
Glial Markers of Suicidal Behavior in the Human Brain—A Systematic Review of Postmortem Studies

Abstract
1. Introduction
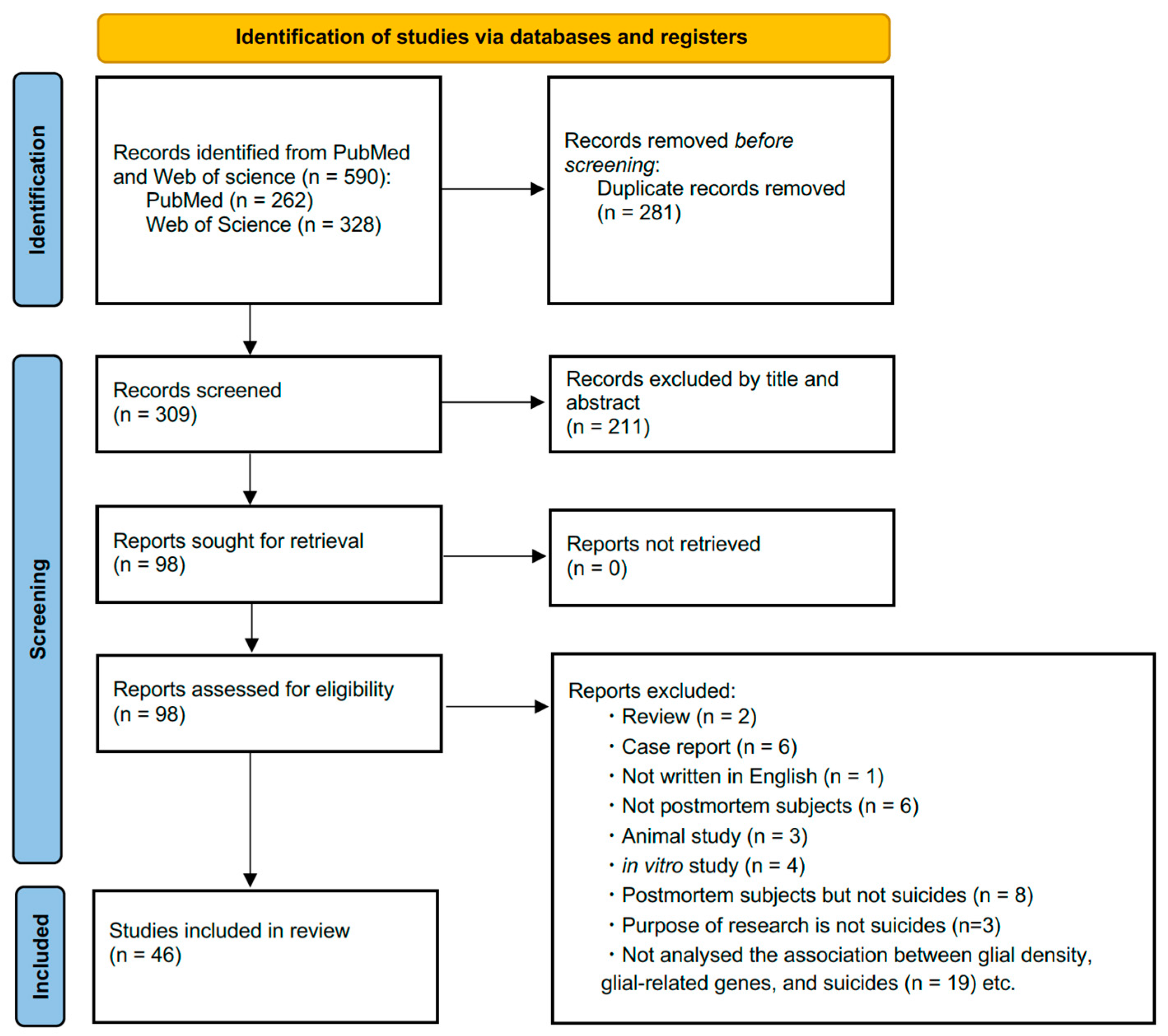
2. Materials and Methods
2.1. Literature Search Strategy
2.2. Eligibility Criteria
2.3. Screening
2.4. Article Selection
2.5. Data Extraction
3. Results
3.1. Glial-Related Alterations in Postmortem Suicidal Brain
3.1.1. Glial Cell Populations

| No. | Cell Type | Region | Changes | Diagnoses | Comparison |
|---|---|---|---|---|---|
| 1 | Glia (Nissl) | OFC | → [34] | MDD (Ad) | Ad suicides vs. Ad non-suicides |
| 2 | Glia (Nissl) | ACC | → vs. non-suicides or ↑ vs. non-Ad suicides [35] | MDD (Ad) | Ad suicides vs. non-Ad suicides Ad suicides vs. non-suicides |
| 3 | Glia (Nissl) | ACC and DLPFC | → [37] | MDD (ELA) and SCZ (ELA) | ELA suicides vs. ELA non-suicides |
| 4 | Glia (Nissl) | AAC and PAC | → [36] | MDD, SCZ | MDD and SCZ suicides vs. H |
| 5 | Glia (Nissl) | CA and DG | → [38] | MDD (ELA) | MDD (ELA) suicides vs. ELA non-suicides |
| 6 | Glia (Nissl) | CA and DG | → [39] | MDD, SCZ | MDD and SCZ suicides vs. H |
| 7 | Glia (Nissl) | anterior DG | ↓ [41] | MDD (As) | MDD (unmedicated) suicides vs. H |
| 8 | Astrocyte (Golgi) | ACC | ↑ [42] | MDD | MDD suicides vs. H |
| 9 | Astrocyte (GFAP) | CA | → [43] | MDD | MDD suicides vs. H |
| 10 | Astrocyte (GFAP and Vimentin) | DCN, DLPFC, and MD | ↓ [44] | MDD | MDD suicides vs. H |
| 11 | Astrocyte (GFAP), synemin-α, synemin-β, Vimentin, nestin) | ACC and DLPFC | ↑↓ [48] | BD and MDD | BD and MDD suicides vs. H |
| 12 | Astrocyte (GFAP) | LC | ↓ [45] | MDD | MDD suicides vs. H |
| 13 | Astrocyte (GFAP, TrkB.T1, Trkb.T1) | DLPFC | → [46] | BD and MDD | MDD suicides vs. H |
| 14 | Microglia (HLA-DR) | ACC, DLPFC, and MD | ↑ [50] $ | AD, SCZ | AD and SCZ suicides vs. AD and SCZ non-suicides |
| 15 | Microglia (HLA-DR) | ACC and MD | ↑ [49] # | SCZ | SCZ suicides vs. SCZ non-suicides |
