
First Evidence of Mineralocorticoid Receptor Gene and Protein Expression in Rat and Human Thyroid Tissues and Cell Cultures

 ,
,  , , , , , , , , , and
, , , , , , , , , and 
Abstract
1. Introduction
2. Results
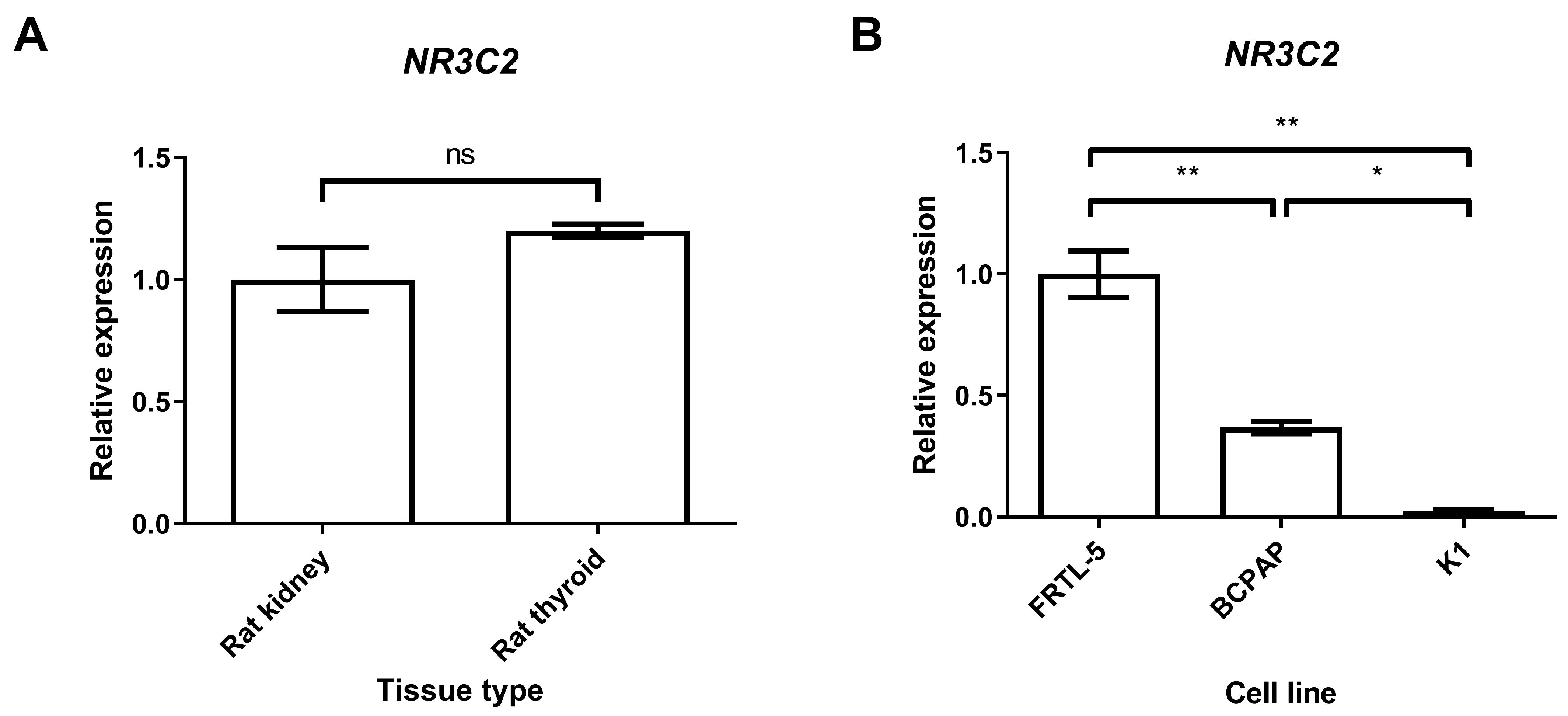
2.1. Gene Expression of Mineralocorticoid Receptor in Thyroid Tissues and Cell Lines
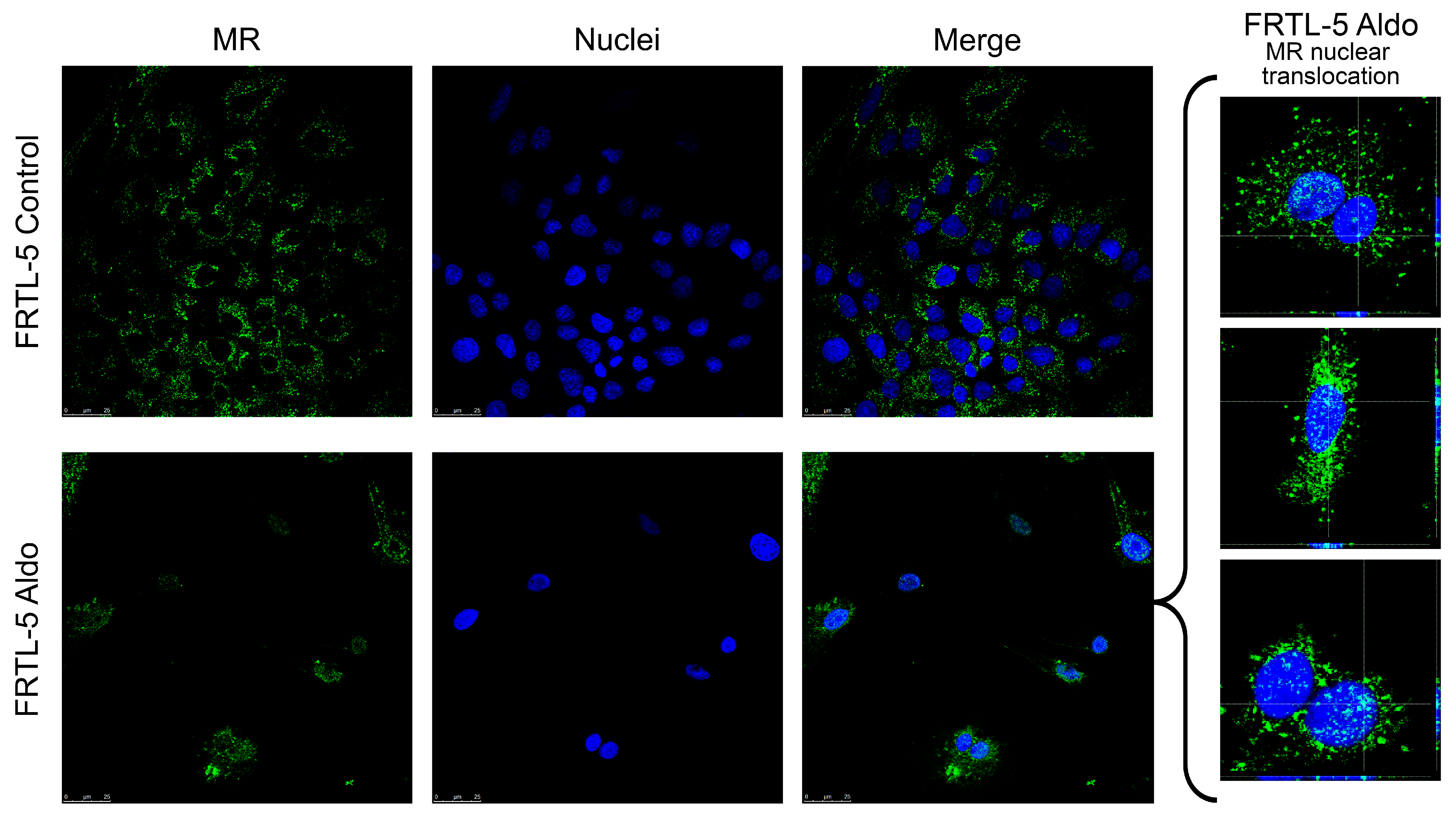
2.2. Mineralocorticoid Receptor Protein Expression, Localisation, and Activation in FRTL-5
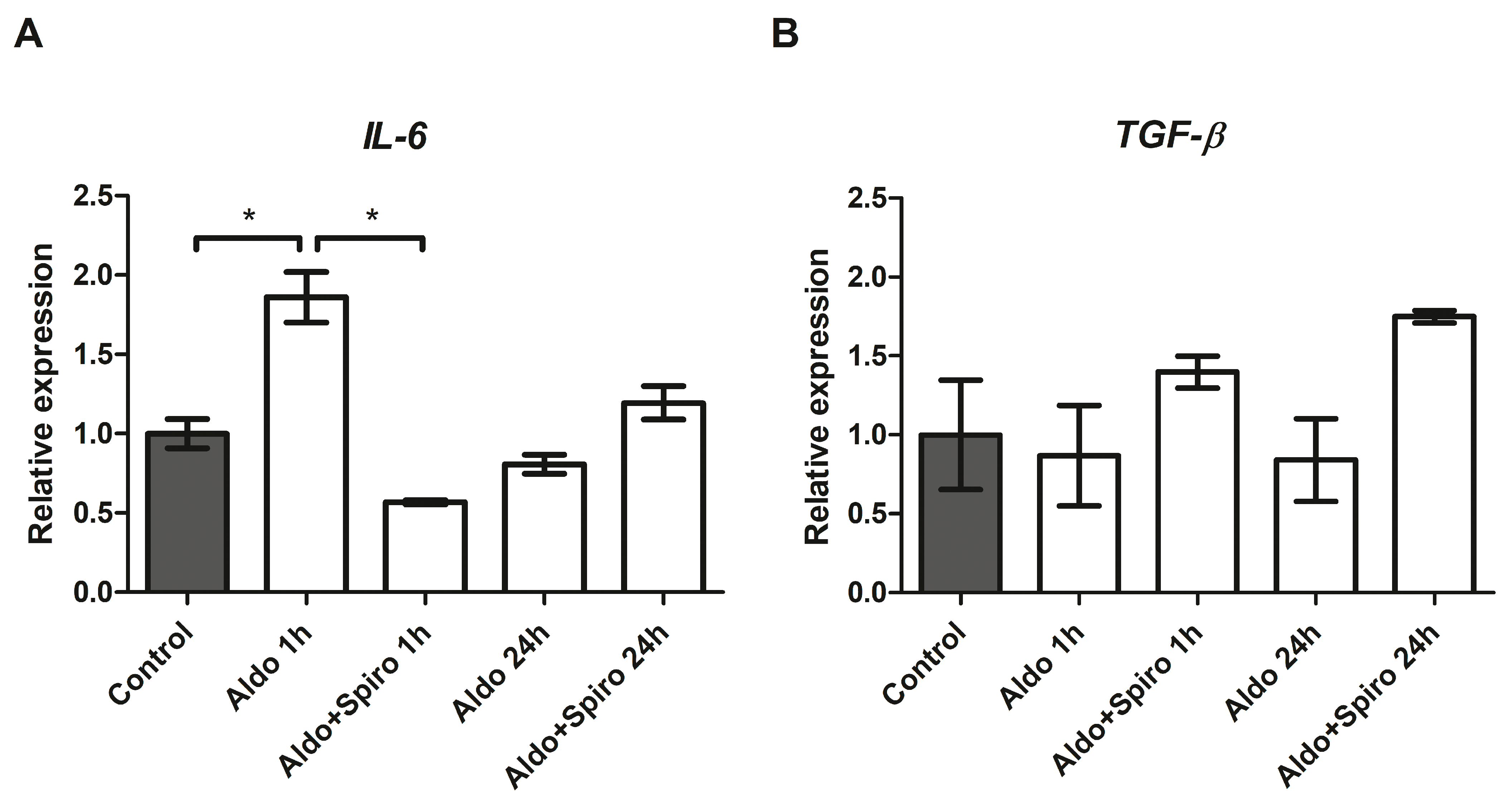
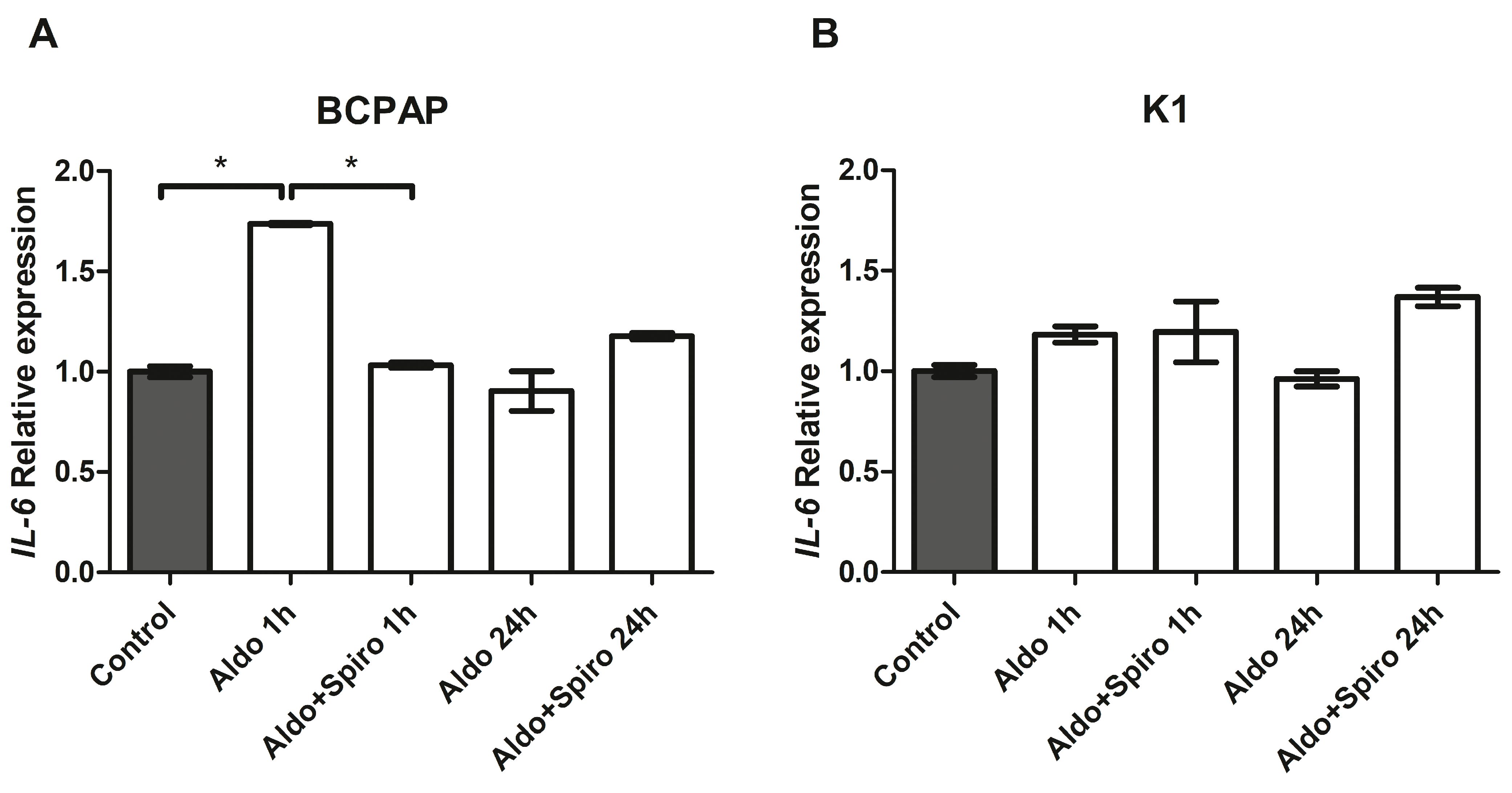
2.3. Effects of Aldosterone on Target Genes Involved in Inflammation in the FRTL-5 Cell Line
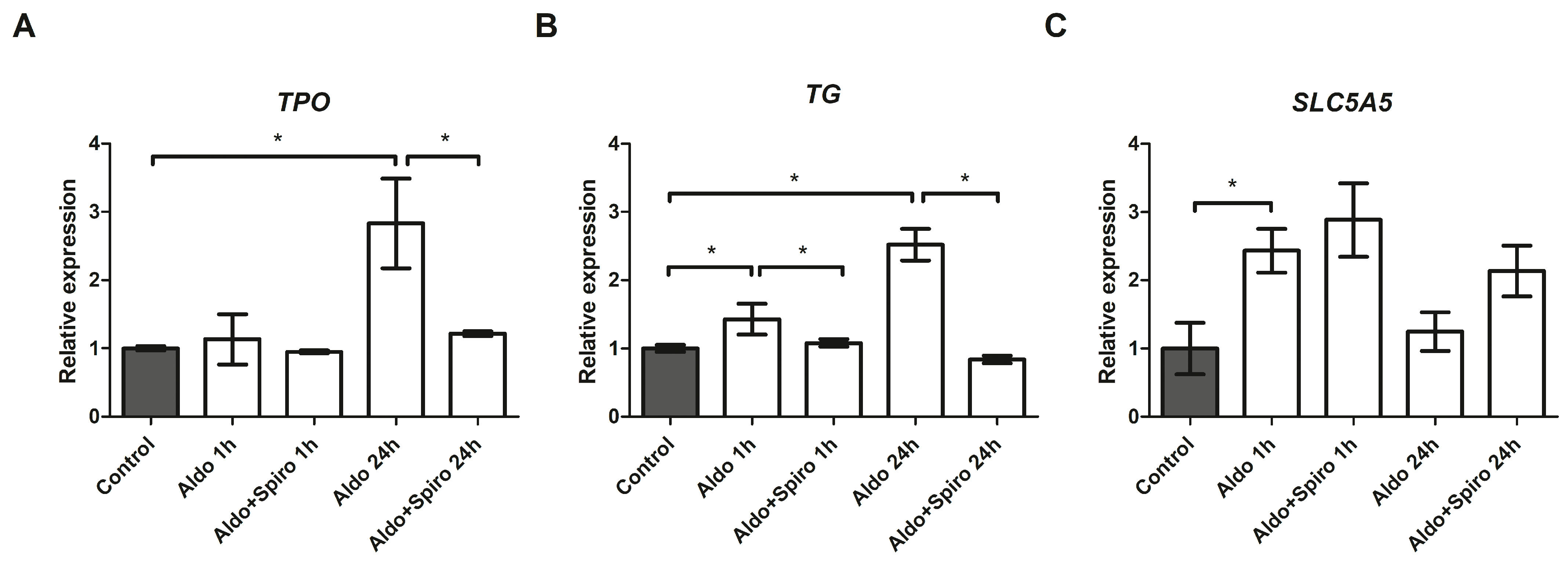
2.4. Effects of Aldosterone on Thyroid-Specific Genes in the FRTL-5 Cell Line
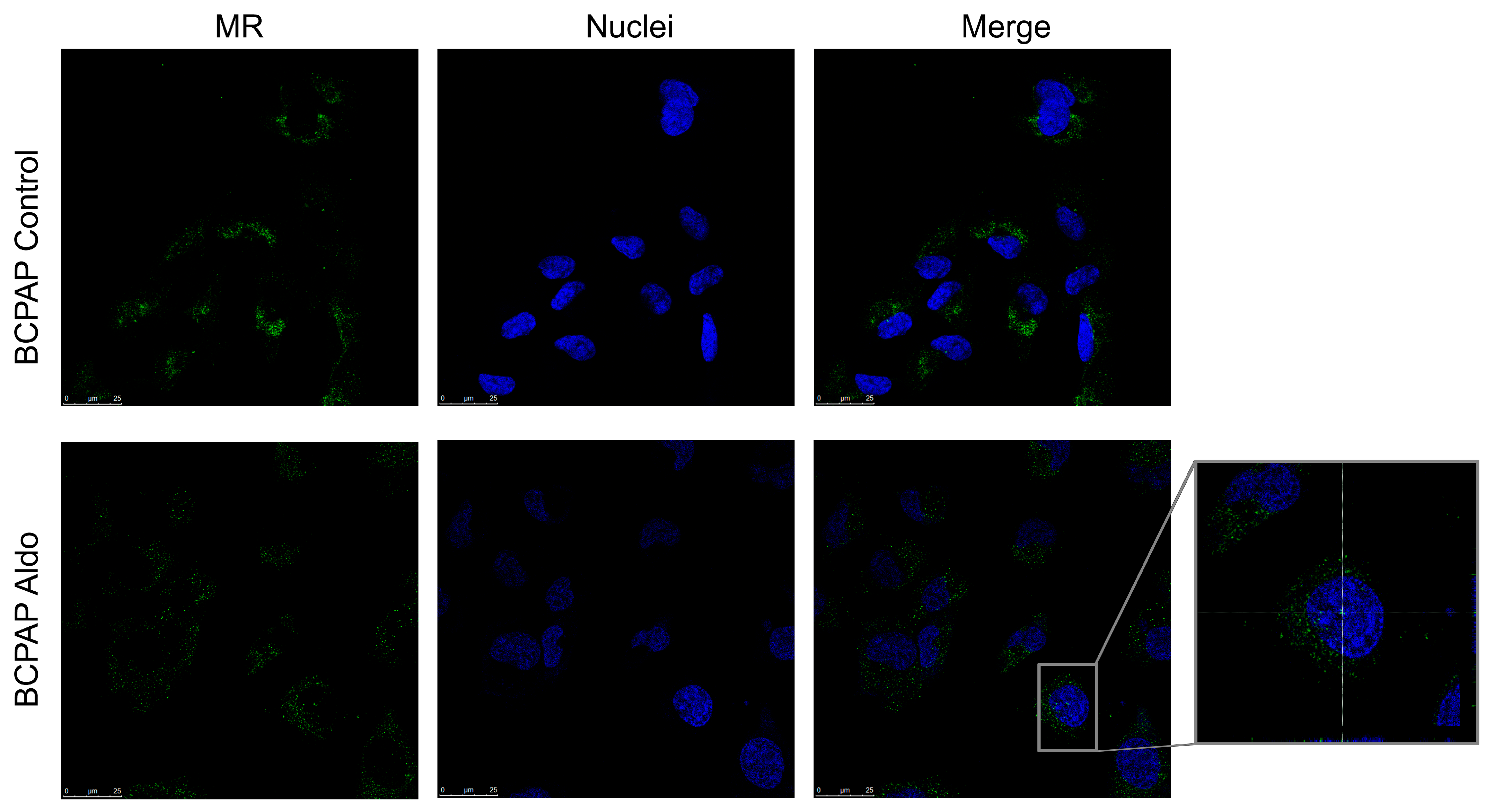
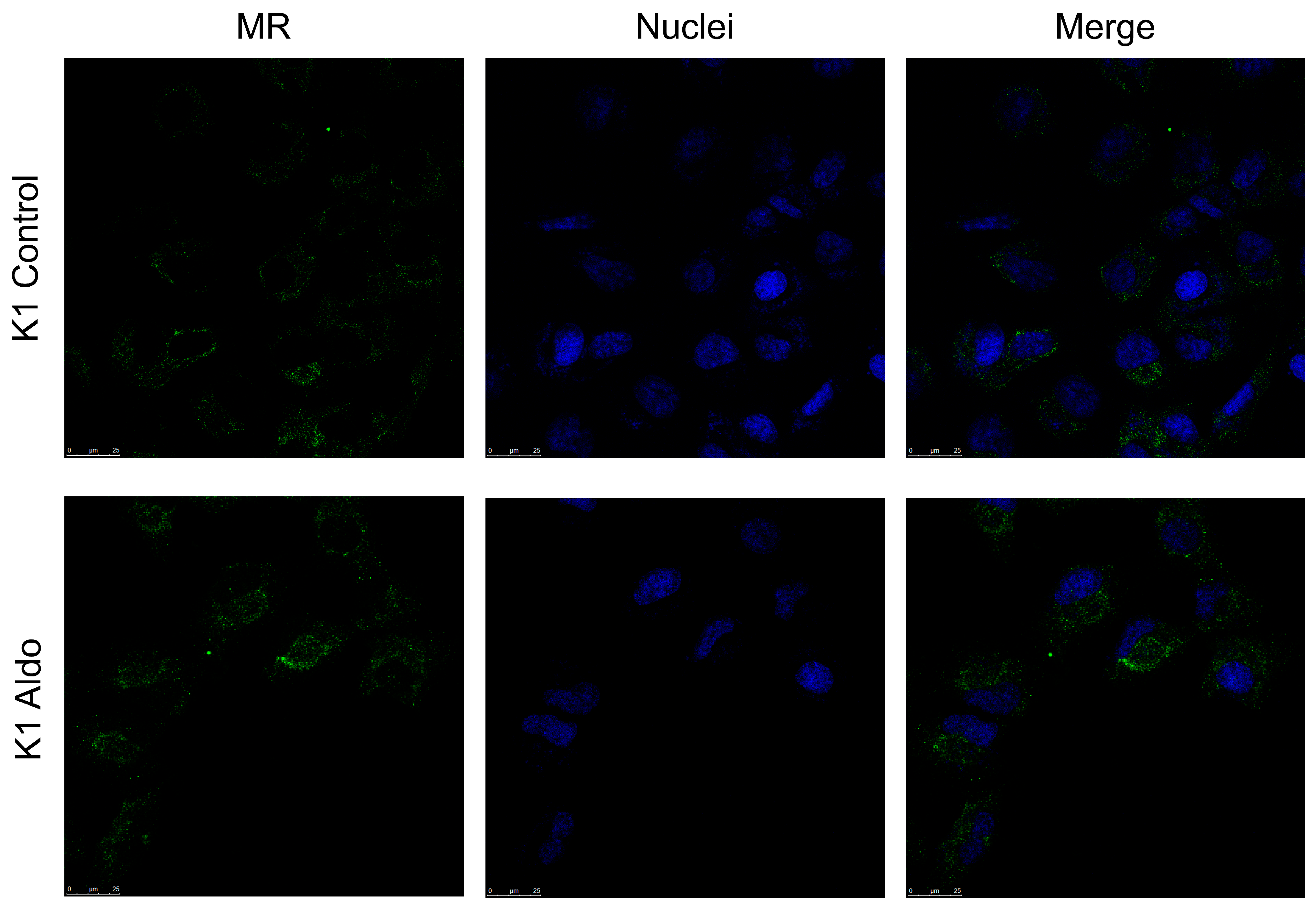
2.5. Mineralocorticoid Receptor Protein Expression, Localisation, and Activation in PTC Cell Lines
2.6. Effects of Aldosterone in PTC Cell Lines
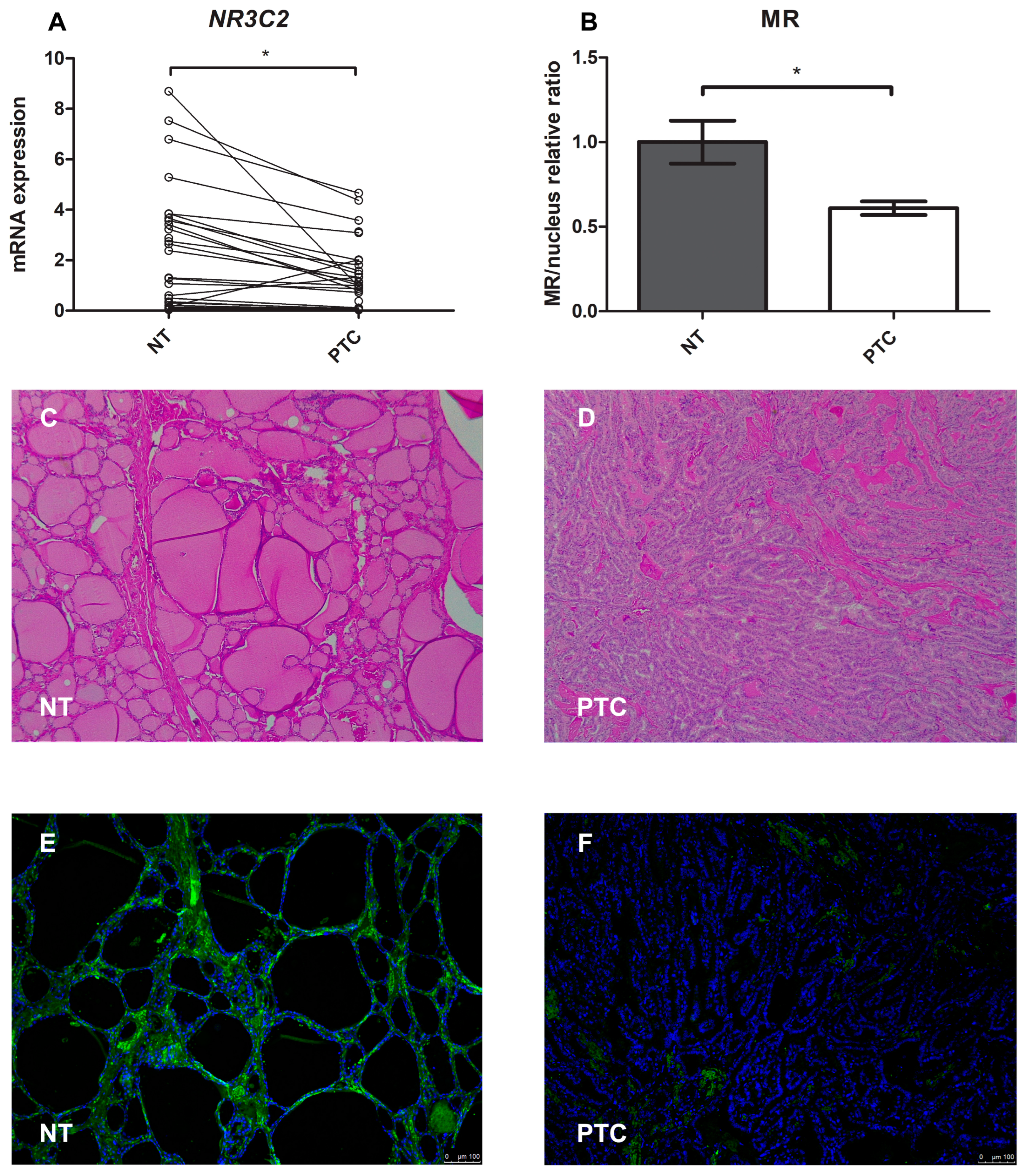
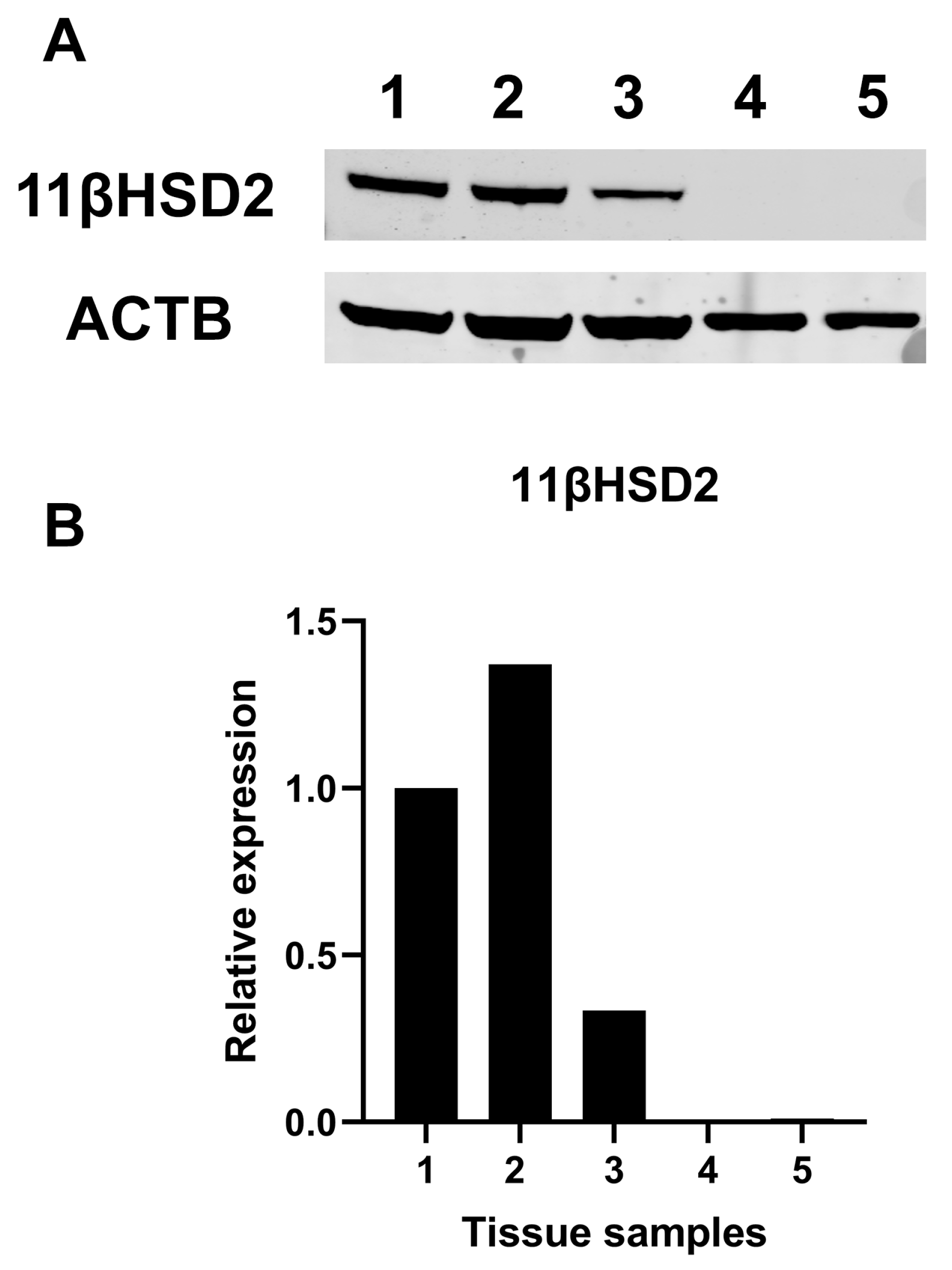
2.7. Mineralocorticoid Receptor Gene and Protein Expression in Human PTC
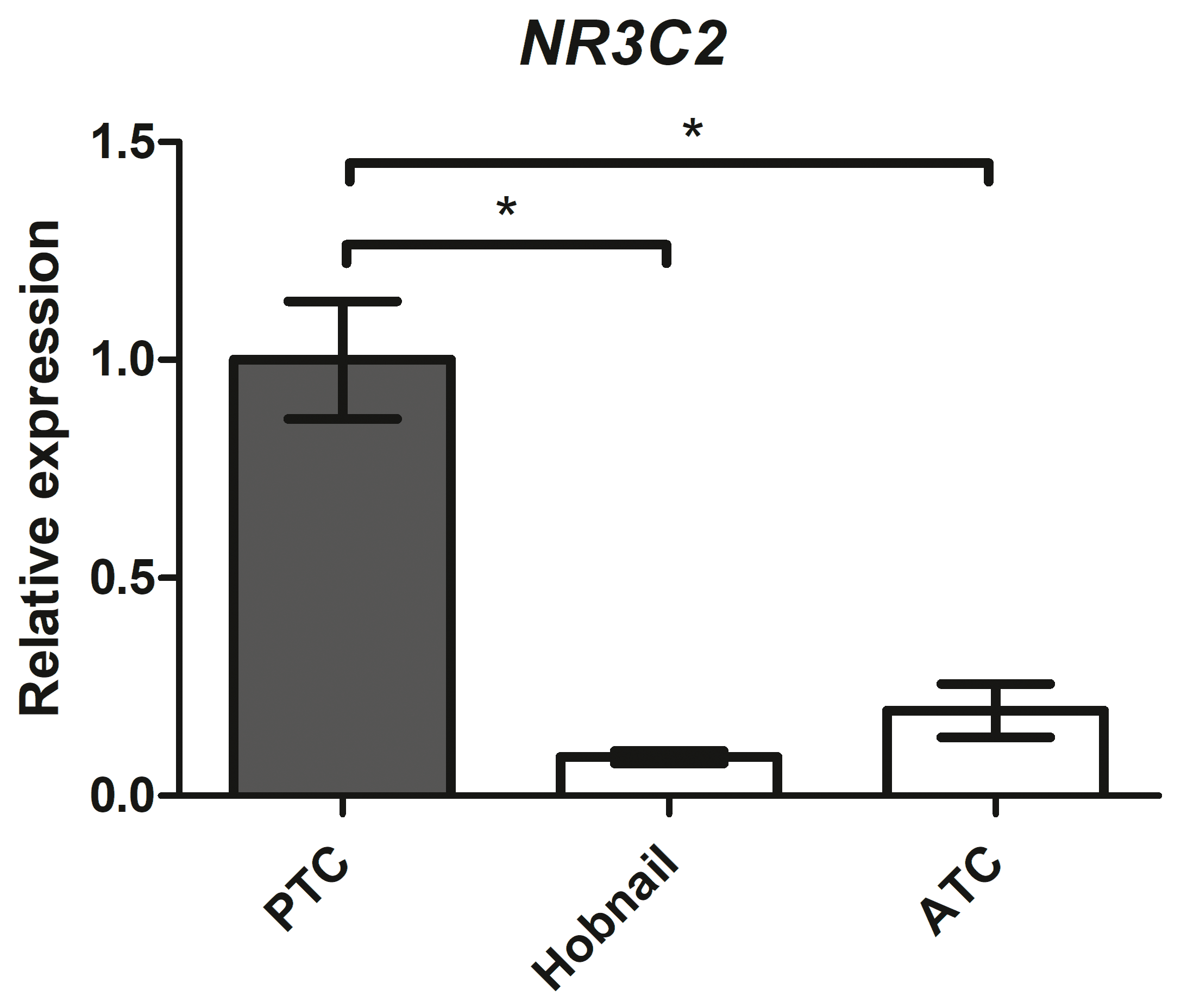
2.8. Mineralocorticoid Receptor Gene Expression in Aggressive Thyroid Cancers
3. Discussion
4. Materials and Method
4.1. Rat and Human Tissues
4.2. Cell Cultures and Maintenance
4.3. Drugs
4.4. RNA Extraction, cDNA Synthesis, and qRT-PCR Analysis
4.5. Immunofluorescence and Confocal Microscopy
4.6. Western Blot Analysis of Human Tissues
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Balsamo, A.; Cicognani, A.; Gennari, M.; Sippell, W.G.; Menabo, S.; Baronio, F.; Riepe, F.G. Functional characterization of naturally occurring NR3C2 gene mutations in Italian patients suffering from pseudohypoaldosteronism type 1. Eur. J. Endocrinol. 