
Ex Vivo Analysis of Cell Differentiation, Oxidative Stress, Inflammation, and DNA Damage on Cutaneous Field Cancerization

 ,
,  ,
,  , , and
, , and 
Abstract
1. Introduction
2. Results
2.1. Expression of p53 and Ki67 Is Altered in CFC
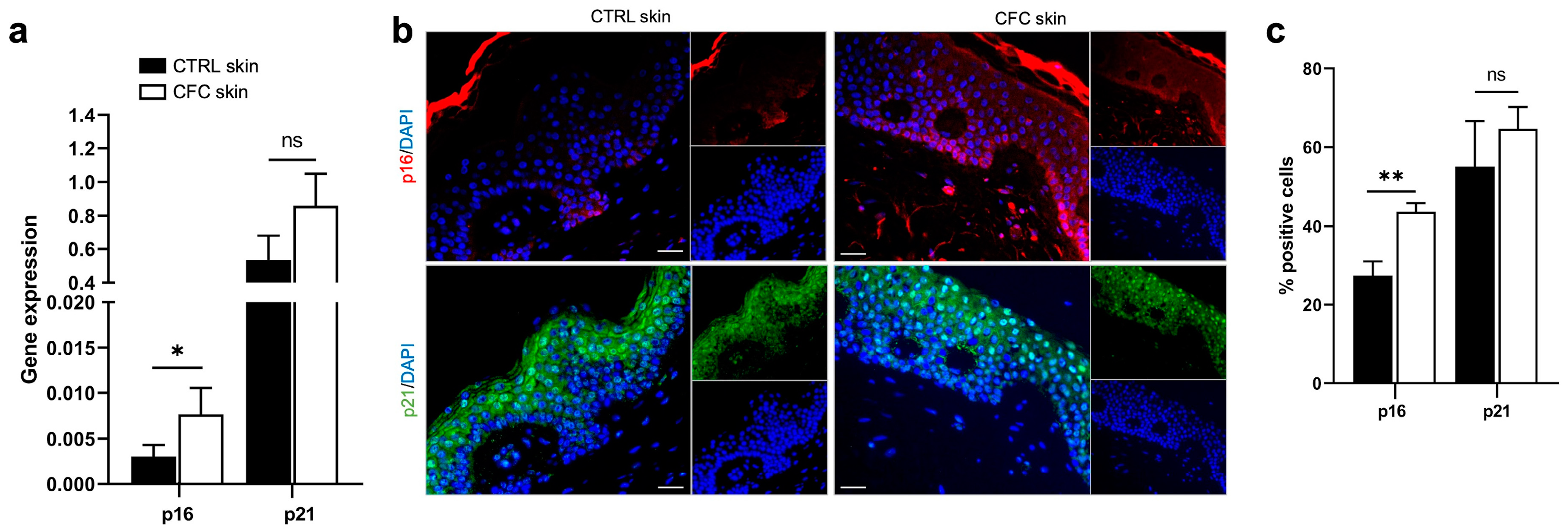
2.2. Cell Cycle Is Impaired in CFC
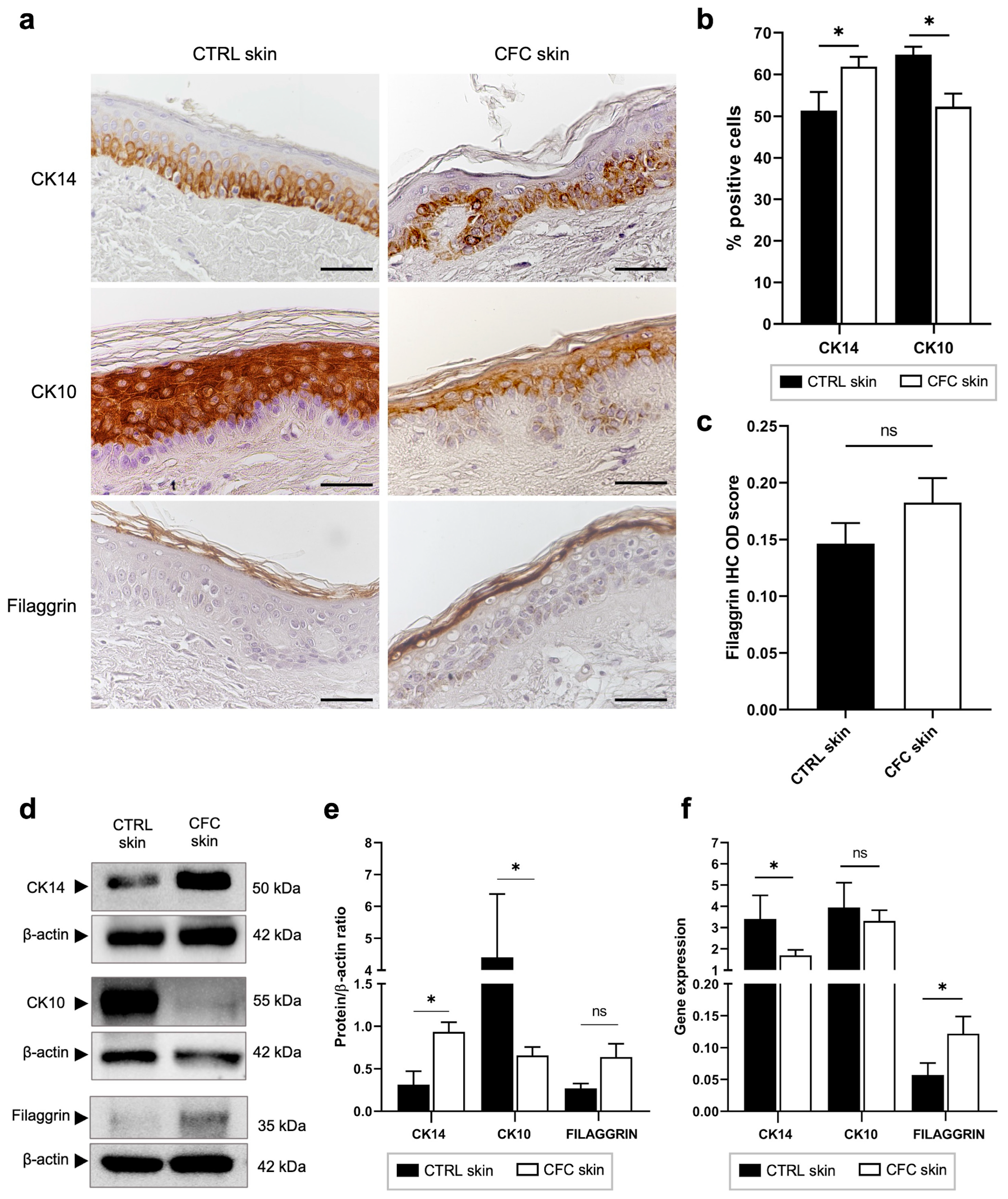
2.3. Analysis of Keratinocytes Differentiation Markers
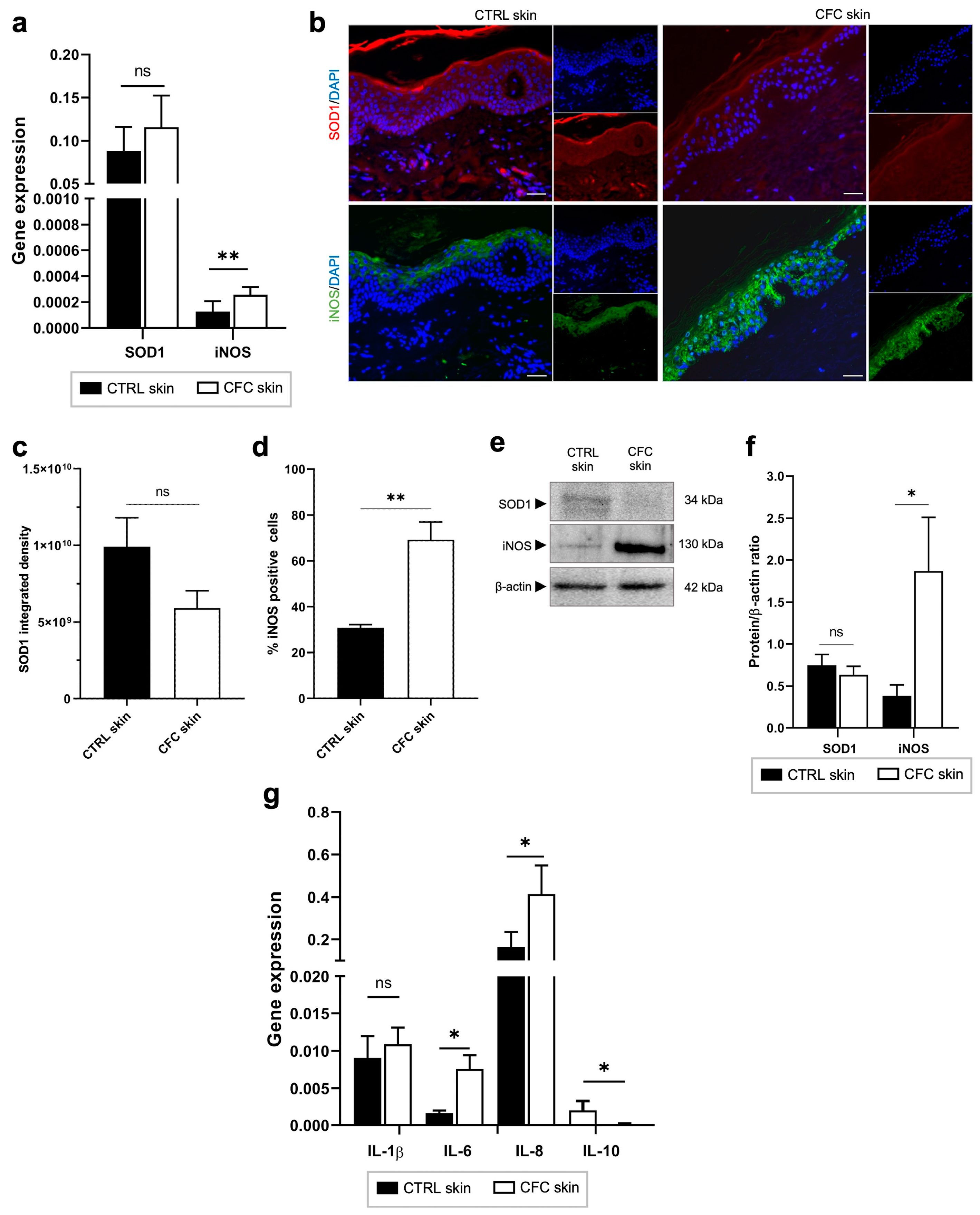
2.4. Oxidative Stress and Inflammation in CFC
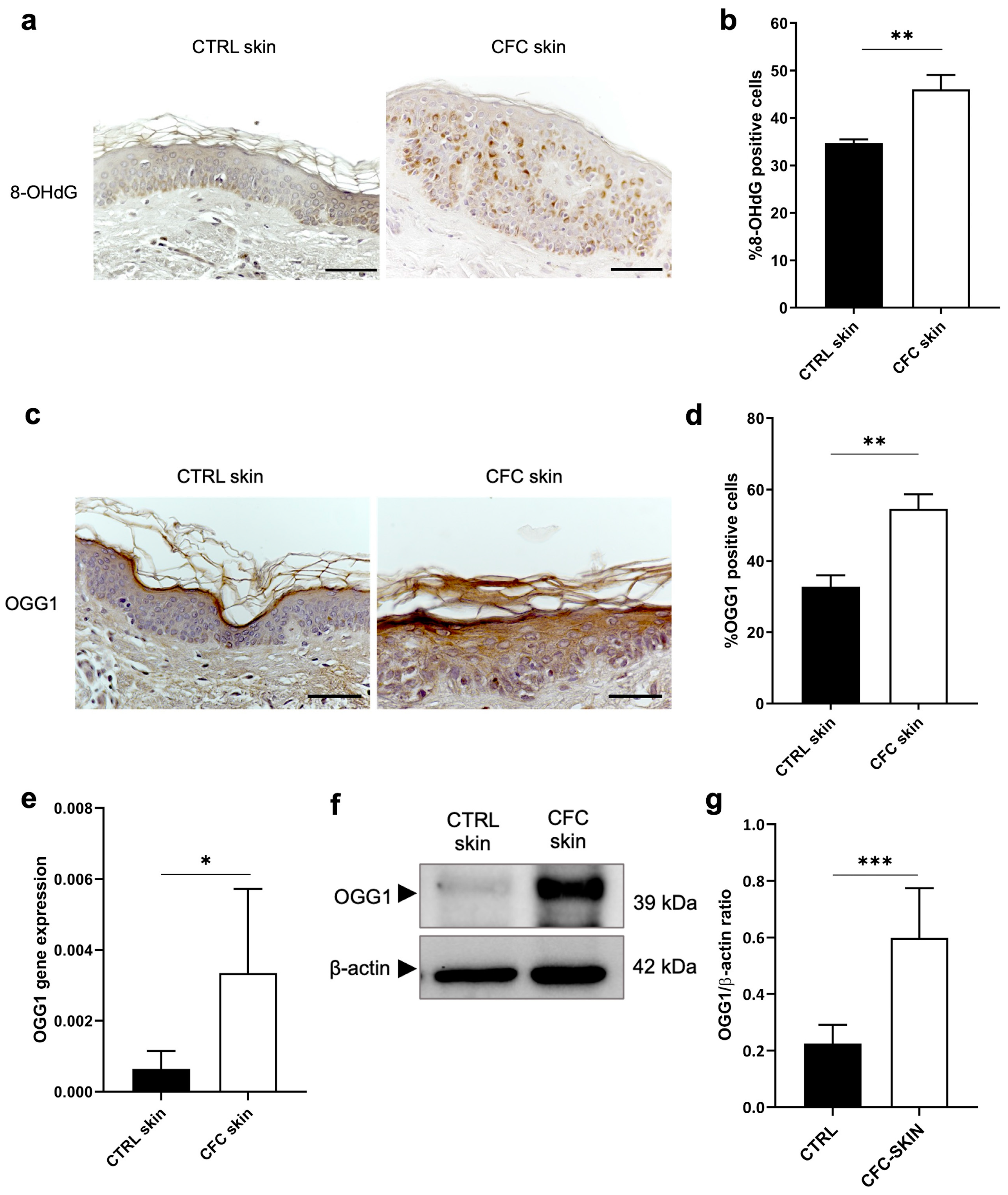
2.5. Analysis of Oxidative DNA Damage
3. Discussion
4. Materials and Methods
4.1. Patient Population and Skin Biopsies
4.2. Separation and Lysis of Epidermis
4.3. Immunohistochemistry (IHC)
4.4. Indirect Immunofluorescence (IF)
4.5. Quantitative Real-Time PCR
4.6. Western Blotting
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Curtius, K.; Wright, N.A.; Graham, T.A. An Evolutionary Perspective on Field Cancerization. Nat. Rev. Cancer 2018, 18, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Slaughter, D.P.; Southwick, H.W.; Smejkal, W. Field Cancerization in Oral Stratified Squamous Epithelium; Clinical Implications of Multicentric Origin. Cancer 1953, 6, 963–968. [Google Scholar] [CrossRef] [PubMed]
- Braakhuis, B.J.M.; Tabor, M.P.; Kummer, J.A.; Leemans, C.R.; Brakenhoff, R.H. A Genetic Explanation of Slaughter’s Concept of Field Cancerization: Evidence and Clinical Implications. Cancer Res. 2003, 63, 1727–1730. [Google Scholar] [PubMed]
- Willenbrink, T.J.; Ruiz, E.S.; Cornejo, C.M.; Schmults, C.D.; Arron, S.T.; Jambusaria-Pahlajani, A. Field Cancerization: Definition, Epidemiology, Risk Factors, and Outcomes. J. Am. Acad. Dermatol. 2020, 83, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Fernandez Figueras, M.T. From Actinic Keratosis to Squamous Cell Carcinoma: Pathophysiology Revisited. J. Eur. Acad. Dermatol. Venereol. 2017, 31 (Suppl. S2), 5–7. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.; Nguyen, J.K.; Austin, E.; Mamalis, A.; Jagdeo, J. Updates on Treatment Approaches for Cutaneous Field Cancerization. Curr. Dermatol. Rep. 2019, 8, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Morton, S.; Muir, J. Field Cancerization in the Skin: Past Errors Repeated. J. Am. Acad. Dermatol. 2021, 85, e41. [Google Scholar] [CrossRef] [PubMed]
