
The CYP80A and CYP80G Are Involved in the Biosynthesis of Benzylisoquinoline Alkaloids in the Sacred Lotus (Nelumbo nucifera)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
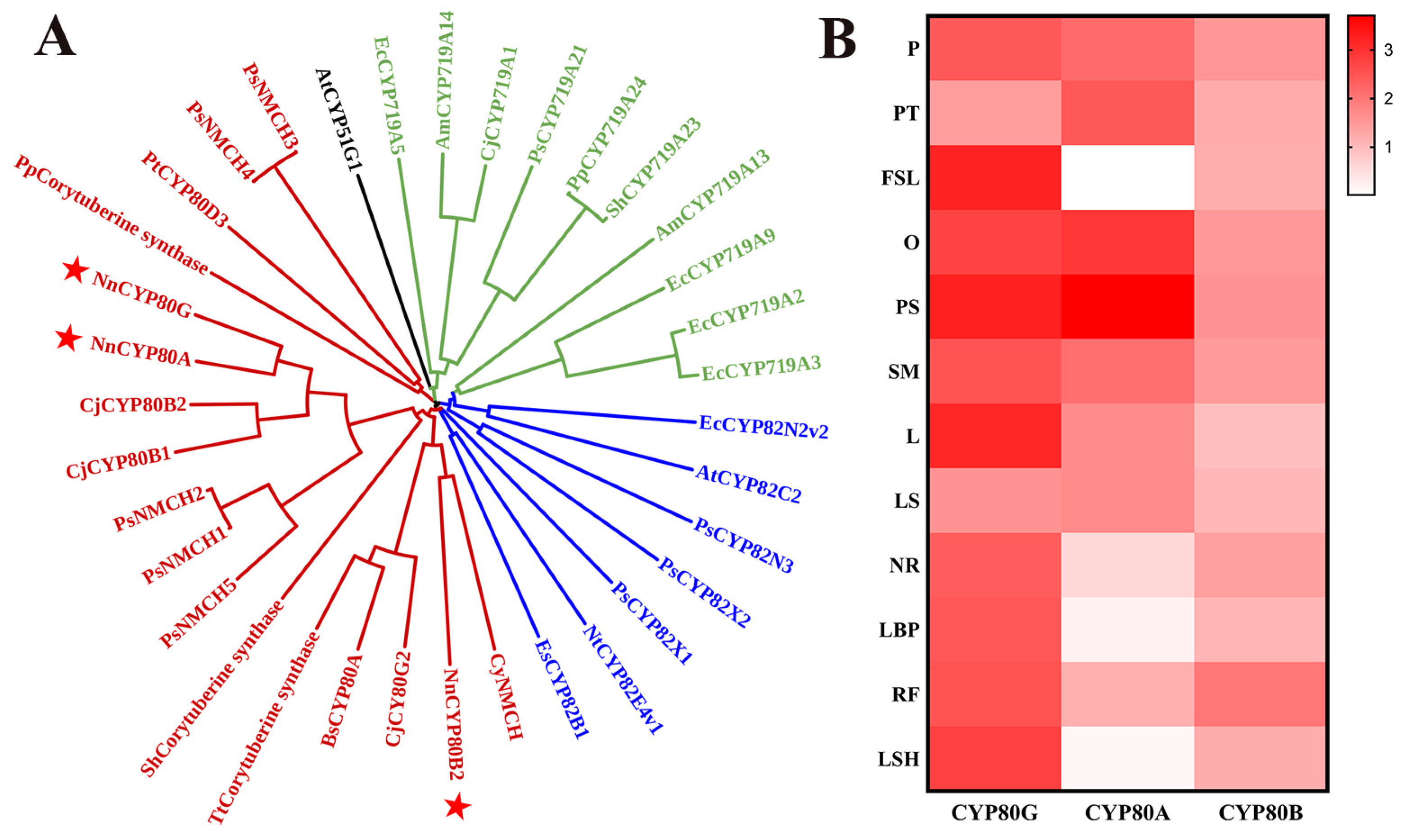
2.1. Cloning and Identification of CYP80 from N. nucifera
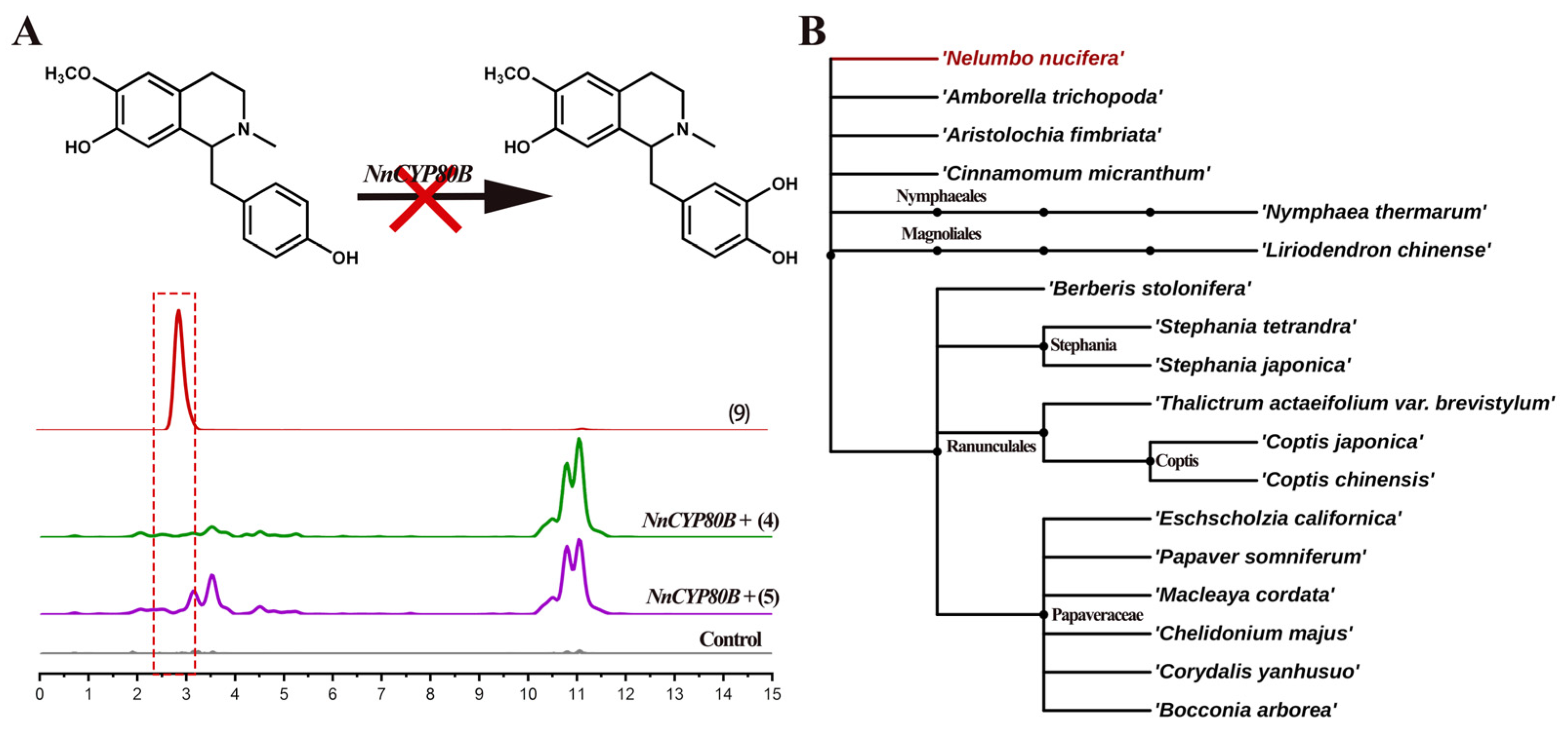
2.2. Functional Characterization of NnCYP80A
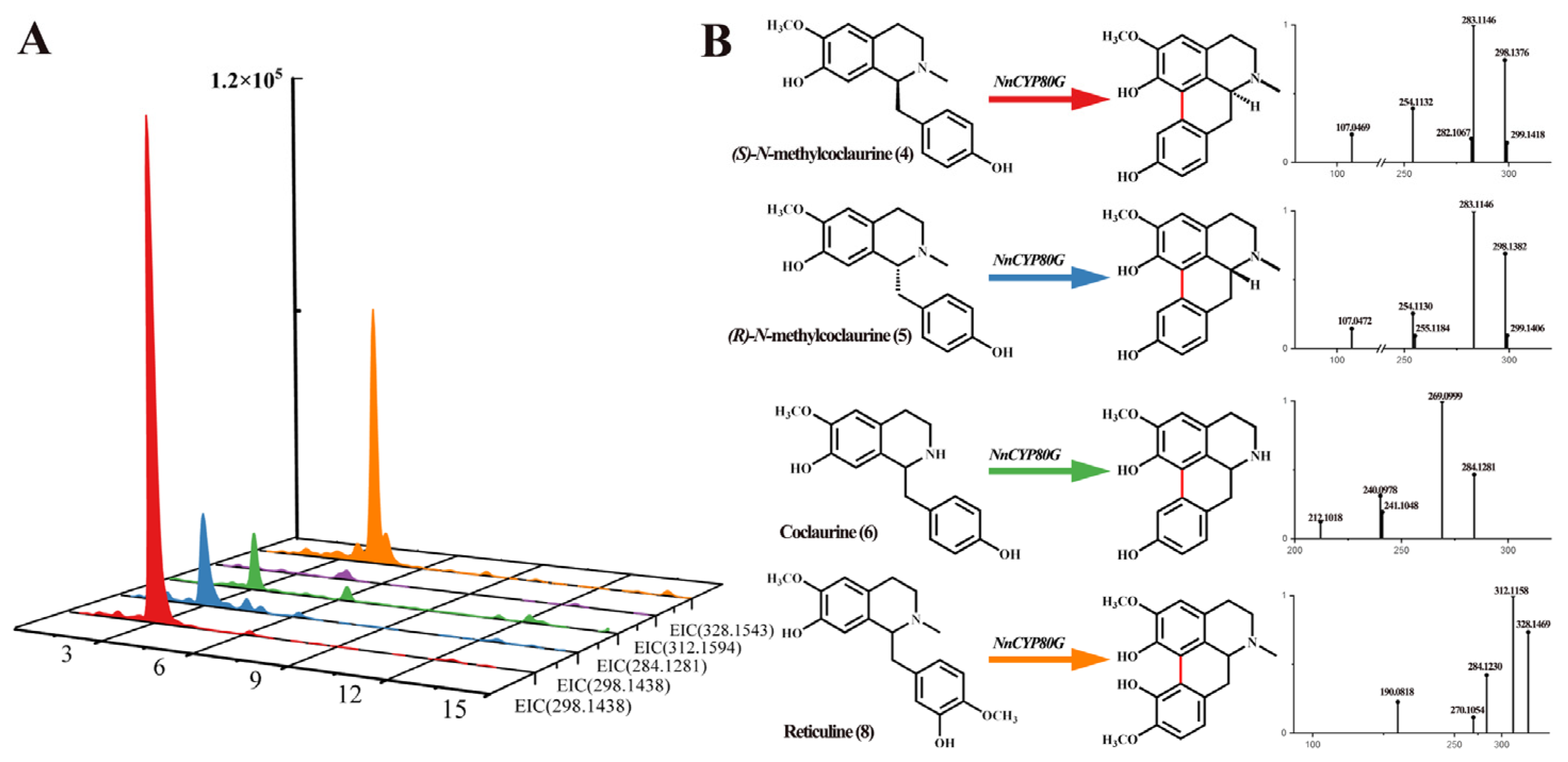
2.3. Functional Characterization of NnCYP80G
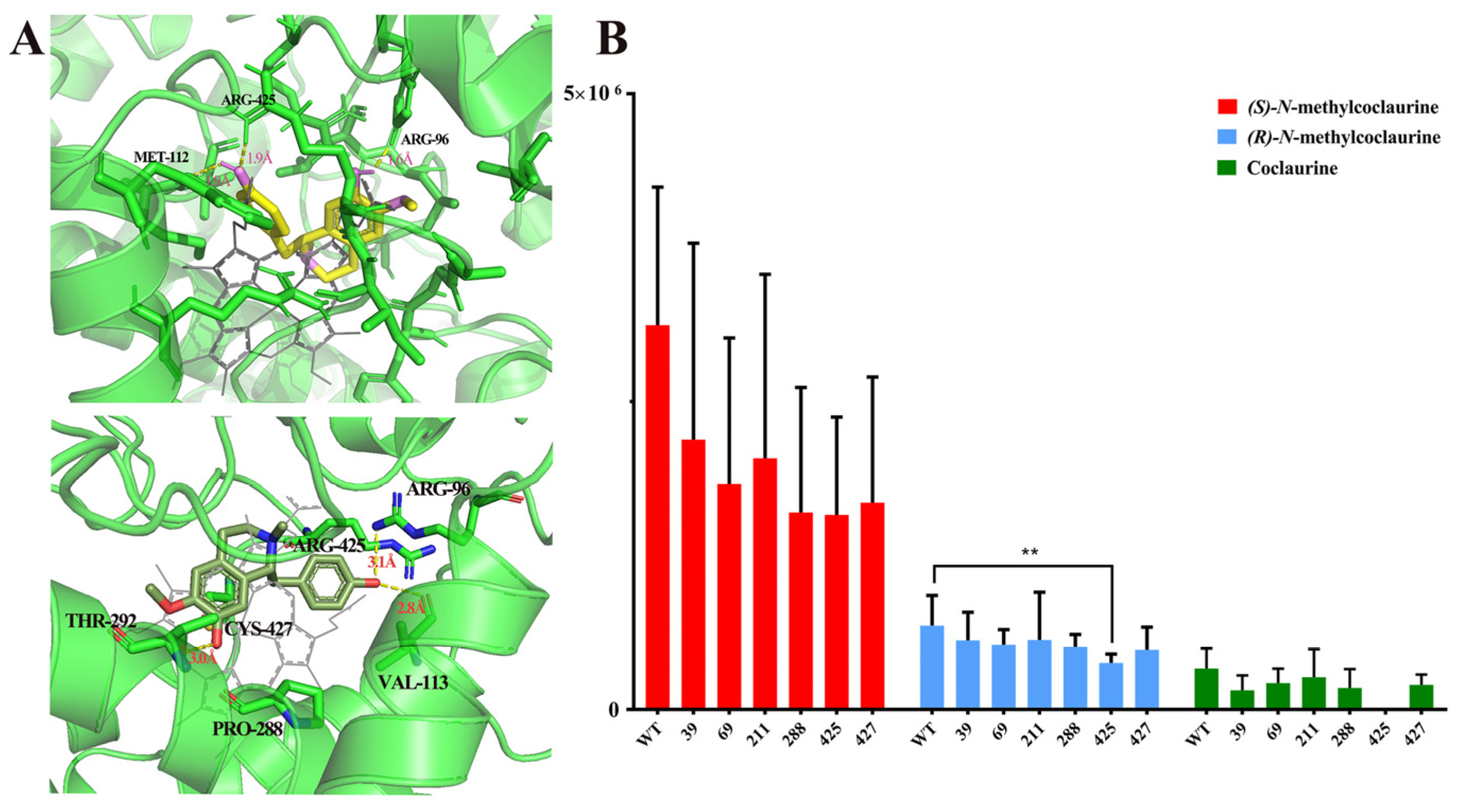
2.4. Mutational Analysis of C-C Coupling by NnCYP80G
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Chemicals
4.2. Gene Cloning and Enzyme Expression in Yeast
4.3. LC–MS Analysis
4.4. Modeling Docking and Mutagenesis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ziegler, J.; Facchini, P.J. Alkaloid Biosynthesis: Metabolism and Trafficking. Annu. Rev. Plant. Biol. 2008, 59, 735–769. [Google Scholar] [CrossRef]
- Menéndez-Perdomo, I.; Facchini, P. Benzylisoquinoline Alkaloids Biosynthesis in Sacred Lotus. Molecules 2018, 23, 2899. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.; Chakraborty, P. A review on sources and health impacts of bisphenol A. Rev. Environ. Health 2020, 35, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Wicks, C.; Hudlicky, T.; Rinner, U. Morphine alkaloids: History, biology, and synthesis. Alkaloids. Chem. Biol. 2021, 86, 145–342. [Google Scholar] [PubMed]
- Liang, D.; Li, Q.; Du, L.; Dou, G. Pharmacological Effects and Clinical Prospects of Cepharanthine. Molecules 2022, 27, 8933. [Google Scholar] [CrossRef] [PubMed]
