
Study of Insulin Aggregation and Fibril Structure under Different Environmental Conditions



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
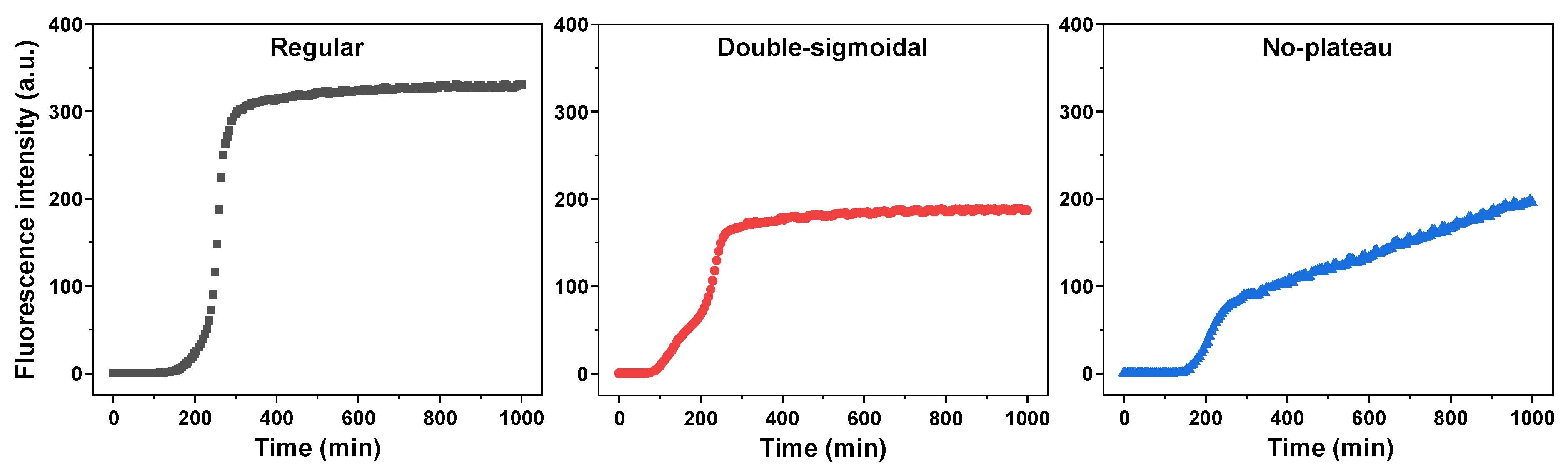
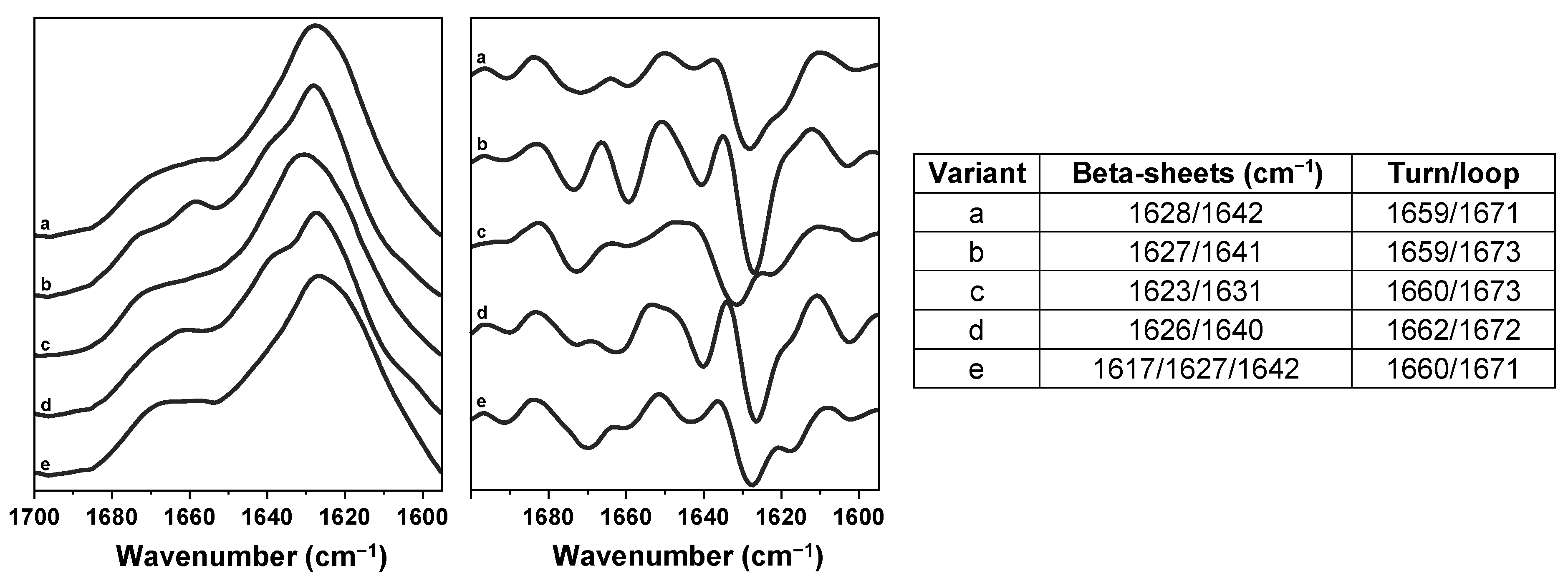
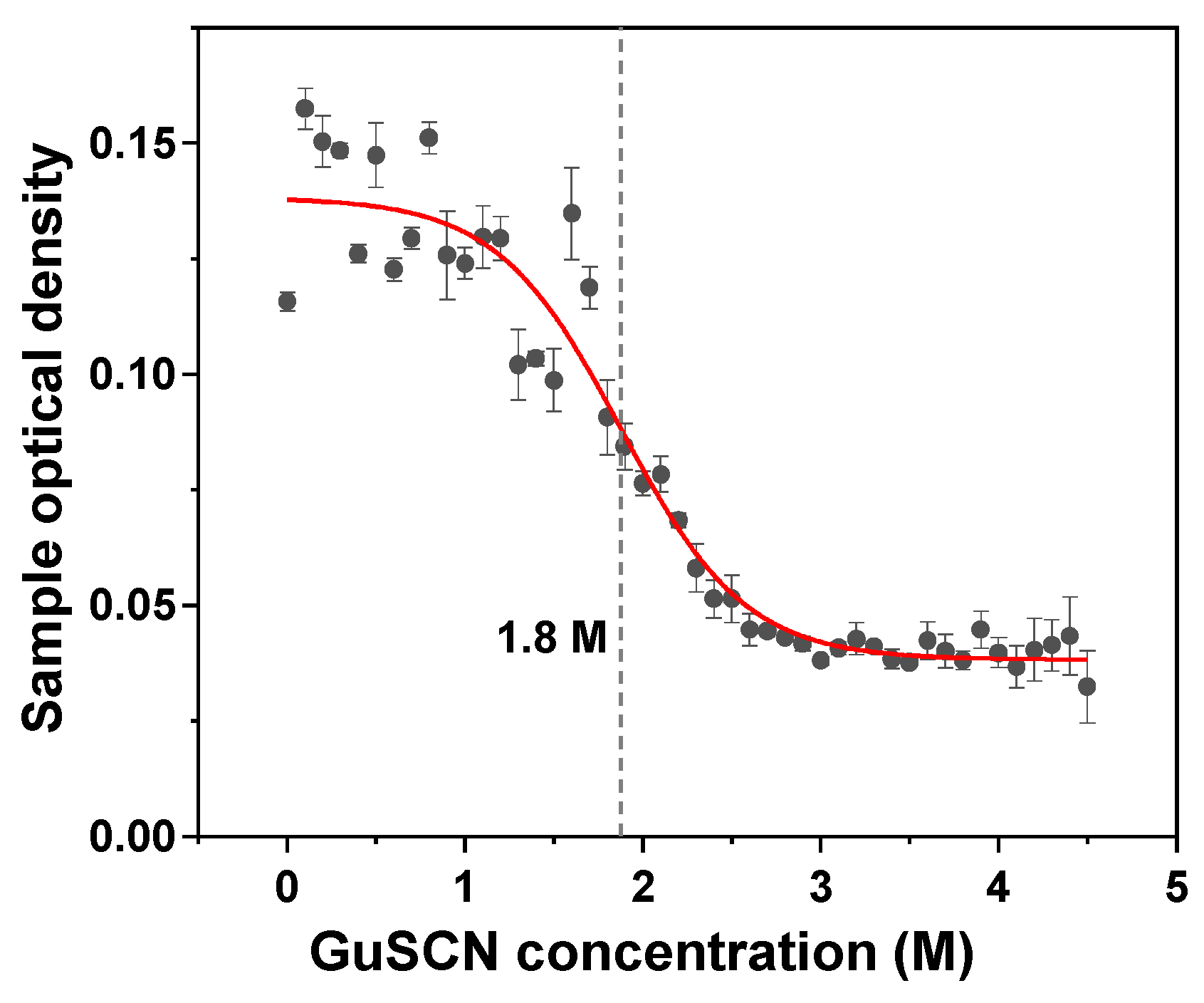
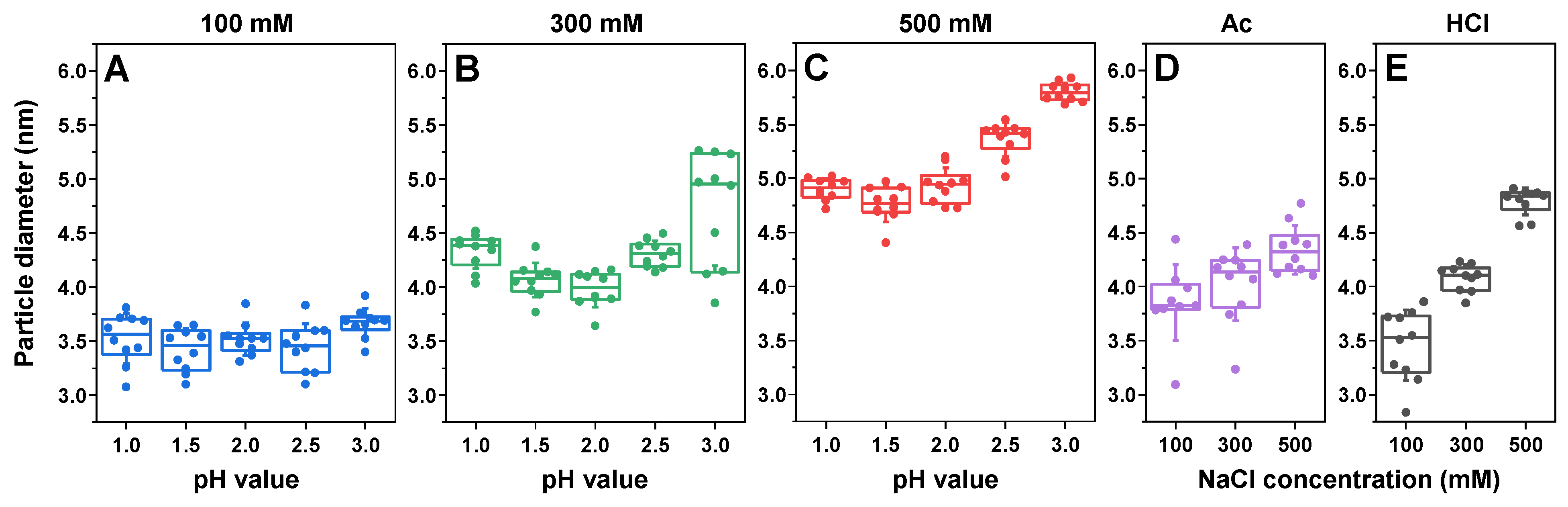
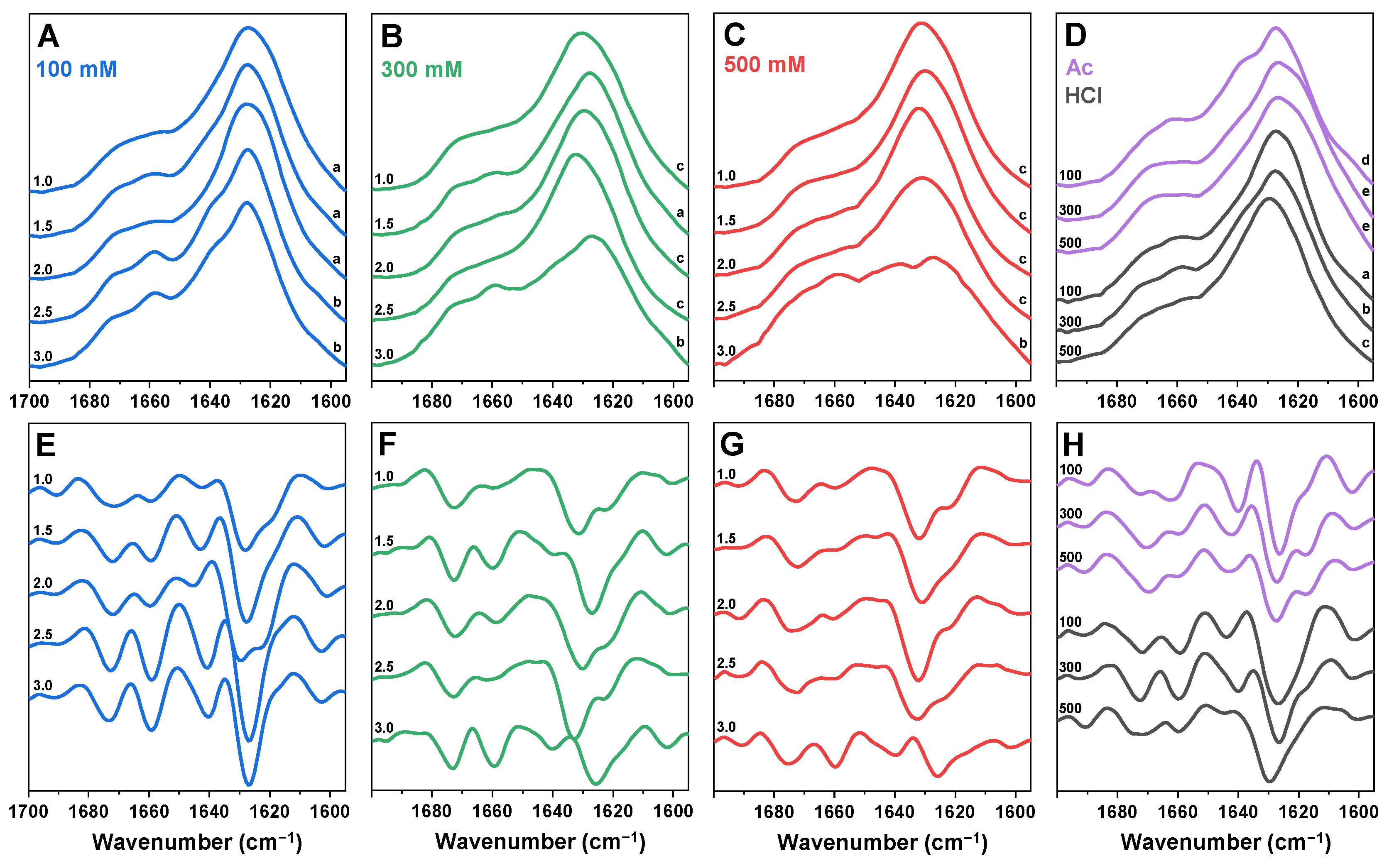
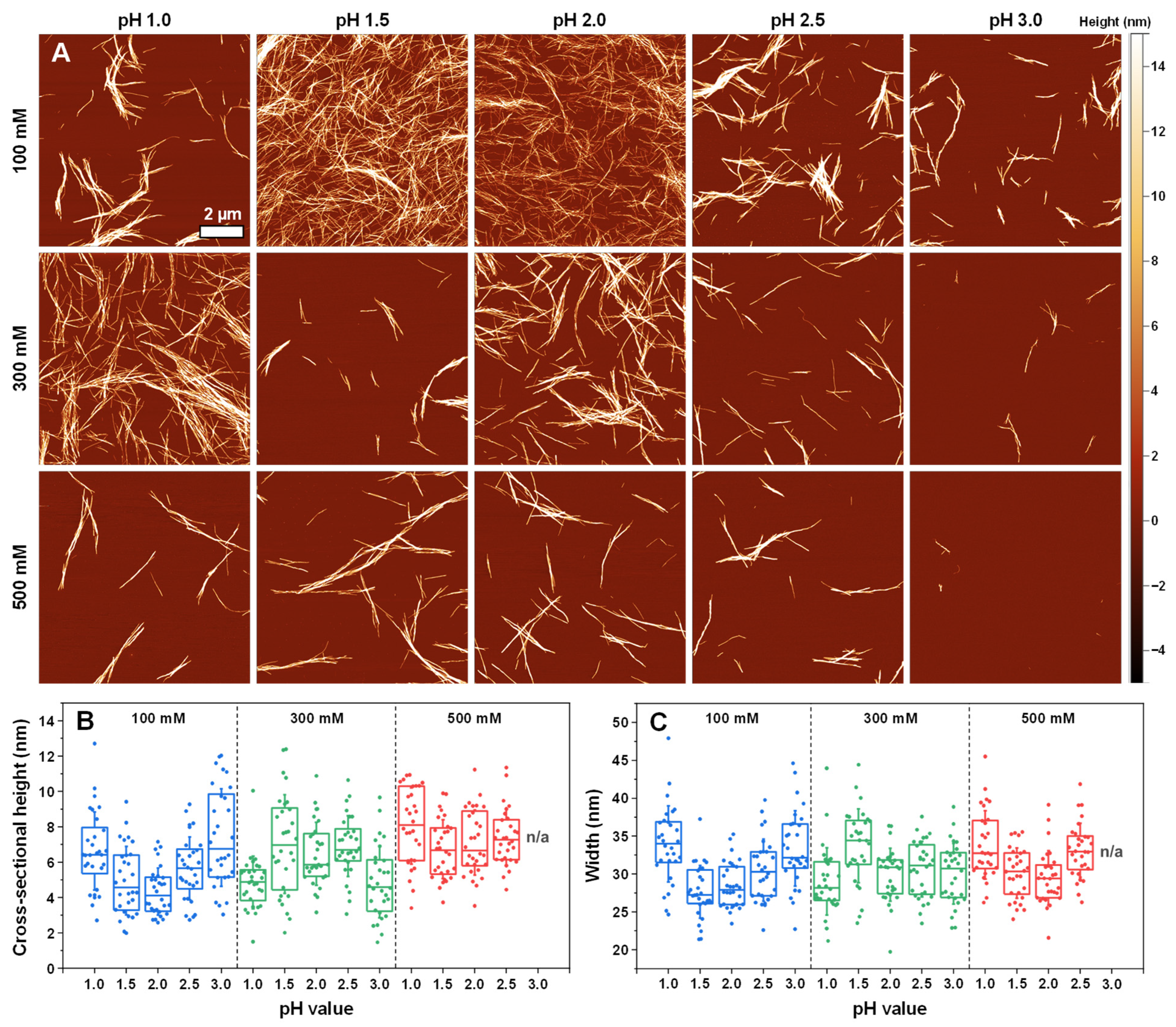
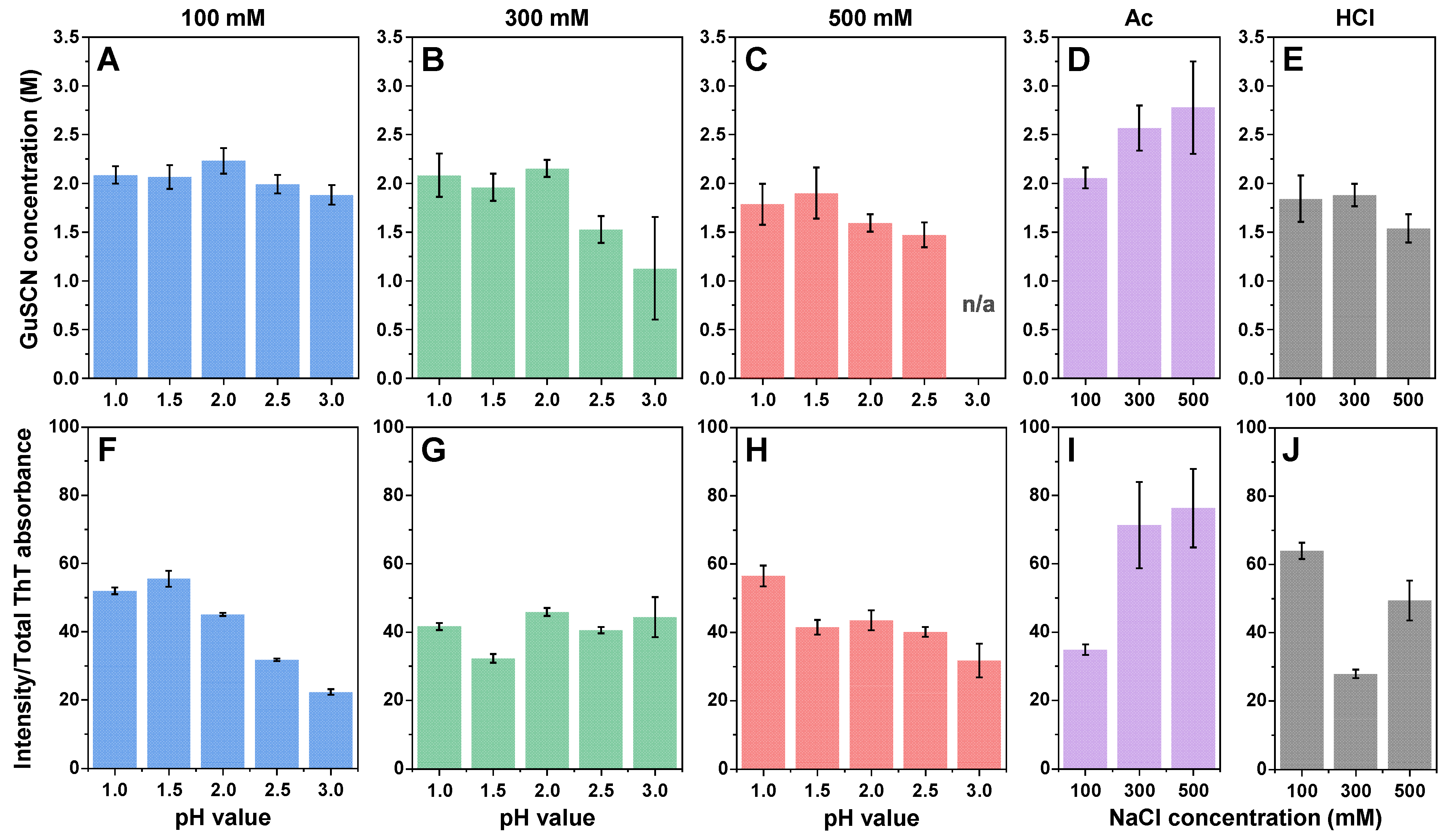
2. Results
3. Discussion
4. Materials and Methods
4.1. Reaction Solution Preparation
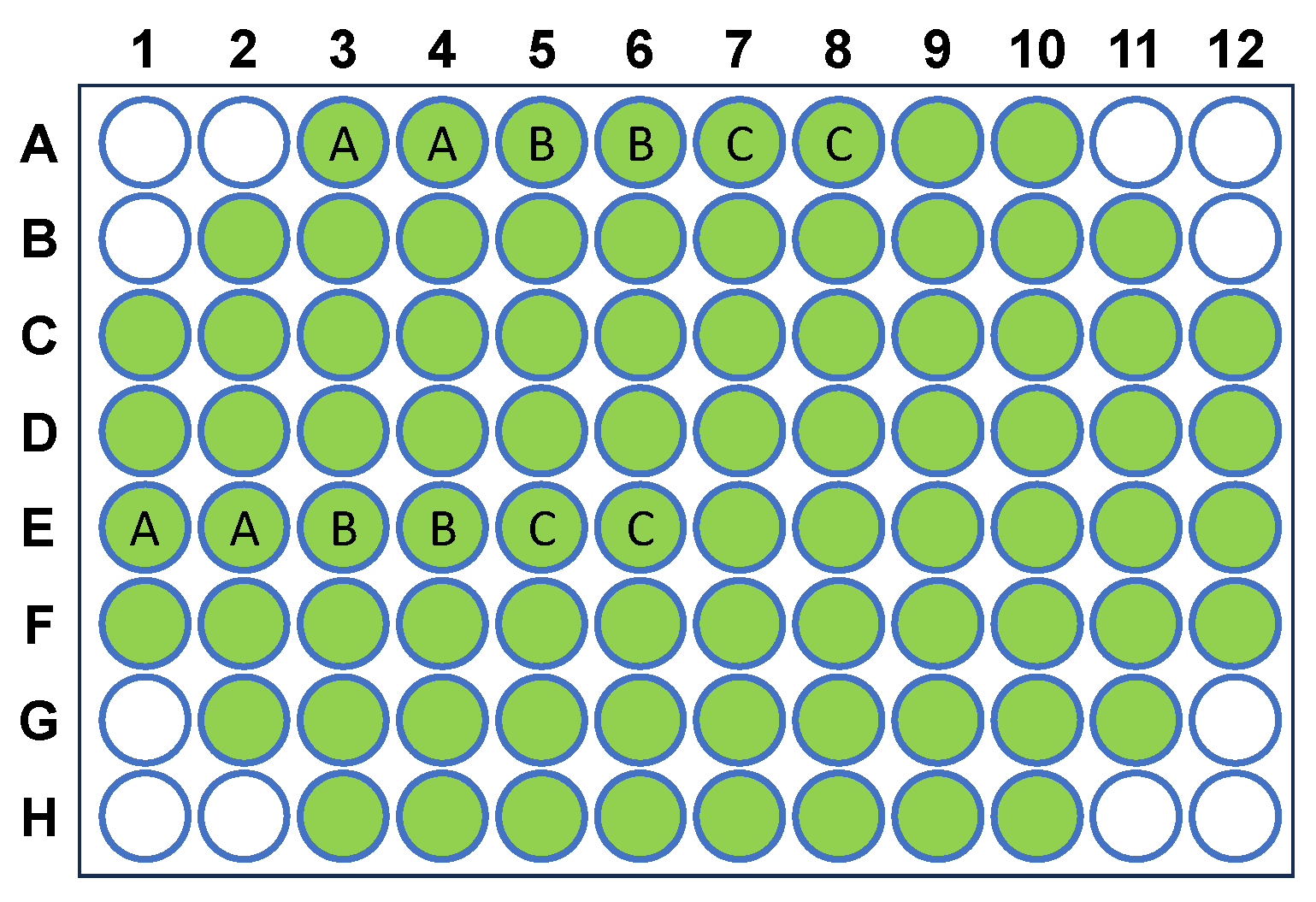
4.2. Insulin Aggregation
4.3. Dynamic Light Scattering
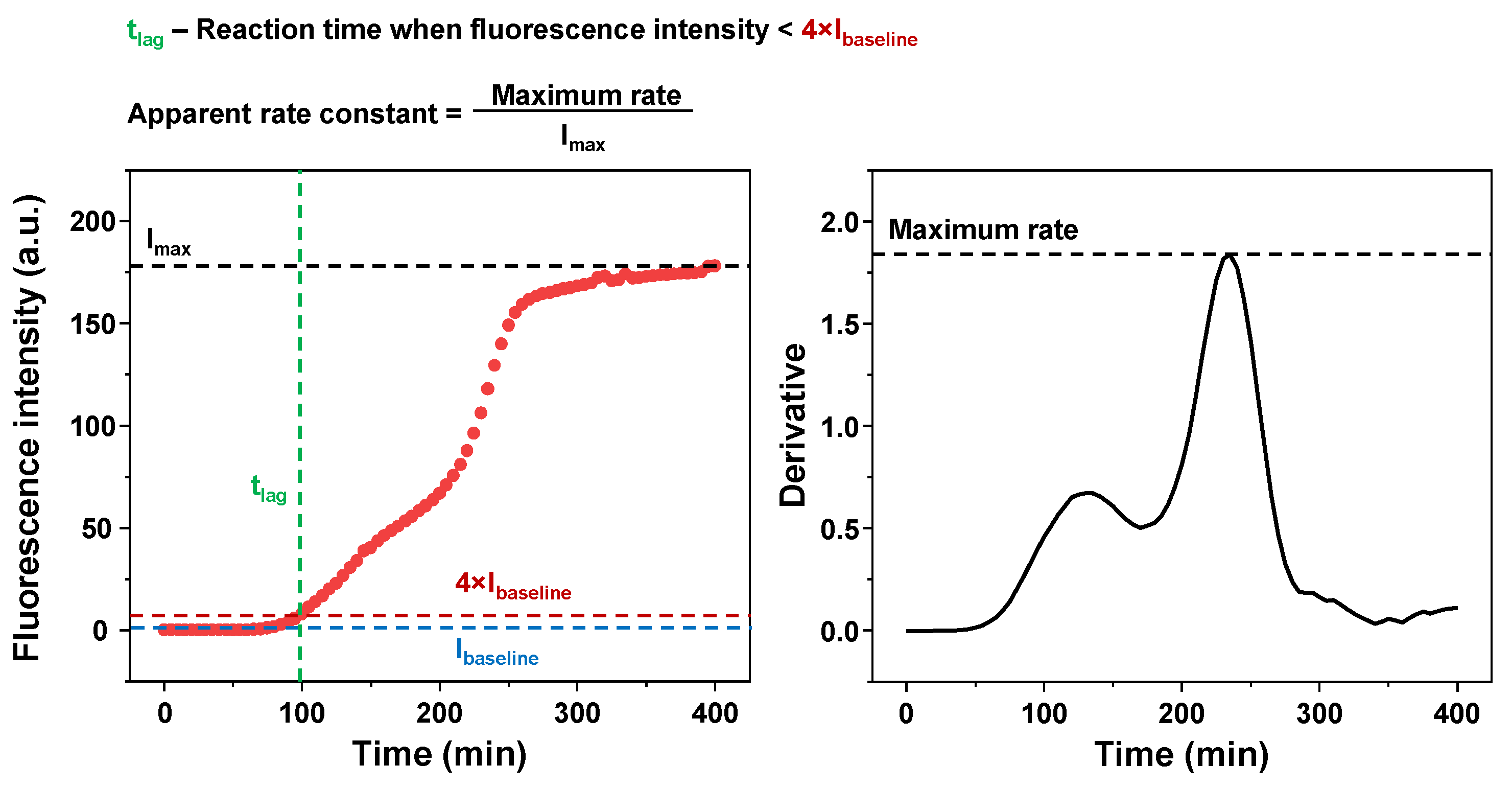
4.4. Kinetic Data Analysis
4.5. Fourier-Transform Infrared Spectroscopy (FTIR)
4.6. ThT-Binding Assay
4.7. Atomic Force Microscopy (AFM)
4.8. Aggregate Denaturation Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
References
- Knowles, T.P.J.; Vendruscolo, M.; Dobson, C.M. The amyloid state and its association with protein misfolding diseases. Nat. Rev. Mol. Cell Biol. 2014, 15, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.; Lee, G.; Nahed, P.; Kambar, M.E.Z.N.; Zhong, K.; Fonseca, J.; Taghva, K. Alzheimer’s disease drug development pipeline: 2022. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2022, 8, e12295. [Google Scholar] [CrossRef]
- Arthur, K.C.; Calvo, A.; Price, T.R.; Geiger, J.T.; Chiò, A.; Traynor, B.J. Projected increase in amyotrophic lateral sclerosis from 2015 to 2040. Nat. Commun. 2016, 7, 12408. [Google Scholar] [CrossRef]
- Brookmeyer, R.; Gray, S.; Kawas, C. Projections of Alzheimer’s disease in the United States and the public health impact of delaying disease onset. Am. J. Public Health 1998, 88, 1337–1342. [Google Scholar] [CrossRef]
