
Exploring the Hemostatic Effects of Platelet Lysate-Derived Vesicles: Insights from Mouse Models

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
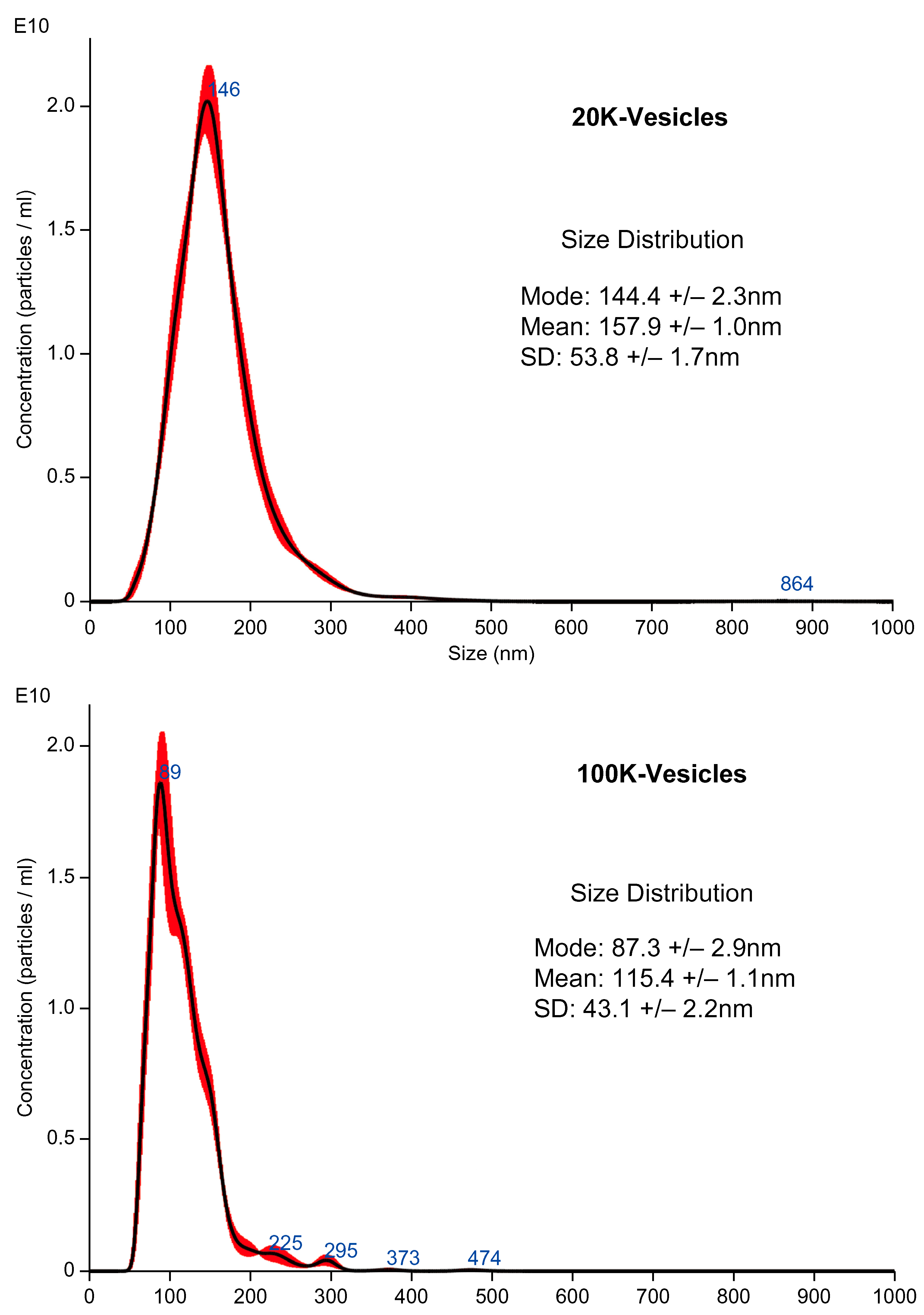
2.1. Particle Size Distribution and Concentration of Mouse PL (mPL)-Derived Vesicles
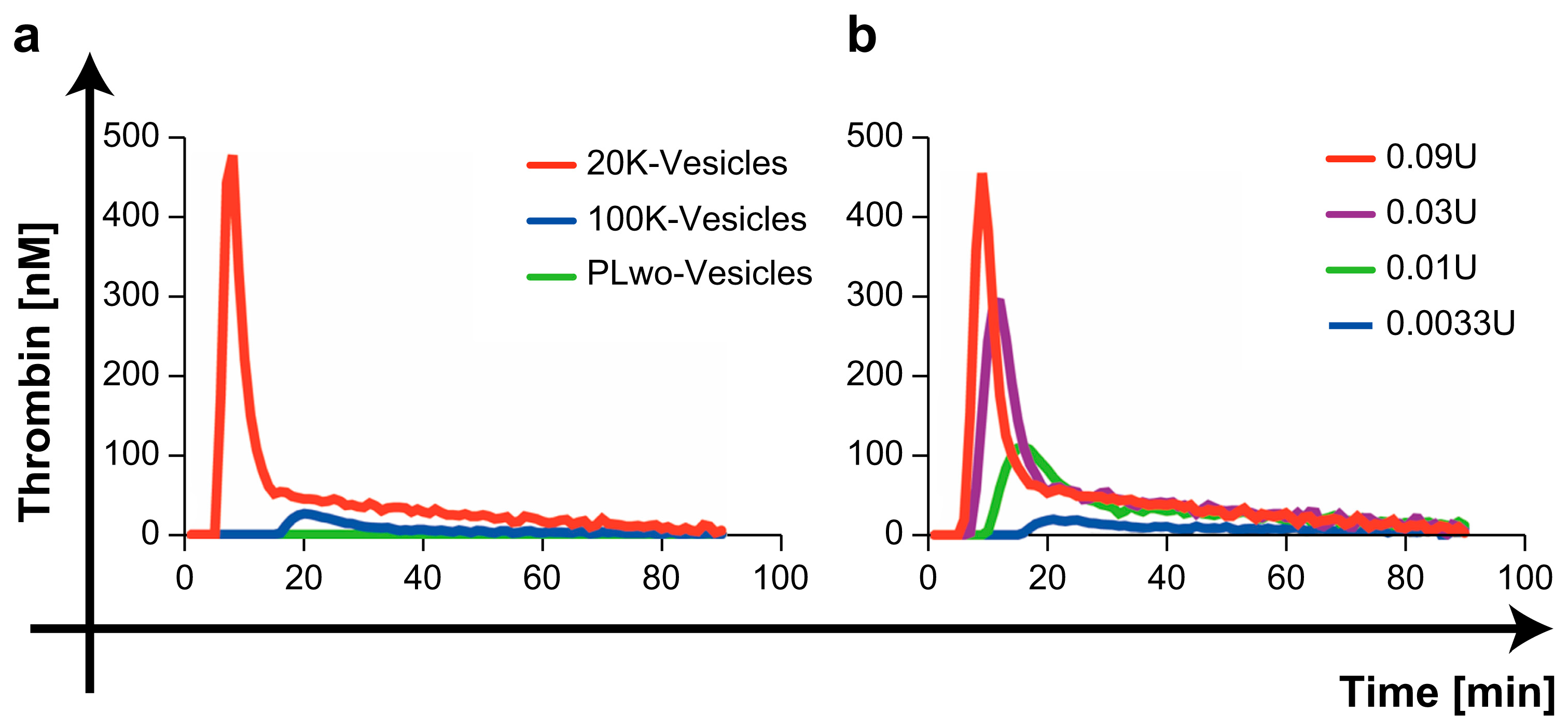
2.2. Comparison of Thrombin Generation Capacity by Three mPL-Derived Fractions
2.3. Platelet Intracellular Proteins in the PLwo-Vesicle Fraction
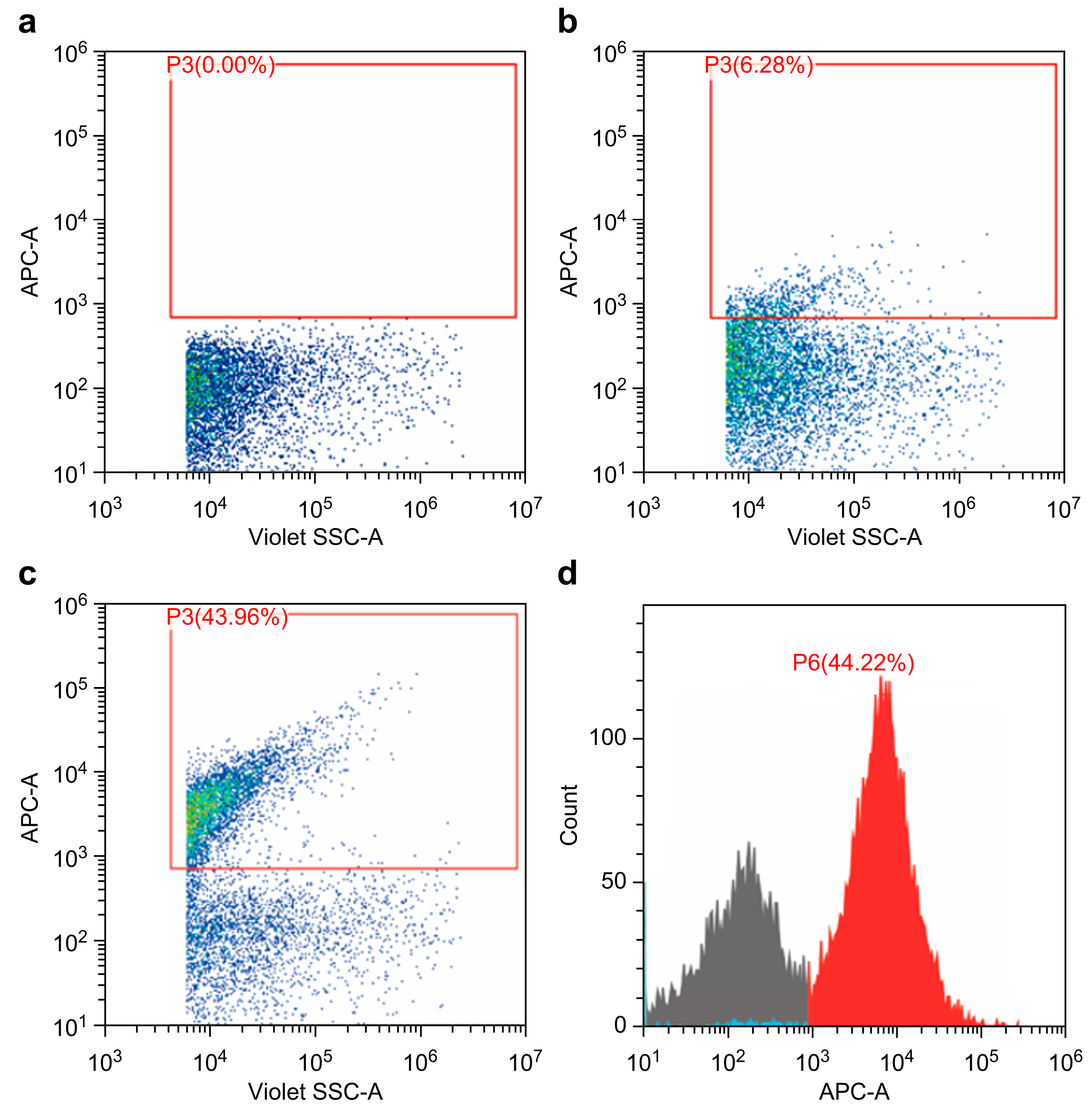
2.4. Phosphatidylserine and Factor X in 20K-Vesicles
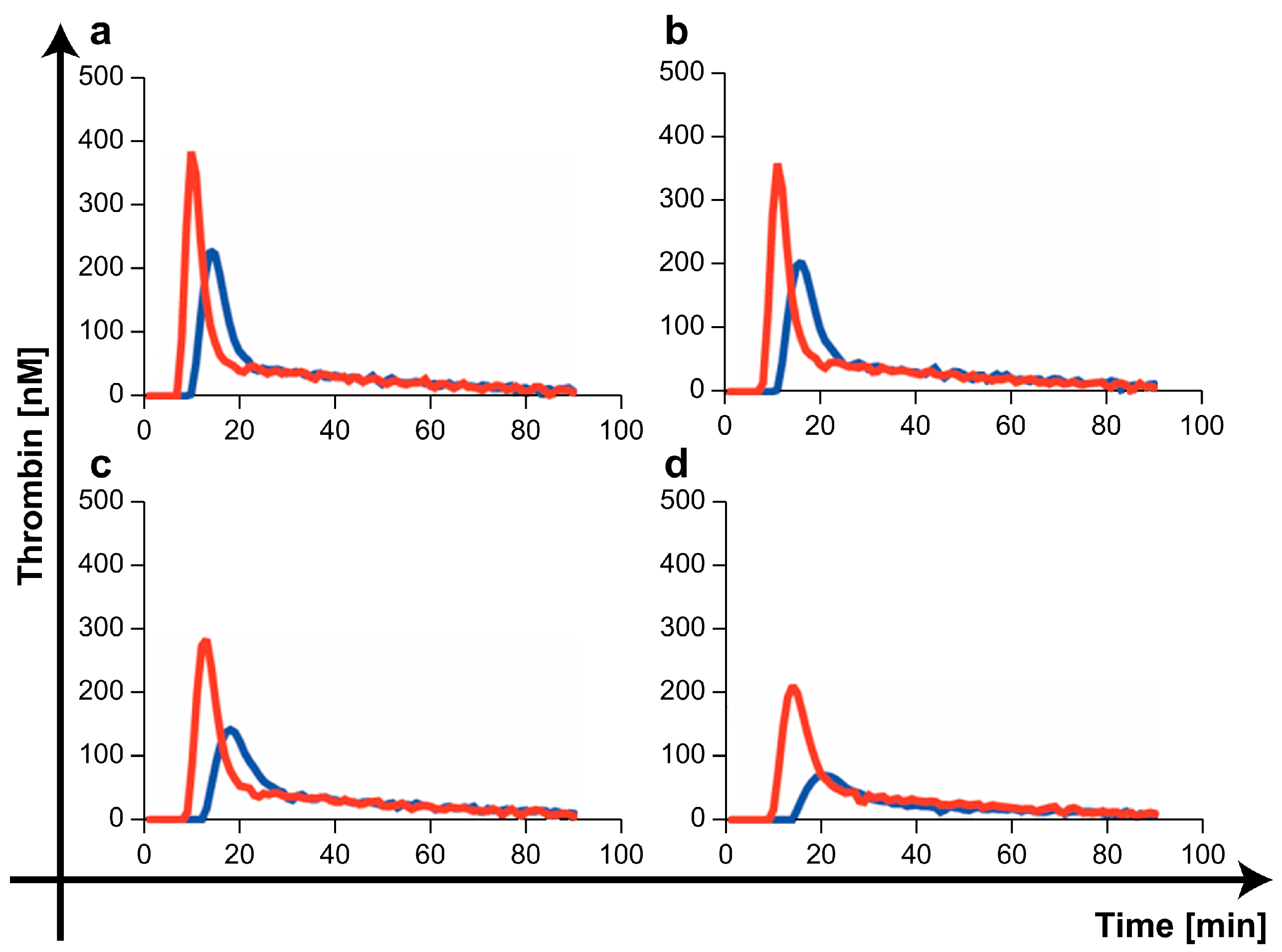
2.5. Procoagulant/Anticoagulant Effects of the Three mPL-Derived Fractions In Vitro
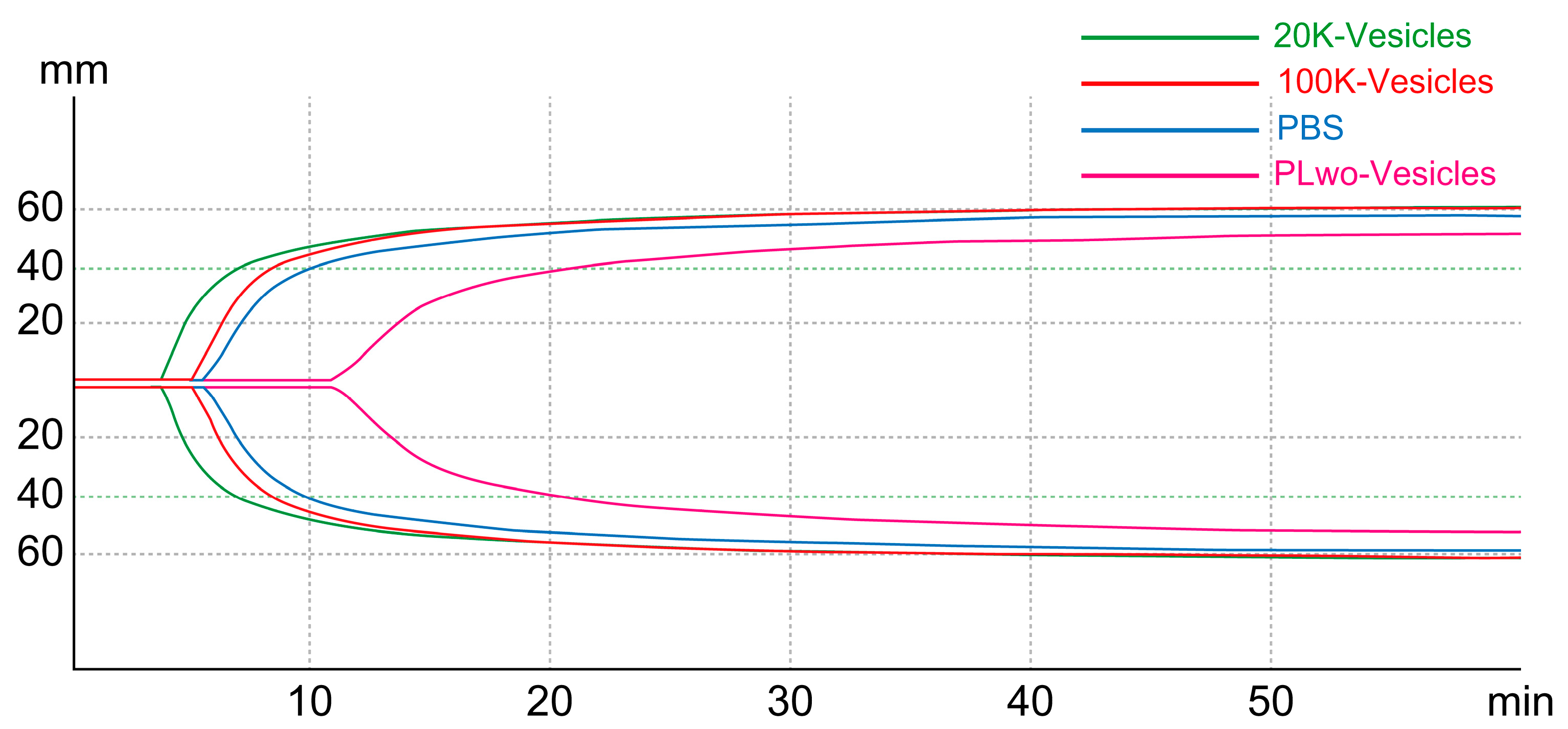
2.6. Hemostatic Effect of 20K-Vesicles in a Bleeding Model (Tail-Snip Bleeding Assay)
2.7. Thrombin–Antithrombin Complex (TAT) and Microthrombus in Various Organs 24 h after the Administration of 20K-Vesicles
2.8. Cytokine Concentration Associated with 20K-Vesicle Administration
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Preparation of PL
4.3. Preparation of mPL-Derived Fractions
4.4. Measurement of Particle Size Distribution of 20K-Vesicles and 100K-Vesicles
4.5. TGA
4.5.1. Preparation of MpFP
4.5.2. TGA Protocol
4.6. Measurement of Various Platelet-Derived Proteins in the PLwo-Vesicle Fraction
4.7. Characteristics of 20K-Vesicles
4.8. Viscoelastic Test
4.9. Tail-Snip Bleeding Assay
4.10. Thrombin–Antithrombin Complex (TAT) and Cytokine Levels after Intraperitoneal Administration of 20K-Vesicles
4.11. Histological Study
