
Molecular Mechanisms for Regulating Stomatal Formation across Diverse Plant Species

 ,
, {kind=link}
{kind=link}
Abstract
1. Introduction
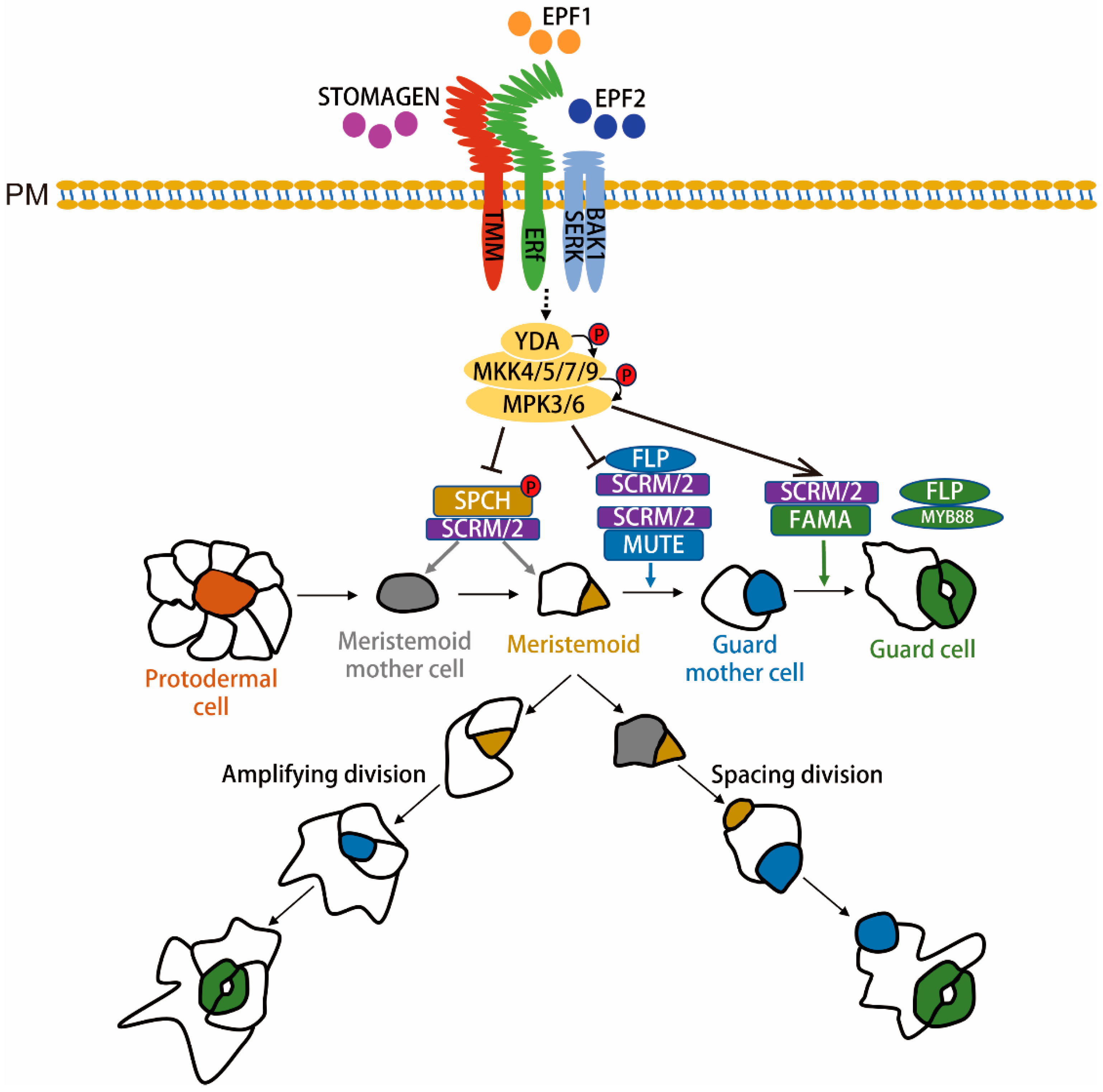
2. Regulation of Stomatal Formation in Arabidopsis
2.1. Transcription Factors Control Stomatal Cell Fate Transitions
2.2. MAPK Signaling Cascade Functions Upstream of Stomatal Transcription Factors to Regulate Stomatal Formation
2.3. Asymmetric Distribution of MAPK Signaling by Scaffold Proteins BASL and POLAR Controls Stomatal Asymmetric Cell Fate
2.4. EPF Ligands and Their Receptors Activate the YDA-MAPK Cascade to Regulate Stomatal Formation
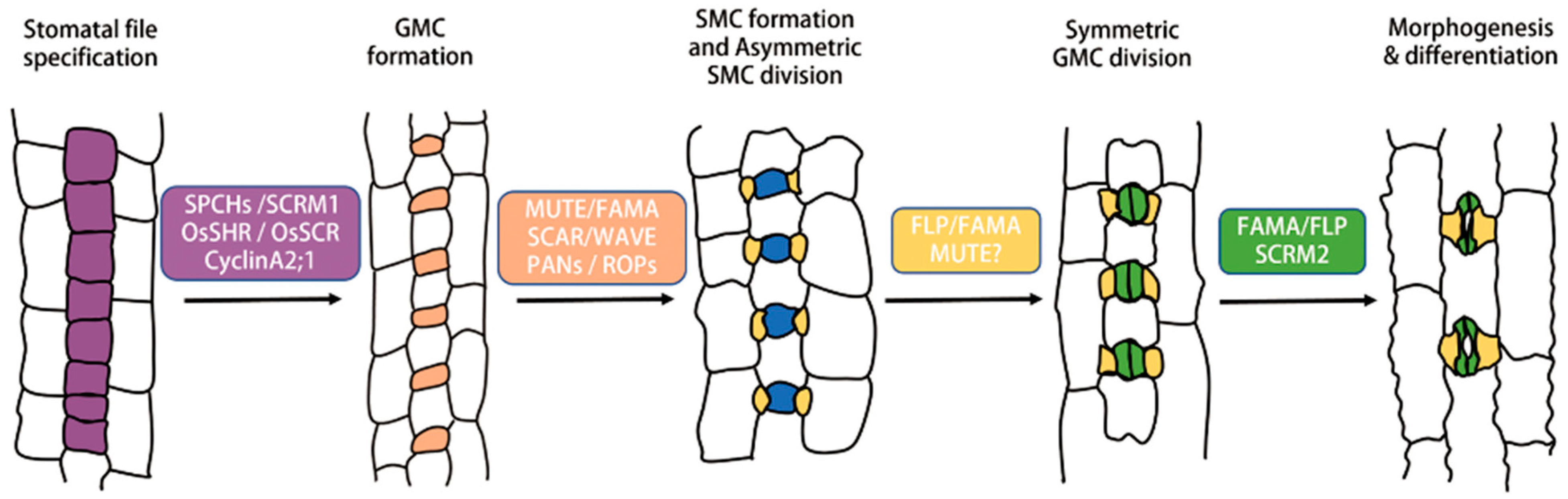
3. Stomatal Formation in Monocot Plants
4. Regulation of Stomatal Formation in Grasses
4.1. Transcription Factors Control Stomatal Cell Differentiation in Grasses
4.2. Regulation of Subdidiary Cell Formation
4.3. Regulation of Grass Stomatal Formation by Other Components
5. The Effect of Plant Hormones on Stomatal Formation
6. The Influence of Environmental Factors on Stomatal Formation
7. Summary and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Rensing, S.A.; Lang, D.; Zimmer, A.D. The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science 2008, 319, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Bennici, A. Origin and early evolution of land plants: Problems and considerations. Commun. Integr. Biol. 2008, 1, 212–218. [Google Scholar] [CrossRef]
- Edwards, D.; Kerp, H.; Hass, H. Stomata in early land plants: An anatomical and ecophysiological approach. J. Exp. Bot. 1998, 49, 255–278. [Google Scholar] [CrossRef]
- Raven, J.A. Selection pressures on stomatal evolution. New Phytol. 2002, 153, 371–386. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.A.; Beerling, D.J.; Franks, P.J. Stomata: Key players in the earth system, past and present. Curr Opin Plant Biol. 2010, 13, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Keeley, J.E.; Osmond, C.B.; Raven, J.A. Stylites, a vascular land plant without stomata absorbs CO2 via its roots. Nature. 1984, 310, 694–695. [Google Scholar] [CrossRef]
- Cuming, A.C.; Cho, S.H.; Kamisugi, Y. Microarray analysis of transcriptional responses to abscisic acid and osmotic, salt, and drought stress in the moss, Physcomitrella patens. New Phytol. 2007, 176, 275–287. [Google Scholar] [CrossRef]
- Sack, F.D.; Paolillo, D.J. Incomplete cytokinesis in Funaria Stomata. Am. J. Bot. 1985, 72, 1325–1333. [Google Scholar] [CrossRef]
- Vaten, A.; Bergmann, D.C. Mechanisms of stomatal development: An evolutionary view. Evodevo. 2012, 3, 11. [Google Scholar] [CrossRef]
- Johnson, R.W.; Riding, R.T. Structure and ontogeny of the stomatal complex in Pinus strobus L. and Pinus banksiana Lamb. Am. J. Bot. 1981, 68, 260–268. [Google Scholar] [CrossRef]