| 16 | Microglia (HLA-DR) | DRN | → [55] | BD, MDD, and SCZ | BD, MDD, and SCZ suicides vs. BD, MDD, and SCZ non-suicides vs. H |
| 17 | Microglia (MHC II, P2RY12) | HIPP | ↑ [54] | BD | BD suicides vs. BD non-suicides |
| 18 | Microglia (IBA1) | AmCC | ↑ [51] | BD, SCZ | BD and SCZ suicides vs. H |
| 19 | Microglia (IBA1) | dACC | → [56] | UD | UD suicides vs. H |
| 20 | Microglia (IBA1) | DLPFC and VLPFC | → [57] | AD, SCZ | AD, SZ, and H suicides vs. AD, SZ, and H non-suicides |
| 21 | Microglia (QUIN) | sACC and AmCC | ↑ [52] | BD, MDD | BD and MDD suicides vs. H |
| 22 | Microglia (QUIN) | right CA1 | ↓ [53] | BD, UD | BD and UD suicides vs. H |
| 23 | Oligodendrocyte (OLIG2, PDGFRA, APC, and NogoA) | VMPFC | ↑↓ [59] | MDD (ELA) | MDD (ELA) suicides vs. MDD suicides MDD (ELA) suicides vs. H |
| 24 | Oligodendrocyte (MBP) | aPFC | ↓ [58] | MDD and SCZ | MDD, SCZ suicides vs. H |
| 25 | Oligodendrocyte population | TC | ↓ [61] | MDD | MDD suicides vs. H |
| 26 | Oligodendrocyte (telomere lengths) | OCC and LC | ↓ [60] | MDD | MDD suicides vs. H |
| 27 | Methylated regions proportions | OFC and vACC | Changed [62] | MDD | MDD suicides vs. H |
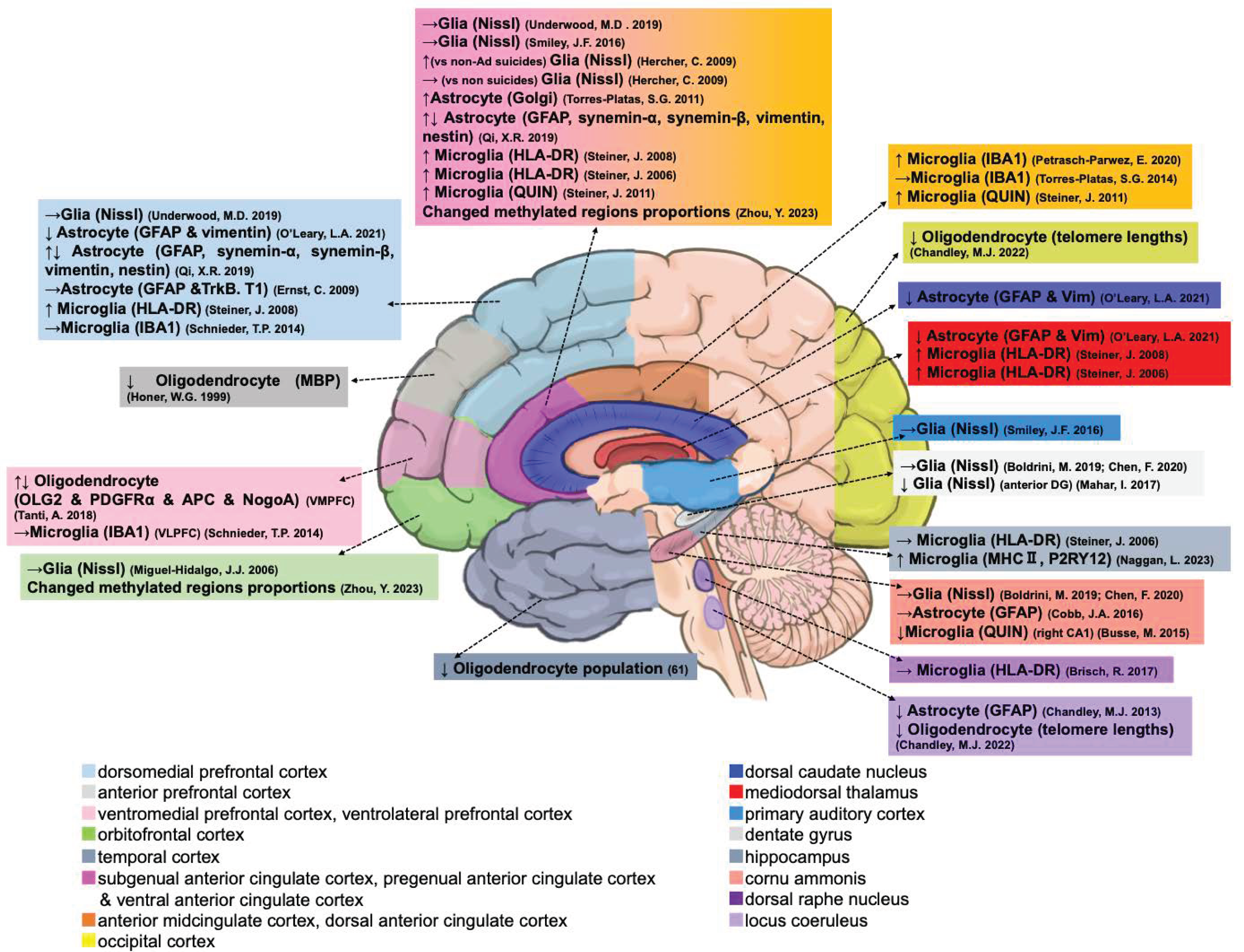
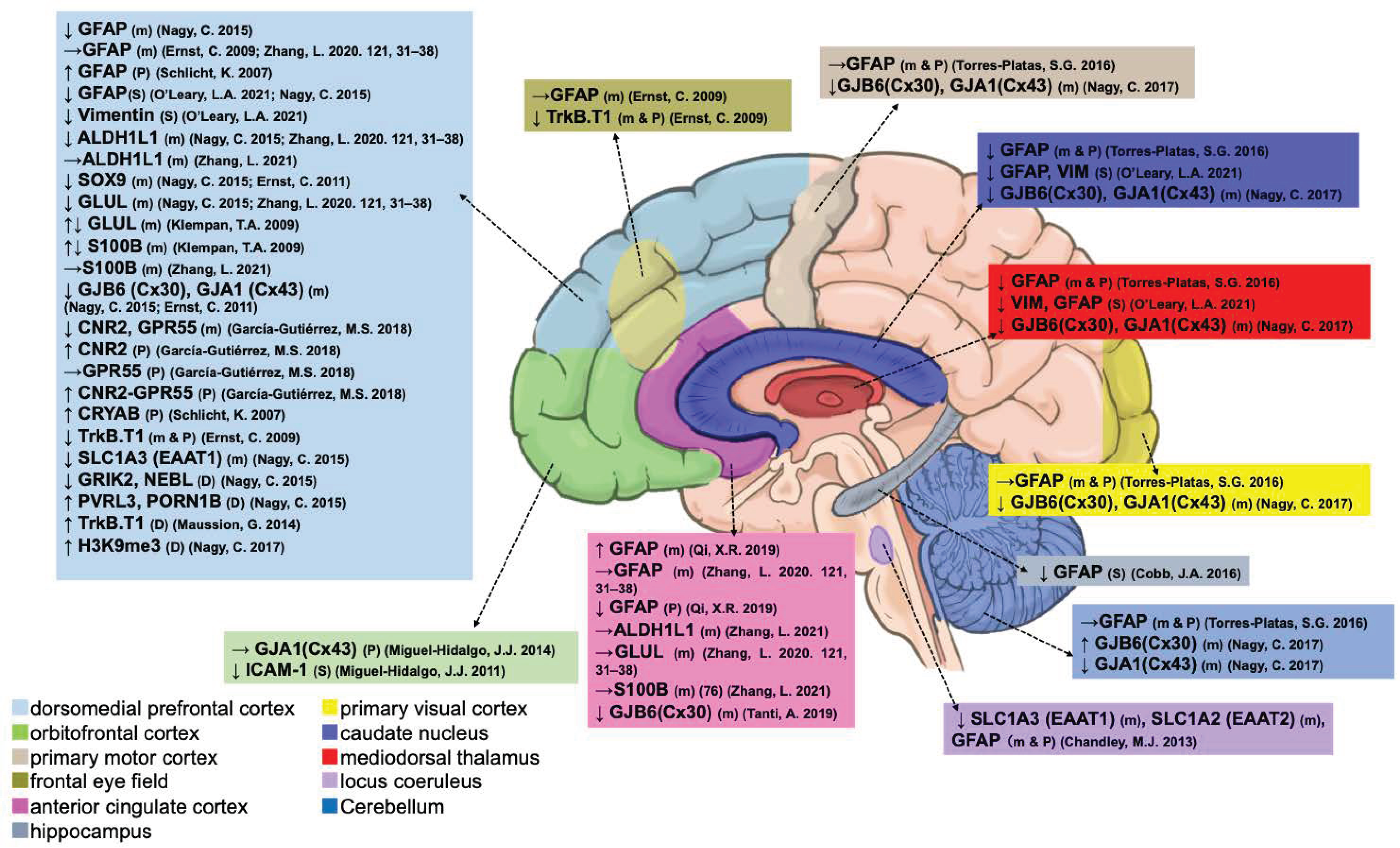
3.1.2. Astrocytes
| Astrocyte | Type | ACC | FEF | DLPFC | OFC | MD | PMC | PVC | CN | HIPP | LC | Cb |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Marker | ||||||||||||
| GFAP | mRNA | ↑ [48] → [65] | → [46] | ↓ [63] → [46,65] | — | ↓ [64] | → [64] | → [64] | ↓ [64] | — | ↓ [45] | → [64] |
| GFAP | Protein | ↓ [48] | — | ↑ [66] | — | ↓ [64] | → [64] | → [64] | ↓ [64] | — | ↓ [45] | → [64] |
| Stain | — | — | ↓ [44,63] | — | ↓ [44] | — | — | ↓ [44] | ↓ [43] | — | — | |
| Vimentin | Stain | — | — | ↓ [44] | — | ↓ [44] | — | — | ↓ [44] | — | — | — |
| ALDH1L1 | mRNA | → [76] | — | ↓ [63,65] → [76] | — | — | — | — | — | — | — | — |