- Willenbrink, T.J.; Ruiz, E.S.; Cornejo, C.M.; Schmults, C.D.; Arron, S.T.; Jambusaria-Pahlajani, A. Field Cancerization: Looking to the Future. J. Am. Acad. Dermatol. 2021, 85, e43–e44. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.B.; Edge, S.B.; Greene, F.L.; Byrd, D.R.; Brookland, R.K.; Washington, M.K.; Gershenwald, J.E.; Compton, C.C.; Hess, K.R.; Sullivan, D.C.; et al. AJCC Cancer Staging Manual; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; ISBN 978-3-319-40617-6. [Google Scholar]
- Torezan, L.A.R.; Festa-Neto, C. Cutaneous Field Cancerization: Clinical, Histopathological and Therapeutic Aspects. An. Bras. Dermatol. 2013, 88, 775–786. [Google Scholar] [CrossRef]
- Matsumura, Y.; Ananthaswamy, H.N. Toxic Effects of Ultraviolet Radiation on the Skin. Toxicol. Appl. Pharmacol. 2004, 195, 298–308. [Google Scholar] [CrossRef]
- Fisher, G.J.; Kang, S.; Varani, J.; Bata-Csorgo, Z.; Wan, Y.; Datta, S.; Voorhees, J.J. Mechanisms of Photoaging and Chronological Skin Aging. Arch. Dermatol. 2002, 138, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV Radiation and the Skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [PubMed]
- Klaunig, J.E. Oxidative Stress and Cancer. Curr. Pharm. Des. 2018, 24, 4771–4778. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-Hydroxy-2′-Deoxyguanosine (8-OHdG): A Critical Biomarker of Oxidative Stress and Carcinogenesis. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef]
- Kim, Y.; He, Y.-Y. Ultraviolet Radiation-Induced Non-Melanoma Skin Cancer: Regulation of DNA Damage Repair and Inflammation. Genes. Dis. 2014, 1, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Halliday, G.M. Inflammation, Gene Mutation and Photoimmunosuppression in Response to UVR-Induced Oxidative Damage Contributes to Photocarcinogenesis. Mutat. Res. 2005, 571, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Berman, B.; Cockerell, C.J. Pathobiology of Actinic Keratosis: Ultraviolet-Dependent Keratinocyte Proliferation. J. Am. Acad. Dermatol. 2013, 68, S10–S19. [Google Scholar] [CrossRef] [PubMed]