- Hara, K.; Kajita, R.; Torii, K.U. The secretory peptide gene EPF1 enforces the stomatal one-cell-spacing rule. J. Gene Dev. 2007, 21, 1720–1725. [Google Scholar] [CrossRef] [PubMed]
- Serna, L.; Fenoll, C. Stomatal development in Arabidopsis: How to make a functional pattern. Trends Plant Sci. 2000, 5, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, D.C.; Sack, F.D. Stomatal development. Annu. Rev. Plant Biol. 2007, 58, 163–181. [Google Scholar] [CrossRef]
- Ohashi-Ito, K.; Bergmann, D.C. Arabidopsis FAMA controls the final proliferation/differentiation switch during stomatal development. Plant Cell 2006, 18, 2493–2505. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.; Sepuru, K.M.; Putarjunan, A.; Aguirre, L.; Burrows, B.A.; Torii, K.U. Intragenic suppressors unravel the role of the SCREAM ACT-like domain for bHLH partner selectivity in stomatal development. Proc. Nat. Acad. Sci. USA 2022, 119, e2117774119. [Google Scholar] [CrossRef]
- MacAlister, C.A.; Ohashi-Ito, K.; Bergmann, D.C. Transcription factor control of asymmetric cell divisions that establish the stomatal lineage. Nature 2007, 445, 537–540. [Google Scholar] [CrossRef]
- Pillitteri, L.J.; Torii, K.U. Mechanisms of stomatal development. Annu. Rev. Plant Biol. 2012, 63, 591–614. [Google Scholar] [CrossRef]
- Kanaoka, M.M.; Pillitteri, L.J.; Fujii, H.; Yoshida, Y.; Bogenschutz, N.L.; Takabayashi, J.; Zhu, J.K.; Torii, K.U. SCREAM/ICE1 and SCREAM2 specify three cell-state transitional steps leading to Arabidopsis stomatal differentiation. Plant Cell 2008, 20, 1775–1785. [Google Scholar] [CrossRef]
- Han, S.K.; Qi, X.; Sugihara, K.; Dang, J.H.; Endo, T.A.; Miller, K.L.; Kim, E.D.; Miura, T.; Torii, K.U. MUTE directly orchestrates cell-State switch and the single symmetric division to create stomata. Dev. Cell 2018, 45, 303–315. [Google Scholar] [CrossRef]
- Chen, L.; Wu, Z.L.; Hou, S.W. SPEECHLESS speaks loudly in stomatal development. Front. Plant Sci. 2020, 11, 114. [Google Scholar] [CrossRef]
- Qi, X.; Han, S.K.; Dang, J.H.; Garrick, J.M.; Ito, M.; Hofstetter, A.K.; Torii, K.U. Autocrine regulation of stomatal differentiation potential by EPF1 and ERECTA-LIKE1 ligand-receptor signaling. Elife 2017, 6, e24102. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Zhu, L.; Wang, H.; Jiang, M.; Xiao, C.; Hu, X.; Vanneste, S.; Dong, J.; Le, J.A. Conserved but plant-specific CDK-mediated regulation of DNA replication protein A2 in the precise control of stomatal terminal division. Proc. Nat. Acad. Sci. USA 2019, 116, 18126–18131. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Lucas, J.R.; Sack, F.D. Deep functional redundancy between FAMA and FOUR LIPS in stomatal development. Plant J. 2014, 78, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Lai, L.B.; Nadeau, J.A.; Lucas, J.; Lee, E.K.; Nakagawa, T.; Zhao, L.; Geisler, M.; Sack, F.D. The Arabidopsis R2R3 MYB proteins FOUR LIPS and MYB88 restrict divisions late in the stomatal cell lineage. Plant Cell 2005, 17, 2754–2767. [Google Scholar] [CrossRef]