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tisherman, S.A.; Schmicker, R.H.; Brasel, K.J.; Bulger, E.M.; Kerby, J.D.; Minei, J.P.; Powell, J.L.; Reiff, D.A.; Rizoli, S.B.; Schreiber, M.A. Detailed Description of All Deaths in Both the Shock and Traumatic Brain Injury Hypertonic Saline Trials of the Resuscitation Outcomes Consortium. Ann. Surg. 2015, 261, 586–590. [Google Scholar] [CrossRef]
- Vinholt, P.J. The Role of Platelets in Bleeding in Patients with Thrombocytopenia and Hematological Disease. Clin. Chem. Lab. Med. 2019, 57, 1808–1817. [Google Scholar] [CrossRef]
- Sharma, S.; Sharma, P.; Tyler, L.N. Transfusion of Blood and Blood Products: Indications and Complications. Am. Fam. Physician 2011, 83, 719–724. [Google Scholar] [PubMed]
- van der Laan, E.A.N.Z.; van der Velden, S.; Porcelijn, L.; Semple, J.W.; van der Schoot, C.E.; Kapur, R. Evaluation of Platelet Responses in Transfusion-Related Acute Lung Injury (TRALI). Transfus. Med. Rev. 2020, 34, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Koupenova, M.; Kehrel, B.E.; Corkrey, H.A.; Freedman, J.E. Thrombosis and Platelets: An Update. Eur. Hear. J. 2016, 38, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, E.; Ghasemzadeh, M.; Nassaji, F.; Jamaat, Z.P. GPVI Modulation during Platelet Activation and Storage: Its Expression Levels and Ectodomain Shedding Compared to Markers of Platelet Storage Lesion. Platelets 2017, 28, 498–508. [Google Scholar] [CrossRef]
- Zhu, C.; Liang, Y.; Luo, Y.; Ma, X. Role of Pyroptosis in Hemostasis Activation in Sepsis. Front. Immunol. 2023, 14, 1114917. [Google Scholar] [CrossRef]
