
Association of the rs1126616 and rs9138 Variants in the SPP1 Gene among Mexican Patients with Systemic Lupus Erythematosus and Lupus Nephritis


 ,
,  ,
,
Abstract
1. Introduction
2. Results
2.1. Demographic and Clinical Characteristics
2.2. Analyses of the Variants
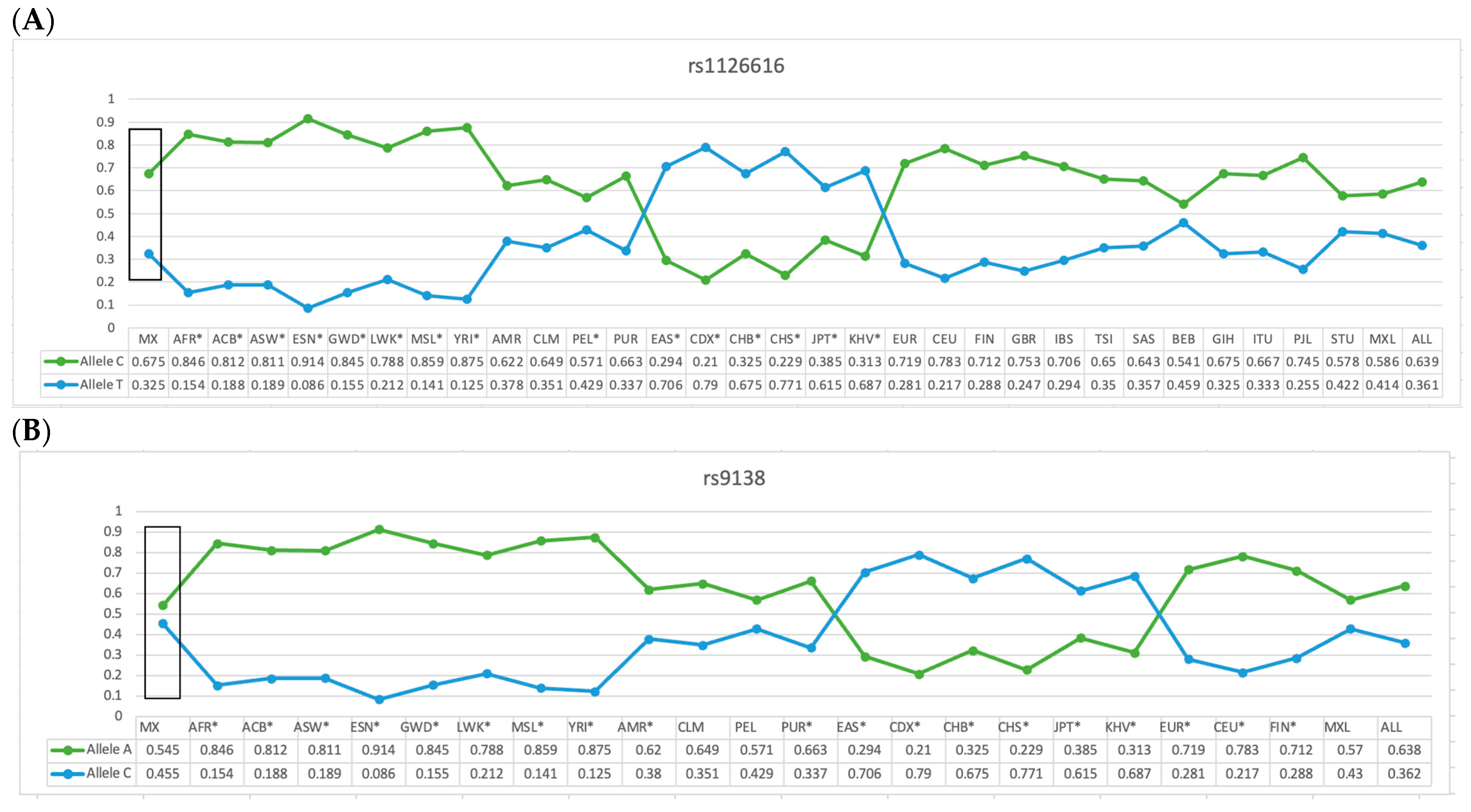
2.3. Comparative Analysis of the Allelic Frequency of the rs11267616 and rs9138 Variants of the SPP1 Gene in the Mexican-Mestizo Population (Reference Group with Different Populations)
2.4. Genotype Distribution of the rs1126616 and rs9138 Variants of the SPP1 Gene in SLE Patients, Stratified by LN
2.5. Comparative Analysis between Patients with SLE Stratified by LN with the Reference Group for the rs1126616 Variant of the SPP1 Gene
