7 cells exposed to C. chinense stem extract at 500 µg/mL and NPs at 250 and 500 µg/mL showed a significantly higher percentage of JC-1 monomers compared to the control group (Table 4). The ratio of JC-1 aggregates to JC-1 monomers decreased with the concentration of C. chinense stem extract and NPs. The ratio of monomers and aggregates significantly increased from 0.066 in the control group to 0.20 in the extract at 500 µg/mL in the treated group and to 1.38 in NPs in the 1000 µg/mL group, indicating a dramatic reduction in MMP and resultant mitochondrial dysfunction in the C. chinense extract and NP-treated groups. These results suggested that the mitochondrial dysfunction that resulted from the extract and NPs was dose dependent. The ratios of monomers to aggregates were increased when cells were treated with NPs compared to the extract at all tested concentrations. Ratios of monomers to aggregates when cells were treated with 250 and 500 µg/mL extract were 0.13 and 0.20, respectively. NPs at 250 and 500 µg/mL increased ratios of monomers to aggregates to 0.185 and 0.233, respectively. This result suggested that NPs decreased mitochondrial membrane potential to a greater extent compared with the extract.

Figure 9.
Flow cytometry analysis of the JC-1 assay (mitochondrial membrane potential) in MCF-7 cells treated with (A) C. chinense stem extract and (B) NPs.
Table 4.
Percentage of JC-1 aggregates, JC-1 monomers, and ratio of monomers to aggregate MCF-7 cells treated with C. chinense stem extract and NPs.
2.10. C. chinense Stem Extract and NP-Induced Cell Cycle Arrest
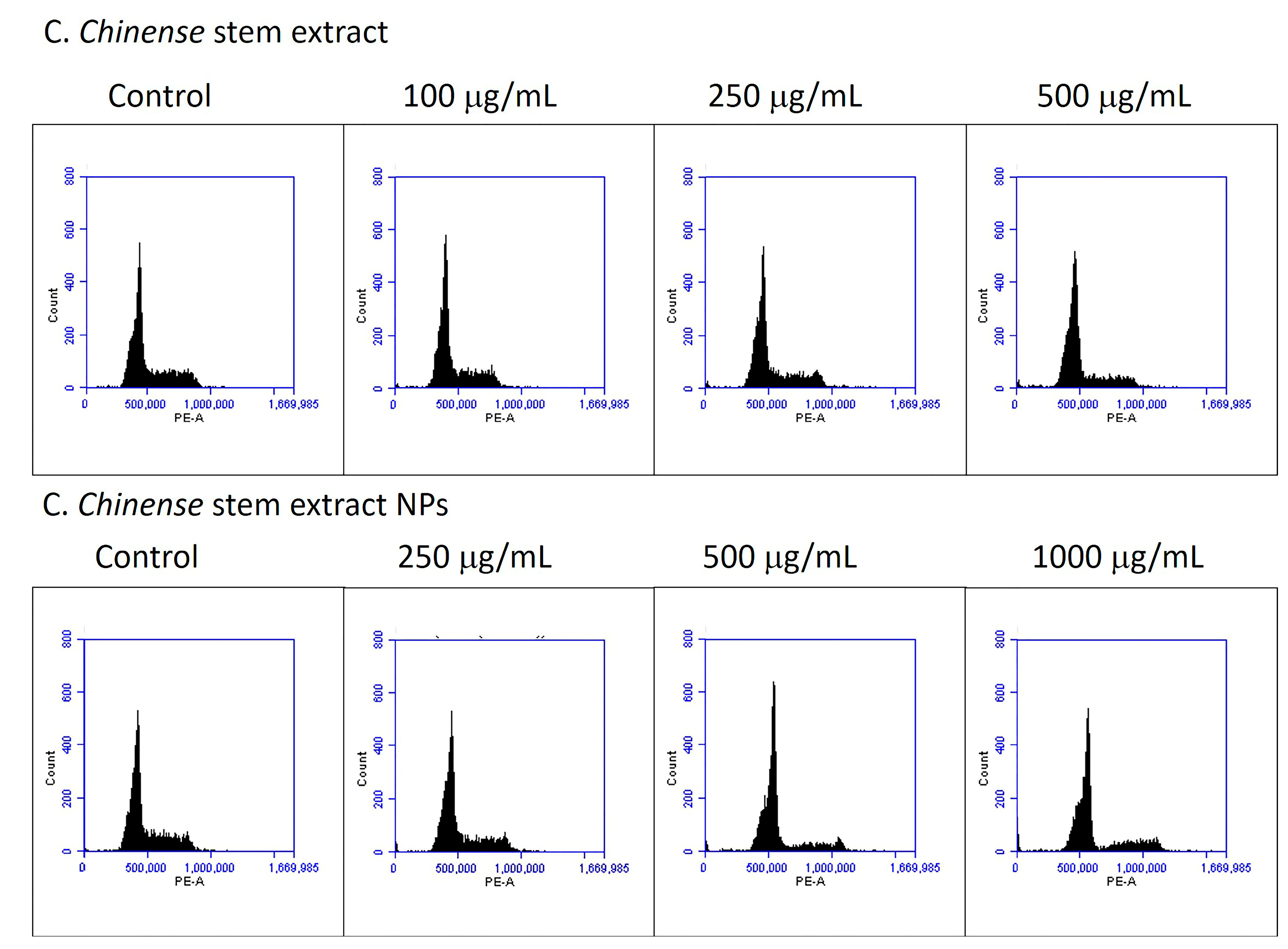
Cell cycle distribution was analyzed to further investigate the potential mechanism by which C. chinense stem extract and NPs depressed MCF-7 cell growth. MCF-7 cell lines were treated with C. chinense stem extract and NPs for 24 h followed by flow cytometry assays. The representative cell cycle distributions of MCF-7 cells exposed to C. chinense stem extract and NPs are presented in Figure 10. The results show that C. chinense stem extract caused G0/G1 phase arrest in MCF-7 cells at a concentration of 500 µg/mL (Table 5). In particular, the proportion of G0/G1 phase cells significantly increased as the concentration of the extract increased, which was accompanied by a decrease in the number of cells in the S phase and the G2/M phase. Therefore, the effect of C. chinense stem extract and NPs on the proliferation of MCF-7 cells is likely associated with cell cycle arrest.
