
Network Analysis of Metabolome and Transcriptome Revealed Regulation of Different Nitrogen Concentrations on Hybrid Poplar Cambium Development

 ,
,
Abstract
1. Introduction
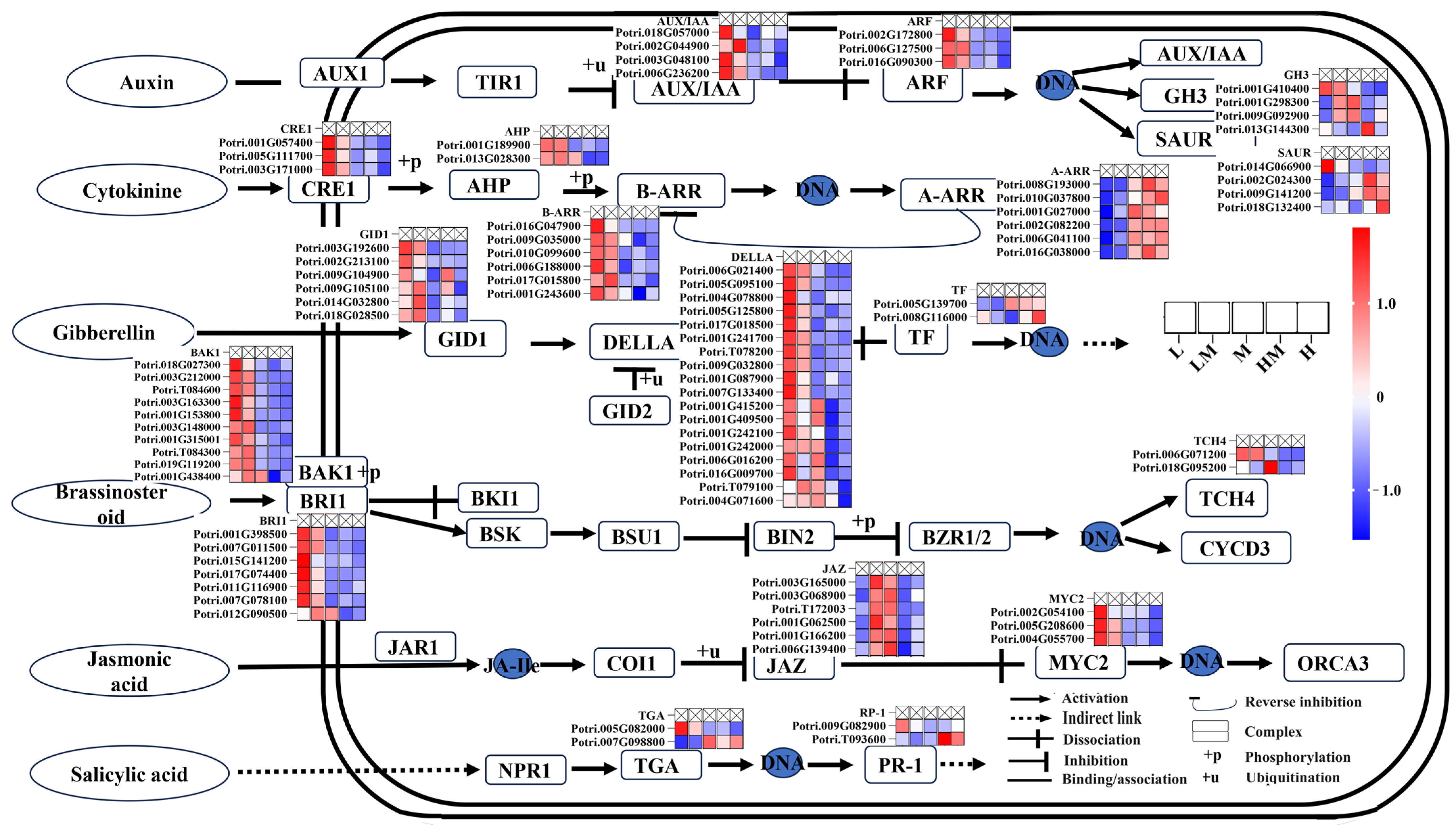
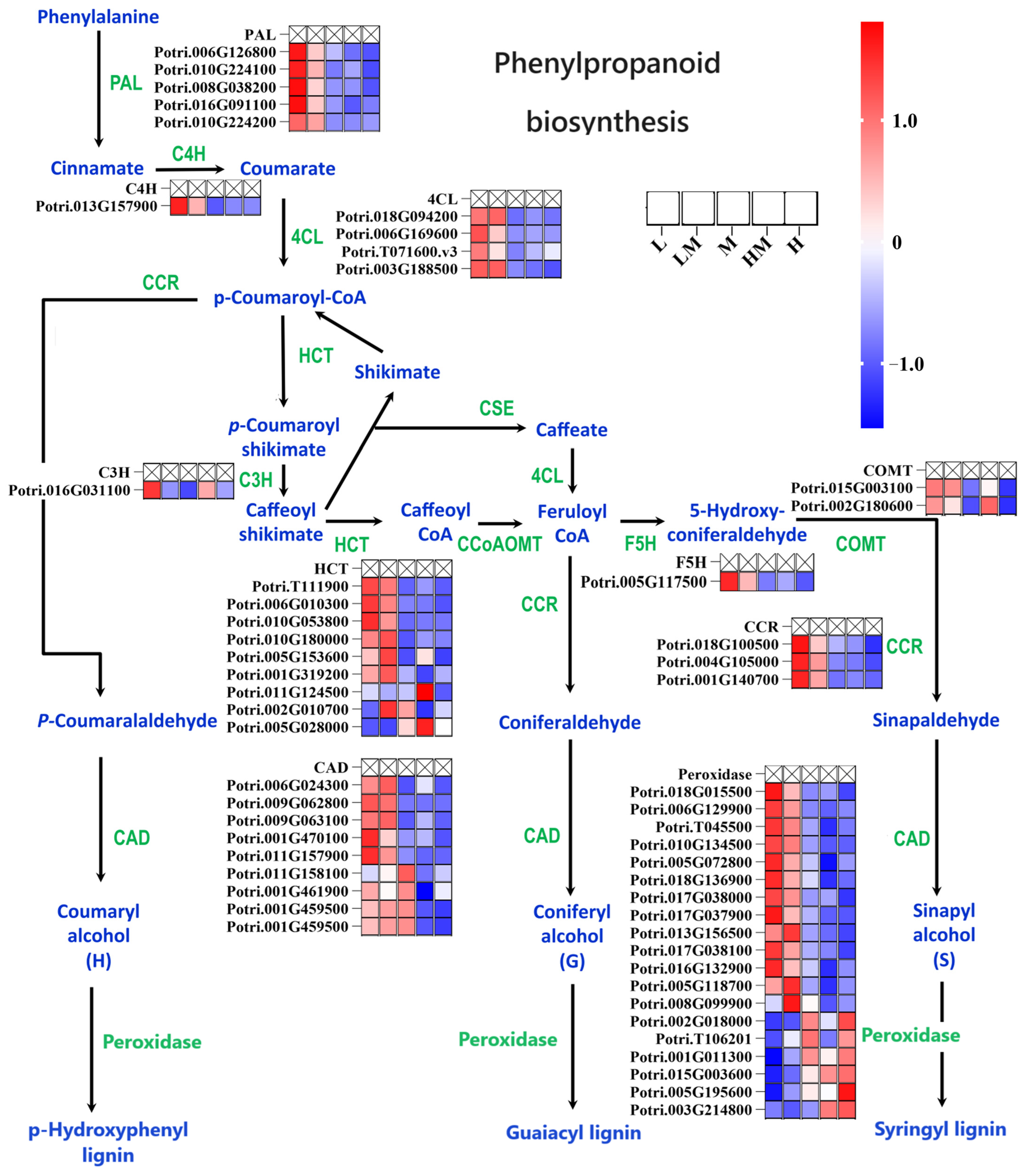
2. Results
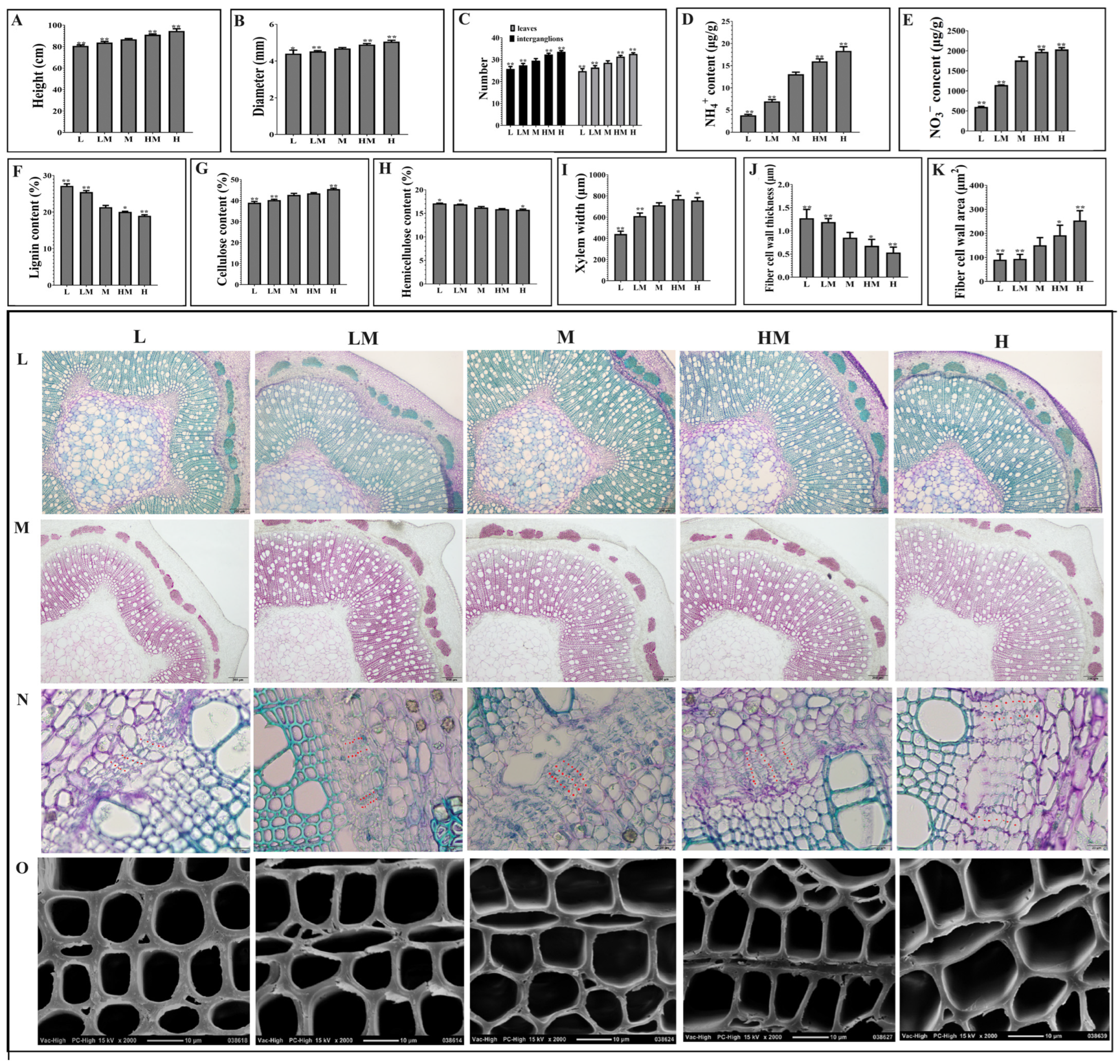
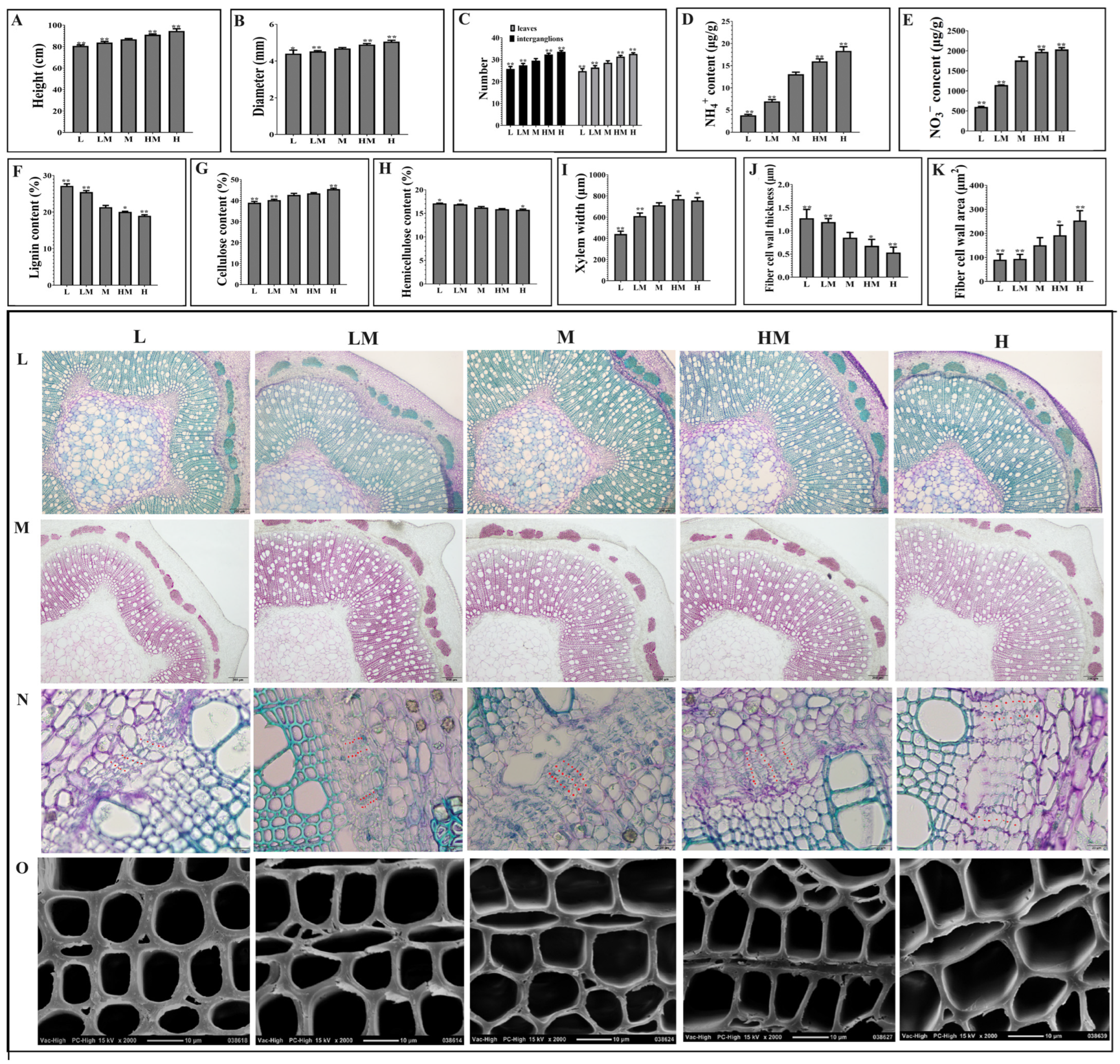
2.1. Effects of Different Nitrogen Concentrations on Growth and Wood Property of Poplar
2.2. Transcriptome Sequencing and Annotation
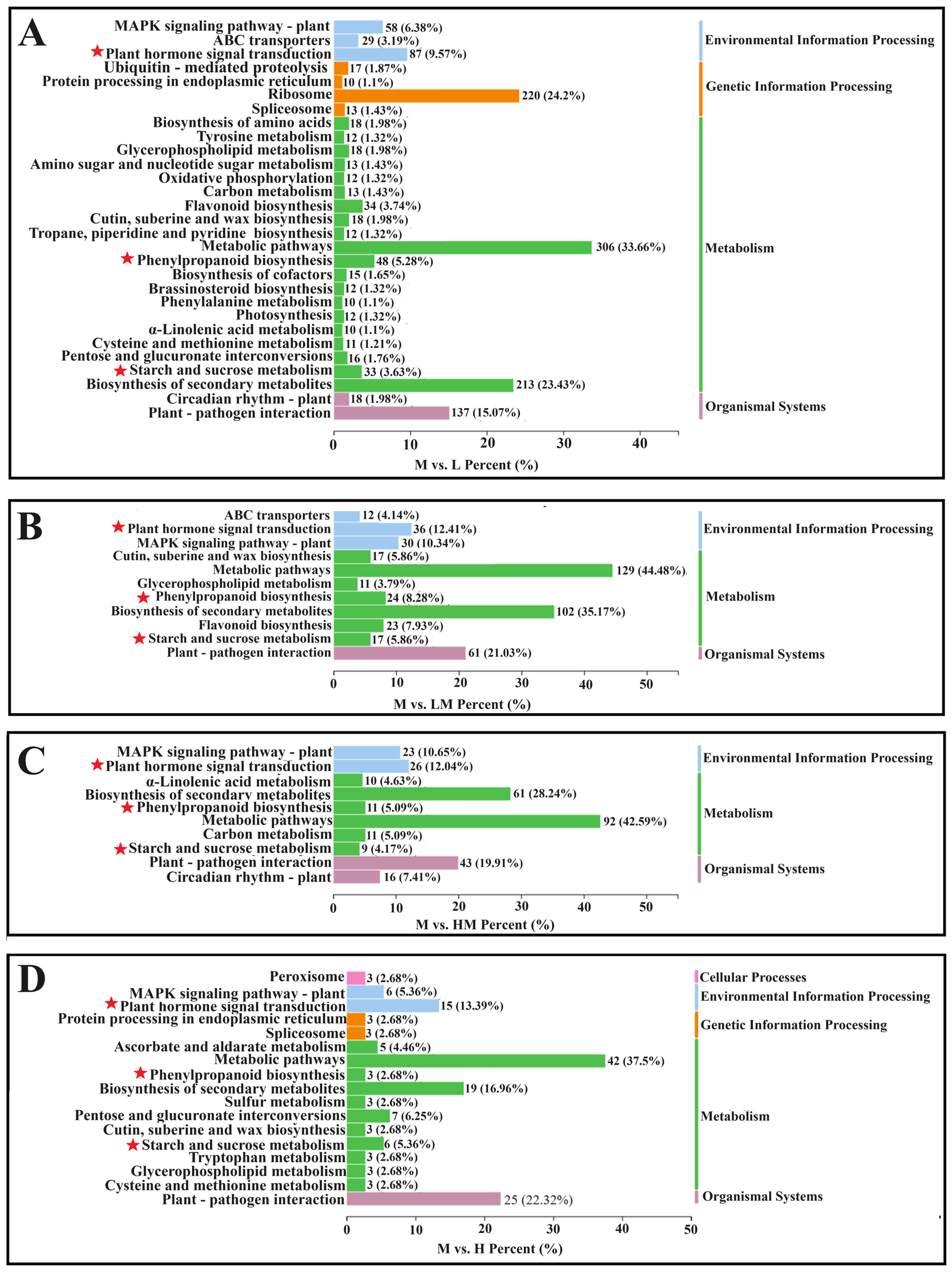
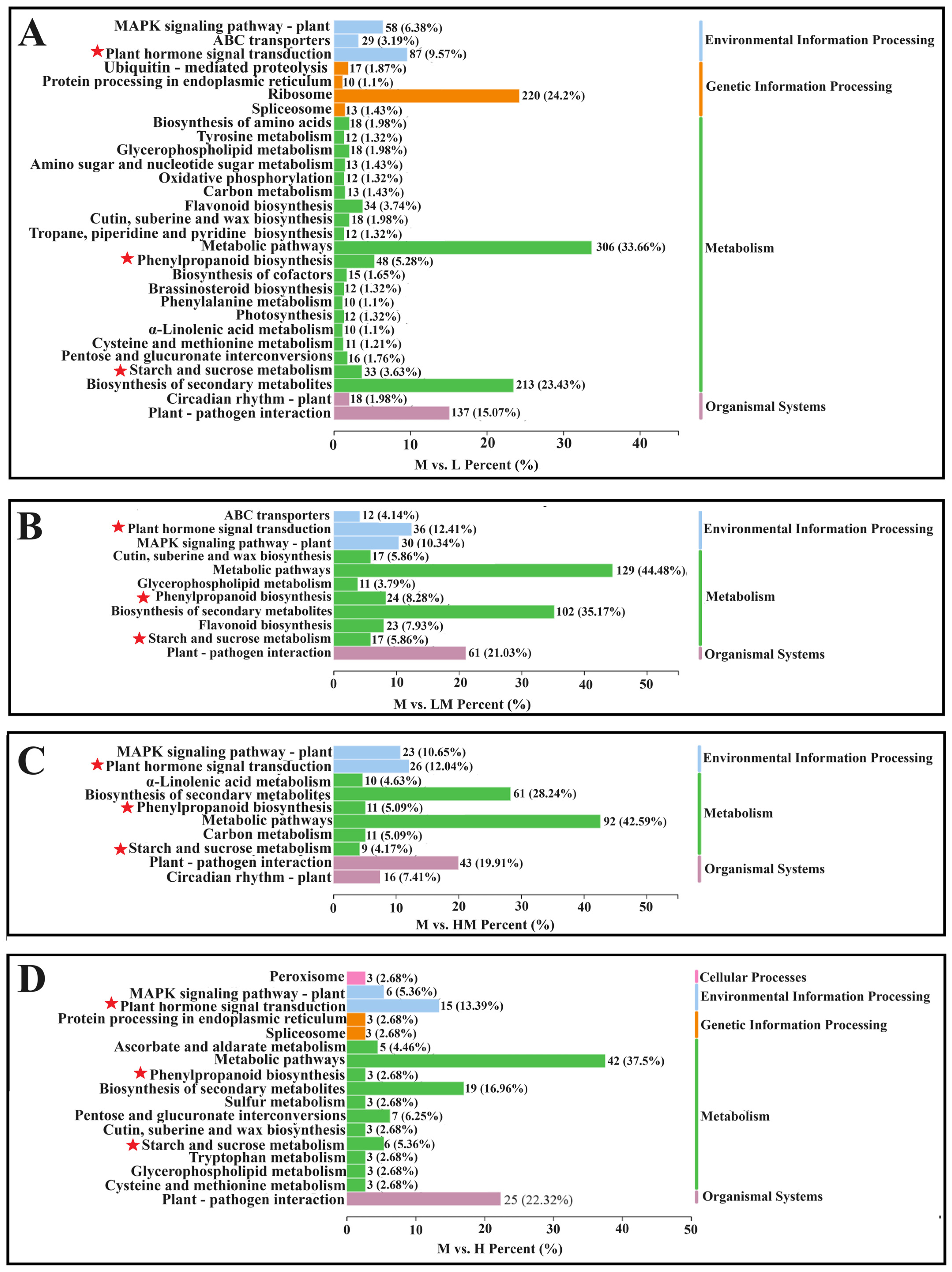
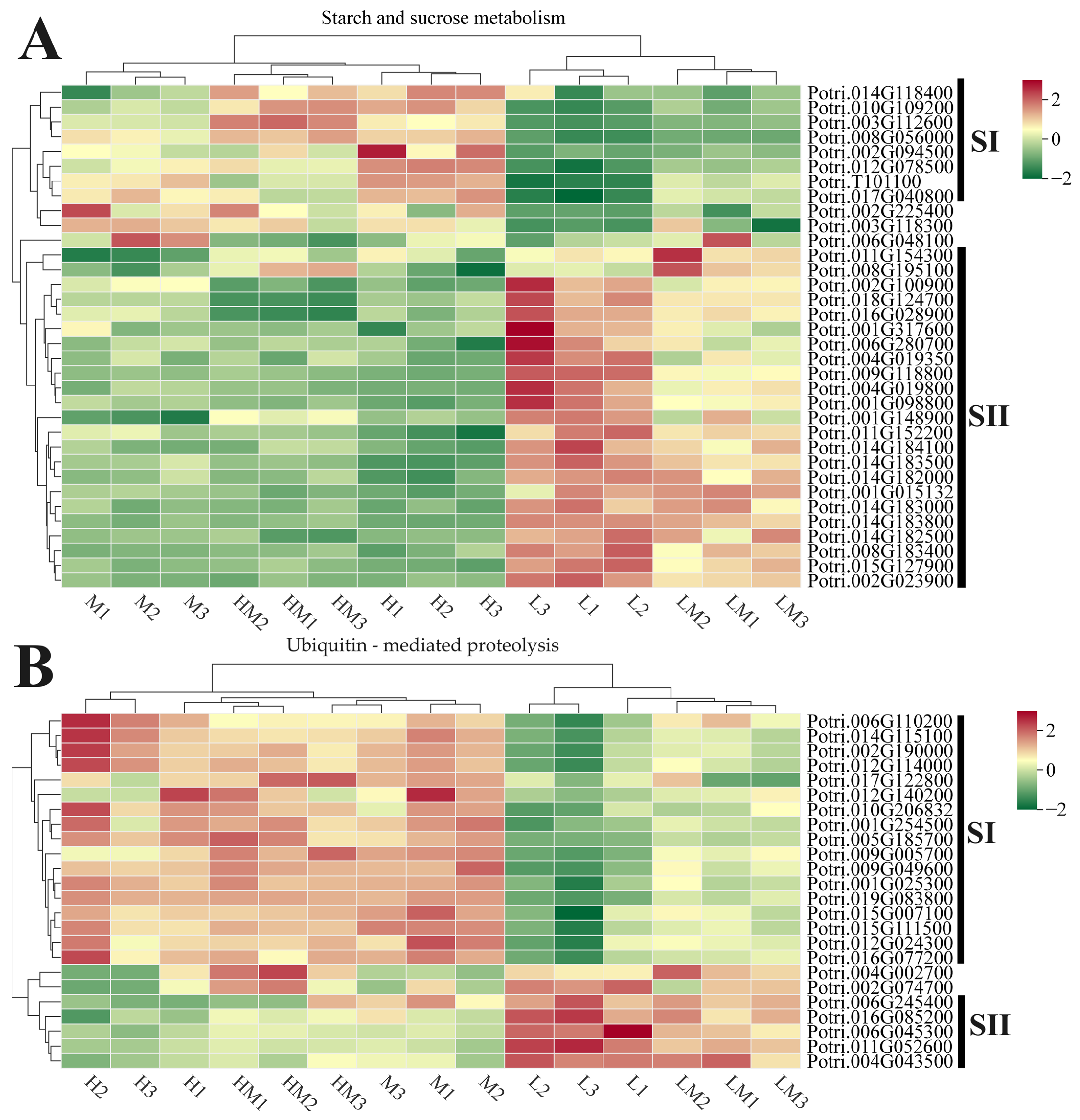
2.3. Analysis of DEGs
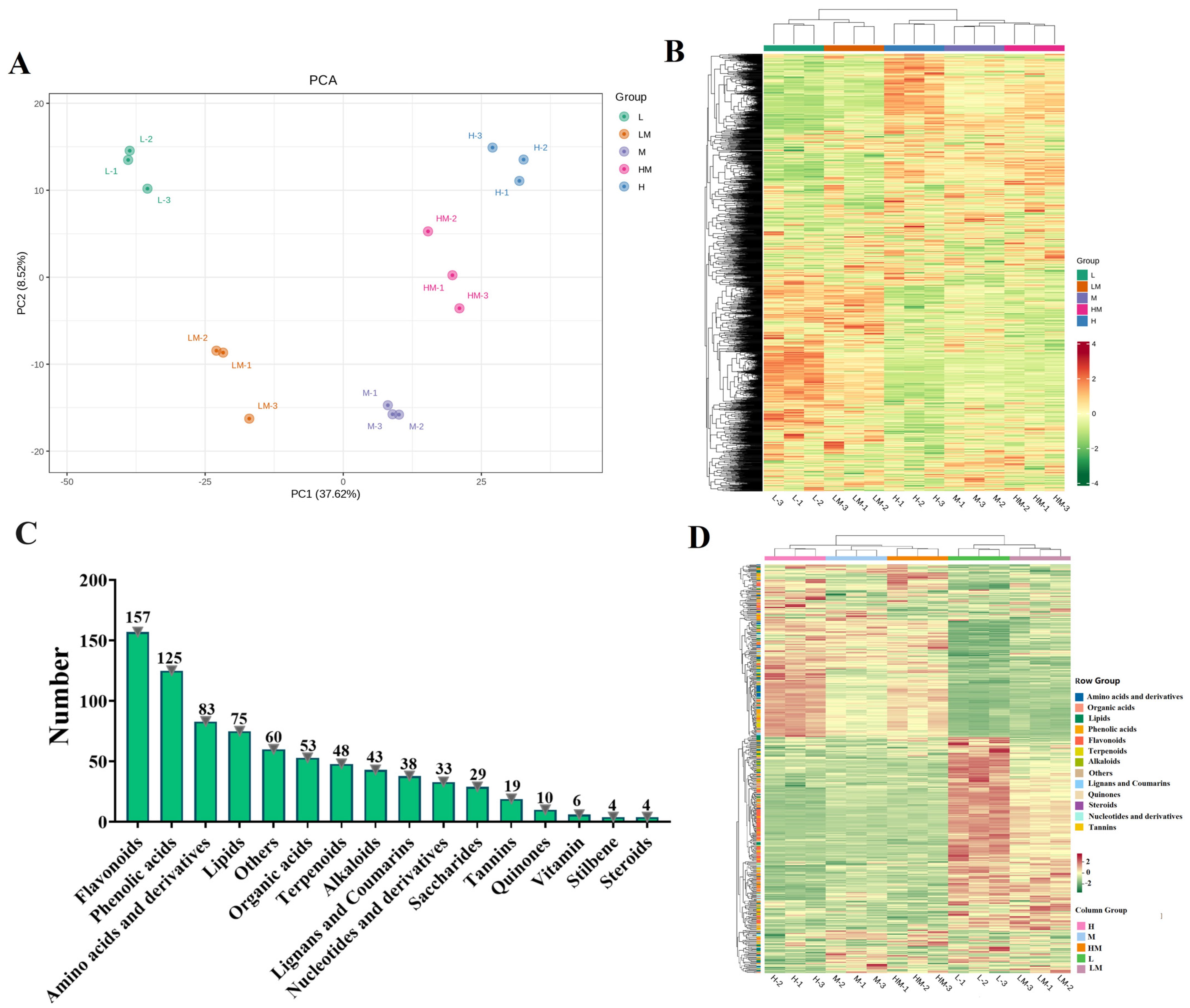
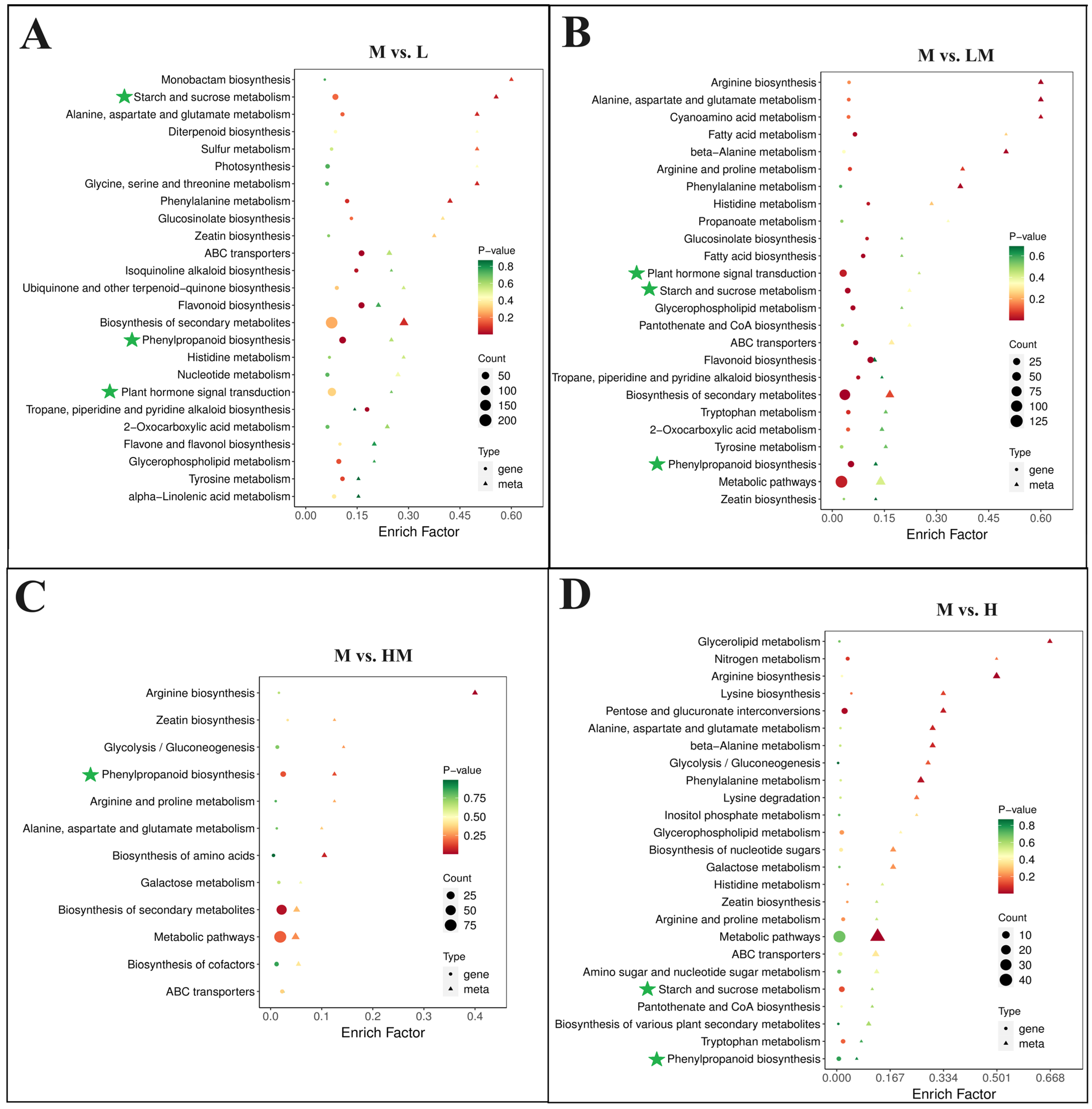
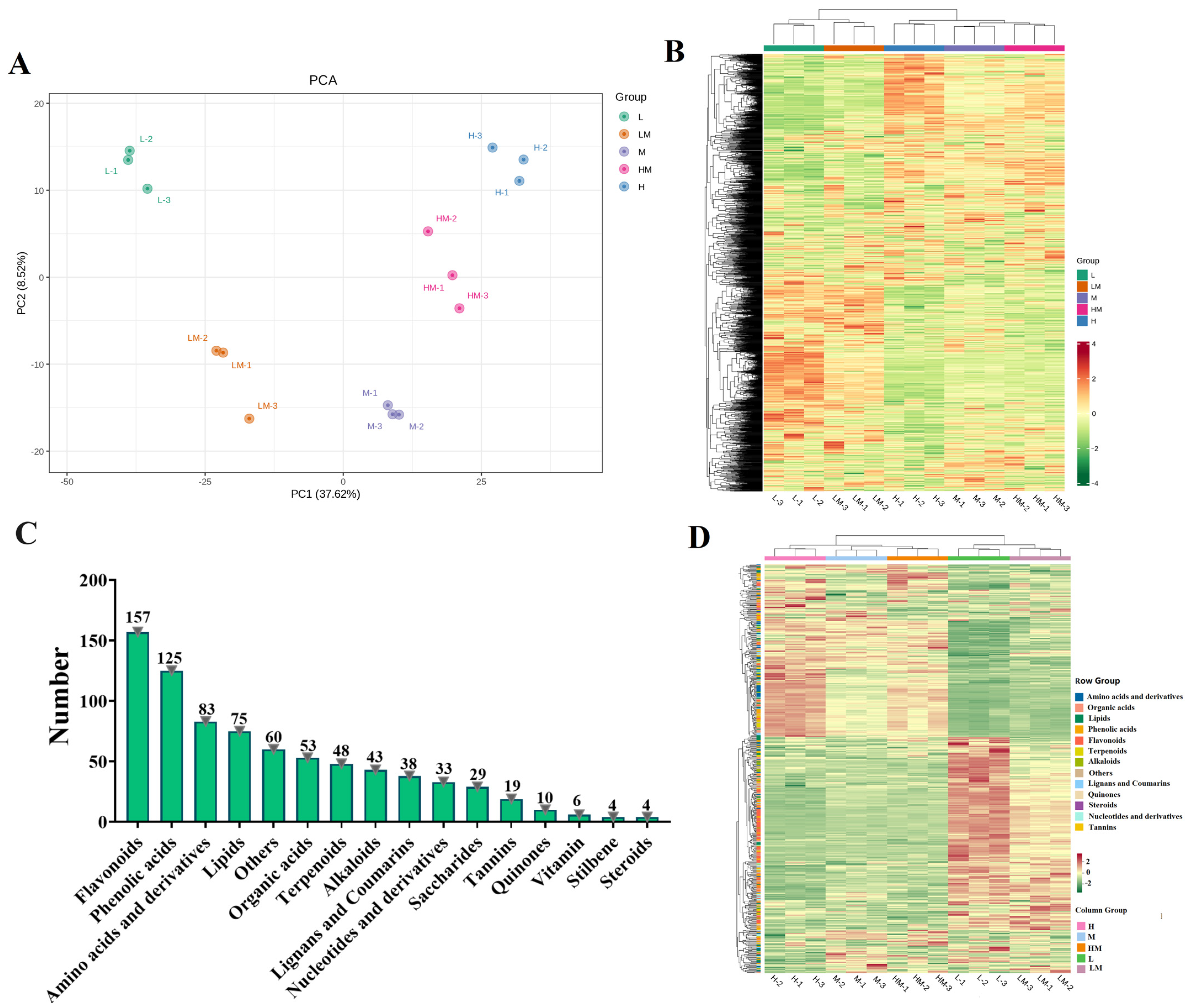
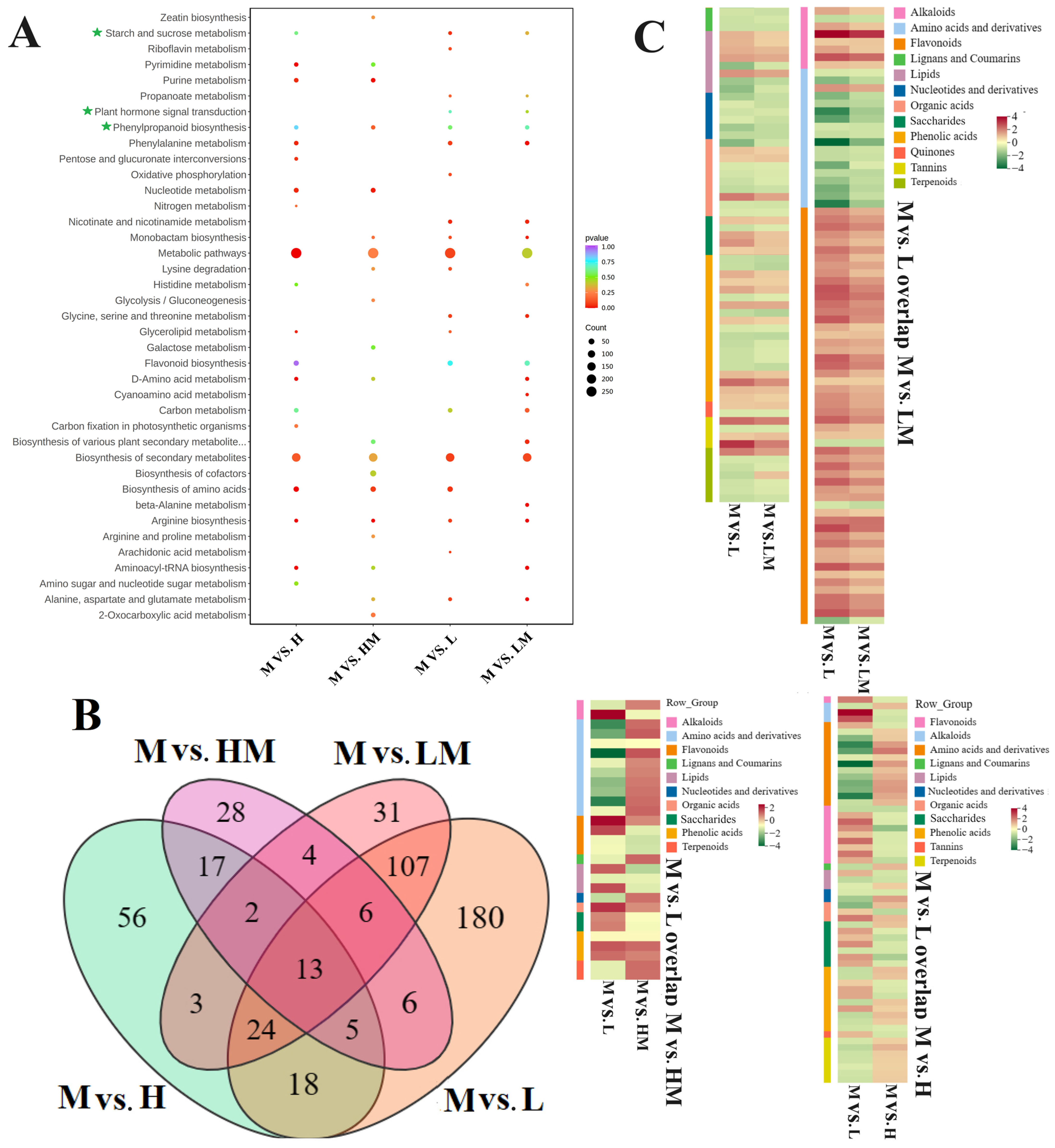
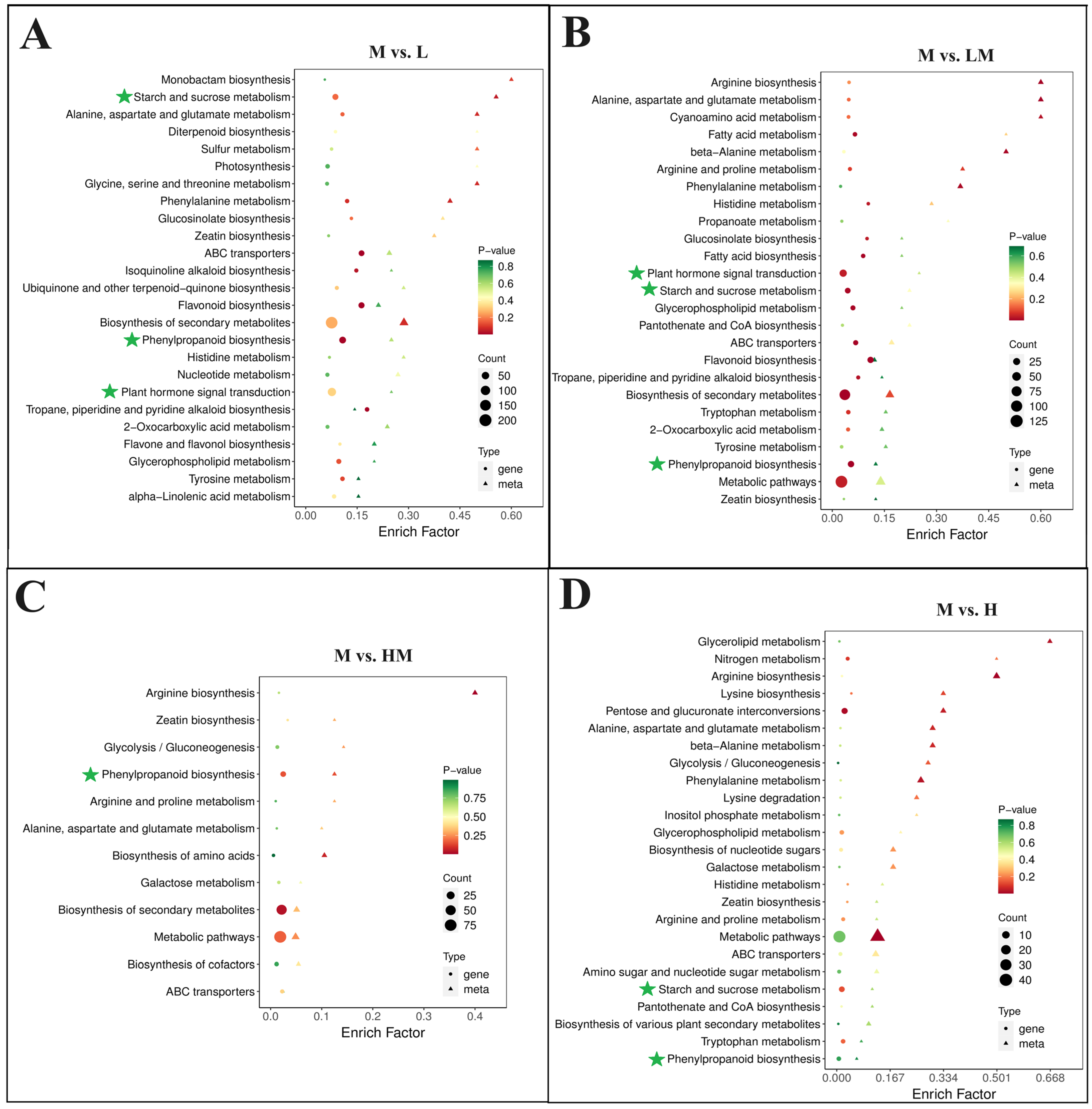
2.4. Metabolome Analysis
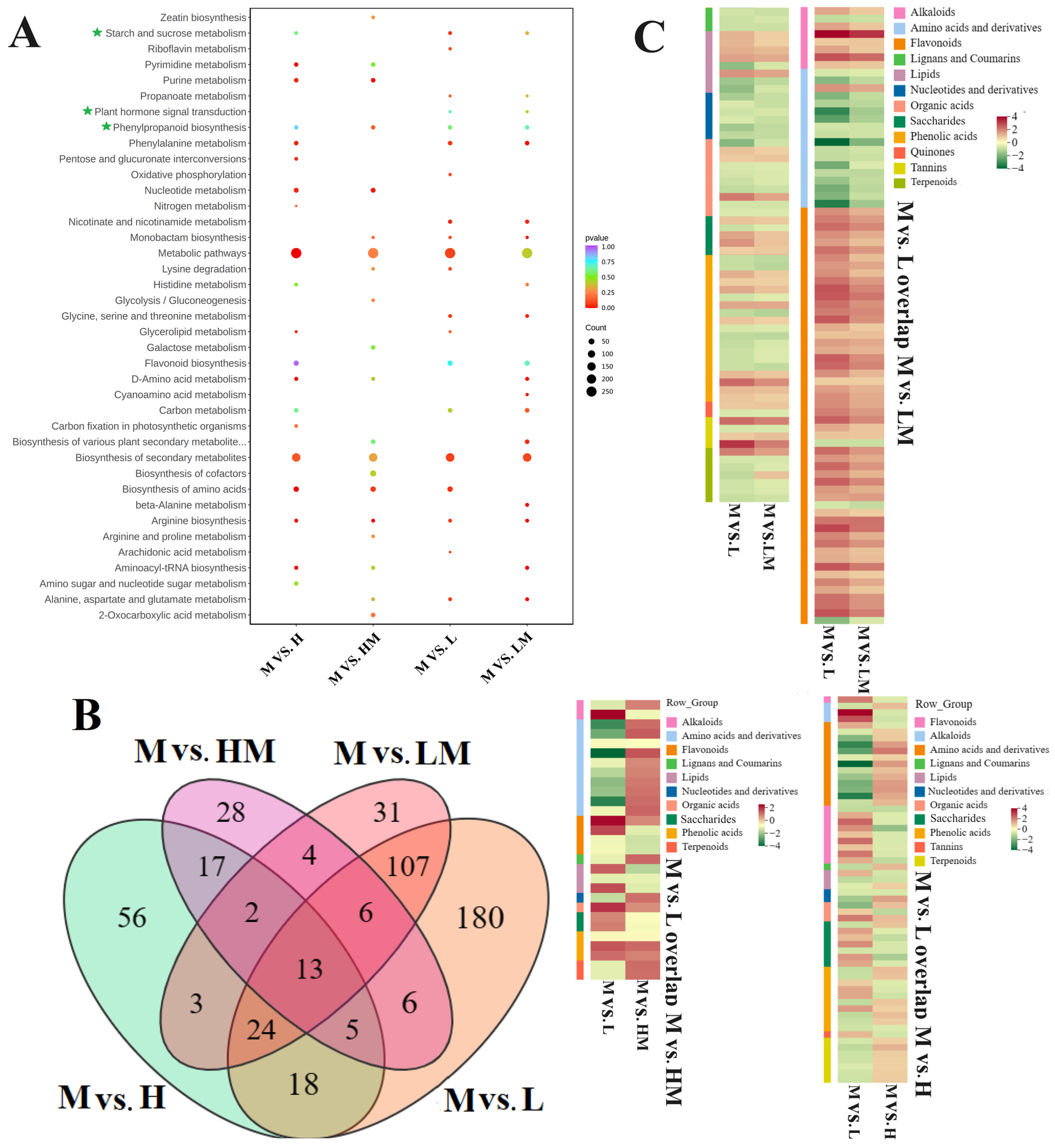
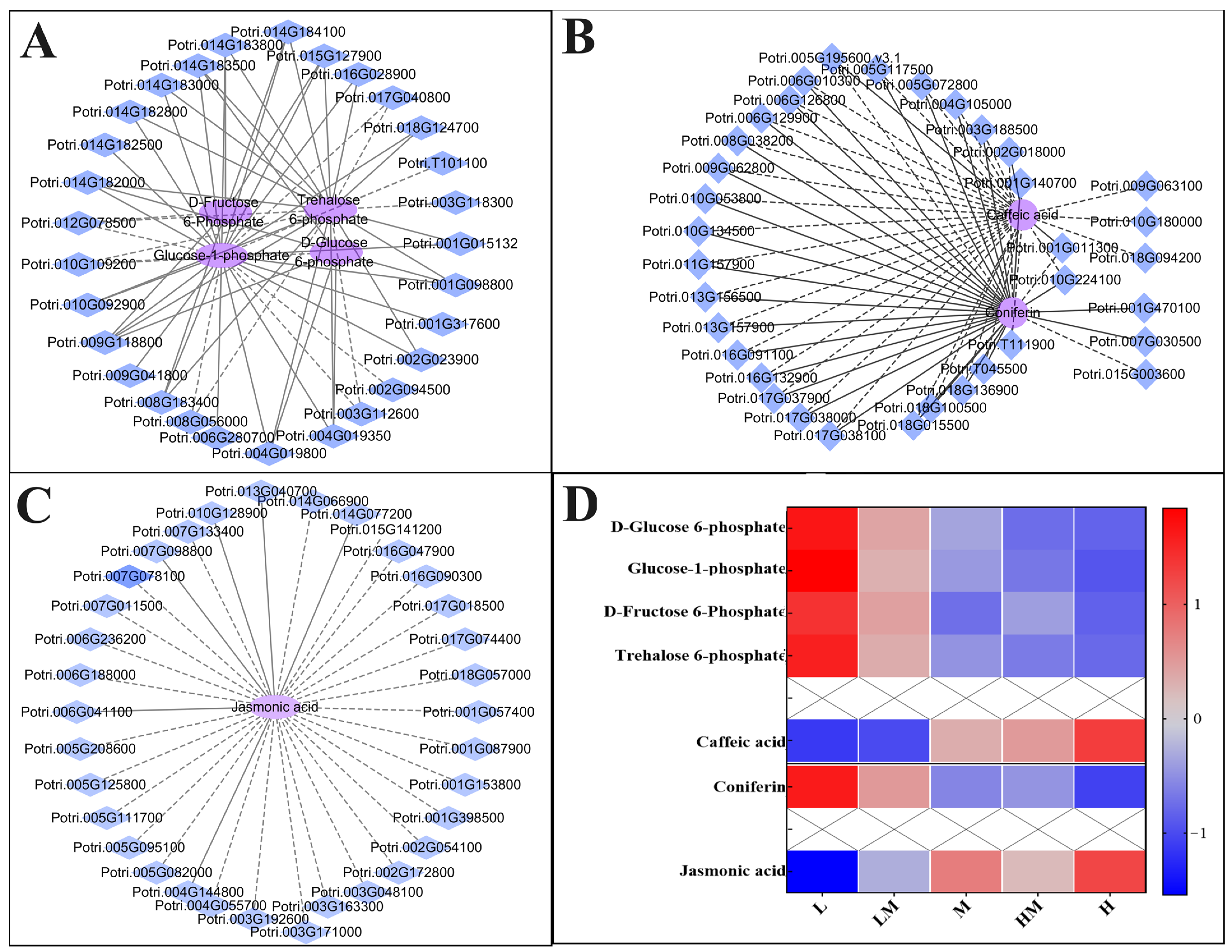
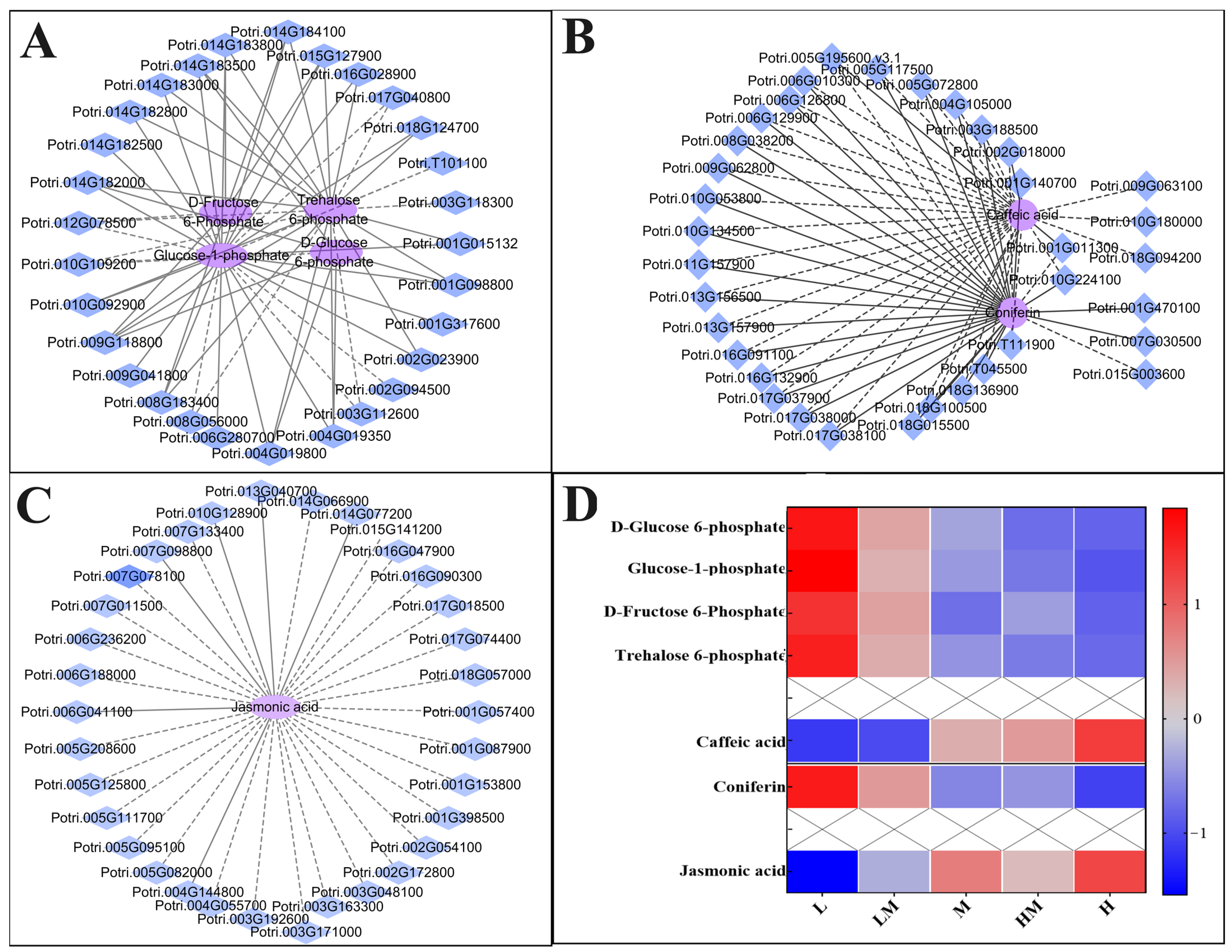
2.5. Conjoint Analysis of Transcriptome and Metabolome
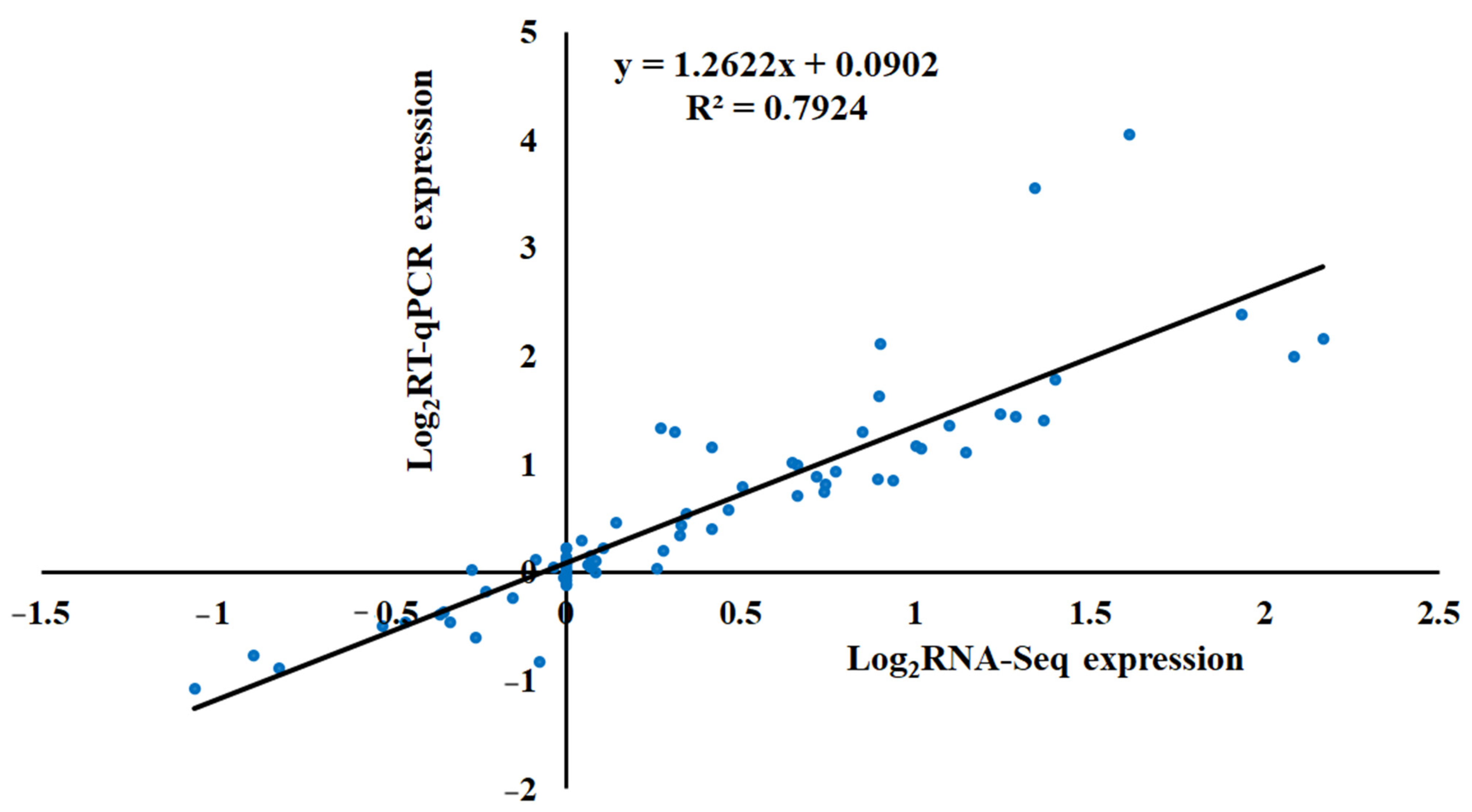
2.6. Correlation between the Results of RT-qPCR and Transcriptome
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Determination of Growth Indices and NH4+ and NO3− Contents
4.3. Wood Composition and Microscopy Assay
4.4. Transcriptome Sequencing and Analysis
4.5. Metabolomics Analysis
4.6. Conjoint Analysis of the Transcriptome and Metabolome
4.7. Real-Time Quantitative PCR (RT-qPCR) Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tan, B.Y.; Xu, M.; Chen, Y.; Huang, M.R. Transient expression for functional gene analysis using Populus protoplasts. Plant Cell Tissue Organ Cult. 2013, 114, 11–18. [Google Scholar] [CrossRef]
- Jansson, S.; Douglas, C.J. Populus: A model system for plant biology. Annu. Rev. Plant. Biol. 2007, 58, 435–458. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, K.; Blomster, T.; Helariutta, Y.; Mahonen, A.P. Vascular cambium development. Arab. Book 2015, 13, e0177. [Google Scholar] [CrossRef]
- Robischon, M.; Du, J.A.; Miura, E.; Groover, A. The Populus class III HD ZIP, popREVOLUTA, influences cambium initiation and patterning of woody stems. Plant Physiol. 2011, 155, 1214–1225. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.Y.; Song, D.L.; Sun, J.Y.; Wang, X.F.; Li, L.G. PtrHB7, a class III HD-Zip gene, plays a critical role in regulation of vascular cambium differentiation in Populus. Mol. Plant 2013, 6, 1331–1343. [Google Scholar] [CrossRef] [PubMed]
- Groover, A.T.; Mansfield, S.D.; DiFazio, S.P.; Dupper, G.; Fontana, J.R.; Millar, R.; Wang, Y. The Populus homeobox gene ARBORKNOX1 reveals overlapping mechanisms regulating the shoot apical meristem and the vascular cambium. Plant Mol. Biol. 2006, 61, 917–932. [Google Scholar] [CrossRef]
