
Function and Global Regulation of Type III Secretion System and Flagella in Entomopathogenic Nematode Symbiotic Bacteria
Abstract
1. Introduction
2. Results
2.1. Flagella Play a Pivotal Role in Governing Both Swarming Motility and Biofilm Formation in Xenorhabdus
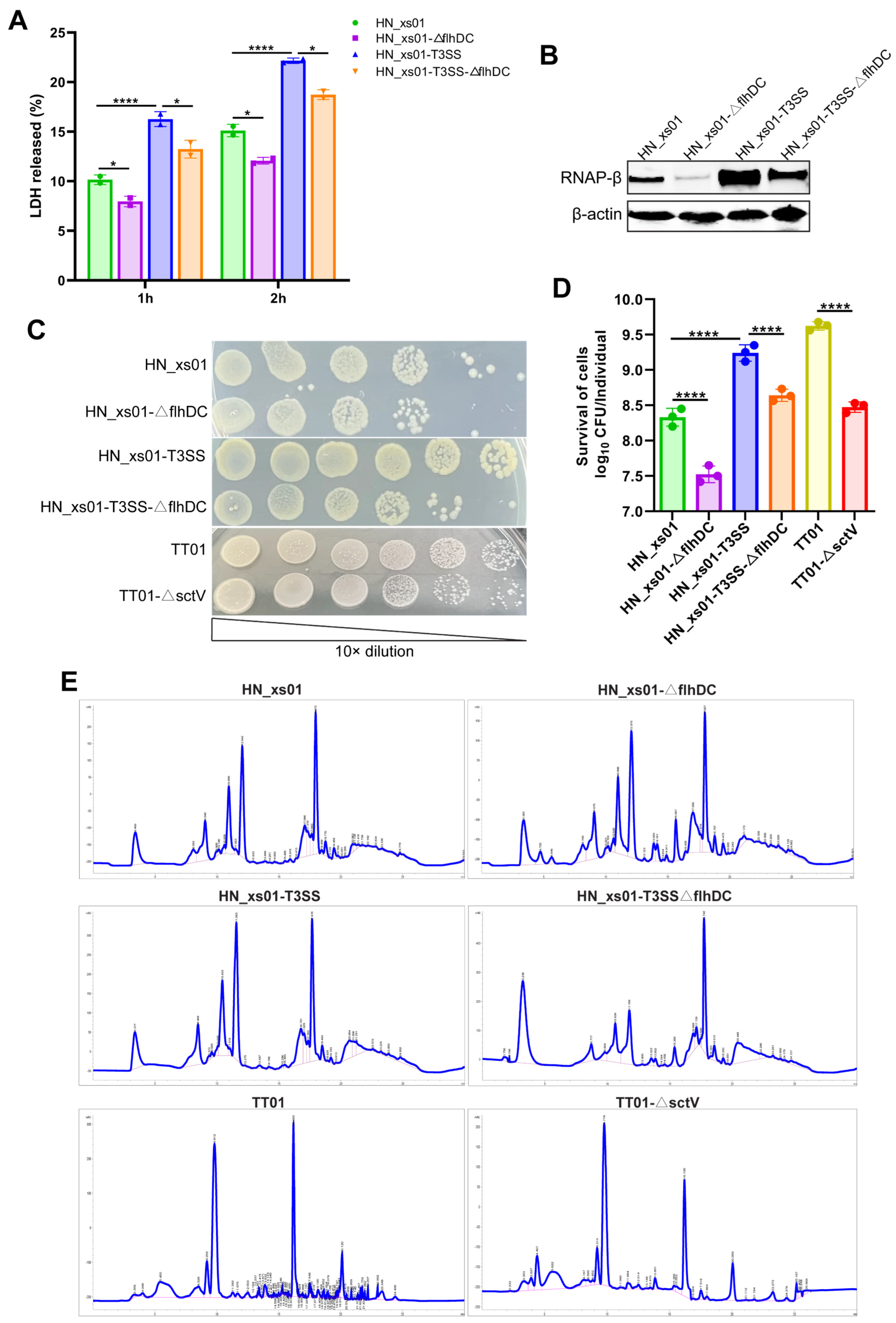
2.2. T3SS and Flagella Collaboratively Facilitate the Invasion and Colonization of ENS While Leaving Secondary Metabolites Unaffected
2.3. Both T3SS and Flagella Play Crucial Roles in Mediating the Chemotaxis of ENS
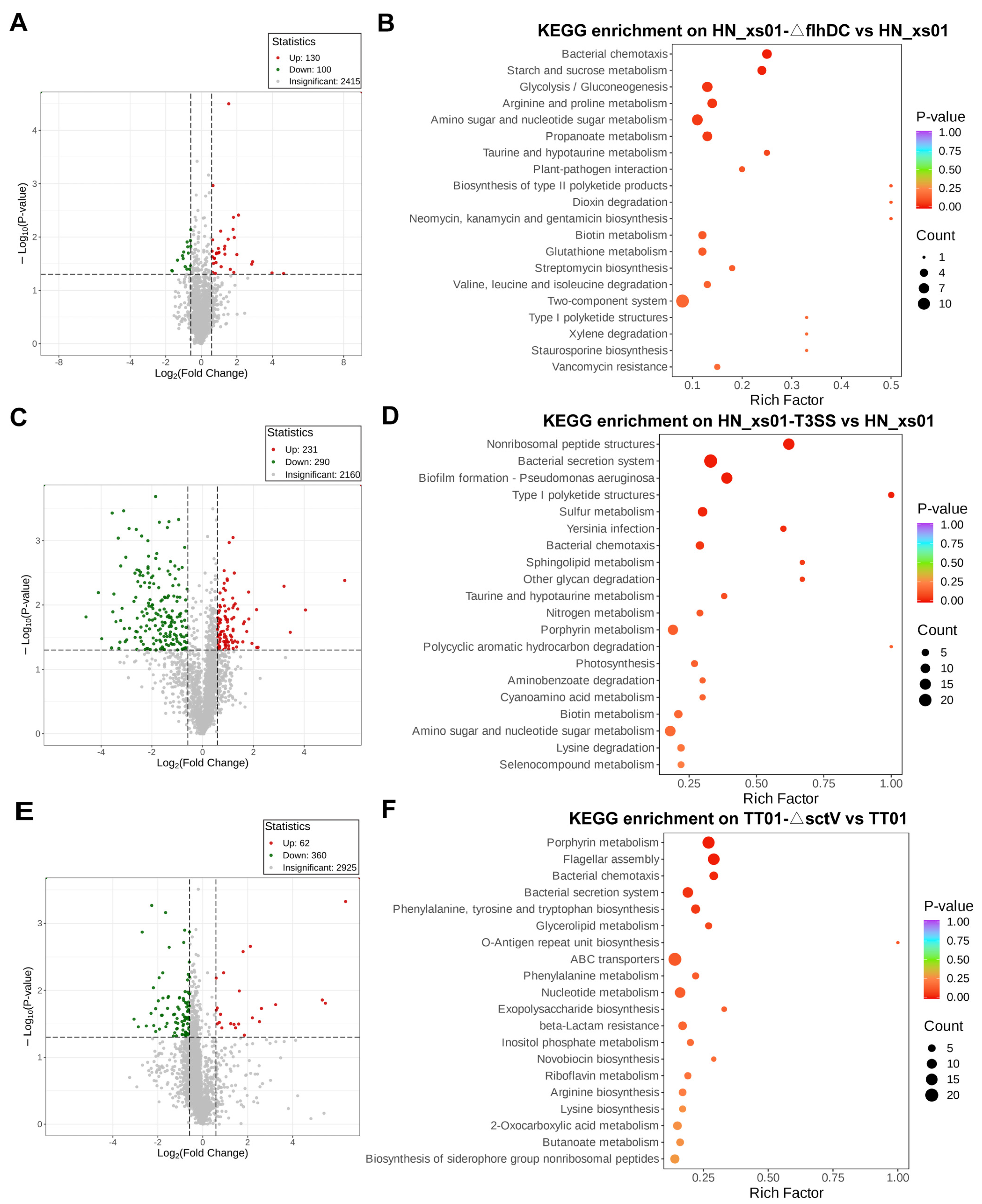
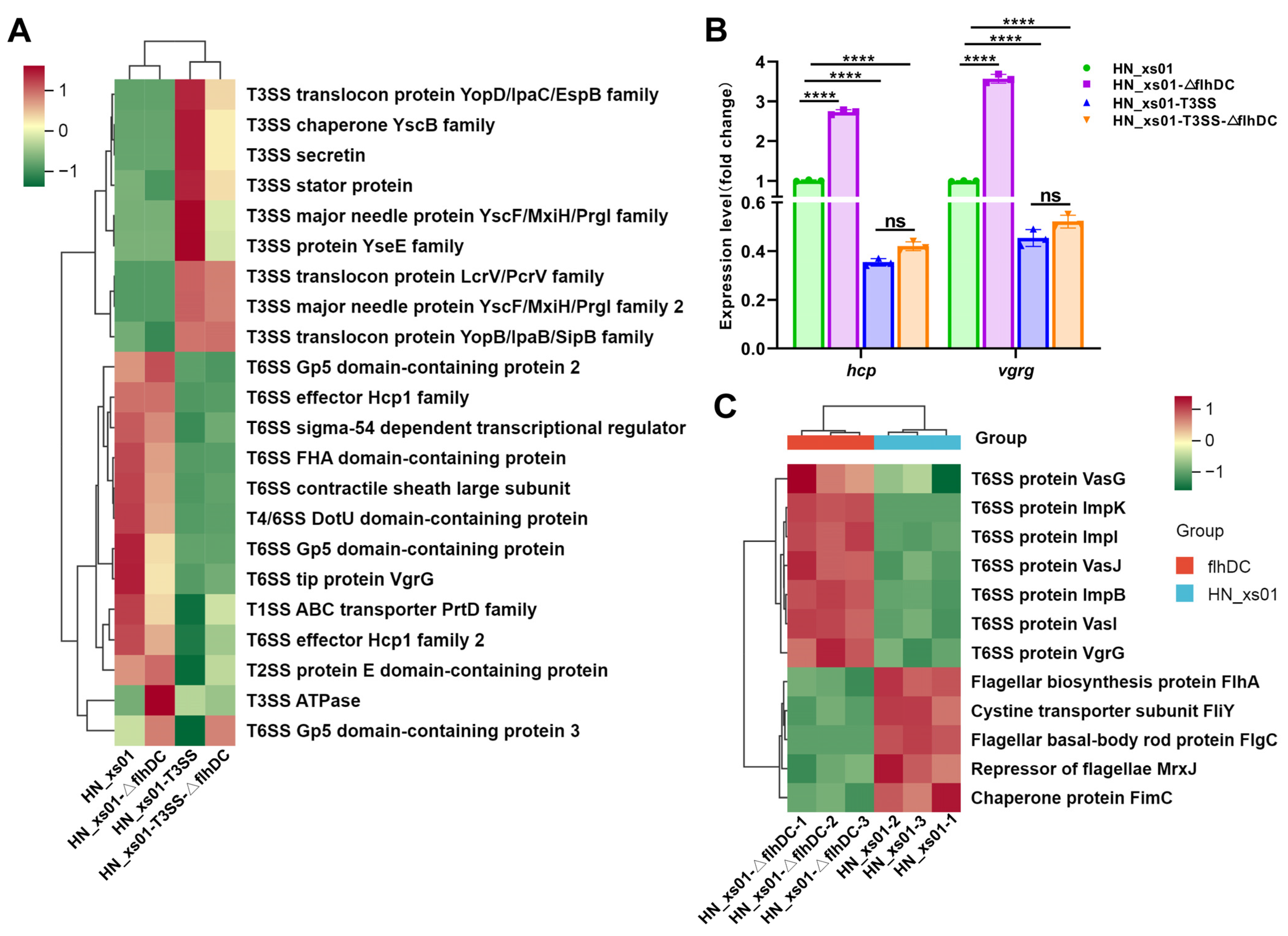
2.4. T3SS and Flagella Inhibited the Expression of T6SS
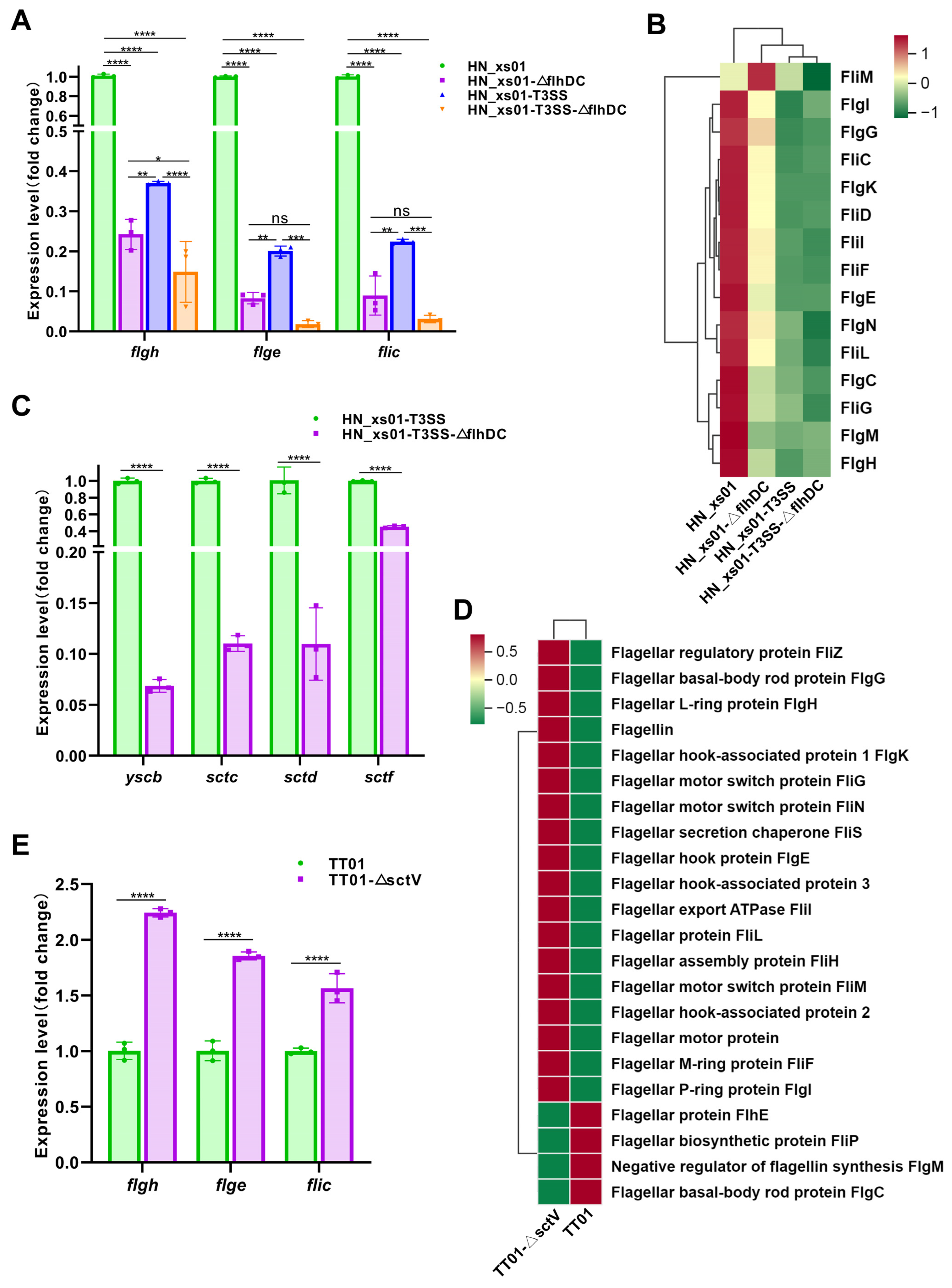
2.5. Flagella Promote the Assembly of the T3SS, While the T3SS Negatively Regulates the Expression of Flagellin
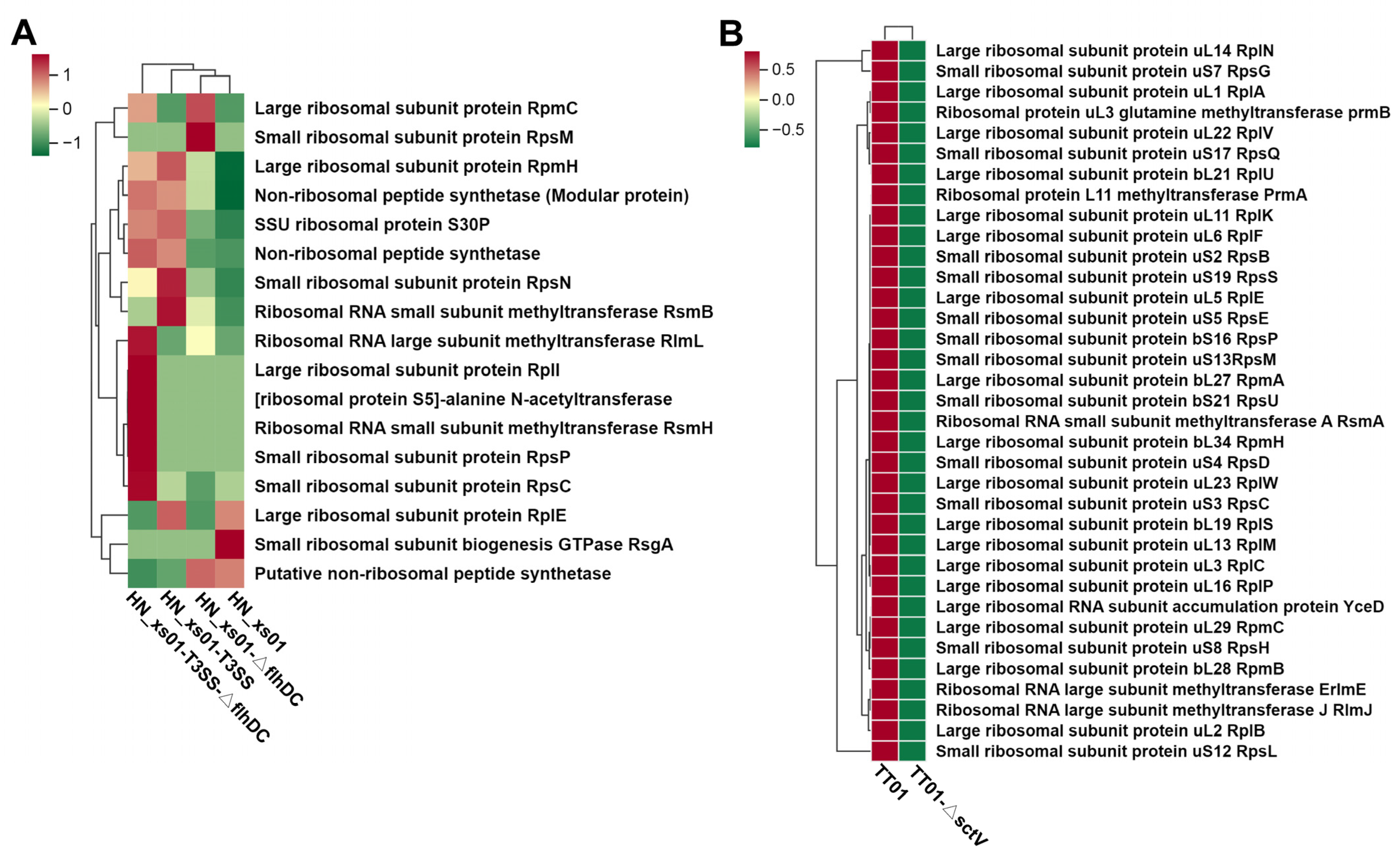
2.6. T3SS Enhances the Expression of Ribosome-Associated Proteins
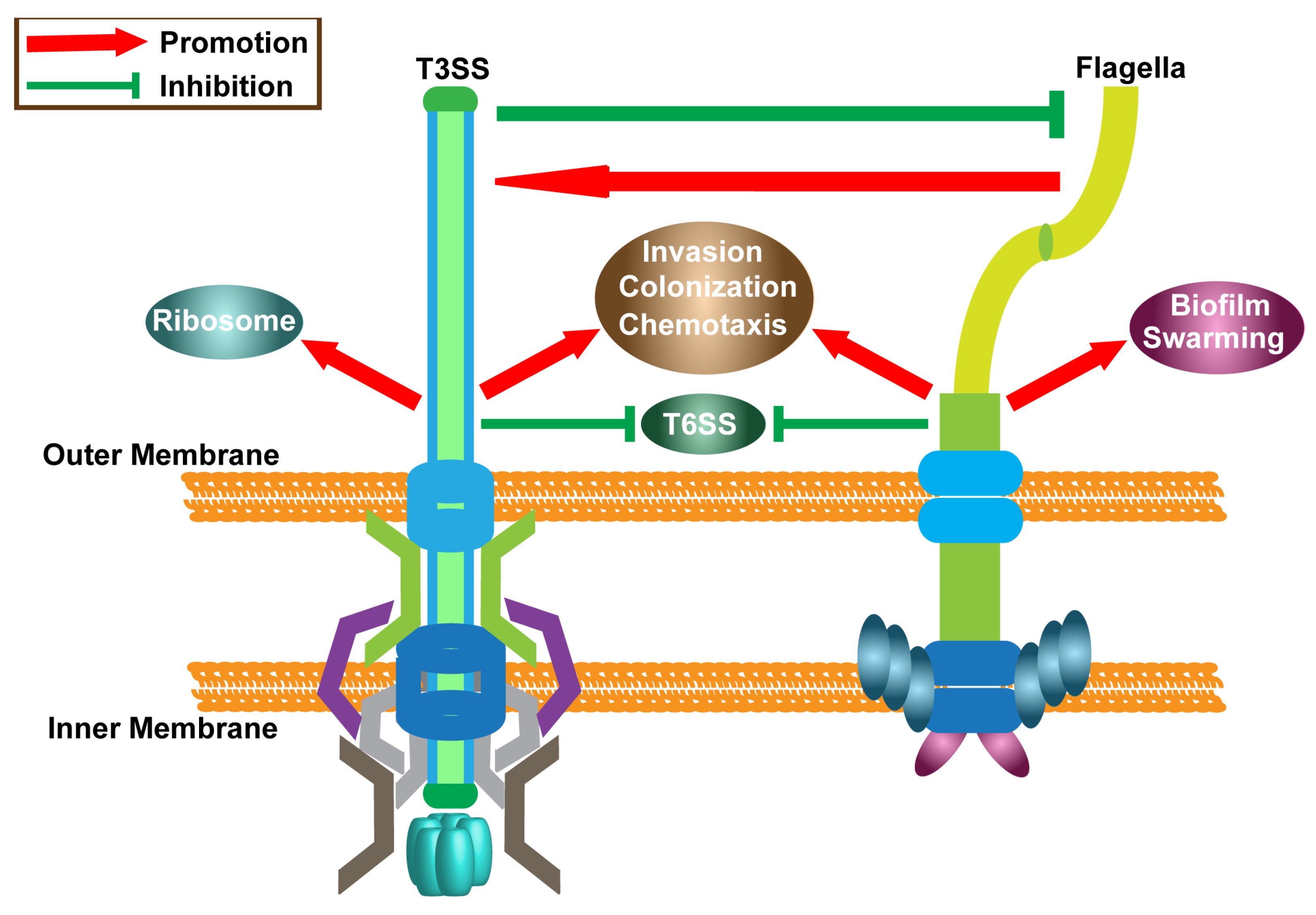
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Cell Lines, and Their Growth Conditions
4.2. Motility Assay
4.3. Biofilm Assays
4.4. LDH Assays
4.5. Immunoblotting
4.6. Detection of ENS Colonization Ability
4.7. HPLC Assays
4.8. Proteomics Assays
4.9. Quantitative Real-Time PCR (RT-PCR) Assays
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Givaudan, A.; Lanois, A. flhDC, the flagellar master operon of Xenorhabdus nematophilus_ requirement for motility, lipolysis, extracellular hemolysis, and full virulence in insects. J. Bacteriol. 2000, 182, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Shulman, A.; Yair, Y.; Biran, D.; Sura, T.; Otto, A.; Gophna, U.; Becher, D.; Hecker, M.; Ron, E.Z. The Escherichia coli Type III Secretion System 2 Has a Global Effect on Cell Surface. mBio 2018, 9, e01070-18. [Google Scholar] [CrossRef] [PubMed]
- Ashida, H.; Suzuki, T.; Sasakawa, C. Sasakawa Shigella infection and host cell death: A double-edged sword for the host and pathogen survival. Curr. Opin. Microbiol. 2021, 59, 1–7. [Google Scholar] [CrossRef]
- Bai, F.; Li, Z.; Umezawa, A.; Terada, N.; Jin, S. Bacterial type III secretion system as a protein delivery tool for a broad range of biomedical applications. Biotechnol. Adv. 2018, 36, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Beeby, M.; Ferreira, J.L.; Tripp, P.; Albers, S.-V.; Mitchell, D.R. Propulsive nanomachines: The convergent evolution of archaella, flagella and cilia. FEMS Microbiol. Rev. 2020, 44, 253–304. [Google Scholar] [CrossRef] [PubMed]