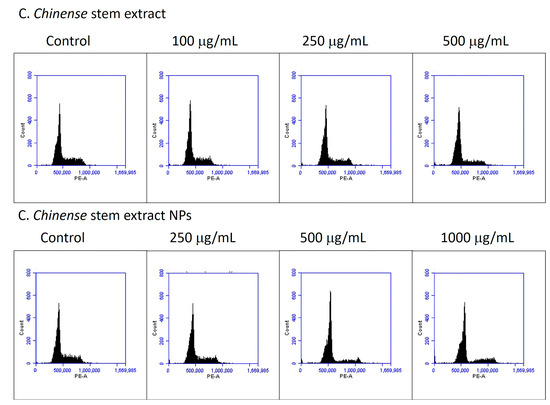
Figure 10.
C. chinense stem extract and NPs induce G0/G1 phase cell cycle arrest.
Table 5.
The percentage of MCF-7 cells treated with C. chinense stem extract and NPs in each phase of the cell cycles.
3. Discussion
The yield of stem extraction was significantly lower than that of the flower and leaves of C. chinense when the same solvent and extraction method were used for extraction. However, the bioactive compound contents in the stem extract were comparable to those of the flower extract. The stem extract contained a lower amount of 9.42 ± 0.15 µg verbascoside/mg of extract, while the flower extract contained 11.27 ± 0.08 µg/mg of extract. Hispidulin was negligible in the stem extract, while the flower extract contained a low amount of hispidulin (0.98 ± 0.01 µg/mg extract). Isoverbascoside content was higher in the stem compared with the flower extract (8.32 ± 0.15 µg/mg extract vs. 4.30 ± 0.27 µg/mg extract). The cytotoxicity investigations demonstrated that the flower extract exhibited lower IC50 values compared to the stem extract, signifying greater cytotoxicity in the flower extract. These findings imply a correlation between yield, bioactive compounds, and cytotoxicity.
The antioxidant activity observed in the C. chinense extract and NPs may be attributed to the presence of phenolic acids and flavonoids. The correlations between TPC and TFC and the antioxidant activities of C. chinense stem extract and NPs were calculated using the Pearson correlation coefficient. The results of the correlation are shown in Table 1. Pearson correlation coefficients revealed that the antioxidant activity of C. chinense extract NPs depended on the total phenolic and flavonoid contents.
The DDPH and ABTS free radical scavenging activity and the ferric-reducing power of the C. chinense extract correlated with the total phenolic content, whereas only the ABTS free radical scavenging activity and ferric-reducing power of the C. chinense extract correlated with the total flavonoid content. These natural phenolic compounds are recognized for their antioxidant properties, which can mitigate the risk of cancer diseases associated with oxidative stress. It is noteworthy that certain phenolic compounds, including phenolic acids, flavonoids, and non-flavonoids, can exhibit prooxidant behavior under specific conditions [26]. A high concentration of phenolic compounds and elevated pH levels may contribute to their prooxidant activity, leading to the induction of apoptosis and necrosis in cancer cells [27]. Hence, it is crucial to note that the behavior of phenolic compounds as either antioxidants or prooxidants depends on the redox homeostasis of the cells.
In this study, it was observed that the C. chinense stem extract NPs exhibited inhibitory effects on the growth of MCF-7 cells at concentrations of 500 and 1000 μg/mL after 24 h of incubation. The cytotoxicity of the NPs on MCF-7 breast cancer cells was found to be dose dependent. The IC50 values of the NPs were consistently higher than those of the extract across all incubation time points. Compared with the NPs, the %cell viability of the 1 mg/mL extract after 72 h of incubation was six-fold. These findings imply that the poloxamer 407 used to stabilize the NPs might mitigate the toxicity of the extract, possibly by delaying its release to the cells.
The cytotoxicity and antiproliferative effects of the C. chinense stem extract and NPs might result from verbascoside, as it has been shown that verbascoside has a cytotoxic effect on MCF-7 breast cancer cells. Senol er al. reported that the IC50 values for the MCF-7 BC cell line after 24, 48, and 72 h of exposure to a different concentration of verbascoside were 0.127, 0.2174, and 0.2828 μM, respectively [28]. Verbascoside isolated from Scrophularia subaphylla L. exhibited cytotoxicity on MCF-7 cells, and the IC50 value was found to be 0.39 ± 0.015 μg/mL after 48 h of exposure [29]. Verbascoside significantly inhibits colorectal cancer cell growth in vivo, represses cell proliferation, and promotes apoptosis by modulating the HIPK2-p53 signaling pathway [30]. Isoverbascoside caused a G0/G1 cell cycle arrest, thereby inhibiting the cell proliferation of MGC 803 gastric cancer [31].
C. chinense stem extract NPs demonstrated better MCF-7 cell colony formation inhibition compared with the extract. This effect might be due to the released bioactive compounds during 15 days of incubation. Several studies have reported the effects of phenolic compounds on cancer cell colony formation. Zhang et al. showed that different doses of glycyrrhizinic acid had a tendency to inhibit or suppress the colony formation tendency of these MCF-7 breast cancer cells [32]. Epigallocatechin gallate (EGCG) was shown to inhibit colony formation and induce apoptosis of MCF-7 breast cancer cells via down-regulation of survivin, a major member of the IAP gene family [24].
The results indicated that at 500 µg/mL, stem extract can induce 12.13% of late-apoptotic cells, while NPs induced 25.57% of apoptotic cells. The enhancement of apoptosis induction by NPs encapsulating C. chinense stem extracts can be attributed to several factors, including enhancing the bioavailability of bioactive compounds present in the extract. The NPs may improve the solubility and stability of the bioactive compounds, thus providing better absorption by the cells. This increased bioavailability can lead to more effective interactions with cellular components, promoting apoptosis. NPs can facilitate the cellular uptake of C. chinense stem extract. The small size and unique surface properties of nanoparticles may enhance their interaction with cell membranes, thus promoting internalization of the encapsulated or surface-bound bioactive substances.
MMP serves as a key indicator of mitochondrial activity, and a decline in MMP is often indicative of mitochondrial dysfunction. To investigate MMP alterations, we utilized JC-1, a dye specifically designed for mitochondria. JC-1 exhibits red fluorescence when forming aggregates in healthy mitochondria. As the membrane potential decreases, JC-1 transitions into monomers, thus resulting in green fluorescence. The alteration in the ratio of red to green fluorescence serves as an indicator of the mitochondrial condition under examination. The results showed that the C. chinense stem extract and the NPs can decrease the mitochondrial membrane potential in a concentration-dependent manner.