- Li, P.; Chen, L.; Gu, X.; Zhao, M.; Wang, W.; Hou, S.W. FOUR LIPS plays a role in meristemoid-to-GMC fate transition during stomatal development in Arabidopsis. Plant J. 2023, 114, 424–436. [Google Scholar] [CrossRef]
- Chen, L.; Zhao, M.; Wu, Z.; Chen, S.; Rojo, E.; Luo, J.; Li, P.; Zhao, L.; Chen, Y.; Deng, J.; et al. RNA polymerase II associated proteins regulate stomatal development through direct interaction with stomatal transcription factors in Arabidopsis thaliana. New Phytol. 2021, 230, 171–189. [Google Scholar] [CrossRef]
- Lampard, G.R.; Lukowitz, W.; Ellis, B.E.; Bergmann, D.C. Novel and expanded roles for MAPK signaling in Arabidopsis stomatal cell fate revealed by cell type-specific manipulations. Plant Cell 2009, 21, 3506–3517. [Google Scholar] [CrossRef]
- Lampard, G.R.; Macalister, C.A.; Bergmann, D.C. Arabidopsis stomatal initiation is controlled by MAPK-mediated regulation of the bHLH SPEECHLESS. Science 2008, 322, 1113–1116. [Google Scholar] [CrossRef] [PubMed]
- Umbrasaite, J.; Schweighofer, A.; Kazanaviciute, V.; Magyar, Z.; Ayatollahi, Z.; Unterwurzacher, V.; Choopayak, C.; Boniecka, J.; Murray, J.A.; Bogre, L.; et al. MAPK phosphatase AP2C3 induces ectopic proliferation of epidermal cells leading to stomata development in Arabidopsis. PLoS One 2010, 5, e15357. [Google Scholar] [CrossRef]
- Tamnanloo, F.; Damen, H.; Jangra, R.; Lee, J.S. MAPKINASE PHOSPHATASE1 controls cell fate transition during stomatal development. Plant Physiol. 2018, 178, 247–257. [Google Scholar] [CrossRef]
- Dong, J.; MacAlister, C.A.; Bergmann, D.C. BASL controls asymmetric cell division in Arabidopsis. Cell 2009, 137, 1320–1330. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, P.; Shao, W.; Zhu, J.K.; Dong, J. The BASL polarity protein controls a MAPK signaling feedback loop in asymmetric cell division. Dev. Cell 2015, 33, 136–149. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, X.; Dong, J. Phosphorylation of the polarity protein BASL differentiates asymmetric cell fate through MAPKs and SPCH. Curr. Biol. 2016, 26, 2957–2965. [Google Scholar] [CrossRef] [PubMed]
- Houbaert, A. POLAR-guided signalling complex assembly and localization drive asymmetric cell division. Nature 2018, 563, 574–578. [Google Scholar] [CrossRef]
- Hara, K.; Yokoo, T.; Kajita, R.; Onishi, T.; Yahata, S.; Peterson, K.M.; Torii, K.U.; Kakimoto, T. Epidermal cell density is autoregulated via a secretory peptide, EPIDERMAL PATTERNING FACTOR 2 in Arabidopsis leaves. Plant Cell Physiol. 2009, 50, 1019–1031. [Google Scholar] [CrossRef]