| SOX9 | mRNA | — | — | ↓ [63,67] | — | — | — | — | — | — | — | — |
| GLUL | mRNA | → [65] | — | ↓ [63,65] ↑↓ [68] | — | — | — | — | — | — | — | — |
| S100B | mRNA | → [76] | — | ↑↓ [68] → [76] | — | — | — | — | — | — | — | — |
| GJA1 (CX43) | mRNA | — | — | ↓ [63,67] | — | ↓ [72] | ↓ [72] | ↓ [72] | ↓ [72] | — | — | ↓ [72] |
| GJA1 (CX43) | Protein | — | — | — | → [74] | — | — | — | — | — | — | — |
| GJB6 (CX30) | mRNA | ↓ [71] | — | ↓ [63,67] | — | ↓ [72] | ↓ [72] | ↓ [72] | ↓ [72] | — | — | ↑ [72] |
| ICAM1 | Stain | — | — | — | ↓ [73] | — | — | — | — | — | — | — |
| CNR2 | mRNA | — | — | ↓ [75] | — | — | — | — | — | — | — | — |
| CNR2 | Protein | — | — | ↑ [75] | — | — | — | — | — | — | — | — |
| GPR55 | mRNA | — | — | ↓ [75] | — | — | — | — | — | — | — | — |
| GPR55 | Protein | — | — | → [75] | — | — | — | — | — | — | — | — |
| CNR2-GPR55 | Protein | — | — | ↑ [75] | — | — | — | — | — | — | — | — |
| CRYAB | Protein | — | — | ↑ [66] | — | — | — | — | — | — | — | — |
| TrkB.T1 | mRNA | — | ↓ [46] | ↓ [46] | — | — | — | — | — | — | — | — |
| TrkB.T1 | Protein | — | ↓ [46] | ↓ [46] | — | — | — | — | — | — | — | — |
| Glutamate–glutamine cycle | ||||||||||||
| SLC1A3 (EAAT1) | mRNA | — | — | ↓ [63] | — | — | — | — | — | — | ↓ [45] | — |
| SLC1A2 (EAAT2) | mRNA | — | — | — | — | — | — | — | — | — | ↓ [45] | — |
| Methylation | ||||||||||||
| GRIK2 | DNA | — | — | ↓ [63] | — | — | — | — | — | — | — | — |
| NEBL | DNA | — | — | ↓ [63] | — | — | — | — | — | — | — | — |
| PVRL3 | DNA | — | — | ↑ [63] | — | — | — | — | — | — | — | — |
| ROPN1B | DNA | — | — | ↑ [63] | — | — | — | — | — | — | — | — |
| TrkB.T1 | DNA | — | — | ↑ [47] | — | — | — | — | — | — | — | — |
| Epigenetic silencing | ||||||||||||
| H3K9me3 | DNA | — | — | ↑ [72] | — | — | — | — | — | — | — | — |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3.1.3. Microglia
3.1.4. Oligodendrocytes
3.2. Alterations of Glial Cell Populations with Major Psychiatric Disorders