Many studies have demonstrated that polyphenols induce apoptosis and dissipate mitochondrial membrane potential in cultured cancer cells. Curcumin induced apoptosis in human gastric adenocarcinoma SGC-7901 cells through dissipation of the mitochondrial membrane potential [33]. Honokiol induced apoptosis and decreased mitochondrial membrane potential of neuroblastoma neuro-2a cells at 40 μM exposure [34]. EGCG inhibited HeLa cervical cancer cells in a concentration- and time-dependent manner by inducing apoptosis, including a decrease in the mitochondrial membrane potential [35]. Qanungo et al. reported that EGCG can indeed promote apoptosis in pancreatic cancer through mitochondrial membrane depolarization [36]. Polyphenols extracted from pinecones of Pinus koraiensis inhibited tumor growth in sarcoma-bearing 180 mice by activating the mitochondrial apoptotic pathway [37].
Both C. chinense stem extract and NPs at all tested concentrations significantly induced cell cycle arrest at the G0/G1 phase. At 500 µg/mL, the C. chinense stem extract NPs increased cells at the G0/G1 phase by 74.63%, which was higher than the extract (71.40 ± 0.56%). The results suggested that the component in NPs did not affect the cell cycle arrest of the extract. Many studies have showed that polyphenolic compounds can suppress the cell cycle. Analysis of a polyphenol-enriched extract from selenium-enriched Ziyang green tea (ZTP) suggested that the treated cells were subjected to a blockage at the G1 phase of the cell cycle, suggesting that ZTP blocked the progression of the cell cycle at the G0/G1 phase by modifying p53 and CDK2 expression [38]. A polyphenol-enriched extract obtained from Salvia chinensis induced G0/G1 cell cycle arrest and induced a significant and concentration-dependent reduction in the mitochondria membrane potential of the MiapaCa-2 human pancreatic cancer cells [39].
The anticancer activity of C. chinense stem extract may result from verbascoside and isoverbascoside. The anticancer activity of verbascoside has been reported. Verbascoside is associated with cell cycle arrest and apoptosis of breast cancer cells, including MCF-7 and MDA-MB-231 cells. Verbascoside exhibited a dose-dependent impact on the modulation of cell cycle and apoptosis-related proteins. The levels of CyclinB1, Cdc2, Bcl-2, and survivin were reduced, whereas cleaved PARP1, BAX, and cleaved caspase3/9 increased. Mechanistically, verbascoside inhibited the PI3K/AKT signaling pathway [40]. In another study, verbascoside elevated the population of subG1 cells and increased cell apoptosis rates. Additionally, it demonstrated time-dependent generation of reactive oxygen species (ROS) in tumor cells within 1–24 h of incubation [41].
4. Materials and Methods
4.1. Preparation of C. chinense Stem Extract
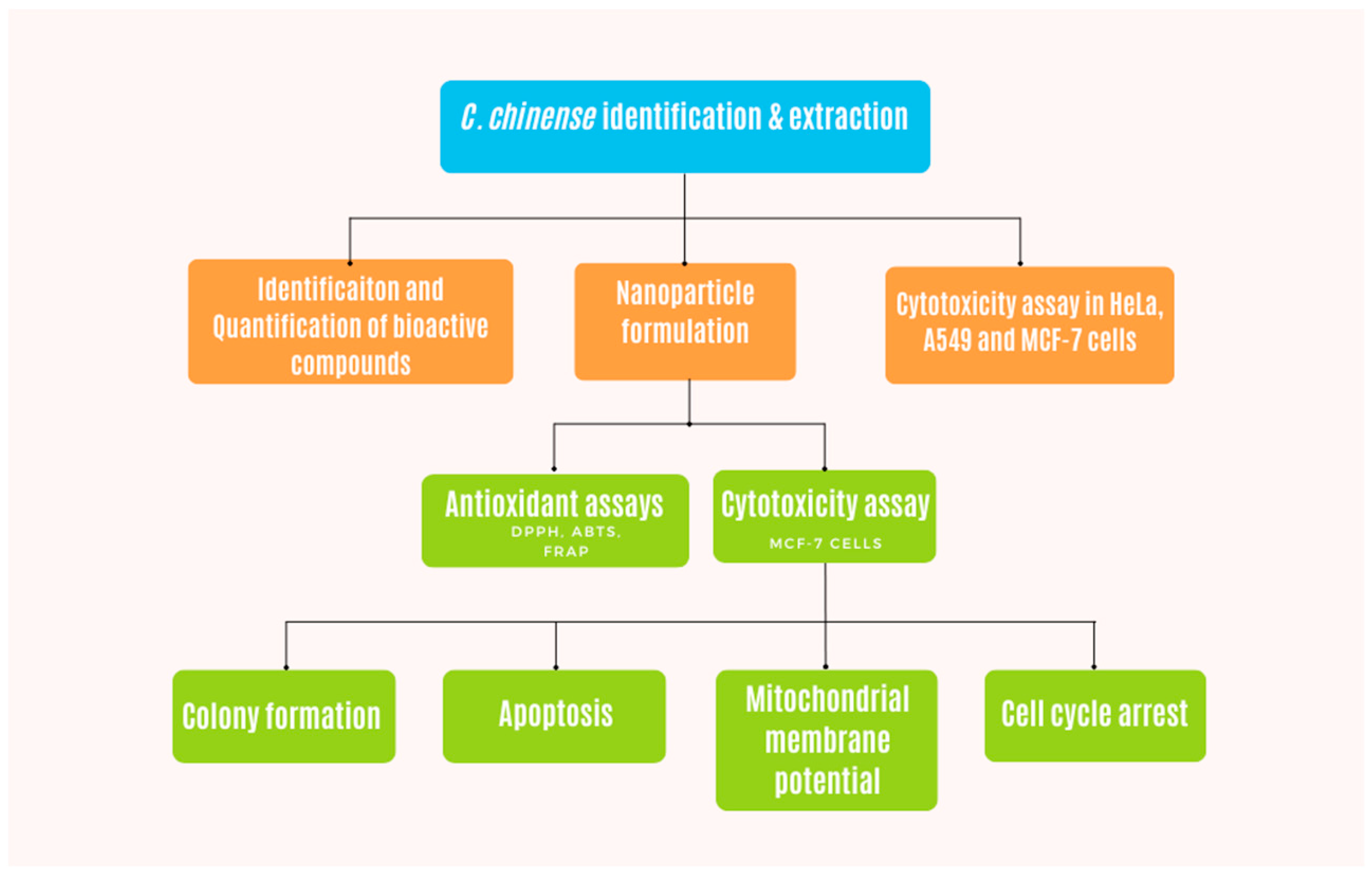
Figure 11 demonstrates the flow chart of experiments performed with C. chinense extract in this study. Stems of C. chinense were collected from Chiang Mai, Thailand (Figure 12). The plant was identified by Assistant Professor Sirivan Athikomkulchai with a voucher specimen named SIRA003. The voucher specimen was stored at the Faculty of Pharmacy, Srinakharinwirot University, Nakhonnayok, Thailand. The stems of C. chinense were thoroughly washed with water and dried at room temperature. Then, the stems were ground to fine powder using a kitchen blender. Next, 10 g of the stem powder was mixed with 100 mL of 95% ethanol. The extract was filtered with Whatman filter paper No 1. The filtrate extract was dried to a crude extract using a rotary evaporator and stored in the vacuum desiccator for further studies. The yield (%w/w) of the extract was calculated using the following equation [19]:

Figure 11.