- Chatani, E.; Yamamoto, N. Recent progress on understanding the mechanisms of amyloid nucleation. Biophys. Rev. 2018, 10, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Bousset, L.; Pieri, L.; Ruiz-Arlandis, G.; Gath, J.; Jensen, P.H.; Habenstein, B.; Madiona, K.; Olieric, V.; Böckmann, A.; Meier, B.H.; et al. Structural and functional characterization of two alpha-synuclein strains. Nat. Commun. 2013, 4, 2575. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Thackray, A.M.; Hopkins, L.; Monie, T.P.; Burke, D.F.; Bujdoso, R. Polymorphisms at amino acid residues 141 and 154 influence conformational variation in ovine PrP. BioMed Res. Int. 2014, 2014, 372491. [Google Scholar] [CrossRef]
- Fändrich, M.; Meinhardt, J.; Grigorieff, N. Structural polymorphism of Alzheimer Aβ and other amyloid fibrils. Prion 2009, 3, 89–93. [Google Scholar] [CrossRef]
- Zhang, W.; Falcon, B.; Murzin, A.G.; Fan, J.; Crowther, R.A.; Goedert, M.; Scheres, S.H.W. Heparin-induced tau filaments are polymorphic and differ from those in Alzheimer’s and pick’s diseases. eLife 2019, 8, e43584. [Google Scholar] [CrossRef]
- Surewicz, W.K.; Apostol, M.I. Prion Protein and Its Conformational Conversion: A Structural Perspective. In Peptide-Based Materials; Springer: Berlin/Heidelberg, Germany, 2011; Volume 310, pp. 135–167. ISBN 978-3-540-73346-1. [Google Scholar]
- Sidhu, A.; Segers-Nolten, I.; Raussens, V.; Claessens, M.M.A.E.; Subramaniam, V. Distinct Mechanisms Determine α-Synuclein Fibril Morphology during Growth and Maturation. ACS Chem. Neurosci. 2017, 8, 538–547. [Google Scholar] [CrossRef]
- Ziaunys, M.; Sakalauskas, A.; Smirnovas, V. Identifying Insulin Fibril Conformational Differences by Thioflavin-T Binding Characteristics. Biomacromolecules 2020, 21, 4989–4997. [Google Scholar] [CrossRef] [PubMed]