- He, C.L.; Huang, L.Y.; Wang, K.; Gu, C.J.; Huang, A.L. Identification of bis-benzylisoquinoline alkaloids as SARS-CoV-2 entry inhibitors from a library of natural products. Signal. Transduct. Target. Ther. 2021, 6, 131. [Google Scholar] [CrossRef]
- Priya, L.B.; Huang, C.Y.; Hu, R.M.; Balasubramanian, B.; Baskaran, R. An updated review on pharmacological properties of neferine—A bisbenzylisoquinoline alkaloid from Nelumbo nucifera. J. Food Biochem. 2021, 45, e13986. [Google Scholar]
- Zhang, C.; Deng, J.; Liu, D.; Tuo, X.; Xiao, L.; Lai, B.; Yao, Q.; Liu, J.; Yang, H.; Wang, N. Nuciferine ameliorates hepatic steatosis in high-fat diet/streptozocin-induced diabetic mice through PPARα/PGC1α pathway. Br. J. Pharmacol. 2018, 17, 4218–4228. [Google Scholar] [CrossRef]
- Liu, J.; Shi, K.; Shi, J.; Feng, Y.; Chen, S. A simple strategy to monitor the temporal and spatial distribution of alkaloids in sacred lotus leaves. Biosci. Biotechnol. Biochem. 2021, 85, 1332–1340. [Google Scholar] [CrossRef]
- Hao, C.; Yu, Y.; Zhang, X.; Dong, G.; Liu, Y.; Chen, S. Nontarget Metabolites of Rhizomes of Edible Sacred Lotus Provide New Insights into Rhizome Browning. J. Food Qual. 2022, 2022, 3943052. [Google Scholar] [CrossRef]
- Li, Y.; Li, S.; Thodey, K.; Trenchard, I.; Cravens, A.; Smolke, C.D. Complete biosynthesis of noscapine and halogenated alkaloids in yeast. Proc. Natl. Acad. Sci. USA 2018, 115, E3922–E3931. [Google Scholar] [CrossRef] [PubMed]