2.6. Comparative Analysis between Patients with SLE with LN and SLE without LN for the rs1126616 and rs9138 Variants of the SPP1 Gene
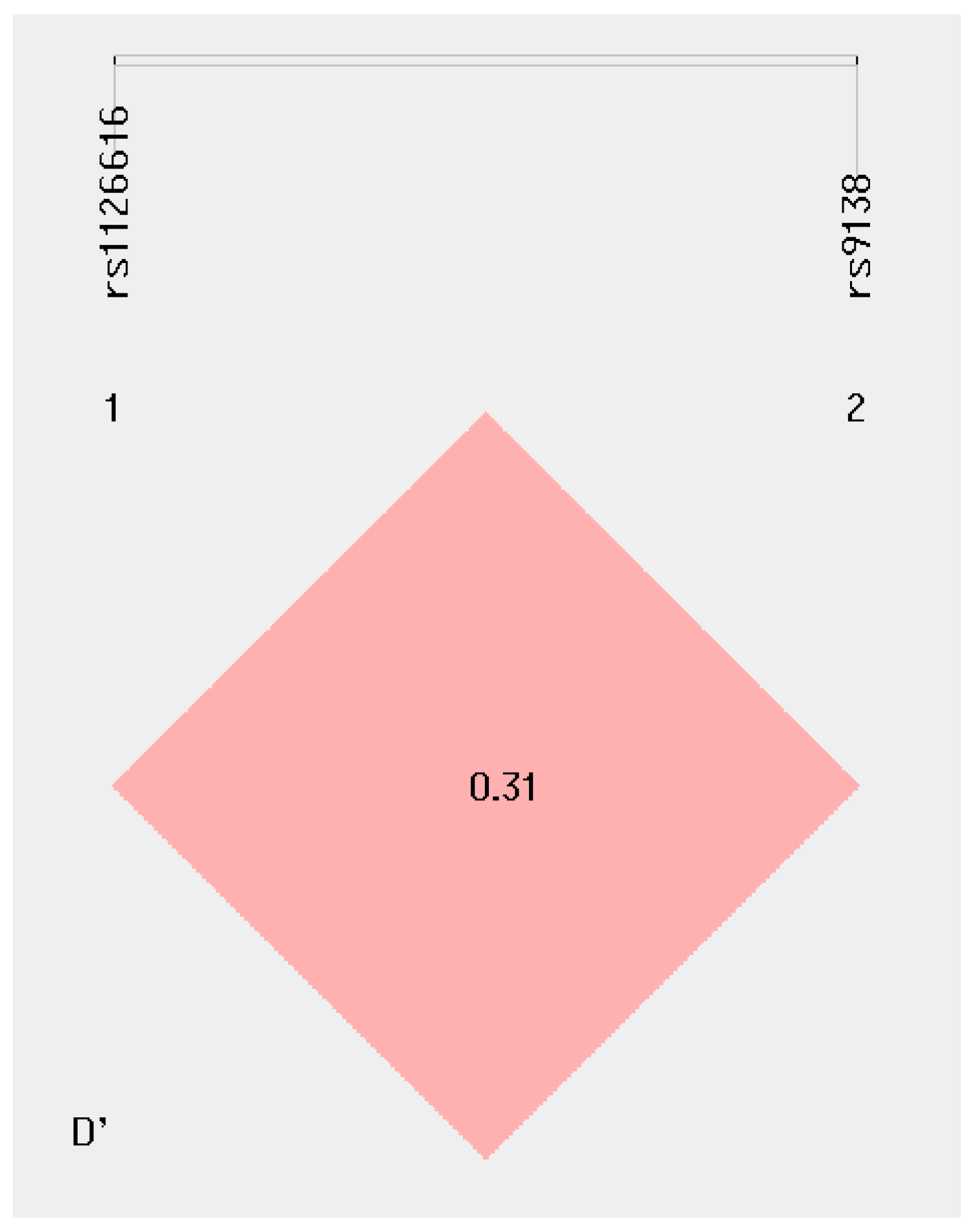
2.7. Haplotype Analyses of rs1126616 and rs9138 Variants of the SPP1 Gene in the Study’s Groups
3. Discussion
4. Materials and Methods
Variant Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anders, H.-J.; Saxena, R.; Zhao, M.-H.; Parodis, I.; Salmon, J.E.; Mohan, C. Lupus nephritis. Nat. Rev. Dis. Primers 2020, 6, 1–25. [Google Scholar] [CrossRef]
- Guía de la Practica CENETEC. Lupus Eritematoso Sistémico. 2015. Available online: https://cenetec-difusion.com/CMGPC/IMSS-752-15/ER.pdf (accessed on 13 November 2023).