- Sinauridze, E.; Kireev, D.; Popenko, N.; Pichugin, A.; Panteleev, M.; Krymskaya, O.; Ataullakhanov, F. Platelet Microparticle Membranes Have 50- to 100-Fold Higher Specific Procoagulant Activity than Activated Platelets. Thromb. Haemost. 2007, 97, 425–434. [Google Scholar] [CrossRef]
- Nebie, O.; Buée, L.; Blum, D.; Burnouf, T. Can the Administration of Platelet Lysates to the Brain Help Treat Neurological Disorders? Cell. Mol. Life Sci. 2022, 79, 379. [Google Scholar] [CrossRef]
- Antich-Rosselló, M.; Forteza-Genestra, M.A.; Monjo, M.; Ramis, J.M. Platelet-Derived Extracellular Vesicles for Regenerative Medicine. Int. J. Mol. Sci. 2021, 22, 8580. [Google Scholar] [CrossRef]
- Johnson, J.; Wu, Y.W.; Blyth, C.; Lichtfuss, G.; Goubran, H.; Burnouf, T. Prospective Therapeutic Applications of Platelet Extracellular Vesicles. Trends Biotechnol. 2021, 39, 598–612. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.C.; Guo, S.C.; Zhang, C.Q. Platelet-Derived Extracellular Vesicles: An Emerging Therapeutic Approach. Int. J. Biol. Sci. 2017, 13, 828–834. [Google Scholar] [CrossRef]
- Raghunathan, S.; Rayes, J.; Gupta, A.S. Platelet-inspired Nanomedicine in Hemostasis Thrombosis and Thromboinflammation. J. Thromb. Haemost. 2022, 20, 1535–1549. [Google Scholar] [CrossRef]
- Bordin, A.; Chirivì, M.; Pagano, F.; Milan, M.; Iuliano, M.; Scaccia, E.; Fortunato, O.; Mangino, G.; Dhori, X.; Marinis, E.D.; et al. Human Platelet Lysate-derived Extracellular Vesicles Enhance Angiogenesis through MiR-126. Cell Prolif. 2022, 55, e13312. [Google Scholar] [CrossRef] [PubMed]