- Nakagaawa, A.; Matsumura, E.K.T.; Katayama, T.; Kawano, N.; Yoshimatsu, K.; Yamamoto, K.; Kumagai, H.; Sato, F.; Mina, H. Efficient Total Biosynthesis of Opiates by Using Engineered Escherichia Coli. Synfacts 2016, 12, 529. [Google Scholar]
- Trenchard, I.J.; Siddiqui, M.S.; Thodey, K.; Smolke, C.D. De novo production of the key branch point benzylisoquinoline alkaloid reticuline in yeast. Metab. Eng. 2015, 31, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Menéndez-Perdomo, I.; Facchini, P.J. Isolation and characterization of two O-methyltransferases involved in benzylisoquinoline alkaloid biosynthesis in sacred lotus (Nelumbo nucifera). J. Biol. Chem. 2019, 295, 1598–1612. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Liu, Y.; Dong, G.; Jiang, J.; Leng, L.; Liu, X.; Zhang, J.; Liu, A.; Chen, S. Functional characterization and key residues engineering of a regiopromiscuity O-methyltransferase involved in benzylisoquinoline alkaloid biosynthesis in Nelumbo nucifera. Hortic. Res. 2022, 10, uhac276. [Google Scholar] [CrossRef]
- Yang, M.; Zhu, L.; Li, L.; Li, J.; Xu, L.; Feng, J.; Liu, Y. Digital Gene Expression Analysis Provides Insight into the Transcript Profile of the Genes Involved in Aporphine Alkaloid Biosynthesis in Lotus (Nelumbo nucifera). Front. Plant. Sci. 2017, 8, 80. [Google Scholar] [CrossRef]