- Sinave, V.E. Protocolo del Sistema Nacional de Vigilancia Epidemiológica SINAVE. 2018. Available online: https://www.gob.mx/cms/uploads/attachment/file/211946/PAE_2013-2018.pdf (accessed on 13 November 2023).
- Gaviria-Garcia, G.; Maidana de Zarza, A.; Aroca-Martinez, G. Características sociodemográficas y clínicas de pacientes con nefritis lúpica. Barranquilla, Colombia. Mem. Inst. Investig. Cienc. Salud 2018, 16, 32–37. [Google Scholar] [CrossRef]
- Briones-Orta, M.A.; Avendaño-Vázquez, S.E.; Aparicio-Bautista, D.I.; Coombes, J.D.; Weber, G.F.; Syn, W.-K. Osteopontin splice variants and polymorphisms in cancer progression and prognosis. Biochim. Biophys. Acta Rev. Cancer 2017, 1868, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Garavito, G.; Egea, E.; Fang, L.; Malagón, C.; Olmos, C.; González, L.; Guarnizo, P.; Aroca, G.; López, G.; Iglesias, A. Association of polymorphic variants of PTPN22, TNF and VDR systems in children with lupus nephritis: A study in trios of Colombian families. Biomedica 2017, 37, 260–266. [Google Scholar] [CrossRef]
- Chauhan, A.; Sun, Y.; Sukumaran, P.; Zangbede, F.O.Q.; Jondle, C.N.; Sharma, A.; Evans, D.L.; Chauhan, P.; Szlabick, R.E.; Aaland, M.O.; et al. M1 macrophage polarization is dependent on TRPC1-mediated calcium entry. iScience 2018, 8, 85–102. [Google Scholar] [CrossRef]
- Icer, M.A.; Gezmen-Karadag, M. The multiple functions and mechanisms of osteopontin. Clin. Biochem. 2018, 59, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Kariuki, S.N.; Moore, J.G.; Kirou, K.A.; Crow, M.K.; Utset, T.O.; Niewold, T.B. Age- and gender-specific modulation of serum osteopontin and interferon-α by osteopontin genotype in systemic lupus erythematosus. Genes Immun. 2009, 10, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Lampe, M.A.; Patarca, R.; Iregui, M.V.; Cantor, H. Polyclonal B cell activation by the Eta-1 cytokine and the development of systemic autoimmune disease. J. Immunol. 1991, 147, 2902–2906. [Google Scholar] [CrossRef]
- Rangaswami, H.; Bulbule, A.; Kundu, G.C. Osteopontin: Role in cell signaling and cancer progression. Trends Cell Biol. 2006, 16, 79–87. [Google Scholar] [CrossRef]
- D’Alfonso, S.; Barizzone, N.; Giordano, M.; Chiocchetti, A.; Magnani, C.; Castelli, L.; Indelicato, M.; Giacopelli, F.; Marchini, M.; Scorza, R.; et al. Two single-nucleotide polymorphisms at the 50 and 30 ends of the osteopontin gene contributes to susceptibility to systemic lupus erythematosus. Arthritis Rheum. 2005, 52, 539–547. [Google Scholar] [CrossRef]
- Cantor, H. The role of eta-1/osteopontin in the pathogenesis of immunological disorders. Ann. N. Y. Acad. Sci. 1995, 760, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Carecchio, M.; Comi, C. The role of osteopontin in neurodegenerative diseases. J. Alzheimers Dis. 2011, 25, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Frenzel, D.F.; Weiss, J.M. Osteopontin and allergic disease: Pathophysiology and implications for diagnoses and therapy. Expert Rev. Clin. Immunol. 2011, 7, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Guthridge, J.M.; Harley, I.T.; Sestak, A.L.; Kim-Howard, X.; Kaufman, K.M.; Namjou, B.; Deshmukh, H.; Bruner, G.; Espinoza, L.R.; et al. Osteopontin and systemic lupus erythematosus association: A probable gene-gender interaction. PLoS ONE 2008, 3, e0001757. [Google Scholar] [CrossRef] [PubMed]