- Hunt, L.; Gray, J.E. The signaling peptide EPF2 controls asymmetric cell divisions during stomatal development. Curr. Biol. 2009, 19, 864–869. [Google Scholar] [CrossRef] [PubMed]
- Sugano, S.S.; Shimada, T.; Imai, Y.; Okawa, K.; Tamai, A.; Mori, M.; Hara-Nishimura, I. Stomagen positively regulates stomatal density in Arabidopsis. Nature 2010, 463, 241–244. [Google Scholar] [CrossRef]
- Lee, J.S.; Kuroha, T.; Hnilova, M.; Khatayevich, D.; Kanaoka, M.M.; Mcabee, J.M.; Sarikaya, M.; Tamerler, C.; Torii, K.U. Direct interaction of ligand-receptor pairs specifying stomatal patterning. Genes Dev. 2012, 26, 126–136. [Google Scholar] [CrossRef]
- Lee, J.S.; Hnilova, M.; Maes, M.; Lin, Y.C.L.; Putarjunan, A.; Han, S.K.; Avila, J.; Torii, K.U. Competitive binding of antagonistic peptides fine-tunes stomatal patterning. Nature 2015, 522, 439–443. [Google Scholar] [CrossRef]
- Abrash, E.B.; Davies, K.A.; Bergmann, D.C. Generation of signaling specificity in Arabidopsis by spatially restricted buffering of ligand-receptor interactions. Plant Cell 2011, 23, 2864–2879. [Google Scholar] [CrossRef]
- Kondo, T.; Kajita, R.; Miyazaki, A.; Hokoyama, M.; Nakamura-Miura, T.; Mizuno, S.; Masuda, Y.; Irie, K.; Tanaka, Y.; Takada, S.; et al. Stomatal density is controlled by a mesophyll-derived signaling molecule. Plant Cell Physiol. 2010, 51, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Zhang, L.; Han, Z.; Yang, X.; Liu, W.; Li, E.; Chang, J.; Qi, Y.; Shpak, E.D.; Chai, J. A receptor-like protein acts as a specificity switch for the regulation of stomatal development. Genes Dev. 2017, 31, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Chen, X.; Mang, H.; Liu, C.; Yu, X.; Gao, X.; Torii, K.U.; He, P.; Shan, L. Differential function of Arabidopsis SERK family receptor-like kinases in stomatal patterning. Curr. Biol. 2015, 25, 2361–2372. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.M.; Paciorek, T.; Abrash, E.; Bergmann, D.C. Modulators of stomatal lineage signal transduction alter membrane contact sites and reveal specialization among ERECTA kinases. Dev. Cell 2016, 38, 345–357. [Google Scholar] [CrossRef]
- Hepworth, C.; Caine, R.S.; Harrison, E.L.; Sloan, J.; Gray, J.E. Stomatal development: Focusing on the grasses. Curr. Opin. Plant Biol. 2018, 41, 1–7. [Google Scholar] [CrossRef]
- Abrash, E.; Anleu Gil, M.X.; Matos, J.L.; Bergmann, D.C. Conservation and divergence of YODA MAPKKK function in regulation of grass epidermal patterning. Development 2018, 145, dev165860. [Google Scholar] [CrossRef]
- Wu, Z.; Chen, L.; Yu, Q.; Zhou, W.Q.; Gou, X.P.; Li, J.; Hou, S.W. Multiple transcriptional factors control stomata development in rice. New Phytol. 2019, 223, 220–232. [Google Scholar] [CrossRef]