- Blocker, A.; Komoriya, K.; Aizawa, S.-I. Type III secretion systems and bacterial flagella: Insights into their function from structural similarities. Proc. Natl. Acad. Sci. USA 2003, 100, 3027–3030. [Google Scholar] [CrossRef]
- Bouteiller, M.; Dupont, C.; Bourigault, Y.; Latour, X.; Barbey, C.; Konto-Ghiorghi, Y.; Merieau, A. Pseudomonas Flagella: Generalities and Specificities. Int. J. Mol. Sci. 2021, 22, 3337. [Google Scholar] [CrossRef]
- Brugirard-Ricaud, K.; Duchaud, E.; Givaudan, A.; Girard, P.A.; Kunst, F.; Boemare, N.; Brehelin, M.; Zumbihl, R. Site-specific antiphagocytic function of the Photorhabdus luminescens type III secretion system during insect colonization. Cell Microbiol. 2005, 7, 363–371. [Google Scholar] [CrossRef]
- Büttner, D.; Bonas, U. Common infection strategies of plant and animal pathogenic bacteria. Curr. Opin. Plant Biol. 2003, 6, 312–319. [Google Scholar] [CrossRef]
- Caimano, M.J.; Groshong, A.M.; Belperron, A.; Mao, J.; Hawley, K.L.; Luthra, A.; Graham, D.E.; Earnhart, C.G.; Marconi, R.T.; Bockenstedt, L.K.; et al. The RpoS Gatekeeper in Borrelia burgdorferi: An Invariant Regulatory Scheme That Promotes Spirochete Persistence in Reservoir Hosts and Niche Diversity. Front. Microbiol. 2019, 10, 1923. [Google Scholar] [CrossRef]
- Cohen, H.; Baram, N.; Fridman, C.M.; Edry-Botzer, L.; Salomon, D.; Gerlic, M. Post-phagocytosis activation of NLRP3 inflammasome by two novel T6SS effectors. eLife 2022, 11, e82766. [Google Scholar] [CrossRef] [PubMed]
- Dadashi, M.; Chen, L.; Nasimian, A.; Ghavami, S.; Duan, K. Putative RNA Ligase RtcB Affects the Switch between T6SS and T3SS in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2021, 22, 12561. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Marshall, N.C.; Rowland, J.L.; McCoy, J.M.; Worrall, L.J.; Santos, A.S.; Strynadka, N.C.J.; Finlay, B.B. Assembly, structure, function and regulation of type III secretion systems. Nature reviews. Microbiology 2017, 15, 323–337. [Google Scholar] [CrossRef] [PubMed]
- A Easom, C.; Clarke, D.J. Motility is required for the competitive fitness of entomopathogenic Photorhabdus luminescens during insect infection. BMC Microbiol. 2008, 8, 168. [Google Scholar] [CrossRef] [PubMed]