- Ling, G.; Persson, A.; Berne, B.; Uhlén, M.; Lundeberg, J.; Ponten, F. Persistent P53 Mutations in Single Cells from Normal Human Skin. Am. J. Pathol. 2001, 159, 1247–1253. [Google Scholar] [CrossRef]
- Jonason, A.S.; Kunala, S.; Price, G.J.; Restifo, R.J.; Spinelli, H.M.; Persing, J.A.; Leffell, D.J.; Tarone, R.E.; Brash, D.E. Frequent Clones of P53-Mutated Keratinocytes in Normal Human Skin. Proc. Natl. Acad. Sci. USA 1996, 93, 14025–14029. [Google Scholar] [CrossRef]
- Martincorena, I.; Roshan, A.; Gerstung, M.; Ellis, P.; Van Loo, P.; McLaren, S.; Wedge, D.C.; Fullam, A.; Alexandrov, L.B.; Tubio, J.M.; et al. Tumor Evolution. High Burden and Pervasive Positive Selection of Somatic Mutations in Normal Human Skin. Science 2015, 348, 880–886. [Google Scholar] [CrossRef]
- South, A.P.; Purdie, K.J.; Watt, S.A.; Haldenby, S.; den Breems, N.; Dimon, M.; Arron, S.T.; Kluk, M.J.; Aster, J.C.; McHugh, A.; et al. NOTCH1 Mutations Occur Early during Cutaneous Squamous Cell Carcinogenesis. J. Investig. Dermatol. 2014, 134, 2630–2638. [Google Scholar] [CrossRef] [PubMed]
- Kanellou, P.; Zaravinos, A.; Zioga, M.; Stratigos, A.; Baritaki, S.; Soufla, G.; Zoras, O.; Spandidos, D.A. Genomic Instability, Mutations and Expression Analysis of the Tumour Suppressor Genes P14(ARF), P15(INK4b), P16(INK4a) and P53 in Actinic Keratosis. Cancer Lett. 2008, 264, 145–161. [Google Scholar] [CrossRef] [PubMed]
- Chitsazzadeh, V.; Coarfa, C.; Drummond, J.A.; Nguyen, T.; Joseph, A.; Chilukuri, S.; Charpiot, E.; Adelmann, C.H.; Ching, G.; Nguyen, T.N.; et al. Cross-Species Identification of Genomic Drivers of Squamous Cell Carcinoma Development across Preneoplastic Intermediates. Nat. Commun. 2016, 7, 12601. [Google Scholar] [CrossRef] [PubMed]
- Miola, A.C.; Castilho, M.A.; Schmitt, J.V.; Marques, M.E.A.; Miot, H.A. Contribution to Characterization of Skin Field Cancerization Activity: Morphometric, Chromatin Texture, Proliferation, and Apoptosis Aspects. An. Bras. Dermatol. 2019, 94, 698–703. [Google Scholar] [CrossRef] [PubMed]