- McKown, K.H.; Bergmann, D.C. Stomatal development in the grasses: Lessons from models and crops (and crop models). New Phytol. 2020, 227, 1636–1648. [Google Scholar] [CrossRef]
- Nunes, T.D.G.; Zhang, D.; Raissig, M.T. Form, development and function of grass stomata. Plant J. 2020, 101, 780–799. [Google Scholar] [CrossRef]
- Wang, H.; Guo, S.; Qiao, X.; Guo, J.; Li, Z.; Zhou, Y.; Bai, S.; Gao, Z.; Wang, D.; Wang, P.; et al. BZU2/ZmMUTE controls symmetrical division of guard mother cell and specifies neighbor cell fate in maize. PLoS Genet. 2019, 15, e1008377. [Google Scholar] [CrossRef]
- McKown, K.H.; Anleu Gil, M.X.; Mair, A.; Xu, S.L.; Raissig, M.T.; Bergmann, D.C. Expanded roles and divergent regulation of FAMA in Brachypodium and Arabidopsis stomatal development. Plant Cell 2023, 35, 756–775. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, K.L.; Benfey, P.N. Both the conserved GRAS domain and nuclear localization are required for SHORT-ROOT movement. Plant J. 2009, 57, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Schuler, M.L.; Sedelnikova, O.V.; Walker, B.J.; Westhoff, P.; Langdale, J.A. SHORTROOT-mediated increase in stomatal density has no impact on photosynthetic efficiency. Plant Physiol. 2018, 176, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Cartwright, H.N.; Humphries, J.A.; Smith, L.G. PAN1: A receptor-like protein that promotes polarization of an asymmetric cell division in maize. Science 2019, 323, 649–651. [Google Scholar] [CrossRef]
- Zhang, X.; Facette, M.; Humphries, J.A.; Shen, Z.; Park, Y.; Sutimantanapi, D.; Sylvester, A.W.; Briggs, S.P.; Smith, L.G. Identification of PAN2 by quantitative proteomics as a leucine-rich repeat-receptor-like kinase acting upstream of PAN1 to polarize cell division in maize. Plant Cell 2012, 24, 4577–4589. [Google Scholar] [CrossRef]
- Humphries, J.A.; Vejlupkova, Z.; Luo, A.; Meeley, R.B.; Sylvester, A.W.; Fowler, J.E.; Smith, L.G. ROP GTPases act with the receptor-like protein PAN1 to polarize asymmetric cell division in maize. Plant Cell 2011, 23, 2273–2284. [Google Scholar] [CrossRef]
- Facette, M.R.; Park, Y.; Sutimantanapi, D.; Luo, A.; Cartwright, H.N.; Yang, B.; Bennett, E.J.; Sylvester, A.W.; Smith, L.G. The SCAR/WAVE complex polarizes PAN receptors and promotes division asymmetry in maize. Nat. Plants 2015, 1, 14024. [Google Scholar] [CrossRef]
- Zhou, W.Q.; Wang, Y.C.; Wu, Z.L.; Luo, L.; Liu, P.; Yan, L.F.; Hou, S.W. Homologs of SCAR/WAVE complex components are required for epidermal cell morphogenesis in rice. J. Exp. Bot. 2016, 67, 4311–4323. [Google Scholar] [CrossRef]
- Zhang, D.; Spiegelhalder, R.P.; Abrash, E.B.; Nunes, T.D.G.; Hidalgo, I.; Anleu Gil, M.X.; Jesenofsky, B.; Lindner, H.; Bergmann, D.C.; Raissig, M.T. Opposite polarity programs regulate asymmetric subsidiary cell divisions in grasses. eLife 2022, 11, e79913. [Google Scholar] [CrossRef] [PubMed]