- Meelaph, T.; Kobtrakul, K.; Chansilpa, N.N.; Han, Y.; Vimolmangkang, S. Coregulation of Biosynthetic Genes and Transcription Factors for Aporphine-Type Alkaloid Production in Wounded Lotus Provides Insight into the Biosynthetic Pathway of Nuciferine. ACS Omega 2018, 3, 8794–8802. [Google Scholar] [CrossRef]
- Pyne, M.E.; Gold, N.D.; Martin, V.J.J. Pathway elucidation and microbial synthesis of proaporphine and bis-benzylisoquinoline alkaloids from sacred lotus (Nelumbo nucifera). Metab. Eng. 2023, 77, 162–173. [Google Scholar] [CrossRef]
- Deng, X.; Zhao, L.; Fang, T.; Xiong, Y.; Ogutu, C.; Yang, D.; Vimolmangkang, S.; Liu, Y.; Han, Y. Investigation of benzylisoquinoline alkaloid biosynthetic pathway and its transcriptional regulation in lotus. Hortic. Res. 2018, 5, 16. [Google Scholar] [CrossRef]
- Liscombe, D.K.; Macleod, B.P.; Loukanina, N.; Nandi, O.I.; Facchini, P.J. Evidence for the monophyletic evolution of benzylisoquinoline alkaloid biosynthesis in angiosperms. Phytochemistry 2005, 66, 1374–1393. [Google Scholar] [CrossRef]
- Singh, A.; Menendez-Perdomo, I.M.; Facchini, P.J. Benzylisoquinoline alkaloid biosynthesis in opium poppy: An update. Phytochem. Rev. 2019, 18, 1457–1482. [Google Scholar] [CrossRef]
- Nguyen, T.D.; Dang, T.T. Cytochrome P450 Enzymes as Key Drivers of Alkaloid Chemical Diversification in Plants. Front. Plant. Sci. 2021, 12, 682181. [Google Scholar] [CrossRef] [PubMed]
- Dastmalchi, M.; Park, M.R.; Morris, J.S.; Facchini, P. Family portraits: The enzymes behind benzylisoquinoline alkaloid diversity. Phytochem. Rev. 2018, 17, 249–277. [Google Scholar] [CrossRef]