Flow chart demonstrating the methodology of this work.

Figure 12.
C. chinense shrub.
4.2. Identification and Quantification of Bioactive Compounds in C. chinense Using High-Performance Liquid Chromatography
In the analysis of C. chinense stem extract, the bioactive compounds verbascoside, isoverbascoside, and hispidulin were identified and quantified. The identification and quantification were achieved through the implementation of a validated HPLC method employing an RP-C18 column (ACE 5 C18-AR, 250 × 4.6 mm, 5 μm) [42]. The mobile phase used for chromatography consisted of a mixture of acetonitrile (A) and 0.085% phosphoric acid in water (B), and a gradient elution mode was employed. Specifically, the elution conditions began with 5% eluent A and followed a linear gradient to reach 40% eluent A over the initial 20.0 min. Subsequently, another linear gradient increased the proportion of eluent A to 80% at 30.0 min and then maintained this ratio for an additional 5.0 min. The analysis was conducted at a constant flow rate of 1.0 mL/min, with an injection volume of 50 μL. The mobile phase was degassed prior to the analysis. The HPLC samples were filtered through a 0.45 μm membrane filter before analysis. Detection and quantification of the peaks were accomplished by monitoring the absorbance at 326 nm using a UV–visible detector (YL9120 UV/VIS). Identification of the peaks was achieved by comparing their retention times with those of standard verbascoside, isoverbascoside, and hispidulin. The quantification of these compounds in the extracts was achieved by constructing calibration curves and plotting the peak area against the standard concentration.
4.3. Formulation of C. chinense Stem Extract Nanoparticles
The preparation of C. chinense stem extract nanoparticles utilized the solvent displacement method. Initially, 200 mg of C. chinense stem extract was dissolved in 2 mL of 95% ethanol, forming the organic phase containing the extract. Afterward, the extract solution was centrifuged at 13,000 rpm for 3 min to remove any undissolved residue. Concurrently, an aqueous phase comprising 15 mL of a 0.1% poloxamer 407 solution was prepared. The organic phase was then introduced into the aqueous phase through gradual infusion at a controlled rate of 1 mL/h while maintaining continuous agitation at 700 rpm [18]. This dispersion process extended for 3 h to facilitate the evaporation of the organic solvent.
The C. chinense stem extract NPs were subjected to storage in different temperatures (4 °C, 30 °C, and 45 °C) and distinct time intervals (0, 0.5, 1, 2, 3, and 4 weeks). Following each incubation period, the samples were analyzed through dynamic light scattering and zeta potential measurements. The results were compared to the initial nanoparticle characterization.
4.4. Determination of Total Phenolic Content (TPC) in C. chinense Stem Extract and NPs
The analysis of the total phenolic content in C. chinense stem extract and NPs was carried out using the Folin–Ciocalteu colorimetric method [43]. In 96-well plates, a 50 µL aliquot of either the stem extract (at concentrations of 156.25–2500 µg/mL, 100 µL per well) or the NPs (at concentrations of 104.16–1666.63 µg/mL, 100 µL per well) was combined with 100 µL of 10% w/v Folin–Ciocalteu phenol reagent. After a 4 min incubation, 50 µL of 7.5% sodium carbonate was added, and the mixture was left at room temperature for 60 min. The absorbance of the resulting solution was measured at 725 nm. A standard calibration curve using gallic acid within the concentration range of 3.9–125 μg/mL was prepared in a similar manner, and the results were expressed as milligrams of gallic acid equivalent (GAE) per gram of extract.
4.5. Determination of Total Flavonoid Content (TFC) in C. chinense Stem Extract and NPs
The determination of the total flavonoid contents in the extract from C. chinense stem extract and NPs was carried out using the aluminum chloride colorimetric method [43]. In 96-well plates, solutions of quercetin (ranging from 3.9 to 1000 µg/mL), the extract (at concentrations of 312.5–20,000 µg/mL, 100 µL per well), and NPs (at concentrations of 52.08–13,333 µg/mL, 100 µL per well) were introduced. Subsequently, 30 µL of 5% NaNO2 was added to each well and incubated for 5 min. This was followed by the addition of 50 µL of 2% w/v aluminum chloride and a 6 min incubation, succeeded by a 10 min incubation with 50 µL of 1 N NaOH. The absorbance was measured at 510 nm using a UV–Vis spectrophotometer (Spectramax M3, Thermo Scientific, Waltham, MA, USA). Total flavonoid contents were determined based on standard curves for quercetin, and the results were expressed as milligrams of quercetin equivalent (QE) per gram of extract.
4.6. Antioxidant Activities of C. chinense Stem Extract and Nanoparticles
4.6.1. DPPH Free Radical Scavenging Activity Assay
The assessment of the free radical scavenging capacity of both C. chinense stem extract and NPs was conducted using the DPPH assay [44]. Ascorbyl glucoside served as the positive control, and it was prepared in a concentration ranging from 3.9 to 2000 μg/mL. In 96-well plates, aliquots of C. chinense stem extract (62.5–2000 µg/mL) and NPs (26.0–3333.33 µg/mL), each measuring 100 µL, were added. Subsequently, 100 µL of a DPPH solution at a concentration of 500 µM was introduced to the wells. The resultant mixture was incubated in darkness for a duration of 30 min, after which the absorbance was measured at 517 nm. The percentage of inhibition for both the standard and the extract was calculated at each concentration, and graphical representations were generated by plotting % inhibition against concentration [45]. The IC50 values were expressed as the concentration in μg/mL of the extract required to reduce the absorbance of DPPH by 50% in comparison to the negative control.
where A is the absorbance of the reaction with solvent control and B is the absorbance of the DPPH with the extract.