- Meftahpour, V.; Malekghasemi, S.; Baghbanzadeh, A.; Aghebati-Maleki, A.; Pourakbari, R.; Fotouhi, A.; Aghebati-Maleki, L. Platelet Lysate: A Promising Candidate in Regenerative Medicine. Regen. Med. 2021, 16, 71–85. [Google Scholar] [CrossRef]
- Yu, D.M.; Zhang, T.; Liu, J.H.; Wang, W.T.; Wang, W.B. The Molecular Mechanism of Platelet Lysate Promotes Transformation of Non-Union Cells into Osteoblasts. Transl. Cancer Res. 2020, 9, 1985–1992. [Google Scholar] [CrossRef]
- Szponder, T.; Latalski, M.; Danielewicz, A.; Krać, K.; Kozera, A.; Drzewiecka, B.; Ngoc, D.N.; Dobko, D.; Wessely-Szponder, J. Osteoarthritis: Pathogenesis, Animal Models, and New Regenerative Therapies. J. Clin. Med. 2022, 12, 5. [Google Scholar] [CrossRef]
- Rossi, S.; Mori, M.; Vigani, B.; Bonferoni, M.C.; Sandri, G.; Riva, F.; Caramella, C.; Ferrari, F. A Novel Dressing for the Combined Delivery of Platelet Lysate and Vancomycin Hydrochloride to Chronic Skin Ulcers: Hyaluronic Acid Particles in Alginate Matrices. Eur. J. Pharm. Sci. 2018, 118, 87–95. [Google Scholar] [CrossRef]
- Jeyaraman, M.; Muthu, S.; Khanna, M.; Jain, R.; Anudeep, T.C.; Muthukanagaraj, P.; Siddesh, S.E.; Gulati, A.; Satish, A.S.; Jeyaraman, N.; et al. Platelet Lysate for COVID-19 Pneumonia—A Newer Adjunctive Therapeutic Avenue. Stem Cell Investig. 2021, 8, 11. [Google Scholar] [CrossRef]
- Lopez, E.; Srivastava, A.K.; Pati, S.; Holcomb, J.B.; Wade, C.E. Platelet-Derived Microvesicles: A Potential Therapy for Trauma-Induced Coagulopathy. Shock 2017, 49, 243–248. [Google Scholar] [CrossRef]