- Kraus, P.F.; Kutchan, T.M. Molecular cloning and heterologous expression of a cDNA encoding berbamunine synthase, a C--O phenol-coupling cytochrome P450 from the higher plant Berberis stolonifera. Proc. Natl. Acad. Sci. USA 1995, 92, 2071–2075. [Google Scholar] [CrossRef] [PubMed]
- Ikezawa, N.; Iwasa, K.; Sato, F. Molecular cloning and characterization of CYP80G2, a cytochrome P450 that catalyzes an intramolecular C-C phenol coupling of (S)-reticuline in magnoflorine biosynthesis from cultured Coptis japonica cells. J. Biol. Chem. 2008, 283, 8810–8821. [Google Scholar] [CrossRef]
- Liu, X.; Bu, J.; Ma, Y.; Chen, Y.; Li, Q.; Jiao, X.; Hu, Z.; Cui, G.; Tang, J.; Guo, J.; et al. Functional characterization of (S)-N-methylcoclaurine 3’-hydroxylase (NMCH) involved in the biosynthesis of benzylisoquinoline alkaloids in Corydalis yanhusuo. Plant. Physiol. Biochem. 2021, 168, 507–515. [Google Scholar] [CrossRef]
- Shahwar, D.; Ahmad, N.; Yasmeen, A.; Khan, M.A.; Ullah, S.; Rahman, A.U. Bioactive constituents from Croton sparsiflorus Morong. Nat. Prod. Res. 2015, 29, 274–276. [Google Scholar] [CrossRef]
- Pauli, H.H.; Kutchan, T.M. Molecular cloning and functional heterologous expression of two alleles encoding (S)-N-methylcoclaurine 3’-hydroxylase (CYP80B1), a new methyl jasmonate-inducible cytochrome P-450-dependent mono-oxygenase of benzylisoquinoline alkaloid biosynthesis. Plant. J. 1998, 13, 793–801. [Google Scholar] [CrossRef]