- Fei, N.; Ji, W.; Yang, L.; Yu, C.; Qiao, P.; Yan, J.; Guan, W.; Yang, Y.; Zhao, T. Hcp of the Type VI Secretion System (T6SS) in Acidovorax citrulli Group II Strain Aac5 Has a Dual Role as a Core Structural Protein and an Effector Protein in Colonization, Growth Ability, Competition, Biofilm Formation, and Ferric Iron Absorption. Int. J. Mol. Sci. 2022, 23, 9632. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Bi, W.; Chen, S.; Zhu, J.; Liu, X. Regulatory function of sigma factors RpoS/RpoN in adaptation and spoilage potential of Shewanella baltica. Food Microbiol. 2021, 97, 103755. [Google Scholar] [CrossRef] [PubMed]
- Gilzer, D.; Baum, E.; Lieske, N.; Kowal, J.L.; Niemann, H.H. Crystals of SctV from different species reveal variable symmetry for the cytosolic domain of the type III secretion system export gate. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2022, 78, 386–394. [Google Scholar] [CrossRef]
- Gilzer, D.; Schreiner, M.; Niemann, H.H. Direct interaction of a chaperone-bound type III secretion substrate with the export gate. Nat. Commun. 2022, 13, 2858. [Google Scholar] [CrossRef]
- Givaudan, A.; Lanois, A. Flagellar Regulation and Virulence in the Entomopathogenic Bacteria—Xenorhabdus nematophila and Photorhabdus luminescens. Curr. Top. Microbiol. Immunol. 2016, 402, 39–51. [Google Scholar] [CrossRef]
- Goodrich-Blair, H.; Clarke, D.J. Mutualism and pathogenesis in Xenorhabdus and Photorhabdus: Two roads to the same destination. Mol. Microbiol. 2007, 64, 260–268. [Google Scholar] [CrossRef]
- Grassmann, A.A.; Tokarz, R.; Golino, C.; McLain, M.A.; Groshong, A.M.; Radolf, J.D.; Caimano, M.J. BosR and PlzA reciprocally regulate RpoS function to sustain Borrelia burgdorferi in ticks and mammals. J. Clin. Investig. 2023, 133, 66710. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Xiao, X.; Xu, S.; Gao, F.; Wang, J.; Wang, T.; Song, Y.; Pan, J.; Shen, X.; Wang, Y. Roles of RpoS in Yersinia pseudotuberculosis stress survival, motility, biofilm formation and type VI secretion system expression. J. Microbiol. 2015, 53, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Spöring, I.; Felgner, S.; Preuße, M.; Eckweiler, D.; Rohde, M.; Häussler, S.; Weiss, S.; Erhardt, M. Regulation of Flagellum Biosynthesis in Response to Cell Envelope Stress in Salmonella enterica Serovar Typhimurium. mBio 2018, 9, e00736-17. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Tachiyama, S.; Zhou, X.; Mathias, R.A.; Bonny, S.Q.; Khan, M.F.; Xin, Y.; Roujeinikova, A.; Liu, J.; Ottemann, K.M. Bacterial flagella hijack type IV pili proteins to control motility. Proc. Natl. Acad. Sci. USA 2024, 121, e2317452121. [Google Scholar] [CrossRef]
- Liu, X.; Zhuo, S.; Jing, X.; Yuan, Y.; Rensing, C.; Zhou, S. Flagella act as Geobacter biofilm scaffolds to stabilize biofilm and facilitate extracellular electron transfer. Biosens. Bioelectron. 2019, 146, 111748. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Pei, T.-T.; Li, H.; Zheng, H.-Y.; Luo, H.; Cui, Y.; Tang, M.-X.; Zhao, Y.-J.; Xu, P.; Dong, T. VgrG-dependent effectors and chaperones modulate the assembly of the type VI secretion system. PLoS Pathog. 2021, 17, e1010116. [Google Scholar] [CrossRef] [PubMed]
- Majewski, D.D.; Lyons, B.J.; Atkinson, C.E.; Strynadka, N.C. Cryo-EM analysis of the SctV cytosolic domain from the enteropathogenic E. coli T3SS injectisome. J. Struct. Biol. 2020, 212, 107660. [Google Scholar] [CrossRef] [PubMed]
- Meir, A.; Macé, K.; Vegunta, Y.; Williams, S.M.; Waksman, G. Substrate recruitment mechanism by gram-negative type III, IV, and VI bacterial injectisomes. Trends Microbiol. 2023, 31, 916–932. [Google Scholar] [CrossRef]
- Michaels, B.; Tisa, L.S. Swarming motility by Photorhabdus temperata is influenced by environmental conditions and uses the same flagella as that used in swimming motilityThis is scientific contribution No. 2431 from the New Hampshire Agricultural Experiment Station. Can. J. Microbiol. 2011, 57, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Feria, J.M.; Valvano, M.A. An Overview of Anti-Eukaryotic T6SS Effectors. Front. Cell Infect. Microbiol. 2020, 10, 584751. [Google Scholar] [CrossRef]
- Nakamura, S.; Minamino, T. Flagella-Driven Motility of Bacteria. Biomolecules 2019, 9, 279. [Google Scholar] [CrossRef] [PubMed]
- Nans, A.; Kudryashev, M.; Saibil, H.R.; Hayward, R.D. Structure of a bacterial type III secretion system in contact with a host membrane in situ. Nat. Commun. 2015, 6, 10114. [Google Scholar] [CrossRef] [PubMed]