4. Discussion
Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Claveria, O. Global economic uncertainty and suicide: Worldwide evidence. Soc. Sci. Med. 2022, 305, 115041. [Google Scholar] [CrossRef]
- Bachmann, S. Epidemiology of Suicide and the Psychiatric Perspective. Int. J. Environ. Res. Public Health 2018, 15, 1425. [Google Scholar] [CrossRef] [PubMed]
- Sechter, D.; Bonin, B.; Bertschy, G.; Vandel, S.; Bizouard, P. Prediction of suicide risk. Encephale 1991, 17, 361–364. [Google Scholar] [PubMed]
- Segal, A.; Parkes, L.; Aquino, K.; Kia, S.M.; Wolfers, T.; Franke, B.; Hoogman, M.; Beckmann, C.F.; Westlye, L.T.; Andreassen, O.A.; et al. Regional, circuit and network heterogeneity of brain abnormalities in psychiatric disorders. Nat. Neurosci. 2023, 26, 1613–1629. [Google Scholar] [CrossRef]
- Turecki, G.; Brent, D.A. Suicide and suicidal behaviour. Lancet 2016, 387, 1227–1239. [Google Scholar] [CrossRef]
- Parpura, V.; Heneka, M.T.; Montana, V.; Oliet, S.H.; Schousboe, A.; Haydon, P.G.; Stout, R.F., Jr.; Spray, D.C.; Reichenbach, A.; Pannicke, T.; et al. Glial cells in (patho)physiology. J. Neurochem. 2012, 121, 4–27. [Google Scholar] [CrossRef]
- Allen, N.J.; Lyons, D.A. Glia as architects of central nervous system formation and function. Science 2018, 362, 181–185. [Google Scholar] [CrossRef]
- Piras, I.S.; Huentelman, M.J.; Pinna, F.; Paribello, P.; Solmi, M.; Murru, A.; Carpiniello, B.; Manchia, M.; Zai, C.C. A review and meta-analysis of gene expression profiles in suicide. Eur. Neuropsychopharmacol. 2022, 56, 39–49. [Google Scholar] [CrossRef]
- Frick, L.R.; Williams, K.; Pittenger, C. Microglial dysregulation in psychiatric disease. Clin. Dev. Immunol. 2013, 2013, 608654. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, A.; De Sousa, A.; Rao, G.P. Brain-Derived Neurotrophic Factor and Suicide in Schizophrenia: Critical Role of Neuroprotective Mechanisms as an Emerging Hypothesis. Indian J. Psychol. Med. 2016, 38, 499–504. [Google Scholar] [CrossRef]