- Tang, M.C.; Zou, Y.; Watanabe, K.; Walsh, C.T.; Tang, Y. Oxidative Cyclization in Natural Product Biosynthesis. Chem. Rev. 2017, 117, 5226–5333. [Google Scholar] [CrossRef]
- Li, Y.; Winzer, T.; He, Z.; Graham, I.A. Over 100 Million Years of Enzyme Evolution Underpinning the Production of Morphine in the Papaveraceae Family of Flowering Plants. Plant. Commun. 2020, 1, 16. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, C.; Yu, Y.; Liu, Y.; Liu, A.; Chen, S. The CYP80A and CYP80G Are Involved in the Biosynthesis of Benzylisoquinoline Alkaloids in the Sacred Lotus (Nelumbo nucifera). Int. J. Mol. Sci. 2024, 25, 702. https://doi.org/10.3390/ijms25020702
Hao C, Yu Y, Liu Y, Liu A, Chen S. The CYP80A and CYP80G Are Involved in the Biosynthesis of Benzylisoquinoline Alkaloids in the Sacred Lotus (Nelumbo nucifera). International Journal of Molecular Sciences. 2024; 25(2):702. https://doi.org/10.3390/ijms25020702
Chicago/Turabian StyleHao, Chenyang, Yuetong Yu, Yan Liu, An Liu, and Sha Chen. 2024. "The CYP80A and CYP80G Are Involved in the Biosynthesis of Benzylisoquinoline Alkaloids in the Sacred Lotus (Nelumbo nucifera)" International Journal of Molecular Sciences 25, no. 2: 702. https://doi.org/10.3390/ijms25020702
APA StyleHao, C., Yu, Y., Liu, Y., Liu, A., & Chen, S. (2024). The CYP80A and CYP80G Are Involved in the Biosynthesis of Benzylisoquinoline Alkaloids in the Sacred Lotus (Nelumbo nucifera). International Journal of Molecular Sciences, 25(2), 702. https://doi.org/10.3390/ijms25020702