- Warino, L.; Tusa, M.; Camacho, F.; Teuschler, H.; Fleischer, A.B.; Feldman, S.R. Frequency and Cost of Actinic Keratosis Treatment. Dermatol. Surg. 2006, 32, 1045–1049. [Google Scholar] [CrossRef] [PubMed]
- Cornejo, C.M.; Jambusaria-Pahlajani, A.; Willenbrink, T.J.; Schmults, C.D.; Arron, S.T.; Ruiz, E.S. Field Cancerization: Treatment. J. Am. Acad. Dermatol. 2020, 83, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Hodges, A.; Smoller, B.R. Immunohistochemical Comparison of P16 Expression in Actinic Keratoses and Squamous Cell Carcinomas of the Skin. Mod. Pathol. 2002, 15, 1121–1125. [Google Scholar] [CrossRef] [PubMed]
- Mohan, M.; Jagannathan, N. Oral Field Cancerization: An Update on Current Concepts. Oncol. Rev. 2014, 8, 244. [Google Scholar] [CrossRef] [PubMed]
- Peralta-Mamani, M.; Terrero-Pérez, Á.; Tucunduva, R.M.A.; Rubira, C.M.F.; Santos, P.S.d.S.; Honório, H.M.; Rubira-Bullen, I.R.F. Occurrence of Field Cancerization in Clinically Normal Oral Mucosa: A Systematic Review and Meta-Analysis. Arch. Oral. Biol. 2022, 143, 105544. [Google Scholar] [CrossRef]
- Sherr, C.J.; McCormick, F. The RB and P53 Pathways in Cancer. Cancer Cell 2002, 2, 103–112. [Google Scholar] [CrossRef]
- Moll, R.; Divo, M.; Langbein, L. The Human Keratins: Biology and Pathology. Histochem. Cell Biol. 2008, 129, 705–733. [Google Scholar] [CrossRef]
- Kezic, S.; Jakasa, I. Filaggrin and Skin Barrier Function. Curr. Probl. Dermatol. 2016, 49, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Montes de Oca, M.K.; Pearlman, R.L.; McClees, S.F.; Strickland, R.; Afaq, F. Phytochemicals for the Prevention of Photocarcinogenesis. Photochem. Photobiol. 2017, 93, 956–974. [Google Scholar] [CrossRef] [PubMed]