- Lopez, E.; Srivastava, A.K.; Burchfield, J.; Wang, Y.W.; Cardenas, J.C.; Togarrati, P.P.; Miyazawa, B.; Gonzalez, E.; Holcomb, J.B.; Pati, S.; et al. Platelet-Derived- Extracellular Vesicles Promote Hemostasis and Prevent the Development of Hemorrhagic Shock. Sci. Rep. 2019, 9, 17676. [Google Scholar] [CrossRef] [PubMed]
- Bazzan, E.; Tinè, M.; Casara, A.; Biondini, D.; Semenzato, U.; Cocconcelli, E.; Balestro, E.; Damin, M.; Radu, C.M.; Turato, G.; et al. Critical Review of the Evolution of Extracellular Vesicles’ Knowledge: From 1946 to Today. Int. J. Mol. Sci. 2021, 22, 6417. [Google Scholar] [CrossRef] [PubMed]
- Matijevic, N.; Wang, Y.-W.W.; Kostousov, V.; Wade, C.E.; Vijayan, K.V.; Holcomb, J.B. Decline in Platelet Microparticles Contributes to Reduced Hemostatic Potential of Stored Plasma. Thromb. Res. 2011, 128, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Arbaeen, A.F.; Serrano, K.; Levin, E.; Devine, D.V. Platelet Concentrate Functionality Assessed by Thromboelastography or Rotational Thromboelastometry. Transfusion 2016, 56, 2790–2798. [Google Scholar] [CrossRef] [PubMed]
- Jerez-Dolz, D.; Torramade-Moix, S.; Palomo, M.; Moreno-Castaño, A.; Lopez-Vilchez, I.; Hernandez, R.; Badimon, J.J.; Zafar, M.U.; Diaz-Ricart, M.; Escolar, G. Internalization of Microparticles by Platelets Is Partially Mediated by Toll-like Receptor 4 and Enhances Platelet Thrombogenicity. Atherosclerosis 2020, 294, 17–24. [Google Scholar] [CrossRef]