- Kaleta, B. The role of osteopontin in kidney diseases. Inflamm. Res. 2019, 68, 93–102. [Google Scholar] [CrossRef]
- Martín-Márquez, B.T.; Sandoval-García, F.; Corona-Meraz, F.I.; Petri, M.H.; Gutiérrez-Mercado, Y.K.; Vázquez-Del Mercado, M. Osteopontin: Another piece in the systemic lupus erythematosus immunopathology puzzle. Clin. Exp. Rheumatol. 2022, 40, 173–182. [Google Scholar] [CrossRef]
- Salimi, S.; Noora, M.; Nabizadeh, S.; Rezaei, M.; Shahraki, H.; Milad, M.K.; Naghavi, A.; Farajian-Mashhadi, F.; Zakeri, Z.; Sandoughi, M. Association of the osteopontin rs1126616 polymorphism and a higher serum osteopontin level with lupus nephritis. Biomed. Rep. 2016, 4, 355–360. [Google Scholar] [CrossRef]
- Xu, A.P.; Bai, J.; Lü, J.; Liang, Y.Y.; Li, J.G.; Lai, D.Y.; Wan, X.; Huang, H.H. Osteopontin gene polymorphism in association with systemic lupus erythematosus in Chinese patients. Chin. Med. J. 2007, 120, 2124–2128. [Google Scholar] [CrossRef]
- Forton, A.C.; Petri, M.A.; Goldman, D.; Sullivan, K.E. An osteopontin (SPP1) polymorphism is associated with systemic lupus erythematosus. Hum. Mutat. 2002, 19, 459. [Google Scholar] [CrossRef]
- Metwally, R.M.; Hasan, A.S. Association of Osteopontin gene single nucleotide polymorphism with lupus nephritis. Int. J. Rheum. Dis. 2022, 25, 571–575. [Google Scholar] [CrossRef]
- Trivedi, T.; Franek, B.S.; Green, S.L.; Kariuki, S.N.; Kumabe, M.; Mikolaitis, R.A.; Jolly, M.; Utset, T.O.; Niewold, T.B. Osteopontin alleles are associated with clinical characteristics in systemic lupus erythematosus. J. Biomed. Biotechnol. 2011, 2011, 802581. [Google Scholar] [CrossRef] [PubMed]
- Comi, C.; Cappellano, G.; Chiocchetti, A.; Orilieri, E.; Buttini, S.; Ghezzi, L.; Galimberti, D.; Guerini, F.; Barizzone, N.; Perla, F.; et al. The impact of osteopontin gene variations on multiple sclerosis development and progression. Clin. Dev. Immunol. 2012, 2012, 212893. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.M.; Buckenberger, J.A.; Jiang, J.; Malana, G.E.; Nuovo, G.J.; Chotani, M.; Feldman, D.S.; Schmittgen, T.D.; Elton, T.S. The human angiotensin II type 1 receptor +1166 A/C polymorphism attenuates microRNA-155 binding. J. Biol. Chem. 2013, 288, 4227. [Google Scholar] [CrossRef]
- Grillo, G.; Turi, A.; Licciulli, F.; Mignone, F.; Liuni, S.; Banfi, S.; Gennarino, V.A.; Horner, D.S.; Pavesi, G.; Picardi, E.; et al. UTRdb and UTRsite (RELEASE 2010): A collection of sequences and regulatory motifs of the untranslated regions of eukaryotic mRNAs. Nucleic Acids Res. 2010, 38, D75–D80. [Google Scholar] [CrossRef] [PubMed]