- Boiteux, S.; Radicella, J.P. The Human OGG1 Gene: Structure, Functions, and Its Implication in the Process of Carcinogenesis. Arch. Biochem. Biophys. 2000, 377, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nishisgori, C. Current Concept of Photocarcinogenesis. Photochem. Photobiol. Sci. 2015, 14, 1713–1721. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S.R. Recent Advances in Field Cancerization and Management of Multiple Cutaneous Squamous Cell Carcinomas. F1000Res 2018, 7, F1000 Faculty Rev-690. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Liang, L.; Matsumoto, M.; Iwata, K.; Umemura, A.; He, F. Reactive Oxygen Species and NRF2 Signaling, Friends or Foes in Cancer? Biomolecules 2023, 13, 353. [Google Scholar] [CrossRef] [PubMed]
- Panieri, E.; Telkoparan-Akillilar, P.; Saso, L. NRF2, a Crucial Modulator of Skin Cells Protection against Vitiligo, Psoriasis, and Cancer. BioFactors 2023, 49, 228–250. [Google Scholar] [CrossRef] [PubMed]
- Zinatizadeh, M.R.; Schock, B.; Chalbatani, G.M.; Zarandi, P.K.; Jalali, S.A.; Miri, S.R. The Nuclear Factor Kappa B (NF-kB) Signaling in Cancer Development and Immune Diseases. Genes. Dis. 2020, 8, 287–297. [Google Scholar] [CrossRef]
- Bang, E.; Kim, D.H.; Chung, H.Y. Protease-Activated Receptor 2 Induces ROS-Mediated Inflammation through Akt-Mediated NF-κB and FoxO6 Modulation during Skin Photoaging. Redox Biol. 2021, 44, 102022. [Google Scholar] [CrossRef]
- Conforti, C.; Giuffrida, R.; Pizzichetta, M.A.; Di Meo, N.; Magaton-Rizzi, G.; Zalaudek, I. Integrating the Concept of Field Cancerization in the Classification and Risk Assessment of Cutaneous Squamous Cell Carcinoma: Proposal for a New Classification and Terminology of Keratinocyte Skin Cancer. J. Eur. Acad. Dermatol. Venereol. 2019, 33, e327–e330. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, T.; Nakagawara, A. P53: The Attractive Tumor Suppressor in the Cancer Research Field. J. Biomed. Biotechnol. 2011, 2011, 603925. [Google Scholar] [CrossRef]
- Berg, R.J.; van Kranen, H.J.; Rebel, H.G.; de Vries, A.; van Vloten, W.A.; Van Kreijl, C.F.; van der Leun, J.C.; de Gruijl, F.R. Early P53 Alterations in Mouse Skin Carcinogenesis by UVB Radiation: Immunohistochemical Detection of Mutant P53 Protein in Clusters of Preneoplastic Epidermal Cells. Proc. Natl. Acad. Sci. USA 1996, 93, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Kramata, P.; Lu, Y.