- Nielsen-LeRoux, C.; Gaudriault, S.; Ramarao, N.; Lereclus, D.; Givaudan, A. How the insect pathogen bacteria Bacillus thuringiensis and Xenorhabdus/Photorhabdus occupy their hosts. Curr. Opin. Microbiol. 2012, 15, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Österberg, S.; del Peso-Santos, T.; Shingler, V. Regulation of Alternative Sigma Factor Use. Annu. Rev. Microbiol. 2011, 65, 37–55. [Google Scholar] [CrossRef] [PubMed]
- Pendergrass, H.A.; May, A.E. Delivery of Heterologous Proteins, Enzymes, and Antigens via the Bacterial Type III Secretion System. Microorganisms 2020, 8, 777. [Google Scholar] [CrossRef] [PubMed]
- Pinaud, L.; Sansonetti, P.J.; Phalipon, A. Host Cell Targeting by Enteropathogenic Bacteria T3SS Effectors. Trends Microbiol. 2018, 26, 266–283. [Google Scholar] [CrossRef] [PubMed]
- Plum, M.T.W.; Cheung, H.C.; Iscar, P.R.; Chen, Y.; Gan, Y.-H.; Basler, M.; Plum, M.T.W.; Cheung, H.C.; Iscar, P.R.; Chen, Y.; et al. Burkholderia thailandensis uses a type VI secretion system to lyse protrusions without triggering host cell responses. Cell Host Microbe 2024, 32, 676–692.e5. [Google Scholar] [CrossRef] [PubMed]
- Pothula, R.; Lee, M.-W.; Stock, S.P. Type 6 secretion system components hcp and vgrG support mutualistic partnership between Xenorhabdus bovienii symbiont and Steinernema jollieti host. J. Invertebr. Pathol. 2023, 198, 107925. [Google Scholar] [CrossRef]
- Potvin, E.; Sanschagrin, F.; Levesque, R.C. Sigma factors inPseudomonas aeruginosa. FEMS Microbiol. Rev. 2008, 32, 38–55. [Google Scholar] [CrossRef]
- Ruano-Gallego, D.; Sanchez-Garrido, J.; Kozik, Z.; Núñez-Berrueco, E.; Cepeda-Molero, M.; Mullineaux-Sanders, C.; Baghshomali, Y.N.; Slater, S.L.; Wagner, N.; Glegola-Madejska, I.; et al. Type III secretion system effectors form robust and flexible intracellular virulence networks. Science 2021, 371, 1122. [Google Scholar] [CrossRef]
- Ruiz, V.M.R.; Ramirez, J.; Naseer, N.; Palacio, N.M.; Siddarthan, I.J.; Yan, B.M.; Boyer, M.A.; Pensinger, D.A.; Sauer, J.-D.; Shin, S. Broad detection of bacterial type III secretion system and flagellin proteins by the human NAIP_NLRC4 inflammasome. Proc. Natl. Acad. Sci. USA 2017, 114, 13242–13247. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Garrido, J.; Ruano-Gallego, D.; Choudhary, J.S.; Frankel, G. The type III secretion system effector network hypothesis. Trends Microbiol. 2022, 30, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Pan, J.; Yang, Y.; Zhang, Z.; Cui, R.; Jia, S.; Wang, Z.; Yang, C.; Xu, L.; Dong, T.G.; et al. Contact-independent killing mediated by a T6SS effector with intrinsic cell-entry properties. Nat. Commun. 2021, 12, 423. [Google Scholar] [CrossRef] [PubMed]