- Deveci, A.; Aydemir, O.; Taskin, O.; Taneli, F.; Esen-Danaci, A. Serum BDNF levels in suicide attempters related to psychosocial stressors: A comparative study with depression. Neuropsychobiology 2007, 56, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Corkrum, M.; Covelo, A.; Lines, J.; Bellocchio, L.; Pisansky, M.; Loke, K.; Quintana, R.; Rothwell, P.E.; Lujan, R.; Marsicano, G.; et al. Dopamine-Evoked Synaptic Regulation in the Nucleus Accumbens Requires Astrocyte Activity. Neuron 2020, 105, 1036–1047.e5. [Google Scholar] [CrossRef] [PubMed]
- Irizarry-Valle, Y.; Parker, A.C. An astrocyte neuromorphic circuit that influences neuronal phase synchrony. IEEE Trans. Biomed. Circuits Syst. 2015, 9, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Kol, A.; Adamsky, A.; Groysman, M.; Kreisel, T.; London, M.; Goshen, I. Astrocytes contribute to remote memory formation by modulating hippocampal-cortical communication during learning. Nat. Neurosci. 2020, 23, 1229–1239. [Google Scholar] [CrossRef] [PubMed]
- Abbott, N.J.; Rönnbäck, L.; Hansson, E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Ono, C.; Kim, H.B.; Komatsu, H.; Tanabe, Y.; Sakae, N.; Nakayama, K.I.; Matsuoka, H.; Sora, I.; Bunney, W.E.; et al. Four mood stabilizers commonly induce FEZ1 expression in human astrocytes. Bipolar Disord. 2011, 13, 486–499. [Google Scholar] [CrossRef]
- Durkee, C.A.; Araque, A. Diversity and Specificity of Astrocyte-neuron Communication. Neuroscience 2019, 396, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Banati, R.B. Neuropathological imaging: In vivo detection of glial activation as a measure of disease and adaptive change in the brain. Br. Med. Bull. 2003, 65, 121–131. [Google Scholar] [CrossRef]
- Mehl, L.C.; Manjally, A.V.; Bouadi, O.; Gibson, E.M.; Tay, T.L. Microglia in brain development and regeneration. Development 2022, 149, dev200425. [Google Scholar] [CrossRef]