-P.; Lou, Y.-R.; Singh, R.N.; Kwon, S.M.; Conney, A.H. Patches of Mutant P53-Immunoreactive Epidermal Cells Induced by Chronic UVB Irradiation Harbor the Same P53 Mutations as Squamous Cell Carcinomas in the Skin of Hairless SKH-1 Mice. Cancer Res. 2005, 65, 3577–3585. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Ramani, P. Histologic and Immunohistochemical Evaluation of Mirror Image Biopsies in Oral Squamous Cell Carcinoma. J. Oral. Biol. Craniofac Res. 2016, 6, 194–197. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.J.; Synnott, N.C.; O’Grady, S.; Crown, J. Targeting P53 for the Treatment of Cancer. Semin. Cancer Biol. 2022, 79, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Marinescu, A.; Mărgăritescu, C.; Stepan, A.E.; Marinescu, A.M.; Simionescu, C.E.; Nitu, F.M. P21 Immunoexpression in Actinic Keratosis and Cutaneous Carcinomas. Curr. Health Sci. J. 2017, 43, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Pavey, S.; Russell, T.; Gabrielli, B. G2 Phase Cell Cycle Arrest in Human Skin Following UV Irradiation. Oncogene 2001, 20, 6103–6110. [Google Scholar] [CrossRef] [PubMed]
- Birajdar, S.S.; Radhika, M.; Paremala, K.; Sudhakara, M.; Soumya, M.; Gadivan, M. Expression of Ki-67 in Normal Oral Epithelium, Leukoplakic Oral Epithelium and Oral Squamous Cell Carcinoma. J. Oral. Maxillofac. Pathol. 2014, 18, 169–176. [Google Scholar] [CrossRef]
- Montebugnoli, L.; Gissi, D.B.; Badiali, G.; Marchetti, C.; Cervellati, F.; Farnedi, A.; Foschini, M.P. Ki-67 from Clinically and Histologically “Normal” Distant Mucosa as Prognostic Marker in Early-Stage (T1-T2N0) Oral Squamous Cell Carcinoma: A Prospective Study. J. Oral Maxillofac. Surg. 2011, 69, 2579–2584. [Google Scholar] [CrossRef]
- Choi, K.H.; Kim, G.M.; Kim, S.Y. The Keratin-14 Expression in Actinic Keratosis and Squamous Cell Carcinoma: Is This a Prognostic Factor for Tumor Progression? Cancer Res. Treat. 2010, 42, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, A.; Liu, X.; Wang, Q.; Bai, Y.; Liu, Z.; Huang, L.; Wu, M.; Li, H.; Miao, J.; et al. A Panel of Biomarkers for Skin Squamous Cell Carcinoma: Various Functional Entities and Differential Responses to Resveratrol. Int. J. Clin. Exp. Pathol. 2019, 12, 1363–1377. [Google Scholar] [PubMed]