- Mocanu, M.-M.; Ganea, C.; Siposova, K.; Filippi, A.; Demjen, E.; Marek, J.; Bednarikova, Z.; Antosova, A.; Baran, I.; Gazova, Z. Polymorphism of hen egg white lysozyme amyloid fibrils influences the cytotoxicity in LLC-PK1 epithelial kidney cells. Int. J. Biol. Macromol. 2014, 65, 176–187. [Google Scholar] [CrossRef]
- Tycko, R. Amyloid Polymorphism: Structural Basis and Neurobiological Relevance. Neuron 2015, 86, 632–645. [Google Scholar] [CrossRef] [PubMed]
- Sternke-Hoffmann, R.; Peduzzo, A.; Bolakhrif, N.; Haas, R.; Buell, A.K. The aggregation conditions define whether EGCG is an inhibitor or enhancer of α-synuclein amyloid fibril formation. Int. J. Mol. Sci. 2020, 21, 1995. [Google Scholar] [CrossRef]
- Sinnige, T. Molecular mechanisms of amyloid formation in living systems. Chem. Sci. 2022, 13, 7080–7097. [Google Scholar] [CrossRef] [PubMed]
- Ow, S.-Y.; Dunstan, D.E. The effect of concentration, temperature and stirring on hen egg white lysozyme amyloid formation. Soft Matter 2013, 9, 9692–9701. [Google Scholar] [CrossRef]
- Wawer, J.; Szociński, M.; Olszewski, M.; Piątek, R.; Naczk, M.; Krakowiak, J. Influence of the ionic strength on the amyloid fibrillogenesis of hen egg white lysozyme. Int. J. Biol. Macromol. 2019, 121, 63–70. [Google Scholar] [CrossRef]
- Sneideris, T.; Darguzis, D.; Botyriute, A.; Grigaliunas, M.; Winter, R.; Smirnovas, V. pH-Driven Polymorphism of Insulin Amyloid-Like Fibrils. PLoS ONE 2015, 10, e0136602. [Google Scholar] [CrossRef]
- Lee, C.F.; Bird, S.; Shaw, M.; Jean, L.; Vaux, D.J. Combined effects of agitation, macromolecular crowding, and interfaces on amyloidogenesis. J. Biol. Chem. 2012, 287, 38006–38019. [Google Scholar] [CrossRef]
- Feng, Z.; Li, Y.; Bai, Y. Elevated temperatures accelerate the formation of toxic amyloid fibrils of hen egg-white lysozyme. Vet. Med. Sci. 2021, 7, 1938–1947. [Google Scholar] [CrossRef]