- Pons-Estel, G.J.; Catoggio, L.J.; Cardiel, M.H.; Bonfa, E.; Caeiro, F.; Sato, E.; Massardo, L.; Molina-Restrepo, J.F.; Toledano, M.G.; Barile-Fabris, L.A.; et al. Lupus in Latin-American patients: Lessons from the GLADEL cohort. Lupus 2015, 24, 536–545. [Google Scholar] [CrossRef]
- Xipell, M.; Lledó, G.M.; Egan, A.C.; Tamirou, F.; Del Castillo, C.S.; Rovira, J.; Gómez-Puerta, J.A.; García-Herrera, A.; Cervera, R.; Kronbichler, A.; et al. From systemic lupus erythematosus to lupus nephritis: The evolving road to targeted therapies. Autoimmun. Rev. 2023, 22, 103404. [Google Scholar] [CrossRef]
- Yen, E.Y.; Singh, R.R. Brief report: Lupus-an unrecognized leading cause of death in young females: A population-based study using nationwide death certificates, 2000–2015. Arthritis Rheumatol. 2018, 70, 1251–1255. [Google Scholar] [CrossRef]
- Kaleta, B. Role of osteopontin in systemic lupus erythematosus. Arch. Immunol. Ther. Exp. 2014, 62, 475–482. [Google Scholar] [CrossRef]
- Chabanon, H.; Mickelburgh, I.; Hesketh, J. Zipcodes and postage stamps: mRNA localization signals and their trans-acting binding proteins. Brief. Funct. Genom. 2004, 3, 240–256. [Google Scholar]
- Conne, B.; Stutz, A.; Vassalli, J.D. The 39 untranslated region of messenger RNA: A molecular “hotspot” for pathology? Nat. Med. 2000, 6, 637–641. [Google Scholar] [CrossRef]
- Gimba, E.R.; Tilli, T.M. Human osteopontin splicing isoforms: Known roles, potential clinical applications and activated signaling pathways. Cancer Lett. 2013, 331, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, Y.; Mori, K.; Hara, T.; Tanaka, K.; Murakami, M.; Uede, T. Functional analysis of the osteopontin molecule. Ann. N. Y. Acad. Sci. 1995, 760, 371–374. [Google Scholar] [CrossRef] [PubMed]
- Tekin, G.; Ertan, P.; Horasan, G.; Berdeli, A. SPP1 gene polymorphisms associated with nephrolithiasis in Turkish pediatric patients. Urol. J. 2012, 9, 640–647. [Google Scholar] [PubMed]
- Kaleta, B.; Lachota, M.; Łukaszkiewicz, J.; Woźniacka, A.; Bogaczewicz, J. Osteopontin gene polymorphisms rs1126616 C>T and rs1126772 A>G are associated with atopic dermatitis in Polish population. Appl. Clin. Genet. 2021, 14, 417–425. [Google Scholar] [CrossRef]
- Lund, S.A.; Giachelli, C.M.; Scatena, M. The role of osteopontin in inflammatory processes. J. Cell Commun. Signal. 2009, 3, 311–322. [Google Scholar] [CrossRef]
- Silva, D.; Pollett, A.P.; Rittling, A.; Denhardt, S.R.; Sodek, D.T.; Zohar, J. Exacerbated tissue destruction in DSS-induced acute colitis of OPN-null mice is associated with downregulation of TNF-alpha expression and non-programmed cell death. J. Cell Physiol. 2006, 208, 629–639. [Google Scholar] [CrossRef]
- Zhao, J.; Jing, J.; Zhao, W.; Li, X.; Hou, L.; Zheng, C.; Kong, Q.; Li, W.; Yao, X.; Chang, L.; et al. Osteopontin exacerbates the progression of experimental autoimmune myasthenia gravis by affecting the differentiation of T cell subsets. Int. Immunopharmacol. 2020, 82, 106335. [Google Scholar] [CrossRef]