- Du, J.; Mansfield, S.D.; Groover, A.T. The Populus homeobox gene ARBORKNOX2 regulates cell differentiation during secondary growth. Plant J. 2009, 60, 1000–1014. [Google Scholar] [CrossRef] [PubMed]
- Etchells, J.P.; Provost, C.M.; Turner, S.R. Plant vascular cell division is maintained by an interaction between PXY and Ethylene Signalling. PLoS Genet. 2012, 8, e1002997. [Google Scholar] [CrossRef]
- Ibañes, M.; Fàbregas, N.; Chory, J.; Caño-Delgado, A.I. Brassinosteroid signaling and auxin transport are required to establish the periodic pattern of Arabidopsis shoot vascular bundles. Proc. Natl. Acad. Sci. USA 2009, 106, 13630–13635. [Google Scholar] [CrossRef]
- Hirakawa, Y.; Kondo, Y.; Fukuda, H. TDIF peptide signaling regulates vascular stem cell proliferation via the WOX4 homeobox gene in Arabidopsis. Plant Cell 2010, 22, 2618–2629. [Google Scholar] [CrossRef]
- Tang, X.F.; Wang, C.P.; Chai, G.H.; Wang, D.A.; Xu, H.; Liu, Y.; He, G.; Liu, S.Q.; Zhang, Y.R.; Kong, Y.Z.; et al. Ubiquitinated DA1 negatively regulates vascular cambium activity through modulating the stability of WOX4 in Populus. Plant Cell 2022, 34, 3364–3382. [Google Scholar] [CrossRef]
- Han, S.; Cho, H.; Noh, J.; Qi, J.Y.; Jung, H.J.; Nam, H.; Lee, S.; Hwang, D.; Greb, T.; Hwang, I. BIL1-mediated MP phosphorylation integrates PXY and cytokinin signalling in secondary growth. Nat. Plants 2018, 4, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Smetana, O.; Makila, R.; Lyu, M.; Amiryousefi, A.; Rodriguez, F.S.; Wu, M.F.; Sole-Gil, A.; Gavarron, M.L.; Siligato, R.; Miyashima, S.; et al. High levels of auxin signalling define the stem-cell organizer of the vascular cambium. Nature 2019, 565, 485–489. [Google Scholar] [CrossRef]
- Nieminen, K.; Immanen, J.; Laxell, M.; Kauppinen, L.; Tarkowski, P.; Dolezal, K.; Tahtiharju, S.; Elo, A.; Decourteix, M.; Ljung, K.; et al. Cytokinin signaling regulates cambial development in poplar. Proc. Natl. Acad. Sci. USA 2008, 105, 20032–20037. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Su, H.L.; Cao, H.; Wei, H.B.; Fu, X.K.; Jiang, X.M.; Song, Q.; He, X.H.; Xu, C.Z.; Luo, K.M. AUXIN RESPONSE FACTOR7 integrates gibberellin and auxin signaling via interactions between DELLA and AUX/IAA proteins to regulate cambial activity in poplar. Plant Cell 2022, 34, 2688–2707. [Google Scholar] [CrossRef]
- Zhu, Y.Y.; Song, D.L.; Zhang, R.; Luo, L.F.; Cao, S.M.; Huang, C.; Sun, J.Y.; Gui, J.S.; Li, L.G. A xylem-produced peptide PtrCLE20 inhibits vascular cambium activity in Populus. Plant Biotechnol. J. 2020, 18, 195–206. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Jiao, Y.; Liu, Z.H.; Zhu, Y.X. ROW1 maintains quiescent centre identity by confining WOX5 expression to specific cells. Nat. Commun. 2015, 6, e6003. [Google Scholar] [CrossRef] [PubMed]