- Lentz, B.R. Exposure of Platelet Membrane Phosphatidylserine Regulates Blood Coagulation. Prog. Lipid Res. 2003, 42, 423–438. [Google Scholar] [CrossRef] [PubMed]
- Holme, P.A.; Brosstad, F.; Solum, N.O. Platelet-Derived Microvesicles and Activated Platelets Express Factor Xa Activity. Blood Coagul. Fibrinolysis 1995, 6, 302–310. [Google Scholar] [CrossRef]
- Maroney, S.A.; Mast, A.E. Platelet Tissue Factor Pathway Inhibitor Modulates Intravascular Coagulation. Thromb. Res. 2012, 129, S21–S22. [Google Scholar] [CrossRef]
- Maroney, S.A.; Hansen, K.G.; Mast, A.E. Cellular Expression and Biological Activities of Alternatively Spliced Forms of Tissue Factor Pathway Inhibitor. Curr. Opin. Hematol. 2013, 20, 403–409. [Google Scholar] [CrossRef]
- Boulaftali, Y.; Adam, F.; Venisse, L.; Ollivier, V.; Richard, B.; Taieb, S.; Monard, D.; Favier, R.; Alessi, M.C.; Bryckaert, M.; et al. Anticoagulant and Antithrombotic Properties of Platelet Protease Nexin-1. Blood 2010, 115, 97–106. [Google Scholar] [CrossRef]
- Bouton, M.C.; Boulaftali, Y.; Richard, B.; Arocas, V.; Michel, J.B.; Jandrot-Perrus, M. Emerging Role of SerpinE2/Protease Nexin-1 in Hemostasis and Vascular Biology. Blood 2012, 119, 2452–2457. [Google Scholar] [CrossRef] [PubMed]
- Maroney, S.A.; Ferrel, J.P.; Pan, S.; White, T.A.; Simari, R.D.; McVey, J.H.; Mast, A.E. Temporal Expression of Alternatively Spliced Forms of Tissue Factor Pathway Inhibitor in Mice. J. Thromb. Haemost. 2009, 7, 1106–1115. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, B.; Trivedi, A.; Togarrati, P.P.; Potter, D.; Baimukanova, G.; Vivona, L.; Lin, M.; Lopez, E.; Callcut, R.; Srivastava, A.K.; et al. Regulation of Endothelial Cell Permeability by Platelet-Derived Extracellular Vesicles. J. Trauma Acute Care 2019, 86, 931–942. [Google Scholar] [CrossRef]