- Borst, K.; Dumas, A.A.; Prinz, M. Microglia: Immune and non-immune functions. Immunity 2021, 54, 2194–2208. [Google Scholar] [CrossRef]
- Sakai, M.; Takeuchi, H.; Yu, Z.; Kikuchi, Y.; Ono, C.; Takahashi, Y.; Ito, F.; Matsuoka, H.; Tanabe, O.; Yasuda, J.; et al. Polymorphisms in the microglial marker molecule CX3CR1 affect the blood volume of the human brain. Psychiatry Clin. Neurosci. 2018, 72, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Sakai, M.; Fukushima, H.; Ono, C.; Kikuchi, Y.; Koyama, R.; Matsui, K.; Furuyashiki, T.; Kida, S.; Tomita, H. Microarray dataset of gene transcription in mouse microglia and peripheral monocytes in contextual fear conditioning. Data Brief. 2023, 46, 108862. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Fukushima, H.; Ono, C.; Sakai, M.; Kasahara, Y.; Kikuchi, Y.; Gunawansa, N.; Takahashi, Y.; Matsuoka, H.; Kida, S.; et al. Microglial production of TNF-alpha is a key element of sustained fear memory. Brain Behav. Immun. 2017, 59, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Sakai, M.; Yu, Z.; Hirayama, R.; Nakasato, M.; Kikuchi, Y.; Ono, C.; Komatsu, H.; Nakanishi, M.; Yoshii, H.; Stellwagen, D.; et al. Deficient Autophagy in Microglia Aggravates Repeated Social Defeat Stress-Induced Social Avoidance. Neural Plast. 2022, 2022, 7503553. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Sakai, M.; Fukushima, H.; Ono, C.; Kikuchi, Y.; Koyama, R.; Matsui, K.; Furuyashiki, T.; Kida, S.; Tomita, H. Contextual fear conditioning regulates synapse-related gene transcription in mouse microglia. Brain Res. Bull. 2022, 189, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Sakai, M.; Yu, Z.; Taniguchi, M.; Picotin, R.; Oyama, N.; Stellwagen, D.; Ono, C.; Kikuchi, Y.; Matsui, K.; Nakanishi, M.; et al. N-Acetylcysteine Suppresses Microglial Inflammation and Induces Mortality Dose-Dependently via Tumor Necrosis Factor-α Signaling. Int. J. Mol. Sci. 2023, 24, 3798. [Google Scholar] [CrossRef] [PubMed]