- Gross-Hardt, R.; Laux, T. Stem cell regulation in the shoot meristem. J. Cell Sci. 2003, 116, 1659–1666. [Google Scholar] [CrossRef]
- Schrader, J.; Nilsson, J.; Mellerowicz, E.; Berglund, A.; Nilsson, P.; Hertzberg, M.; Sandberg, G. A high-resolution transcript profile across the wood-forming meristem of poplar identifies potential regulators of cambial stem cell identity. Plant Cell 2004, 16, 2278–2292. [Google Scholar] [CrossRef]
- Prassinos, C.; Ko, J.H.; Han, K.H. Transcriptome profiling of vertical stem segments provides insights into the genetic regulation of secondary growth in hybrid aspen trees. Plant Cell Physiol. 2005, 46, 1213–1225. [Google Scholar] [CrossRef]
- van Raemdonck, D.; Pesquet, E.; Cloquet, S.; Beeckman, H.; Boerjan, W.; Goffner, D.; El Jaziri, M.; Baucher, M. Molecular changes associated with the setting up of secondary growth in aspen. J. Exp. Bot. 2005, 56, 2211–2227. [Google Scholar] [CrossRef] [PubMed]
- Stitt, M. Nitrate regulation of metabolism and growth. Curr. Opin. Plant Biol. 1999, 2, 178–186. [Google Scholar] [CrossRef]
- Paul, M.J.; Foyer, C.H. Sink regulation of photosynthesis. J. Exp. Bot. 2001, 52, 1383–1400. [Google Scholar] [CrossRef] [PubMed]
- Pitre, F.E.; Cooke, J.E.K.; Mackay, J.J. Short-term effects of nitrogen availability on wood formation and fibre properties in hybrid poplar. Trees-Struct. Funct. 2007, 21, 249–259. [Google Scholar] [CrossRef]
- Palenchar, P.M.; Kouranov, A.; Lejay, L.V.; Coruzzi, G.M. Genome-wide patterns of carbon and nitrogen regulation of gene expression validate the combined carbon and nitrogen (CN)-signaling hypothesis in plants. Genome Biol. 2004, 5, R91. [Google Scholar] [CrossRef]
- Mellerowicz, E.J.; Baucher, M.; Sundberg, B.; Boerjan, W. Unravelling cell wall formation in the woody dicot stem. Plant Mol. Biol. 2001, 47, 239–274. [Google Scholar] [CrossRef] [PubMed]
- Moreau, C.; Aksenov, N.; Lorenzo, M.G.; Segerman, B.; Funk, C.; Nilsson, P.; Jansson, S.; Tuominen, H. A genomic approach to investigate developmental cell death in woody tissues of Populus trees. Genome Biol. 2005, 6, R34. [Google Scholar] [CrossRef]
- Boerjan, W.; Ralph, J.; Baucher, M. Lignin biosynthesis. Annu. Rev. Plant Biol. 2003, 54, 519–546. [Google Scholar] [CrossRef]
- Plavcova, L.; Hacke, U.G.; Almeida-Rodriguez, A.M.; Li, E.; Douglas, C.J. Gene expression patterns underlying changes in xylem structure and function in response to increased nitrogen availability in hybrid poplar. Plant Cell Environ. 2013, 36, 186–199. [Google Scholar] [CrossRef]