2007, 156, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Booth, R.E.; Johnson, J.P.; Stockand, J.D. Aldosterone. Adv. Physiol. Educ. 2002, 26, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.D.; Sherman, T.G.; Goldman, D.J.; Watson, S.J. Molecular cloning of a mineralocorticoid (type I) receptor complementary DNA from rat hippocampus. Mol. Endocrinol. 1989, 3, 1877–1885. [Google Scholar] [CrossRef] [PubMed]
- Pearce, P.; Funder, J.W. High affinity aldosterone binding sites (type I receptors) in rat heart. Clin. Exp. Pharmacol. Physiol. 1987, 14, 859–866. [Google Scholar] [CrossRef]
- Funder, J.W.; Pearce, P.T.; Smith, R.; Smith, A.I. Mineralocorticoid action: Target tissue specificity is enzyme, not receptor, mediated. Science 1988, 242, 583–585. [Google Scholar] [CrossRef] [PubMed]
- Caprio, M.; Newfell, B.G.; la Sala, A.; Baur, W.; Fabbri, A.; Rosano, G.; Mendelsohn, M.E.; Jaffe, I.Z. Functional mineralocorticoid receptors in human vascular endothelial cells regulate intercellular adhesion molecule-1 expression and promote leukocyte adhesion. Circ. Res. 2008, 102, 1359–1367. [Google Scholar] [CrossRef] [PubMed]
- Armanini, D.; Endres, S.; Kuhnle, U.; Weber, P.C. Parallel determination of mineralocorticoid and glucocorticoid receptors in T- and B-lymphocytes of human spleen. Acta Endocrinol. 1988, 118, 479–482. [Google Scholar] [CrossRef]
- Penfornis, P.; Viengchareun, S.; Le Menuet, D.; Cluzeaud, F.; Zennaro, M.C.; Lombès, M. The mineralocorticoid receptor mediates aldosterone-induced differentiation of T37i cells into brown adipocytes. Am. J. Physiol. Endocrinol. Metab. 2000, 279, E386–E394. [Google Scholar] [CrossRef]
- Caprio, M.; Feve, B.; Claes, A.; Viengchareun, S.; Lombes, M.; Zennaro, M.C. Pivotal role of the mineralocorticoid receptor in corticosteroid-induced adipogenesis. FASEB J. 2007, 21, 2185–2194. [Google Scholar] [CrossRef]