- Wang, F.; Wang, X.; Orrú, C.D.; Groveman, B.R.; Surewicz, K.; Abskharon, R.; Imamura, M.; Yokoyama, T.; Kim, Y.S.; Vander Stel, K.J.; et al. Self-propagating, protease-resistant, recombinant prion protein conformers with or without in vivo pathogenicity. PLoS Pathog. 2017, 13, e1006491. [Google Scholar] [CrossRef] [PubMed]
- Whittingham, J.L.; Scott, D.J.; Chance, K.; Wilson, A.; Finch, J.; Brange, J.; Guy Dodson, G. Insulin at pH 2: Structural analysis of the conditions promoting insulin fibre formation. J. Mol. Biol. 2002, 318, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Nettleton, E.J.; Tito, P.; Sunde, M.; Bouchard, M.; Dobson, C.M.; Robinson, C.V. Characterization of the Oligomeric States of Insulin in Self-Assembly and Amyloid Fibril Formation by Mass Spectrometry. Biophys. J. 2000, 79, 1053–1065. [Google Scholar] [CrossRef] [PubMed]
- Tokarz, V.L.; MacDonald, P.E.; Klip, A. The cell biology of systemic insulin function. J. Cell Biol. 2018, 217, 2273–2289. [Google Scholar] [CrossRef]
- Chiti, F.; Dobson, C.M. Protein Misfolding, Amyloid Formation, and Human Disease: A Summary of Progress over the Last Decade. Annu. Rev. Biochem. 2017, 86, 27–68. [Google Scholar] [CrossRef]
- Foderà, V.; Librizzi, F.; Groenning, M.; Van De Weert, M.; Leone, M. Secondary nucleation and accessible surface in insulin amyloid fibril formation. J. Phys. Chem. B 2008, 112, 3853–3858. [Google Scholar] [CrossRef]
- Noormägi, A.; Valmsen, K.; Tõugu, V.; Palumaa, P. Insulin Fibrillization at Acidic and Physiological pH Values is Controlled by Different Molecular Mechanisms. Protein J. 2015, 34, 398–403. [Google Scholar] [CrossRef]