- Nan, Q.; Char, S.N.; Yang, B.; Bennett, E.J.; Yang, B.; Facette, M.R. Polarly localized WPR proteins interact with PAN receptors and the actin cytoskeleton during maize stomatal development. Plant Cell 2023, 35, 469–487. [Google Scholar] [CrossRef]
- Yin, X.; Biswal, A.K.; Dionora, J.; Perdigon, K.M.; Balahadia, C.P.; Mazumdar, S.; Chater, C.; Lin, H.C.; Coe, R.A.; Kretzschmar, T.; et al. CRISPR-Cas9 and CRISPR-Cpf1 mediated targeting of a stomatal developmental gene EPFL9 in rice. Plant Cell Rep. 2017, 36, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.; Hepworth, C.; Dutton, C.; Dunn, J.A.; Hunt, L.; Stephens, J.; Waugh, R.; Cameron, D.D.; Gray, J.E. Reducing stomatal density in barley improves drought tolerance without impacting on yield. Plant Physiol. 2017, 174, 776–787. [Google Scholar] [CrossRef]
- Lu, J.; He, J.; Zhou, X.; Zhong, J.; Li, J.; Liang, Y.K. Homologous genes of epidermal patterning factor regulate stomatal development in rice. J. Plant Physiol. 2019, 234–235, 18–27. [Google Scholar] [CrossRef]
- Dunn, J.; Hunt, L.; Afsharinafar, M.; Meselmani, M.; Al Mitchell, A.; Howells, R.; Wallington, E.; Fleming, A.J.; Gray, J.E. Reduced stomatal density in bread wheat leads to increased water-use efficiency. J. Exp. Bot. 2019, 70, 4737–4747. [Google Scholar] [CrossRef]
- Jangra, R.; Brunetti, S.C.; Wang, X.; Kaushik, P.; Gulick, P.J.; Foroud, N.A.; Wang, S.; Lee, J.S. Duplicated antagonistic EPF peptides optimize grass stomatal initiation. Development 2021, 148, dev199780. [Google Scholar] [CrossRef] [PubMed]
- Caine, R.S.; Yin, X.; Sloan, J.; Harrison, E.L.; Mohammed, U.; Fulton, T.; Biswal, A.K.; Dionora, J.; Chater, C.C.; Coe, R.A.; et al. Rice with reduced stomatal density conserves water and has improved drought tolerance under future climate conditions. New Phytol. 2019, 221, 371–384. [Google Scholar] [CrossRef]
- Qu, X.; Yan, M.; Zou, J.; Jiang, M.; Yang, K.; Le, J. A2-type cyclin is required for the asymmetric entry division in rice stomatal development. J. Exp. Bot. 2018, 69, 3587–3599. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, T.; Wang, X.; Wu, W.; Xing, J.; Li, Z.; Qiao, X.; Zhang, C.; Wang, X.; Wang, G.; et al. A maize epimerase modulates cell wall synthesis and glycosylation during stomatal morphogenesis. Nat. Commun. 2023, 14, 4384. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.W.; Michniewicz, M.; Bergmann, D.C.; Wang, Z.Y. Brassinosteroid regulates stomatal development by GSK3-mediated inhibition of a MAPK pathway. Nature 2012, 482, 419–422. [Google Scholar] [CrossRef]
- Tanaka, Y.; Nose, T.; Jikumaru, Y.; Kamiya, Y. ABA inhibits entry into stomatal-lineage development in Arabidopsis leaves. Plant J. 2013, 74, 448–457. [Google Scholar] [CrossRef]