- Cooke, J.E.K.; Brown, K.A.; Wu, R.; Davis, J.M. Gene expression associated with N-induced shifts in resource allocation in poplar. Plant Cell Environ. 2003, 26, 757–770. [Google Scholar] [CrossRef]
- Fritz, C.; Palacios-Rojas, N.; Feil, R.; Stitt, M. Regulation of secondary metabolism by the carbon-nitrogen status in tobacco: Nitrate inhibits large sectors of phenylpropanoid metabolism. Plant J. 2006, 46, 533–548. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Yu, R.; Li, J.; Wang, K.; Liu, G. Preparation of leaf protoplasts from Populus (Populus × xiaohei T. S. Hwang et Liang) and establishment of transient expression system. J. Plant Physiol. 2023, 291, 154122. [Google Scholar] [CrossRef]
- Hefer, C.A.; Mizrachi, E.; Myburg, A.A.; Douglas, C.J.; Mansfield, S.D. Comparative interrogation of the developing xylem transcriptomes of two wood-forming species: Populus trichocarpa and Eucalyptus grandis. New Phytol. 2015, 206, 1391–1405. [Google Scholar] [CrossRef]
- Kraiser, T.; Gras, D.E.; Gutierrez, A.G.; Gonzalez, B.; Gutierrez, R.A. A holistic view of nitrogen acquisition in plants. J. Exp. Bot. 2011, 62, 1455–1466. [Google Scholar] [CrossRef]
- Canton, F.R.; Suarez, M.F.; Canovas, F.M. Molecular aspects of nitrogen mobilization and recycling in trees. Photosynth. Res. 2005, 83, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Euring, D.; Bai, H.; Janz, D.; Polle, A. Nitrogen-driven stem elongation in poplar is linked with wood modification and gene clusters for stress, photosynthesis and cell wall formation. BMC Plant Biol. 2014, 14, 391. [Google Scholar] [CrossRef]
- Euring, D.; Lofke, C.; Teichmann, T.; Polle, A. Nitrogen fertilization has differential effects on N allocation and lignin in two Populus species with contrasting ecology. Trees-Struct. Funct. 2012, 26, 1933–1942. [Google Scholar] [CrossRef]
- Pitre, F.E.; Lafarguette, F.; Boyle, B.; Pavy, N.; Caron, S.; Dallaire, N.; Poulin, P.L.; Ouellet, M.; Morency, M.J.; Wiebe, N.; et al. High nitrogen fertilization and stem leaning have overlapping effects on wood formation in poplar but invoke largely distinct molecular pathways. Tree Physiol. 2010, 30, 1273–1289. [Google Scholar] [CrossRef]
- Camargo, E.L.; Nascimento, L.C.; Soler, M.; Salazar, M.M.; Lepikson-Neto, J.; Marques, W.L.; Alves, A.; Teixeira, P.J.; Mieczkowski, P.; Carazzolle, M.F.; et al. Contrasting nitrogen fertilization treatments impact xylem gene expression and secondary cell wall lignification in Eucalyptus. BMC Plant Biol. 2014, 14, 256. [Google Scholar] [CrossRef]
- Barnett, J.R. Reactivation of the cambium in Aesculus hippocastanum L.: A transmission Electron Microscope Study. Ann. Bot. 1992, 70, 169–177. [Google Scholar] [CrossRef]