- Soria-Bustos, J.; Saldaña-Ahuactzi, Z.; Samadder, P.; Yañez-Santos, J.A.; Laguna, Y.M.; Cedillo-Ramírez, M.L.; Girón, J.A. The Assembly of Flagella in Enteropathogenic Escherichia coli Requires the Presence of a Functional Type III Secretion System. Int. J. Mol. Sci. 2022, 23, 13705. [Google Scholar] [CrossRef]
- Subramanian, S.; Kearns, D.B. Functional Regulators of Bacterial Flagella. Annu. Rev. Microbiol. 2019, 73, 225–246. [Google Scholar] [CrossRef] [PubMed]
- Tobias, N.J.; Mishra, B.; Gupta, D.K.; Sharma, R.; Thines, M.; Stinear, T.P.; Bode, H.B. Genome comparisons provide insights into the role of secondary metabolites in the pathogenic phase of the Photorhabdus life cycle. BMC Genom. 2016, 17, 537. [Google Scholar] [CrossRef] [PubMed]
- Viducic, D.; Murakami, K.; Amoh, T.; Ono, T.; Miyake, Y. RpoN Promotes Pseudomonas aeruginosa Survival in the Presence of Tobramycin. Front. Microbiol. 2017, 8, 839. [Google Scholar] [CrossRef]
- Wang, Y.; Zeng, M.; Xia, L.; Olovo, C.V.; Su, Z.; Zhang, Y. Bacterial strategies for immune systems—Role of the type VI secretion system. Int. Immunopharmacol. 2023, 114, 109550. [Google Scholar] [CrossRef] [PubMed]
- Westerman, T.L.; McClelland, M.; Elfenbein, J.R. YeiE Regulates Motility and Gut Colonization in Salmonella enterica Serotype Typhimurium. mBio 2021, 12, e03680-20. [Google Scholar] [CrossRef]
- Liu, X.; Lertsethtakarn, P.; Mariscal, V.T.; Yildiz, F.; Ottemann, K.M. Counterclockwise rotation of the flagellum promotes biofilm initiation in Helicobacter pylori. mBio 2024, 3, e00440-24. [Google Scholar] [CrossRef]
- Yan, J.; Guo, X.; Li, J.; Li, Y.; Sun, H.; Li, A.; Cao, B. RpoN is required for the motility and contributes to the killing ability of Plesiomonas shigelloides. BMC Microbiol. 2022, 22, 299. [Google Scholar] [CrossRef] [PubMed]