4.6.2. ABTS Free Radical Scavenging Activity Assay
The ABTS radical scavenging assay was performed to confirm the free radical scavenging activity of the extract [44]. The stock solution of ABTS radical was prepared by mixing 7 mM of ABTS solution and 2.45 mM of potassium persulfate at a 1:1 ratio and allowing it to react for 12 h at room temperature in the dark. The ascorbyl glucoside (3.9–2000 µg/mL), C. chinense stem extract (31.25–2000 µg/mL), and NPs (26.0–6666.5 µg/mL) (20 µL) were placed in the 96-well plates, followed by adding ABTS solution (180 µL). The plates were kept in the dark for 30 min. The absorbance was read at 734 nm. The %inhibition of the standard and the extract was calculated [18]. The IC50 values of the standard and the sample were calculated from the graph.
where A is the absorbance of the reaction with solvent control and B is the absorbance of the reaction with the extract.
4.6.3. Ferric-Reducing Antioxidant Power (FRAP) Assay
FRAP reagent was prepared by mixing 300 mM of acetate buffer (pH 3.6), 10 mM of 2,4,6-tripyridyl-s-triazine (TPTZ) solution in 40 mM of HCl, and 20 mM of FeCl3·6H2O solution at a 10:1:1 ratio [44]. Ascorbyl glucoside solution (3.9–2000 µg/mL) was used as the standard solution, and C. chinense stem extract solution (39–20,000 µg/mL) and NPs (26–13,333 µg/mL) were prepared. Samples (20 μL) were allowed to react with FRAP solution (180 μL) in 96-well plates at 37 °C for 30 min in the dark. The absorbance of ferrous tripyridyltriazine complex of the standard and the extract was read at 595 nm. The concentration of FRAP content in the extract was reported as μM Fe (II) equivalent. The FRAP content was calculated from the standard curve constructed using a ferrous sulfate solution (9.8–5000 μM).
In this study, three distinct antioxidant assays were employed, each with specific applications. The DPPH assay was utilized to evaluate the antioxidant activity of hydrophobic compounds present in both the C. chinense stem extract and NPs. In contrast, the ABTS assay is versatile and capable of measuring the antioxidant capacity of both hydrophilic and hydrophobic antioxidants. The FRAP assay, on the other hand, specifically measures the antioxidant activity of hydrophilic compounds in the extract and NPs. Given that the extraction solvent for the C. chinense stem was 95% ethanol, it facilitated the extraction of both hydrophobic and hydrophilic antioxidants.
The DPPH test is a widely employed method for assessing the antioxidant activity of plant extracts. This assay relies on the ability of antioxidants to donate electrons, thus neutralizing the DPPH radical. The ABTS test evaluates the antioxidants’ capacity to neutralize the stable radical cation ABTS•+. The presence of antioxidants leads to a decrease in the intensity of the blue-green chromophore. The FRAP test follows a Single Electron Transfer (SET) mechanism, measuring the reduction of ferric ions (Fe3+) ligand to the intensely blue ferrous complex (Fe2+) in acidic conditions. Antioxidants in the solution can either reduce Fe3+ to Fe2+, binding with ferricyanide to produce Prussian blue, or reduce ferricyanide to ferrocyanide, which binds free Fe3+ to form Prussian blue [46].
The correlations between total phenolic and total flavonoid contents and the antioxidant activities of C. chinense stem extract and the NPs were calculated using the Pearson correlation coefficient.
4.7. Cell Proliferation Assay Using a Sulforhodamine B (SRB) Assay
The anticancer activities of the C. chinense stem extract were tested for the first time in this study. To confirm the anticancer activity of the C. chinense stem extract, the cytotoxicity of the extract was tested in several cancer cell lines, including HeLa, A549, MCF-7, and SKOV-3 cells. The most sensitive cell line to this extract was further investigated for the anticancer activity mechanisms. The C. chinense stem extract was exposed to three distinct cancer cell lines: MCF-7, HeLa, and A549 cells. The cytotoxicity of NPs was investigated in the most sensitive cells, MCF-7 cells, and it was confirmed in A549 cells. After a 24, 48, and 72 h incubation period, the cytotoxicity of both the extract and the NPs (25, 50, 100, 250, 500, and 1000 µg/mL) was assessed via the SRB assay [19]. To fix the cells, ice-cold 10% trichloroacetic acid at 4 °C was added. Subsequently, the cells were stained with a 0.4% SRB dye for 30 min at room temperature. Removal of unbound dye was achieved through three washes with 1% acetic acid. Then, 200 µL of a 10 mm Tris base buffer was added to each well to dissolve the bound dye. The absorbance was measured using a spectrophotometer microplate reader at a wavelength of 540 nm. The percentage of cell viability was calculated by employing the following equation, with untreated cells serving as the control [19]:
where the A540 sample is the absorbance of treated cells and the A540 control is the absorbance of untreated cells. The IC50 value indicates the concentration of C. chinense stem extract and NPs that reduce cancer cell viability or induce cell death by 50%. The dose–response curve was created by plotting the percentage of cell viability against the log-transformed concentrations of the extract or NPs. The IC50 was calculated using GraphPad Prism 7.0.
C. chinense stem extract and NPs were treated with C2C12 normal cells, and the cell viability was measured using an SRB assay following the protocol above. The selectivity index was calculated according to the following formula: Selectivity index = (IC50 of a sample in SKOV-3 noncancerous cell)/(IC50 of a sample in cancerous cell lines).
4.8. Colony Formation Assay
The colony formation assay was used to assess the anchorage-dependent growth of MCF-7 cells. MCF-7 cells were seeded in 6-well plates at 2000 cells/well. Then, the cells were subjected to treatment with a concentration range of 25, 50, 100, 250, and 500 µg/mL of C. chinense stem extract and 50, 100, 250, 500, and 1000 µg/mL of NPs for 24 h. After a 15-day incubation period, the cells were stained with 0.5% crystal violet for 5 min at room temperature, followed by a thorough wash with deionized water and air-drying [18]. The colonies that had formed a minimum of 50 cells were observed and quantified using a light microscope at 40× magnification.
4.9. Annexin V/PI Apoptosis Detection Assay
Cell death mechanisms were examined using the Annexin V-FITC apoptosis detection kit (Sigma-Aldrich, St. Louis, MO, USA) by following the manufacturer’s instructions. MCF-7 cells were initially seeded in 6-well plates at a density of 2.5 × 105 cells/well and left to incubate for 24 h before being subjected to treatment with C. chinense stem extract (100, 250, and 500 μg/mL) and NPs (250, 500, and 1000 μg/mL) for an additional 24 h. Following treatment, both the treated and untreated (negative control) cells were trypsinized and subsequently washed three times with PBS. The cells were then re-suspended in a 1× apoptosis binding buffer. To label the cells, 5 μL of Annexin V-FITC and 10 μL of PI solution were added to each suspension and allowed to incubate for 15 min at room temperature in the absence of light [18]. Cell apoptosis was assessed using a flow cytometer (BD Biosciences, San Jose, CA, USA) within 60 min, employing BD Accuri C6 Plus software version 227.4.