- Dyer, M.R.; Alexander, W.; Hassoune, A.; Chen, Q.; Brzoska, T.; Alvikas, J.; Liu, Y.; Haldeman, S.; Plautz, W.; Loughran, P.; et al. Platelet-derived Extracellular Vesicles Released after Trauma Promote Hemostasis and Contribute to DVT in Mice. J. Thromb. Haemost. 2019, 17, 1733–1745. [Google Scholar] [CrossRef]
- Noulsri, E. Effects of Cell-Derived Microparticles on Immune Cells and Potential Implications in Clinical Medicine. Lab. Med. 2020, 52, 122–135. [Google Scholar] [CrossRef]
- Fendl, B.; Eichhorn, T.; Weiss, R.; Tripisciano, C.; Spittler, A.; Fischer, M.B.; Weber, V. Differential Interaction of Platelet-Derived Extracellular Vesicles with Circulating Immune Cells: Roles of TAM Receptors, CD11b, and Phosphatidylserine. Front. Immunol. 2018, 9, 2797. [Google Scholar] [CrossRef] [PubMed]
- Somiya, M.; Yoshioka, Y.; Ochiya, T. Biocompatibility of Highly Purified Bovine Milk-Derived Extracellular Vesicles. J. Extracell. Vesicles 2018, 7, 1440132. [Google Scholar] [CrossRef]
- Refaai, M.A.; Conley, G.W.; Hudson, C.A.; Spinelli, S.L.; Phipps, R.P.; Morrell, C.N.; Blumberg, N.; McRae, H.L. Evaluation of the Procoagulant Properties of a Newly Developed Platelet Modified Lysate Product. Transfusion 2020, 60, 1579–1589. [Google Scholar] [CrossRef]