- Le, J.; Liu, X.G.; Yang, K.Z.; Chen, X.L.; Zou, J.J.; Wang, H.Z.; Wang, M.; Vanneste, S.; Morita, M.; Tasaka, M.; et al. Auxin transport and activity regulate stomatal patterning and development. Nat. Commun. 2014, 5, 3090. [Google Scholar] [CrossRef] [PubMed]
- Balcerowicz, M.; Hoecker, U. Auxin—A novel regulator of stomata differentiation. Trends Plant Sci. 2014, 19, 747–749. [Google Scholar] [CrossRef] [PubMed]
- Vatén, A.; Soyars, C.L.; Tarr, P.T.; Nimchuk, Z.L.; Bergmann, D.C. Modulation of asymmetric division diversity through cytokinin and SPEECHLESS regulatory interactions in the Arabidopsis stomatal lineage. Dev. Cell 2018, 47, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Rovira, A.; Veciana, N.; Basté-Miquel, A.; Quevedo, M.; Locascio, A.; Yenush, L.; Toledo-Ortiz, G.; Leivar, P.; Monte, E. PIF transcriptional regulators are required for rhythmic stomatal movements. Nat. Commun. 2024, 15, 4540. [Google Scholar] [CrossRef]
- Kang, C.Y.; Lian, H.L.; Wang, F.F.; Huang, J.R.; Yang, H.Q. Cryptochromes, phytochromes, and COP1 regulate light-controlled stomatal development in Arabidopsis. Plant Cell 2009, 21, 2624–2641. [Google Scholar] [CrossRef]
- Lee, J.H.; Jung, J.H.; Park, C.M. Light inhibits COP1-mediated degradation of ICE transcription factors to induce stomatal development in Arabidopsis. Plant Cell 2017, 29, 2817–2830. [Google Scholar] [CrossRef]
- Casson, S.A.; Franklin, K.A.; Gray, J.E.; Grierson, C.S.; Whitelam, G.C.; Hetherington, A.M. Phytochrome B and PIF4 regulate stomatal development in response to light quantity. Curr. Biol. 2009, 19, 229–234. [Google Scholar] [CrossRef]
- Klermund, L.; Riederer, A.; Hunger, A.; Castiglione, K. Protein engineering of a bacterial N-acyl-d-glucosamine 2-epimerase for improved stability under process conditions. Enzym. Microb. Technol. 2016, 87–88, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.E.; Holroyd, G.H.; van der Lee, F.M.; Bahrami, A.R.; Sijmons, P.C.; Woodward, F.I.; Schuch, W.; Hetherington, A.M. The HIC signalling pathway links CO2 perception to stomatal development. Nature 2000, 408, 713–716. [Google Scholar] [CrossRef]
- Engineer, C.B.; Ghassemian, M.; Anderson, J.C.; Peck, S.C.; Hu, H.; Schroeder, J.I. Carbonic anhydrases, EPF2 and a novel protease mediate CO2 control of stomatal development. Nature 2014, 513, 246–250, Erratum in Nature 2015, 526, 458. [Google Scholar] [CrossRef]
- Koini, M.A.; Alvey, L.; Allen, T.; Tilley, C.A.; Harberd, N.P.; Whitelam, G.C.; Franklin, K.A. High temperature-mediated adaptations in plant architecture require the bHLH transcription factor PIF4. Curr. Biol. 2009, 19, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Lau, O.S.; Song, Z.; Zhou, Z.; Davies, K.A.; Chang, J.; Yang, X.; Wang, S.; Lucyshyn, D.; Tay, I.H.Z.; Wigge, P.A.; et al. Direct control of SPEECHLESS by PIF4 in the high-temperature response of stomatal development. Curr. Biol. 2018, 28, 1273–1280.e3. [Google Scholar] [CrossRef] [PubMed]