4.10. Mitochondrial Membrane Potential (MMP) Assay
The effect of C. chinense stem extract and NPs on mitochondrial function was determined using the JC-1 staining method. For mitochondrial function, cancer cells (2.5 × 105 cells/well) were seeded into 6-well cultured plate for 24 h and treated with C. chinense stem extract (100, 250, and 500 µg/mL) and NPs (250, 500, and 1000 µg/mL) for 24 h. MCF-7 cells were harvested and washed, and the pellet cancer cells were collected. The cells were added to 100 µL of medium containing 5 µL of JC-1 assay reagents for 30 min at 37 °C in the dark; finally, 400 µL of DMEM medium was added. The mitochondrial membrane potential was determined through flow cytometric analysis (BD Biosciences, CA, USA) using BD Accuri C6 Plus software.
4.11. Cell Cycle Arrest Assay
Flow cytometry was used to investigate how C. chinense stem extract and NPs affect the cell cycle distribution of MCF-7 cells [18]. The cells, initially seeded at a density of 2.5 × 105 cells/mL, were cultured in 6-well plates for 24 h before exposure to C. chinense stem extract (100, 250, and 500 µg/mL) and NPs (250, 500, and 1000 µg/mL) for an additional 24 h incubation. Following trypsinization, the cells were washed with PBS buffer and then fixed with ice-cold 70% ethanol at −20 °C. Subsequently, the cells were treated with PI solution (BD Biosciences, CA, USA) for 30 min at 4 °C. Flow cytometry, with the aid of BD Accuri C6 Plus software, was employed to analyze the cell cycle phases, and the resulting fluorescent signals were presented as histograms. Gated cells were manually classified into their respective cell cycle stages.
4.12. Statistical Analysis
The data were expressed as mean ± S.D. Statistical analysis was conducted using one-way analysis of variance (ANOVA) and Tukey’s post hoc test. Additionally, a t-test was employed to assess the significance of the difference between the means of the two groups. A p-value < 0.05 was deemed statistically significant.
5. Conclusions
This study explored the cytotoxic impact of C. chinense stem extract on four human cancer cell lines, including MCF-7, A549, HeLa, and SKOV-3 cells. The MCF-7 cell line was mostly sensitive to the extract with the highest selectivity index. While nanoparticles derived from the extract exhibited reduced cytotoxicity against MCF-7 cells, they did not disrupt the extract’s anticancer mechanism. Investigations into the underlying mechanisms revealed that both the C. chinense stem extract and its nanoparticles significantly impeded the growth of MCF-7 cells by inducing apoptosis, inhibiting colony formation, reducing mitochondrial membrane potential, and causing cell cycle arrest through the regulation of the G0/G1 transition.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms25020978/s1.
Author Contributions
Conceptualization, C.C., S.O. and S.A.; Methodology, C.C., W.S., M.N.T. and S.A.; Validation, C.C. and S.N.; Formal analysis, C.C., W.S., M.N.T. and S.A.; Investigation, C.C., W.S., M.N.T. and S.A.; Resources, W.S., M.N.T. and S.O.; Data curation, M.N.T.; Writing—original draft, C.C., M.N.T. and S.N.; Writing—review & editing, W.S., M.N.T., S.N. and S.A.; Visualization, S.N.; Funding acquisition, S.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Chiang Mai University grant number RG01/2566 and The APC was funded by the Center of Excellence in Pharmaceutical Nanotechnology, Chiang Mai University.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article and Supplementary Material.
Acknowledgments
This research was supported by the Center of Excellence in Pharmaceutical Nanotechnology, Faculty of Pharmacy, Chiang Mai University, Thailand. The authors acknowledge Chiang Mai University and Srinakharinwirot University, Thailand.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| A549 | Human lung carcinoma |
| ABTS | 2,2′-Azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) |
| ANOVA | Analysis of variance |
| DPPH | 2,2-Diphenyl-1-picrylhydrazyl |
| EGCG | Epigallocatechin gallate |
| FRAP | Ferric-Reducing Antioxidant Power |
| GAE | Gallic acid equivalent |
| HeLa | Human epithelial cervical cancer |
| HPLC | High-performance liquid chromatography |
| JC-1 | 5,5,6,6-Tetrachloro-1,1,3,3-tetraethylbenzimidazolylcarbocyanine iodide |
| MCF-7 | Human breast adenocarcinoma |
| MMP | Mitochondrial membrane potential |
| NAPIs | Natural active pharmaceutical ingredients |
| NPs | Nanoparticles |
| PDI | Polydispersity index |
| QE | Quercetin equivalent |
| RP | Reversed Phase |
| SRB | Sulforhodamine B |
| TEM | Transmission electron microscopy |
| TFC | Total flavonoid content |
| TPC | Total phenolic content |
| ZTP | Ziyang green tea |
References
- Debela, D.T.; Muzazu, S.G.; Heraro, K.D.; Ndalama, M.T.; Mesele, B.W.; Haile, D.C.; Kitui, S.K.; Manyazewal, T. New approaches and procedures for cancer treatment: Current perspectives. SAGE Open Med. 2021, 9, 20503121211034366. [Google Scholar] [CrossRef]
- Arnold, M.; Morgan, E.; Rumgay, H.; Mafra, A.; Singh, D.; Laversanne, M.; Vignat, J.; Gralow, J.R.; Cardoso, F.; Siesling, S.; et al. Current and future burden of breast cancer: Global statistics for 2020 and 2040. Breast 2022, 66, 15–23. [Google Scholar] [CrossRef]
- Williams, A.; Moo, T.-A. The Impact of Socioeconomic Status and Social Determinants of Health on Disparities in Breast Cancer Incidence, Treatment, and Outcomes. Curr. Breast Cancer Rep. 2023, 15, 30–36. [Google Scholar] [CrossRef]