- Bordin, L.; Dona, G.; Sabbadin, C.; Ragazzi, E.; Andrisani, A.; Ambrosini, G.; Brunati, A.M.; Clari, G.; Armanini, D. Human red blood cells alterations in primary aldosteronism. J. Clin. Endocrinol. Metab. 2013, 98, 2494–2501. [Google Scholar] [CrossRef][Green Version]
- McCurley, A.; McGraw, A.; Pruthi, D.; Jaffe, I.Z. Smooth muscle cell mineralocorticoid receptors: Role in vascular function and contribution to cardiovascular disease. Pflug. Arch. 2013, 465, 1661–1670. [Google Scholar] [CrossRef]
- Calo, L.A.; Zaghetto, F.; Pagnin, E.; Davis, P.A.; De Mozzi, P.; Sartorato, P.; Martire, G.; Fiore, C.; Armanini, D. Effect of aldosterone and glycyrrhetinic acid on the protein expression of PAI-1 and p22(phox) in human mononuclear leukocytes. J. Clin. Endocrinol. Metab. 2004, 89, 1973–1976. [Google Scholar] [CrossRef] [PubMed]
- Bene, N.C.; Alcaide, P.; Wortis, H.H.; Jaffe, I.Z. Mineralocorticoid receptors in immune cells: Emerging role in cardiovascular disease. Steroids 2014, 91, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Carvajal, C.A.; Herrada, A.A.; Castillo, C.R.; Contreras, F.J.; Stehr, C.B.; Mosso, L.M.; Kalergis, A.M.; Fardella, C.E. Primary aldosteronism can alter peripheral levels of transforming growth factor beta and tumor necrosis factor alpha. J. Endocrinol. Invest. 2009, 32, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Herrada, A.A.; Contreras, F.J.; Marini, N.P.; Amador, C.A.; González, P.A.; Cortés, C.M.; Riedel, C.A.; Carvajal, C.A.; Figueroa, F.; Michea, L.F.; et al. Aldosterone promotes autoimmune damage by enhancing Th17-mediated immunity. J. Immunol. 2010, 184, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, M.; Schott, M. Hashimoto’s thyroiditis and papillary thyroid cancer: Are they immunologically linked? Trends. Endocrinol. Metab. 2014, 25, 656–664. [Google Scholar] [CrossRef]