- Jayamani, J.; Shanmugam, G. Gallic acid, one of the components in many plant tissues, is a potential inhibitor for insulin amyloid fibril formation. Eur. J. Med. Chem. 2014, 85, 352–358. [Google Scholar] [CrossRef]
- Nie, R.; Zhu, W.; Peng, J.; Ge, Z.; Li, C. A-type dimeric epigallocatechin-3-gallate (EGCG) is a more potent inhibitor against the formation of insulin amyloid fibril than EGCG monomer. Biochimie 2016, 125, 204–212. [Google Scholar] [CrossRef]
- Gong, H.; He, Z.; Peng, A.; Zhang, X.; Cheng, B.; Sun, Y.; Zheng, L.; Huang, K. Effects of several quinones on insulin aggregation. Sci. Rep. 2015, 4, 5648. [Google Scholar] [CrossRef]
- Deckert-Gaudig, T.; Deckert, V. High resolution spectroscopy reveals fibrillation inhibition pathways of insulin. Sci. Rep. 2016, 6, 39622. [Google Scholar] [CrossRef]
- Katebi, B.; Mahdavimehr, M.; Meratan, A.A.; Ghasemi, A.; Nemat-Gorgani, M. Protective effects of silibinin on insulin amyloid fibrillation, cytotoxicity and mitochondrial membrane damage. Arch. Biochem. Biophys. 2018, 659, 22–32. [Google Scholar] [CrossRef]
- Herranz-Trillo, F.; Groenning, M.; van Maarschalkerweerd, A.; Tauler, R.; Vestergaard, B.; Bernadó, P. Structural Analysis of Multi-Component Amyloid Systems by Chemometric SAXS Data Decomposition. Structure 2017, 25, 5–15. [Google Scholar] [CrossRef]
- Nie, R.; Zhu, W.; Peng, J.; Ge, Z.; Li, C. Comparison of disaggregative effect of A-type EGCG dimer and EGCG monomer on the preformed bovine insulin amyloid fibrils. Biophys. Chem. 2017, 230, 1–9. [Google Scholar] [CrossRef]
- Zheng, Q.; Lazo, N.D. Mechanistic Studies of the Inhibition of Insulin Fibril Formation by Rosmarinic Acid. J. Phys. Chem. B 2018, 122, 2323–2331. [Google Scholar] [CrossRef]