- Geyer, F.C.; Lopez-Garcia, M.A.; Lambros, M.B.; Reis-Filho, J.S. Genetic characterization of breast cancer and implications for clinical management. J. Cell. Mol. Med. 2009, 13, 4090–4103. [Google Scholar] [CrossRef]
- Elenbaas, B.; Spirio, L.; Koerner, F.; Fleming, M.D.; Zimonjic, D.B.; Donaher, J.L.; Popescu, N.C.; Hahn, W.C.; Weinberg, R.A. Human breast cancer cells generated by oncogenic transformation of primary mammary epithelial cells. Genes Dev. 2001, 15, 50–65. [Google Scholar] [CrossRef]
- Dhyani, P.; Quispe, C.; Sharma, E.; Bahukhandi, A.; Sati, P.; Attri, D.C.; Szopa, A.; Sharifi-Rad, J.; Docea, A.O.; Mardare, I.; et al. Anticancer potential of alkaloids: A key emphasis to colchicine, vinblastine, vincristine, vindesine, vinorelbine and vincamine. Cancer Cell Int. 2022, 22, 206. [Google Scholar] [CrossRef] [PubMed]
- Greenwell, M.; Rahman, P.K. Medicinal Plants: Their Use in Anticancer Treatment. Int. J. Pharm. Sci. Res. 2015, 6, 4103–4112. [Google Scholar] [CrossRef]
- Ganesan, K.; Du, B.; Chen, J. Effects and mechanisms of dietary bioactive compounds on breast cancer prevention. Pharmacol. Res. 2022, 178, 105974. [Google Scholar] [CrossRef] [PubMed]
- Mileo, A.M.; Miccadei, S. Polyphenols as Modulator of Oxidative Stress in Cancer Disease: New Therapeutic Strategies. Oxid. Med. Cell. Longev. 2016, 2016, 6475624. [Google Scholar] [CrossRef] [PubMed]
- Bhosale, P.B.; Ha, S.E.; Vetrivel, P.; Kim, H.H.; Kim, S.M.; Kim, G.S. Functions of polyphenols and its anticancer properties in biomedical research: A narrative review. Transl. Cancer Res. 2020, 9, 7619–7631. [Google Scholar] [CrossRef]
- Cháirez-Ramírez, M.H.; de la Cruz-López, K.G.; García-Carrancá, A. Polyphenols as Antitumor Agents Targeting Key Players in Cancer-Driving Signaling Pathways. Front. Pharmacol. 2021, 12, 710304. [Google Scholar] [CrossRef] [PubMed]
- Chojnacka, K.; Lewandowska, U. The Antiangiogenic Activity of Polyphenol-Rich Extracts and Its Implication on Cancer Chemoprevention. Food Rev. Int. 2020, 36, 77–103. [Google Scholar] [CrossRef]
- Bag, N.; Bag, A. Antimetastatic Properties of Tea Polyphenols. Nutr. Cancer 2020, 72, 365–376. [Google Scholar] [CrossRef]
- Azqueta, A.; Collins, A. Polyphenols and DNA Damage: A Mixed Blessing. Nutrients 2016, 8, 785. [Google Scholar] [CrossRef]
- Kar, P.; Goyal, A.; Das, A.; Sen, A. Antioxidant and pharmaceutical potential of Clerodendrum L.: An overview. Int. J. Green Pharm. 2014, 8, 210–216. [Google Scholar] [CrossRef]
- Qi, J.; Zhang, Y.; Liu, Q.; Liu, H.; Fan, Y.; Yue, J. Clerodenoids A—F: C-ring Aromatized and/or Rearranged Abietane Diterpenoids from Clerodendrum chinense var. simplex. Chin. J. Chem. 2021, 39, 1891–1897. [Google Scholar] [CrossRef]
- Barung, E.N.; Kalonio, D.E.; Banne, Y.; Kambuno, N.T. Anticancer Activities of Sesewanua Leaf Extracts (Clerodendrum fragrans (Vent.) Willd) Against A549 Lung Cancer Cell. Open Access Maced. J. Med. Sci. 2021, 9, 1226–1230. [Google Scholar] [CrossRef]
- Chittasupho, C.; Samee, W.; Tadtong, S.; Jittachai, W.; Managit, C.; Athikomkulchai, S. Cytotoxicity, Apoptosis Induction, Oxidative Stress, and Cell Cycle Arrest of Clerodendrum chinense Flower Extract Nanoparticles in HeLa Cells. Nat. Life Sci. Commun. 2023, 22, e2023057. [Google Scholar] [CrossRef]
- Chittasupho, C.; Athikomkulchai, S.; Samee, W.; Na Takuathung, M.; Yooin, W.; Sawangrat, K.; Saenjum, C. Phenylethanoid Glycoside-Enriched Extract Prepared from Clerodendrum chinense Leaf Inhibits A549 Lung Cancer Cell Migration and Apoptosis Induction through Enhancing ROS Production. Antioxidants 2023, 12, 461. [Google Scholar] [CrossRef]
- Gavas, S.; Quazi, S.; Karpiński, T.M. Nanoparticles for Cancer Therapy: Current Progress and Challenges. Nanoscale Res. Lett. 2021, 16, 173. [Google Scholar] [CrossRef]
- Altemimi, A.; Lakhssassi, N.; Baharlouei, A.; Watson, D.G.; Lightfoot, D.A. Phytochemicals: Extraction, Isolation, and Identification of Bioactive Compounds from Plant Extracts. Plants 2017, 6, 42. [Google Scholar] [CrossRef]
- Payne, A.C.; Mazzer, A.; Clarkson, G.J.; Taylor, G. Antioxidant assays—Consistent findings from FRAP and ORAC reveal a negative impact of organic cultivation on antioxidant potential in spinach but not watercress or rocket leaves. Food Sci. Nutr. 2013, 1, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Funes, L.; Fernández-Arroyo, S.; Laporta, O.; Pons, A.; Roche, E.; Segura-Carretero, A.; Fernández-Gutiérrez, A.; Micol, V. Correlation between plasma antioxidant capacity and verbascoside levels in rats after oral administration of lemon verbena extract. Food Chem. 2009, 117, 589–598. [Google Scholar] [CrossRef]
- Tang, Y.; Zhao, D.Y.; Elliott, S.; Zhao, W.; Curiel, T.J.; Beckman, B.S.; Burow, M.E. Epigallocatechin-3 gallate induces growth inhibition and apoptosis in human breast cancer cells through survivin suppression. Int. J. Oncol. 2007, 31, 705–711. [Google Scholar] [CrossRef]
- Patel, V.A.; Longacre, A.; Hsiao, K.; Fan, H.; Meng, F.; Mitchell, J.E.; Rauch, J.; Ucker, D.S.; Levine, J.S. Apoptotic cells, at all stages of the death process, trigger characteristic signaling events that are divergent from and dominant over those triggered by necrotic cells: Implications for the delayed clearance model of autoimmunity. J. Biol. Chem. 2006, 281, 4663–4670. [Google Scholar] [CrossRef]