- Kuwabara, T.; Ishikawa, F.; Kondo, M.; Kakiuchi, T. The Role of IL-17 and Related Cytokines in Inflammatory Autoimmune Diseases. Mediat. Inflamm. 2017, 2017, 3908061. [Google Scholar] [CrossRef]
- Sabbadin, C.; Mian, C.; Nacamulli, D.; Dona, G.; Presotto, F.; Betterle, C.; Boscaro, M.; Bordin, L.; Armanini, D. Association of primary aldosteronism with chronic thyroiditis. Endocrine 2017, 55, 303–306. [Google Scholar] [CrossRef]
- Armanini, D.; Nacamulli, D.; Scaroni, C.; Lumachi, F.; Selice, R.; Fiore, C.; Favia, G.; Mantero, F. High prevalence of thyroid ultrasonographic abnormalities in primary aldosteronism. Endocrine 2003, 22, 155–160. [Google Scholar] [CrossRef]
- Krysiak, R.; Okopien, B. Coexistence of primary aldosteronism and Hashimoto’s thyroiditis. Rheumatol. Int. 2012, 32, 2561–2563. [Google Scholar] [CrossRef][Green Version]
- Jaisser, F.; Farman, N. Emerging Roles of the Mineralocorticoid Receptor in Pathology: Toward New Paradigms in Clinical Pharmacology. Pharmacol. Rev. 2016, 68, 49–75. [Google Scholar] [CrossRef] [PubMed]
- Barrera-Chimal, J.; Girerd, S.; Jaisser, F. Mineralocorticoid receptor antagonists and kidney diseases: Pathophysiological basis. Kidney Int. 2019, 96, 302–319. [Google Scholar] [CrossRef] [PubMed]
- Kolkhof, P.; Barfacker, L. 30 Years of the Mineralocorticoid Receptor: Mineralocorticoid receptor antagonists: 60 years of research and development. J. Endocrinol. 2017, 234, T125–T140. [Google Scholar] [CrossRef]
- Walker, B.R.; Yau, J.L.; Brett, L.P.; Seckl, J.R.; Monder, C.; Williams, B.C.; Edwards, C.R. 11 beta-hydroxysteroid dehydrogenase in vascular smooth muscle and heart: Implications for cardiovascular responses to glucocorticoids. Endocrinology 1991, 129, 3305–3312. [Google Scholar] [CrossRef] [PubMed]