- Yin, Y.F.; Jiang, X.M.; Cui, K.M. Seasonal changes in the ultrastructure of the vascular cambium in shoots of Populus tomentosa. Acta Bot. Sin. 2002, 44, 1268–1277. [Google Scholar]
- Qiu, Z.B.; Wan, L.C.; Chen, T.; Wan, Y.L.; He, X.Q.; Lu, S.F.; Wang, Y.W.; Lin, J.X. The regulation of cambial activity in Chinese fir (Cunninghamia lanceolata) involves extensive transcriptome remodeling. New Phytol. 2013, 199, 708–719. [Google Scholar] [CrossRef]
- Mishima, K.; Fujiwara, T.; Iki, T.; Kuroda, K.; Yamashita, K.; Tamura, M.; Fujisawa, Y.; Watanabe, A. Transcriptome sequencing and profiling of expressed genes in cambial zone and differentiating xylem of Japanese cedar (Cryptomeria japonica). BMC Genom. 2014, 15, 219. [Google Scholar] [CrossRef] [PubMed]
- Weijers, D.; Wagner, D. Transcriptional responses to the auxin hormone. Annu. Rev. Plant Biol. 2016, 67, 539–574. [Google Scholar] [CrossRef] [PubMed]
- De Rybel, B.; Adibi, M.; Breda, A.S.; Wendrich, J.R.; Smit, M.E.; Novák, O.; Yamaguchi, N.; Yoshida, S.; Van Isterdael, G.; Palovaara, J.; et al. Plant development. Integration of growth and patterning during vascular tissue formation in Arabidopsis. Science 2014, 345, 6197. [Google Scholar] [CrossRef]
- Ohashi-Ito, K.; Saegusa, M.; Iwamoto, K.; Oda, Y.; Katayama, H.; Kojima, M.; Sakakibara, H.; Fukuda, H. A bHLH Complex activates vascular cell division via cytokinin action in root apical meristem. Curr. Biol. 2014, 24, 2053–2058. [Google Scholar] [CrossRef] [PubMed]
- Weijers, D.; Schlereth, A.; Ehrismann, J.S.; Schwank, G.; Kientz, M.; Jürgens, G. Auxin triggers transient local signaling for cell specifcation in Arabidopsis embryogenesis. Dev. Cell 2006, 10, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Schrader, J.; Baba, K.; May, S.T.; Palme, K.; Bennett, M.; Bhalerao, R.P.; Sandberg, G. Polar auxin transport in the wood-forming tissues of hybrid aspen is under simultaneous control of developmental and environmental signals. Proc. Natl. Acad. Sci. USA 2003, 100, 10096–10101. [Google Scholar] [CrossRef]
- Nilsson, J.; Karlberg, A.; Antti, H.; Lopez-Vernaza, M.; Mellerowicz, E.; Perrot-Rechenmann, C.; Sandberg, G.; Bhalerao, R.P. Dissecting the molecular basis of the regulation of wood formation by auxin in hybrid aspen. Plant Cell 2008, 20, 843–855. [Google Scholar] [CrossRef]
- Suer, S.; Agusti, J.; Sanchez, P.; Schwarz, M.; Greb, T. WOX4 imparts auxin responsiveness to cambium cells in Arabidopsis. Plant Cell 2011, 23, 3247–3259. [Google Scholar] [CrossRef]
- Kucukoglu, M.; Nilsson, J.; Zheng, B.; Chaabouni, S.; Nilsson, O. WUSCHEL-RELATED HOMEOBOX4 (WOX4)-like genes regulate cambial cell division activity and secondary growth in Populus trees. New Phytol. 2017, 215, 642–657. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Shen, Y.; He, F.; Fu, X.; Yu, H.; Lu, W.; Li, Y.; Li, C.; Fan, D.; Wang, H.C. Auxin-mediated Aux/IAA-ARF-HB signaling cascade regulates secondary xylem development in Populus. New Phytol. 2019, 222, 752–767. [Google Scholar] [CrossRef] [PubMed]