- Nusrat, S.; Zaman, M.; Masroor, A.; Siddqi, M.K.; Zaidi, N.; Neelofar, K.; Abdelhameed, A.S.; Khan, R.H. Deciphering the enhanced inhibitory, disaggregating and cytoprotective potential of promethazine towards amyloid fibrillation. Int. J. Biol. Macromol. 2018, 106, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Tsuhara, S.; Tamura, A.; Chatani, E. A specific form of prefibrillar aggregates that functions as a precursor of amyloid nucleation. Sci. Rep. 2018, 8, 62. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Jing, B.; Sorci, M.; Belfort, G.; Zhu, Y. Accelerated insulin aggregation under alternating current electric fields: Relevance to amyloid kinetics. Biomicrofluidics 2015, 9, 044123. [Google Scholar] [CrossRef]
- Saithong, T.; Thilavech, T.; Adisakwattana, S. Cyanidin-3-rutinoside reduces insulin fibrillation and attenuates insulin fibrils-induced oxidative hemolysis of human erythrocytes. Int. J. Biol. Macromol. 2018, 113, 259–268. [Google Scholar] [CrossRef]
- Silva, A.; Sárkány, Z.; Fraga, J.S.; Taboada, P.; Macedo-Ribeiro, S.; Martins, P.M. Probing the occurrence of soluble oligomers through amyloid aggregation scaling laws. Biomolecules 2018, 8, 108. [Google Scholar] [CrossRef] [PubMed]
- Flint, Z.; Grannemann, H.; Baffour, K.; Koti, N.; Taylor, E.; Grier, E.; Sutton, C.; Johnson, D.; Dandawate, P.; Patel, R.; et al. Mechanistic Insights Behind the Self-Assembly of Human Insulin under the Influence of Surface-Engineered Gold Nanoparticles. ACS Chem. Neurosci. 2024, 15, 2359–2371. [Google Scholar] [CrossRef]
- Joshi, R.; Zhaliazka, K.; Holman, A.P.; Kurouski, D. Elucidation of the Role of Lipids in Late Endosomes on the Aggregation of Insulin. ACS Chem. Neurosci. 2023, 14, 3551–3559. [Google Scholar] [CrossRef] [PubMed]