- Whorwood, C.B.; Franklyn, J.A.; Sheppard, M.C.; Stewart, P.M. Tissue localization of 11 beta-hydroxysteroid dehydrogenase and its relationship to the glucocorticoid receptor. J. Steroid Biochem. Mol. Biol. 1992, 41, 21–28. [Google Scholar] [CrossRef]
- Gray, G.A.; White, C.I.; Castellan, R.F.; McSweeney, S.J.; Chapman, K.E. Getting to the heart of intracellular glucocorticoid regeneration: 11beta-HSD1 in the myocardium. J. Mol. Endocrinol. 2017, 58, R1–R13. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pitt, B.; Remme, W.; Zannad, F.; Neaton, J.; Martinez, F.; Roniker, B.; Bittman, R.; Hurley, S.; Kleiman, J.; Gatlin, M. Eplerenone, a selective aldosterone blocker, in patients with left ventricular dysfunction after myocardial infarction. N. Engl. J. Med. 2003, 348, 1309–1321. [Google Scholar] [CrossRef]
- Patil, S.; Rojulpote, C.; Amanullah, A. Primary Aldosteronism and Ischemic Heart Disease. Front. Cardiovasc. Med. 2022, 9, 882330. [Google Scholar] [CrossRef]
- Affinito, O.; Orlandella, F.M.; Luciano, N.; Salvatore, M.; Salvatore, G.; Franzese, M. Evolution of intra-tumoral heterogeneity across different pathological stages in papillary thyroid carcinoma. Cancer Cell Int. 2022, 22, 263. [Google Scholar] [CrossRef]
- Saiselet, M.; Floor, S.; Tarabichi, M.; Dom, G.; Hebrant, A.; van Staveren, W.C.; Maenhaut, C. Thyroid cancer cell lines: An overview. Front. Endocrinol. 2012, 3, 133. [Google Scholar] [CrossRef]
- Barrera-Chimal, J.; Bonnard, B.; Jaisser, F. Roles of Mineralocorticoid Receptors in Cardiovascular and Cardiorenal Diseases. Annu. Rev. Physiol. 2022, 84, 585–610. [Google Scholar] [CrossRef] [PubMed]