- Safarinejad, M.R.; Shafiei, N.; Safarinejad, S. Association between polymorphisms in osteopontin gene (SPP1) and first episode calcium oxalate urolithiasis. Urolithiasis 2013, 41, 303–313. [Google Scholar] [CrossRef]
- Fan, K.; Dai, J.; Wang, H.; Wei, H.; Cao, Z.; Hou, S.; Qian, W.; Wang, H.; Li, B.; Zhao, J.; et al. Treatment of collagen-induced arthritis with an anti-osteopontin monoclonal antibody through promotion of apoptosis of both murine and human activated T cells. Arthritis Rheum. 2008, 58, 2041–2052. [Google Scholar] [CrossRef]
- Lee, Y.H.; Song, G.G. Associations Between Osteopontin Variants and Systemic Lupus Erythematosus: A Meta-Analysis. Genet. Test. Mol. Biomark. 2023, 27, 277–283. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| SLE without LN (n = 111) | SLE with LN (n = 60) | Reference Group (n = 100) | |
|---|---|---|---|
| Age in years, mean (range) | 42 (18–82) | 42 (18–82) | 41 (18–80) |
| Gender n, (%) | 111 (100) | 60 (100) | 100 (100) |
| Female n (%) | 98 (88.28) | 48 (80) | 50 (50%) |
| Male n (%) | 13 (11.71) | 12 (20) | 50 (50%) |
| Clinical Criteria EULAR/ACR 2019 * | |||
| Constitutional: | |||
| Fever n (%) | 1 (0.90) | 3 (5) | - |
| Mucocutaneous n (%) | 80 (72.07) | 41 (68.33) | - |
| Arthritis n (%) | 67 (60.3) | 50 (83.33) | - |
| Neuropsychiatric n (%) | 2 (1.80) | 5 (8.3) | - |
| Serositis n (%) | 30 (27.02) | 20 (33.33) | - |
| Hematological n (%) | 62 (55.85) | 49 (81.66) | - |
| Asymptomatic n (%) | 28 (25.22) | 10 (16.66) | - |
| Variants | SLE * | SLE with NL ** | SLE without NL ** | Reference Group * | OR * | 95%(CI) * | p Value * | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| rs1126616 | Genotype | (n = 171) | % | (n = 60) | % | (n = 111) | % | (n = 100) | % | |||
| Model | CC | −82 | 48 | −23 | 39 | −59 | 53 | −46 | 46 | |||
| CT | −52 | 30 | −17 | 28 | −35 | 32 | −43 | 43 | 1 | |||
| Model | TT | −37 | 22 | −20 | 33 | −17 | 15 | −11 | 11 | 0.57 | (0.34–0.96) | 0.049 |
| CC | −82 | 48 | −23 | 39 | −59 | 53 | −46 | 46 | 2.23 | (1.08–4.61) | 0.04 | |
| Dominant | CT+TT | −89 | 52 | −37 | 61 | −52 | 47 | −54 | 54 | |||
| TT | −37 | 22 | −20 | 33 | −17 | 15 | −11 | 11 | 0.92 | (0.56–1.57) | 0.756 | |
| Recessive | CC+CT | −134 | 78 | −40 | 67 | −94 | 85 | −89 | 89 | 2.23 | (1.08–4.61) | 0.04 |
| Alleles (2n = 342) | (2n = 120) | (2n = 222) | (2n = 200) | |||||||||
| C | −216 | 0.6315 | −63 | 0.525 | −153 | 0.689 | −135 | 0.675 | 0.82 | (0.57–1.19) | 0.353 | |
| T | −126 | 0.3685 | −57 | 0.475 | −69 | 0.311 | −65 | 0.325 | 1.21 | (0–83–1.75) | 0.353 | |
| rs9138 | (n = 100) | % | ||||||||||
| AA | −38 | 22 | −17 | 28 | −28 | 25 | −30 | 30 | 1 | |||
| AC | −96 | 56 | −34 | 57 | −62 | 56 | −49 | 49 | 1.33 | (0.81–2.18) | 0.312 | |
| CC | −37 | 22 | −9 | 15 | −21 | 19 | −21 | 21 | 1.03 | (0.56–1.89) | 1 | |
| Dominant | AA | −38 | 22 | −38 | 22 | −30 | 30 | −30 | 30 | |||
| AC+CC | −133 | 78 | −133 | 78 | −70 | 70 | −70 | 70 | 1.5 | (0.86–2.62) | 0.154 | |
| Recessive | CC | −37 | 22 | −37 | 22 | −21 | 21 | −21 | 21 | 1.03 | (0.56–1.89) | 1 |