- Guan, C.N.; Xue, Y.; Jiang, P.F.; He, C.C.; Zhuge, X.L.; Lan, T.; Yang, H.L. Overexpression of PtoCYCD3;3 promotes growth and causes leaf wrinkle and branch appearance in Populus. Int. J. Mol. Sci. 2021, 22, 1288. [Google Scholar] [CrossRef] [PubMed]
- Mähönen, A.P.; Higuchi, M.; Törmäkangas, K.; Miyawaki, K.; Pischke, M.S.; Sussman, M.R.; Helariutta, Y.; Kakimoto, T. Cytokinins regulate a bidirectional phosphorelay network in Arabidopsis. Curr. Biol. 2006, 16, 1116–1122. [Google Scholar] [CrossRef] [PubMed]
- Bishopp, A.; Help, H.; El-Showk, S.; Weijers, D.; Scheres, B.; Friml, J.; Benková, E.; Mähönen, A.P.; Helariutta, Y. A mutually inhibitory interaction between auxin and cytokinin specifies vascular pattern in roots. Curr. Biol. 2011, 21, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Funada, R.; Miura, T.; Shimizu, Y.; Kinase, T.; Nakaba, S.; Kubo, T.; Sano, Y. Gibberellin-induced formation of tension wood in angiosperm trees. Planta 2008, 227, 1409–1414. [Google Scholar] [CrossRef]
- Eriksson, M.E.; Israelsson, M.; Olsson, O.; Moritz, T. Increased gibberellin biosynthesis in transgenic trees promotes growth, biomass production and xylem fiber length. Nat. Biotechnol. 2000, 18, 784–788. [Google Scholar] [CrossRef]
- Immanen, J.; Nieminen, K.; Smolander, O.P.; Kojima, M.; Alonso Serra, J.; Koskinen, P.; Zhang, J.; Elo, A.; Mähönen, A.P.; Street, N. Cytokinin and auxin display distinct but Interconnected distribution and signaling profiles to stimulate cambial activity. Curr. Biol. 2016, 26, 1990–1997. [Google Scholar] [CrossRef]
- Jeon, H.W.; Cho, J.S.; Park, E.J.; Han, K.H.; Choi, Y.I.; Ko, J.H. Developing xylem-preferential expression of PdGA20ox1, a gibberellin 20-oxidase 1 from Pinus densiflora, improves woody biomass production in a hybrid poplar. Plant Biotechnol. J. 2016, 14, 1161–1170. [Google Scholar] [CrossRef]
- Israelsson, M.; Sundberg, B.R.; Moritz, T. Tissue-specific localization of gibberellins and expression of gibberellin-biosynthetic and signaling genes in wood-forming tissues in aspen. Plant J. 2010, 44, 494–504. [Google Scholar] [CrossRef]
- Mauriat, M.; Moritz, T. Analyses of GA20ox- and GID1-over-expressing aspen suggest that gibberellins play two distinct roles in wood formation. Plant J. 2009, 58, 989–1003. [Google Scholar] [CrossRef] [PubMed]
- Do; JiHye; Kwon; Mi; Choi; JoonWeon; Shin; SooJeong; Choi; YoungIm, Exogenously applied 24-epi brassinolide reduces lignification and alters cell wall carbohydrate biosynthesis in the secondary xylem of Liriodendron tulipifera. Phytochemistry 2014, 101, 40–51. [CrossRef]