- Zazeri, G.; Povinelli, A.P.R.; Pavan, N.M.; Jones, A.M.; Ximenes, V.F. Solvent-Induced Lag Phase during the Formation of Lysozyme Amyloid Fibrils Triggered by Sodium Dodecyl Sulfate: Biophysical Experimental and In Silico Study of Solvent Effects. Molecules 2023, 28, 6891. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.M.; Malik, A.; Alresaini, S.M. Molecular mechanism of insulin aggregation in the presence of a cationic surfactant. Int. J. Biol. Macromol. 2023, 230, 123370. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Shah, M.; Saraogi, I. Molecular Aspects of Insulin Aggregation and Various Therapeutic Interventions. ACS Bio Med Chem Au 2022, 2, 205–221. [Google Scholar] [CrossRef]
- Ziaunys, M.; Mikalauskaite, K.; Sakalauskas, A.; Smirnovas, V. Interplay between epigallocatechin-3-gallate and ionic strength during amyloid aggregation. PeerJ 2021, 9, e12381. [Google Scholar] [CrossRef]
- Banga, A.K. Therapeutic Peptides and Proteins; CRC Press: Boca Raton, FL, USA, 2015; ISBN 9780429099908. [Google Scholar]
- Grudzielanek, S.; Smirnovas, V.; Winter, R. Solvation-assisted Pressure Tuning of Insulin Fibrillation: From Novel Aggregation Pathways to Biotechnological Applications. J. Mol. Biol. 2006, 356, 497–509. [Google Scholar] [CrossRef]
- Barth, A. Infrared spectroscopy of proteins. Biochim. Biophys. Acta-Bioenerg. 2007, 1767, 1073–1101. [Google Scholar] [CrossRef]
- Sakalauskas, A.; Ziaunys, M.; Smirnovas, V. Concentration-dependent polymorphism of insulin amyloid fibrils. PeerJ 2019, 7, e8208. [Google Scholar] [CrossRef]
- Ziaunys, M.; Sakalauskas, A.; Mikalauskaite, K.; Smirnovas, V. Exploring the occurrence of thioflavin-T-positive insulin amyloid aggregation intermediates. PeerJ 2021, 9, e10918. [Google Scholar] [CrossRef]
- Mansoury, M.; Hamed, M.; Karmustaji, R.; Al Hannan, F.; Safrany, S.T. The edge effect: A global problem. The trouble with culturing cells in 96-well plates. Biochem. Biophys. Rep. 2021, 26, 100987. [Google Scholar] [CrossRef]