- Dotto, G.P. Multifocal Epithelial Tumors and Field Cancerization: Stroma as a Primary Determinant. J. Clin. Investig. 2014, 124, 1446–1453. [Google Scholar] [CrossRef] [PubMed]
- Yoshifuku, A.; Fujii, K.; Kanekura, T. Comparison of Oxidative Stress on DNA, Protein and Lipids in Patients with Actinic Keratosis, Bowen’s Disease and Squamous Cell Carcinoma. J. Dermatol. 2018, 45, 1319–1323. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | Sex | Anatomical Site | |
|---|---|---|---|
| Ctrl_1 | 51 | M | Trunk |
| Ctrl_2 | 41 | M | Trunk |
| Ctrl_3 | 79 | M | Trunk |
| Ctrl_4 | 67 | M | Trunk |
| Ctrl_5 | 88 | M | Trunk |
| Ctrl_6 | 86 | M | Lower limb |
| Ctrl_7 | 84 | F | Scalp |
| Ctrl_8 | 61 | F | Head |
| Ctrl_9 | 39 | F | Scalp |
| Ctrl_10 | 79 | M | Face |
| Ctrl_11 | 62 | M | Lower limb |
| Ctrl_12 | 47 | F | Scalp |
| Ctrl_13 | 51 | M | Scalp |
| Ctrl_14 | 76 | F | Neck |
| Ctrl_15 | 75 | M | Scalp |
| Ctrl_16 | 88 | F | Trunk |
| Ctrl_17 | 90 | M | Trunk |
| Ctrl_18 | 67 | M | Upper limb |
| Ctrl_19 | 71 | F | Scalp |
| Ctrl_20 | 65 | M | Face |
| Ctrl_21 | 79 | F | Scalp |
| Ctrl_22 | 85 | F | Neck |
| Ctrl_23 | 89 | M | Trunk |
| Ctrl_24 | 86 | F | Scalp |
| Ctrl_25 | 75 | M | Lower limb |
| Patient | Age | Sex | Anatomical Site | Skin Cancer |
|---|---|---|---|---|
| CFC_1 | 83 | M | Head | BCC |
| CFC_2 | 83 | M | Trunk | BCC |
| CFC_3 | 82 | M | Face | BCC |
| CFC_4 | 87 | M | Face | BCC |
| CFC_5 | 91 | M | Head | Keratoacanthomas |
| CFC_6 | 78 | F | Face | BCC |
| CFC_7 | 84 | M | Trunk | BCC |
| CFC_8 | 80 | M | Trunk | BCC |
| CFC_9 | 83 | M | Trunk | BCC |
| CFC_10 | 75 | M | Head | SCC |
| CFC_11 | 82 | M | Face | BCC |
| CFC_12 | 87 | M | Neck | BCC |
| CFC_13 | 91 | M | Head | Keratoacanthomas |
| CFC_14 | 78 | F | Face | BCC |
| CFC_15 | 87 | F | Face | BCC |
| CFC_16 | 67 | M | Lower limb | BCC |
| CFC_17 | 86 | M | Lower limb | Melanoma |
| CFC_18 | 87 | M | Face | SCC |
| CFC_19 | 90 | F | Face | BCC |