- Noh, S.A.; Choi, Y.I.; Cho, J.S.; Lee, H. The poplar basic helix-loop-helix transcription factor BEE3—Like gene affects biomass production by enhancing proliferation of xylem cells in poplar. Biochem. Biophys. Res. Commun. 2015, 462, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.L.; Tang, R.J.; Wang, H.H.; Jiang, C.M.; Bao, Y.; Yang, Y.; Liang, M.X.; Sun, Z.C.; Kong, F.J.; Li, B. Overexpression of Populus trichocarpa CYP85A3 promotes growth and biomass production in transgenic trees. Plant Biotechnol. J. 2017, 15, 1309–1321. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Gerttula, S.; Li, Z.H.; Zhao, S.T.; Li-Liu, Y.; Liu, Y.; Lu, M.Z.; Groover, A.T. Brassinosteroid regulation of wood formation in poplar. New Phytol. 2020, 225, 1516–1530. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Deng, S.R.; Li, Z.R.; Wu, J.T.; Liu, Q.F.; Liu, W.Z.; Yu, W.J.; Zhang, Y.H.; Shi, W.G.; Zhou, J.; et al. Competing endogenous RNA networks underlying anatomical and physiological characteristics of poplar wood in acclimation to low nitrogen availability. Plant Cell Physiol. 2019, 60, 2478–2495. [Google Scholar] [CrossRef] [PubMed]
- Kraft, C.; Vodermaier, H.C.; Maurer-Stroh, S.; Eisenhaber, F.; Peters, J.M. The WD40 propeller domain of Cdh1 functions as a destruction box receptor for APC/C substrates. Mol. Cell 2005, 18, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Marrocco, K.; Thomann, A.; Parmentier, Y.; Genschik, P.; Criqui, M.C. The APC/C E3 ligase remains active in most post-mitotic Arabidopsis cells and is required for proper vasculature development and organization. Development 2009, 136, 1475–1485. [Google Scholar] [CrossRef]
- Zhang, S.; Cao, L.; Sun, X.; Yu, J.; Xu, X.; Chang, R.; Suo, J.; Liu, G.; Xu, Z.; Qu, C. Genome-wide analysis of UGDH genes in Populus trichocarpa and responsiveness to nitrogen treatment. 3 Biotech 2021, 11, 149. [Google Scholar] [CrossRef]
- Xu, Z.; Ma, J.; Qu, C.; Hu, Y.; Hao, B.; Sun, Y.; Liu, Z.; Yang, H.; Yang, C.; Wang, H.; et al. Identification and expression analyses of the alanine aminotransferase (AlaAT) gene family in poplar seedlings. Sci. Rep. 2017, 7, e45933. [Google Scholar] [CrossRef]
- Ni, J.; Su, S.; Li, H.; Geng, Y.; Zhou, H.; Feng, Y.; Xu, X. Distinct physiological and transcriptional responses of leaves of paper mulberry (Broussonetia kazinoki x B. papyrifera) under different nitrogen supply levels. Tree Physiol. 2020, 40, 667–682. [Google Scholar] [CrossRef] [PubMed]
- Foster, C.E.; Martin, T.M.; Pauly, M. Comprehensive compositional analysis of plant cell walls (lignocellulosic biomass) part I: Lignin. J. Vis. Exp. 2010, 37, e1745. [Google Scholar]