- Kan, Y.; Zhang, Y.; Lin, W.; Dong, T. Differential plant cell responses to Acidovorax citrulli T3SS and T6SS reveal an effective strategy for controlling plant-associated pathogens. mBio 2023, 14, e00459-23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Riba, A.; Nietschke, M.; Torow, N.; Repnik, U.; Pütz, A.; Fulde, M.; Dupont, A.; Hensel, M.; Hornef, M. Minimal SPI1-T3SS effector requirement for Salmonella enterocyte invasion and intracellular proliferation in vivo. PLoS Pathog. 2018, 14, e1006925. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.-H.; Shaw, J.G. Cross-Talk between the Aeromonas hydrophila Type III Secretion System and Lateral Flagella System. Front. Microbiol. 2016, 7, 1434. [Google Scholar] [CrossRef]
- Zhu, S.; Gao, B. Bacterial Flagella Loss under Starvation. Trends Microbiol. 2020, 28, 785–788. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Genotype or Description | Source |
|---|---|---|
| Strains | ||
| HN_xs01 | wild-type X. stockiae HN_xs01 | laboratory preservation |
| HN_xs01-ΔflhDC | flagella inactivation (Cmr) | this study |
| HN_xs01-T3SS | heterogeneous synthesis of the T3SS (Ermr) | this study |
| HN_xs01-T3SS-ΔflhDC | T3SS activity without flagella (Cmr Ermr) | this study |
| TT01 | wild-type P. luminescens TT01 | laboratory preservation |
| TT01–ΔsctV | T3SS inactivation (Cmr) | this study |
| Plasmids | ||
| pEVS107-T3SS | T3SS gene cluster (Ermr) | laboratory preservation |
| pRE112::flhDC | knockout flhDC (Cmr) | this study |
| pRE112::sctV | knockoutsctV (Cmr) | this study |
| Primer | Sequence | Information |
|---|---|---|
| X-16s-F | CAGATGGGATTAGCTAGTAG | the 16s of X. stockiae HN_xs01 |
| X-16s-R | TGCAATATTCCCCACTGCTG | the 16s of X. stockiae HN_xs01 |
| X-Flge-F | GATGGTTCAACCACTACGAC | the flge of X. stockiae HN_xs01 |
| X-Flge-R | AGAGTACGGTCTGAGTCCAT | the flge of X. stockiae HN_xs01 |
| X-Flgh-F | GTACTCGCATTGTCGGTACT | the flgh of X. stockiae HN_xs01 |
| X-Flgh-R | AACAGAACCATTCGGCGCTG | the flgh of X. stockiae HN_xs01 |
| X-Flic-F | CAACCTGTACAGCCATTGAG | the flic of X. stockiae HN_xs01 |
| X-Flic-R | TGGCAGCAACTCTAAGAGTG | the flic of X. stockiae HN_xs01 |
| X-Vgrg-F | CATCAATCACCACTTTCGCG | the vgrg of X. stockiae HN_xs01 |
| X-Vgrg-R | CGATAATGAATACCGGGTAC | the vgrg of X. stockiae HN_xs01 |
| X-Hcp-F | CTCTGCTGGATGTTCAACAC | the hcp of X. stockiae HN_xs01 |
| X-Hcp-R | GTATTGGGTGGTGACTTAC | the hcp of X. stockiae HN_xs01 |
| YscB-F | CAGTCGTTTAGGCCAGAAGC | the yscb of X. stockiae HN_xs01 |
| YscB-R | CTTAGCGGGCTGGATAACAC | the yscb of X. stockiae HN_xs01 |