- Melotto, M.; Underwood, W.; Koczan, J.; Nomura, K.; He, S.Y. Plant stomata function in innate immunity against bacterial invasion. Cell 2006, 126, 969–980. [Google Scholar] [CrossRef]
- Melotto, M.; Zhang, L.; Oblessuc, P.R.; He, S.Y. Stomatal defense a decade later. Plant Physiol. 2017, 174, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Chen, L. Emerging roles of protein phosphorylation in regulation of stomatal development. Plant Physiol. 2023, 280, 153882. [Google Scholar] [CrossRef]
- Chen, L. Regulation of stomatal development by epidermal, subepidermal and long-distance signals. Plant Mol. Biol. 2024, 114, 80. [Google Scholar] [CrossRef]
- Le Gloanec, C.; Gómez-Felipe, A.; Alimchandani, V.; Branchini, E.; Bauer, A.; Routier-Kierzkowska, A.L.; Kierzkowski, D. Modulation of cell differentiation and growth underlies the shift from bud protection to light capture in cauline leaves. Plant Physiol. 2024, kiae408. [Google Scholar] [CrossRef]
- Wu, M.; Wang, S.; Ma, P.; Li, B.; Hu, H.; Wang, Z.; Qiu, Q.; Qiao, Y.; Niu, D.; Lukowitz, W.; et al. Dual roles of the MPK3 and MPK6 mitogen-activated protein kinases in regulating Arabidopsis stomatal development. Plant Cell 2024, koae225. [Google Scholar] [CrossRef]
- Zhang, K.; Xue, M.; Qin, F.; He, Y.; Zhou, Y. Natural polymorphisms in ZmIRX15A affect water-use efficiency by modulating stomatal density in maize. Plant Biotechnol. J. 2023, 21, 2560–2573. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, W.; Liu, J.; Wang, W.; Li, Y.; Ma, Z.; He, H.; Wang, X.; Lian, X.; Dong, X.; Zhao, X.; et al. Molecular Mechanisms for Regulating Stomatal Formation across Diverse Plant Species. Int. J. Mol. Sci. 2024, 25, 10403. https://doi.org/10.3390/ijms251910403
Zhou W, Liu J, Wang W, Li Y, Ma Z, He H, Wang X, Lian X, Dong X, Zhao X, et al. Molecular Mechanisms for Regulating Stomatal Formation across Diverse Plant Species. International Journal of Molecular Sciences. 2024; 25(19):10403. https://doi.org/10.3390/ijms251910403
Chicago/Turabian StyleZhou, Wenqi, Jieshan Liu, Wenjin Wang, Yongsheng Li, Zixu Ma, Haijun He, Xiaojuan Wang, Xiaorong Lian, Xiaoyun Dong, Xiaoqiang Zhao, and et al. 2024. "Molecular Mechanisms for Regulating Stomatal Formation across Diverse Plant Species" International Journal of Molecular Sciences 25, no. 19: 10403. https://doi.org/10.3390/ijms251910403
APA StyleZhou, W., Liu, J., Wang, W., Li, Y., Ma, Z., He, H., Wang, X., Lian, X., Dong, X., Zhao, X., & Zhou, Y. (2024). Molecular Mechanisms for Regulating Stomatal Formation across Diverse Plant Species. International Journal of Molecular Sciences, 25(19), 10403. https://doi.org/10.3390/ijms251910403