- Turchi, F.; Ronconi, V.; di Tizio, V.; Boscaro, M.; Giacchetti, G. Blood pressure, thyroid-stimulating hormone, and thyroid disease prevalence in primary aldosteronism and essential hypertension. Am. J. Hypertens 2011, 24, 1274–1279. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, S.B.; Limbutara, K.; Fenton, R.A.; Pisitkun, T.; Christensen, B.M. RNA sequencing of kidney distal tubule cells reveals multiple mediators of chronic aldosterone action. Physiol. Genom. 2018, 50, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Verlander, J.W.; Hassell, K.A.; Royaux, I.E.; Glapion, D.M.; Wang, M.E.; Everett, L.A.; Green, E.D.; Wall, S.M. Deoxycorticosterone upregulates PDS (Slc26a4) in mouse kidney: Role of pendrin in mineralocorticoid-induced hypertension. Hypertension 2003, 42, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, M.; Inada, M.; Kurata, S.; Oishi, M.; Naito, K.; Ishii, H.; Tanaka, K.; Mashio, Y.; Imura, H. Primary aldosteronism associated with a primary double cancer of the thyroid and rectum: Report of a case. Endocrinol. Jpn. 1981, 28, 735–739. [Google Scholar] [CrossRef] [PubMed]
- Wanta, S.M.; Basina, M.; Chang, S.D.; Chang, D.T.; Ford, J.M.; Greco, R.; Kingham, K.; Merritt, R.E.; Kunz, P.L. A rare case of an aldosterone secreting metastatic adrenocortical carcinoma and papillary thyroid carcinoma in a 31-year-old male. Rare Tumors 2011, 3, e45. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Ishimori, M.; Yamakita, N. Association of Papillary Thyroid Carcinoma with Primary Aldosteronism. Intern. Med. 2019, 58, 497–504. [Google Scholar] [CrossRef]
- Yamada, Y.; Haga, H.; Yamada, Y. Concise review: Dedifferentiation meets cancer development: Proof of concept for epigenetic cancer. Stem. Cells Transl. Med. 2014, 3, 1182–1187. [Google Scholar] [CrossRef]