| SctC-F | CTGCTTATAGCCAAGATCTG | the sctc of X. stockiae HN_xs01 |
| SctC-R | CTTGTCGCTGACAACCACTG | the sctc of X. stockiae HN_xs01 |
| SctD-F | GCACCTGATCTTAGAAGTCG | the sctd of X. stockiae HN_xs01 |
| SctD-R | CACAATAACGGGCCAACTTC | the sctd of X. stockiae HN_xs01 |
| SctF-F | TCCGCAACAACAGTGAATAC | the sctf of X. stockiae HN_xs01 |
| SctF-R | GCTAGCAAAGCTGGGTTATC | the sctf of X. stockiae HN_xs01 |
| P-16s-F | GAAGAAGCACCGGCTAACTC | the 16s of P. luminescens TT01 |
| P-16s-R | GATGCCATTCCCAGGTTGAG | the 16s of P. luminescens TT01 |
| P-Flge-F | CAACGACGACTAACCGAACA | the flge of P. luminescens TT01 |
| P-Flge-R | TGCGGTTCTCATCCATCTTG | the flge of P. luminescens TT01 |
| P-Flgh-F | CACCTTCTGTGGCAACTGCA | the flgh of P. luminescens TT01 |
| P-Flgh-R | CCTGCAATGTAATCGTCAGG | the flgh of P. luminescens TT01 |
| P-Flic-F | GAGCGTCTATCTTCTGGTCT | the flic of P. luminescens TT01 |
| P-Flic-R | CTGTGCAATAGAGATACCGT | the flic of P. luminescens TT01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Li, C.; Zhang, K.; Li, K.; Xie, J.; Peng, Y.; Quan, M.; Sun, Y.; Hu, Y.; Xia, L.; et al. Function and Global Regulation of Type III Secretion System and Flagella in Entomopathogenic Nematode Symbiotic Bacteria. Int. J. Mol. Sci. 2024, 25, 7579. https://doi.org/10.3390/ijms25147579
Huang X, Li C, Zhang K, Li K, Xie J, Peng Y, Quan M, Sun Y, Hu Y, Xia L, et al. Function and Global Regulation of Type III Secretion System and Flagella in Entomopathogenic Nematode Symbiotic Bacteria. International Journal of Molecular Sciences. 2024; 25(14):7579. https://doi.org/10.3390/ijms25147579
Chicago/Turabian StyleHuang, Xiyin, Chen Li, Ke Zhang, Kunyan Li, Jiajie Xie, Yuyuan Peng, Meifang Quan, Yunjun Sun, Yibo Hu, Liqiu Xia, and et al. 2024. "Function and Global Regulation of Type III Secretion System and Flagella in Entomopathogenic Nematode Symbiotic Bacteria" International Journal of Molecular Sciences 25, no. 14: 7579. https://doi.org/10.3390/ijms25147579
APA StyleHuang, X., Li, C., Zhang, K., Li, K., Xie, J., Peng, Y., Quan, M., Sun, Y., Hu, Y., Xia, L., & Hu, S. (2024). Function and Global Regulation of Type III Secretion System and Flagella in Entomopathogenic Nematode Symbiotic Bacteria. International Journal of Molecular Sciences, 25(14), 7579. https://doi.org/10.3390/ijms25147579