- Price, J.; Gardiner, C.; Harrison, P. Platelet-enhanced Plasma: Characterization of a Novel Candidate Resuscitation Fluid’s Extracellular Vesicle Content, Clotting Parameters, and Thrombin Generation Capacity. Transfusion 2021, 61, 2179–2194. [Google Scholar] [CrossRef]
- Aurbach, K.; Spindler, M.; Haining, E.J.; Bender, M.; Pleines, I. Blood Collection, Platelet Isolation and Measurement of Platelet Count and Size in Mice—A Practical Guide. Platelets 2019, 30, 698–707. [Google Scholar] [CrossRef]
- Spiel, A.O.; Mayr, F.B.; Firbas, C.; Quehenberger, P.; Jilma, B. Validation of Rotation Thrombelastography in a Model of Systemic Activation of Fibrinolysis and Coagulation in Humans. J. Thromb. Haemost. 2006, 4, 411–416. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirayu, N.; Takasu, O. Exploring the Hemostatic Effects of Platelet Lysate-Derived Vesicles: Insights from Mouse Models. Int. J. Mol. Sci. 2024, 25, 1188. https://doi.org/10.3390/ijms25021188
Hirayu N, Takasu O. Exploring the Hemostatic Effects of Platelet Lysate-Derived Vesicles: Insights from Mouse Models. International Journal of Molecular Sciences. 2024; 25(2):1188. https://doi.org/10.3390/ijms25021188
Chicago/Turabian StyleHirayu, Nobuhisa, and Osamu Takasu. 2024. "Exploring the Hemostatic Effects of Platelet Lysate-Derived Vesicles: Insights from Mouse Models" International Journal of Molecular Sciences 25, no. 2: 1188. https://doi.org/10.3390/ijms25021188
APA StyleHirayu, N., & Takasu, O. (2024). Exploring the Hemostatic Effects of Platelet Lysate-Derived Vesicles: Insights from Mouse Models. International Journal of Molecular Sciences, 25(2), 1188. https://doi.org/10.3390/ijms25021188