- Foster, C.E.; Martin, T.M.; Pauly, M. Comprehensive compositional analysis of plant cell walls (lignocellulosic biomass) part II: Carbohydrates. J. Vis. Exp. 2010, 37, e1837. [Google Scholar]
- Liu, J.W.; Hai, G.H.; Wang, C.; Cao, S.Q.; Xu, W.J.; Jia, Z.G.; Yang, C.P.; Wang, J.P.; Dai, S.J.; Cheng, Y.X. Comparative proteomic analysis of Populus trichocarpa early stem from primary to secondary growth. J. Proteom. 2015, 126, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Gambino, G.; Perrone, I.; Gribaudo, I. A Rapid and effective method for RNA extraction from different tissues of grapevine and other woody plants. Phytochem. Anal. 2008, 19, 520–525. [Google Scholar] [CrossRef]
- Khan, M.K.; Pandey, A.; Hamurcu, M.; Rajpal, V.R.; Vyhnanek, T.; Topal, A.; Raina, S.N.; Gezgin, S. Insight into the boron toxicity stress-responsive genes in boron-tolerant Triticum dicoccum shoots using RNA sequencing. Agronomy 2023, 13, 631–649. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Database | KEGG | Nr | Swissport | TrEMBL | KOG | GO | Pfam | Total |
|---|---|---|---|---|---|---|---|---|
| Number | 21,624 | 36,551 | 27,253 | 37,756 | 19,089 | 31,083 | 29,829 | 38,285 |
| Group | M vs. L | M vs. LM | M vs. HM | M vs. H |
|---|---|---|---|---|
| Up | 195 | 120 | 40 | 67 |
| Down | 164 | 70 | 41 | 71 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Cao, L.; Chang, R.; Zhang, H.; Yu, J.; Li, C.; Liu, G.; Yan, J.; Xu, Z. Network Analysis of Metabolome and Transcriptome Revealed Regulation of Different Nitrogen Concentrations on Hybrid Poplar Cambium Development. Int. J. Mol. Sci. 2024, 25, 1017. https://doi.org/10.3390/ijms25021017
Zhang S, Cao L, Chang R, Zhang H, Yu J, Li C, Liu G, Yan J, Xu Z. Network Analysis of Metabolome and Transcriptome Revealed Regulation of Different Nitrogen Concentrations on Hybrid Poplar Cambium Development. International Journal of Molecular Sciences. 2024; 25(2):1017. https://doi.org/10.3390/ijms25021017
Chicago/Turabian StyleZhang, Shuang, Lina Cao, Ruhui Chang, Heng Zhang, Jiajie Yu, Chunming Li, Guanjun Liu, Junxin Yan, and Zhiru Xu. 2024. "Network Analysis of Metabolome and Transcriptome Revealed Regulation of Different Nitrogen Concentrations on Hybrid Poplar Cambium Development" International Journal of Molecular Sciences 25, no. 2: 1017. https://doi.org/10.3390/ijms25021017
APA StyleZhang, S., Cao, L., Chang, R., Zhang, H., Yu, J., Li, C., Liu, G., Yan, J., & Xu, Z. (2024). Network Analysis of Metabolome and Transcriptome Revealed Regulation of Different Nitrogen Concentrations on Hybrid Poplar Cambium Development. International Journal of Molecular Sciences, 25(2), 1017. https://doi.org/10.3390/ijms25021017