- Mikalauskaite, K.; Ziaunys, M.; Sneideris, T.; Smirnovas, V. Effect of Ionic Strength on Thioflavin-T Affinity to Amyloid Fibrils and Its Fluorescence Intensity. Int. J. Mol. Sci. 2020, 21, 8916. [Google Scholar] [CrossRef] [PubMed]
- Mikalauskaite, K.; Ziaunys, M.; Smirnovas, V. Lysozyme Amyloid Fibril Structural Variability Dependence on Initial Protein Folding State. Int. J. Mol. Sci. 2022, 23, 5421. [Google Scholar] [CrossRef] [PubMed]
- Ziaunys, M.; Smirnovas, V. Additional Thioflavin-T Binding Mode in Insulin Fibril Inner Core Region. J. Phys. Chem. B 2019, 123, 8727–8732. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziaunys, M.; Mikalauskaite, K.; Sakalauskas, A.; Smirnovas, V. Study of Insulin Aggregation and Fibril Structure under Different Environmental Conditions. Int. J. Mol. Sci. 2024, 25, 9406. https://doi.org/10.3390/ijms25179406
Ziaunys M, Mikalauskaite K, Sakalauskas A, Smirnovas V. Study of Insulin Aggregation and Fibril Structure under Different Environmental Conditions. International Journal of Molecular Sciences. 2024; 25(17):9406. https://doi.org/10.3390/ijms25179406
Chicago/Turabian StyleZiaunys, Mantas, Kamile Mikalauskaite, Andrius Sakalauskas, and Vytautas Smirnovas. 2024. "Study of Insulin Aggregation and Fibril Structure under Different Environmental Conditions" International Journal of Molecular Sciences 25, no. 17: 9406. https://doi.org/10.3390/ijms25179406
APA StyleZiaunys, M., Mikalauskaite, K., Sakalauskas, A., & Smirnovas, V. (2024). Study of Insulin Aggregation and Fibril Structure under Different Environmental Conditions. International Journal of Molecular Sciences, 25(17), 9406. https://doi.org/10.3390/ijms25179406