| CFC_20 | 72 | M | Trunk | AK |
| CFC_21 | 83 | M | Trunk | Epithelioma |
| CFC_22 | 85 | M | Upper limb | BCC |
| CFC_23 | 83 | M | Head | BCC |
| CFC_24 | 86 | M | Scalp | KA |
| CFC_25 | 83 | M | Head | Epithelioma |
| CFC_26 | 75 | M | Face | BCC |
| CFC_27 | 77 | F | Trunk | BCC |
| CFC_28 | 51 | M | Trunk | BCC |
| CFC_29 | 74 | M | Trunk | Melanoacanthoma |
| CFC_30 | 63 | M | Trunk | BCC |
| CFC_31 | 89 | M | Lower limb | SCC |
| CFC_32 | 86 | M | Head | AK |
| CFC_33 | 67 | M | Trunk | BCC |
| CFC_34 | 54 | M | Trunk | Displastic nevi |
| CFC_35 | 78 | F | Trunk | BCC |
| CFC_36 | 82 | M | Upper limb | BCC |
| CFC_37 | 66 | F | Trunk | BCC |
| CFC_38 | 64 | F | Face | BCC |
| CFC_39 | 83 | M | Head | BCC |
| CFC_40 | 83 | M | Head | BCC |
| CFC_41 | 74 | M | Lower limb | Melanoma |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camillo, L.; Zavattaro, E.; Veronese, F.; Gironi, L.C.; Cremona, O.; Savoia, P. Ex Vivo Analysis of Cell Differentiation, Oxidative Stress, Inflammation, and DNA Damage on Cutaneous Field Cancerization. Int. J. Mol. Sci. 2024, 25, 5775. https://doi.org/10.3390/ijms25115775
Camillo L, Zavattaro E, Veronese F, Gironi LC, Cremona O, Savoia P. Ex Vivo Analysis of Cell Differentiation, Oxidative Stress, Inflammation, and DNA Damage on Cutaneous Field Cancerization. International Journal of Molecular Sciences. 2024; 25(11):5775. https://doi.org/10.3390/ijms25115775
Chicago/Turabian StyleCamillo, Lara, Elisa Zavattaro, Federica Veronese, Laura Cristina Gironi, Ottavio Cremona, and Paola Savoia. 2024. "Ex Vivo Analysis of Cell Differentiation, Oxidative Stress, Inflammation, and DNA Damage on Cutaneous Field Cancerization" International Journal of Molecular Sciences 25, no. 11: 5775. https://doi.org/10.3390/ijms25115775
APA StyleCamillo, L., Zavattaro, E., Veronese, F., Gironi, L. C., Cremona, O., & Savoia, P. (2024). Ex Vivo Analysis of Cell Differentiation, Oxidative Stress, Inflammation, and DNA Damage on Cutaneous Field Cancerization. International Journal of Molecular Sciences, 25(11), 5775. https://doi.org/10.3390/ijms25115775