- Khan, H.; Zubair, H.; Ullah, M.F.; Ahmad, A.; Hadi, S. A Prooxidant Mechanism for the Anticancer and Chemopreventive Properties of Plant Polyphenols. Curr. Drug Targets 2012, 13, 1738–1749. [Google Scholar] [CrossRef] [PubMed]
- Rajashekar, C. Dual Role of Plant Phenolic Compounds as Antioxidants and Prooxidants. Am. J. Plant Sci. 2023, 14, 15–28. [Google Scholar] [CrossRef]
- Şenol, H.; Tulay, P.; Ergören, M.; Hanoğlu, A.; Çalış, İ.; Mocan, G. Cytotoxic Effects of Verbascoside on MCF-7 and MDA-MB-231. Turk. J. Pharm. Sci. 2021, 18, 637–644. [Google Scholar] [CrossRef]
- Delazar, A.; Asnaashari, S.; Nikkhah, E.; Asgharian, P. Phytochemical analysis and antiproliferative activity of the aerial parts of Scrophularia subaphylla. Res. Pharm. Sci. 2019, 14, 263–272. [Google Scholar] [CrossRef]
- Zhou, L.; Feng, Y.; Jin, Y.; Liu, X.; Sui, H.; Chai, N.; Chen, X.; Liu, N.; Ji, Q.; Wang, Y.; et al. Verbascoside promotes apoptosis by regulating HIPK2-p53 signaling in human colorectal cancer. BMC Cancer 2014, 14, 747. [Google Scholar] [CrossRef]
- Chen, R.C.; Su, J.H.; Yang, S.M.; Li, J.; Wang, T.J.; Zhou, H. Effect of isoverbascoside, a phenylpropanoid glycoside antioxidant, on proliferation and differentiation of human gastric cancer cell. Acta Pharmacol. Sin. 2002, 23, 997–1001. [Google Scholar]
- Zhang, Z.; Feng, Y.; Li, Z.Y.; Cao, X.Z. Antiproliferative and apoptotic activity of glycyrrhizinic acid in MCF-7 human breast cancer cells and evaluation of its effect on cell cycle, cell migration and m-TOR/PI3K/Akt signalling pathway. Arch. Med. Sci. 2019, 15, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Yu, J.L.; Sun, D.Q.; Kong, F.; Qu, X.J.; Zou, W.; Wu, J.; Wang, R.M. Curcumin induces apoptosis in SGC-7901 gastric adenocarcinoma cells via regulation of mitochondrial signaling pathways. Asian Pac. J. Cancer Prev. 2014, 15, 3987–3992. [Google Scholar] [CrossRef]
- Lin, J.W.; Chen, J.T.; Hong, C.Y.; Lin, Y.L.; Wang, K.T.; Yao, C.J.; Lai, G.M.; Chen, R.M. Honokiol traverses the blood-brain barrier and induces apoptosis of neuroblastoma cells via an intrinsic bax-mitochondrion-cytochrome c-caspase protease pathway. Neuro Oncol. 2012, 14, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Q.; Lu, J.L.; Liang, Y.R.; Li, Q.S. Suppressive Effects of EGCG on Cervical Cancer. Molecules 2018, 23, 2334. [Google Scholar] [CrossRef]
- Qanungo, S.; Das, M.; Haldar, S.; Basu, A. Epigallocatechin-3-gallate induces mitochondrial membrane depolarization and caspase-dependent apoptosis in pancreatic cancer cells. Carcinogenesis 2005, 26, 958–967. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Qu, H.; Wu, Y.; Wang, Z.; Wang, L. Study on antitumor, antioxidant and immunoregulatory activities of the purified polyphenols from pinecone of Pinus koraiensis on tumor-bearing S180 mice in vivo. Int. J. Biol. Macromol. 2017, 94, 735–744. [Google Scholar] [CrossRef]
- Li, W.; He, N.; Tian, L.; Shi, X.; Yang, X. Inhibitory effects of polyphenol-enriched extract from Ziyang tea against human breast cancer MCF-7 cells through reactive oxygen species-dependent mitochondria molecular mechanism. J. Food Drug Anal. 2016, 24, 527–538. [Google Scholar] [CrossRef]
- Zhao, Q.; Huo, X.C.; Sun, F.D.; Dong, R.Q. Polyphenol-rich extract of Salvia chinensis exhibits anticancer activity in different cancer cell lines, and induces cell cycle arrest at the G0/G1-phase, apoptosis and loss of mitochondrial membrane potential in pancreatic cancer cells. Mol. Med. Rep. 2015, 12, 4843–4850. [Google Scholar] [CrossRef]
- Ou, Y.; Zhang, Y.; Zhu, X.; Zhu, D. Verbascoside induces the cell cycle arrest and apoptosis of breast cancer via suppression of the PI3K/AKT signaling pathway. Res. Sq. 2022. preprint. [Google Scholar] [CrossRef]
- Sciandra, F.; Bottoni, P.; De Leo, M.; Braca, A.; Brancaccio, A.; Bozzi, M. Verbascoside Elicits Its Beneficial Effects by Enhancing Mitochondrial Spare Respiratory Capacity and the Nrf2/HO-1 Mediated Antioxidant System in a Murine Skeletal Muscle Cell Line. Int. J. Mol. Sci. 2023, 24, 15276. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, L.; Yang, G.; Shi, H.; Jiang, C.; Liu, W.; Zhang, Y. Study on the determination of polyphenols in tobacco by HPLC coupled with ESI-MS after solid-phase extraction. J. Chromatogr. Sci. 2003, 41, 36–40. [Google Scholar] [CrossRef]
- Tunit, P.; Thammarat, P.; Okonogi, S.; Chittasupho, C. Hydrogel Containing Borassus flabellifer L. Male Flower Extract for Antioxidant, Antimicrobial, and Anti-Inflammatory Activity. Gels 2022, 8, 126. [Google Scholar] [CrossRef]
- Chittasupho, C.; Chaobankrang, K.; Sarawungkad, A.; Samee, W.; Singh, S.; Hemsuwimon, K.; Okonogi, S.; Kheawfu, K.; Kiattisin, K.; Chaiyana, W. Antioxidant, Anti-Inflammatory and Attenuating Intracellular Reactive Oxygen Species Activities of Nicotiana tabacum var. Virginia Leaf Extract Phytosomes and Shape Memory Gel Formulation. Gels 2023, 9, 78. [Google Scholar] [CrossRef] [PubMed]
- Athikomkulchai, S.; Tunit, P.; Tadtong, S.; Jantrawut, P.; Sommano, S.R.; Chittasupho, C. Moringa oleifera Seed Oil For-mulation Physical Stability and Chemical Constituents for Enhancing Skin Hydration and Antioxidant Activity. Cosmetics 2021, 8, 2. [Google Scholar] [CrossRef]
- Munteanu, I.G.; Apetrei, C. Analytical Methods Used in Determining Antioxidant Activity: A Review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).