- Censi, S.; Bertazza, L.; Piva, I.; Manso, J.; Benna, C.; Iacobone, M.; Mondin, A.; Plebani, M.; Faggian, D.; Galuppini, F.; et al. Serum miR-375 for Diagnostic and Prognostic Purposes in Medullary Thyroid Carcinoma. Front. Endocrinol. 2021, 12, 647369. [Google Scholar] [CrossRef]
- Chai, W.; Garrelds, I.M.; de Vries, R.; Batenburg, W.W.; van Kats, J.P.; Danser, A.H. Nongenomic effects of aldosterone in the human heart: Interaction with angiotensin II. Hypertension 2005, 46, 701–706. [Google Scholar] [CrossRef]
- Wang, B.H.; Bertucci, M.C.; Ma, J.Y.; Adrahtas, A.; Cheung, R.Y.; Krum, H. Celecoxib, but not rofecoxib or naproxen, attenuates cardiac hypertrophy and fibrosis induced in vitro by angiotensin and aldosterone. Clin. Exp. Pharmacol. Physiol. 2010, 37, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Shui, F.; Qiu, G.; Pan, S.; Wang, X.; Jia, F.; Jiang, T.; Li, Y.; Geng, Z.; Jin, S. Identification of stable reference genes for quantitative gene expression analysis in the duodenum of meat-type ducks. Front. Vet. Sci. 2023, 10, 1160384. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay ID | Gene | Gene Symbol | Cat. No. |
|---|---|---|---|
| Hs01031809_m1 | Human Mineralocorticoid Receptor (MR) | NR3C2 | 4331182 |
| Hs001741341_m1 | Human Interluekin 6 | IL-6 | 4331182 |
| Hs00998133_m1 | Human Transforming Growth Factor-β | TGFβ | 4331182 |
| Hs00892519_m1 | Human Thyroid Peroxidase | TPO | 4331182 |
| Hs00174974_m1 | Human Thyreoglobulin | TG | 4331182 |
| Hs00166567_m1 | Human Sodium/Iodide symporter (NIS) | SLC5A5 | 4331182 |
| Hs00388669_m1 | Human Hydroxysteroid 11-β dehydrogenase 2 | HSD11B2 | 4331182 |
| Rn00565562_m1 | Rat Mineralocorticoid Receptor (MR) | NR3C2 | 4448892 |
| Rn01410330_m1 | Rat Interluekin 6 | IL-6 | 4448892 |
| Rn00572010_m1 | Rat Transforming Growth Factor-β | TGFβ | 4448892 |
| Rn00571159_m1 | Rat Thyroid Peroxidase | TPO | 4448892 |