| AC+CC | −134 | 78 | −134 | 78 | −79 | 79 | −79 | 79 | ||||
| Alleles | (2n = 342) | (2n = 222) | (2n = 200) | |||||||||
| A | −172 | 0.5029 | −68 | 0.566 | −118 | 0.5315 | −109 | 0.545 | 0.84 | (0.59–1.19) | 0.391 | |
| C | −170 | 0.4971 | −52 | 0.434 | −104 | 0.4685 | −91 | 0.455 | 1.18 | (0.83–1.67) | 0.391 | |
| Haplotype | SLE (2n = 342) | Reference Group (2n = 200) | |||||
|---|---|---|---|---|---|---|---|
| rs1126616 | rs9138 | n | % | n | % | OR 95% (CI) | p-Value |
| T | C | (82) | 24 | (41) | 21 | 1.2 (0.80–1.8) | 0.73 |
| T | A | (44) | 13 | (24) | 12 | 1.0 (0.63–1.8) | 0.87 |
| C | A | (128) | 37 | (85) | 43 | 0.8 (0.56–1.1) | 0.28 |
| C | C | (88) | 26 | (50) | 25 | 1.0 (0.69–1.5) | 0.93 |
| Haplotype | SLE with LN (2n = 120) | SLE without LN (2n = 222) | |||||
|---|---|---|---|---|---|---|---|
| rs1126616 | rs9138 | n | % | n | % | OR 95% (CI) | p-Value |
| T | C | (32) | 27 | (50) | 23 | 1.2 (0.74–2.0) | 0.39 |
| T | A | (25) | 21 | (19) | 9 | 2.8 (1.4–5.3) | 0.002 |
| C | A | (43) | 36 | (85) | 38 | 0.9 (0.56–1.4) | 0.72 |
| C | C | (20) | 16 | (68) | 30 | 0.4 (0.25–0.79) | 0.004 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera-Cameras, A.; Gallegos-Arreola, M.P.; Morán-Moguel, M.C.; Salazar-Páramo, M.; Alcaraz-López, M.F.; Echeverría-González, G.; Topete-Reyes, J.F.; Franco-Chávez, S.A.; Dávalos-Rodríguez, I.P. Association of the rs1126616 and rs9138 Variants in the SPP1 Gene among Mexican Patients with Systemic Lupus Erythematosus and Lupus Nephritis. Int. J. Mol. Sci. 2024, 25, 1000. https://doi.org/10.3390/ijms25021000
Rivera-Cameras A, Gallegos-Arreola MP, Morán-Moguel MC, Salazar-Páramo M, Alcaraz-López MF, Echeverría-González G, Topete-Reyes JF, Franco-Chávez SA, Dávalos-Rodríguez IP. Association of the rs1126616 and rs9138 Variants in the SPP1 Gene among Mexican Patients with Systemic Lupus Erythematosus and Lupus Nephritis. International Journal of Molecular Sciences. 2024; 25(2):1000. https://doi.org/10.3390/ijms25021000
Chicago/Turabian StyleRivera-Cameras, Alicia, Martha Patricia Gallegos-Arreola, María Cristina Morán-Moguel, Mario Salazar-Páramo, Miriam Fabiola Alcaraz-López, Gustavo Echeverría-González, Jorge Fernando Topete-Reyes, Sergio Adalberto Franco-Chávez, and Ingrid Patricia Dávalos-Rodríguez. 2024. "Association of the rs1126616 and rs9138 Variants in the SPP1 Gene among Mexican Patients with Systemic Lupus Erythematosus and Lupus Nephritis" International Journal of Molecular Sciences 25, no. 2: 1000. https://doi.org/10.3390/ijms25021000
APA StyleRivera-Cameras, A., Gallegos-Arreola, M. P., Morán-Moguel, M. C., Salazar-Páramo, M., Alcaraz-López, M. F., Echeverría-González, G., Topete-Reyes, J. F., Franco-Chávez, S. A., & Dávalos-Rodríguez, I. P. (2024). Association of the rs1126616 and rs9138 Variants in the SPP1 Gene among Mexican Patients with Systemic Lupus Erythematosus and Lupus Nephritis. International Journal of Molecular Sciences, 25(2), 1000. https://doi.org/10.3390/ijms25021000