| Rn00667257_g1 | Rat Thyreoglobulin | TG | 4448892 |
| Rn00583900_m1 | Rat Sodium/Iodide Symporter (NIS) | SLC5A5 | 4448892 |
| Rn00492539_m1 | Rat Hydroxysteroid 11-β Dehydrogenase 2 | HSD11B2 | 4448892 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manso, J.; Pedron, M.C.; Mondin, A.; Censi, S.; Pennelli, G.; Galuppini, F.; Barollo, S.; Bertazza, L.; Radu, C.M.; Ghini, F.; et al. First Evidence of Mineralocorticoid Receptor Gene and Protein Expression in Rat and Human Thyroid Tissues and Cell Cultures. Int. J. Mol. Sci. 2024, 25, 754. https://doi.org/10.3390/ijms25020754
Manso J, Pedron MC, Mondin A, Censi S, Pennelli G, Galuppini F, Barollo S, Bertazza L, Radu CM, Ghini F, et al. First Evidence of Mineralocorticoid Receptor Gene and Protein Expression in Rat and Human Thyroid Tissues and Cell Cultures. International Journal of Molecular Sciences. 2024; 25(2):754. https://doi.org/10.3390/ijms25020754
Chicago/Turabian StyleManso, Jacopo, Maria Chiara Pedron, Alberto Mondin, Simona Censi, Gianmaria Pennelli, Francesca Galuppini, Susi Barollo, Loris Bertazza, Claudia Maria Radu, Francesca Ghini, and et al. 2024. "First Evidence of Mineralocorticoid Receptor Gene and Protein Expression in Rat and Human Thyroid Tissues and Cell Cultures" International Journal of Molecular Sciences 25, no. 2: 754. https://doi.org/10.3390/ijms25020754
APA StyleManso, J., Pedron, M. C., Mondin, A., Censi, S., Pennelli, G., Galuppini, F., Barollo, S., Bertazza, L., Radu, C. M., Ghini, F., Simioni, P., Sabbadin, C., Ceccato, F., Armanini, D., & Mian, C. (2024). First Evidence of Mineralocorticoid Receptor Gene and Protein Expression in Rat and Human Thyroid Tissues and Cell Cultures. International Journal of Molecular Sciences, 25(2), 754. https://doi.org/10.3390/ijms25020754