- Barateiro, A.; Brites, D.; Fernandes, A. Oligodendrocyte Development and Myelination in Neurodevelopment: Molecular Mechanisms in Health and Disease. Curr. Pharm. Des. 2016, 22, 656–679. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, S.; Gritti, L.; Crooks, D.; Dombrowski, Y. Oligodendrocytes in Development, Myelin Generation and Beyond. Cells 2019, 8, 1424. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Zhu, Z.; Ransom, B.R.; Tong, X. Oligodendrocyte lineage cells and depression. Mol. Psychiatry 2021, 26, 103–117. [Google Scholar] [CrossRef]
- Pandey, G.N.; Rizavi, H.S.; Zhang, H.; Bhaumik, R.; Ren, X. Abnormal protein and mRNA expression of inflammatory cytokines in the prefrontal cortex of depressed individuals who died by suicide. J. Psychiatry Neurosci. 2018, 43, 376–385. [Google Scholar] [CrossRef]
- Monji, A.; Kato, T.; Kanba, S. Cytokines and schizophrenia: Microglia hypothesis of schizophrenia. Psychiatry Clin. Neurosci. 2009, 63, 257–265. [Google Scholar] [CrossRef]
- Courtet, P.; Giner, L.; Seneque, M.; Guillaume, S.; Olie, E.; Ducasse, D. Neuroinflammation in suicide: Toward a comprehensive model. World J. Biol. Psychiatry 2016, 17, 564–586. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; The PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. J. Clin. Epidemiol. 2009, 62, 1006–1012. [Google Scholar] [CrossRef] [PubMed]
- Miguel-Hidalgo, J.J.; Overholser, J.C.; Meltzer, H.Y.; Stockmeier, C.A.; Rajkowska, G. Reduced glial and neuronal packing density in the orbitofrontal cortex in alcohol dependence and its relationship with suicide and duration of alcohol dependence. Alcohol. Clin. Exp. Res. 2006, 30, 1845–1855. [Google Scholar] [CrossRef] [PubMed]
- Hercher, C.; Parent, M.; Flores, C.; Canetti, L.; Turecki, G.; Mechawar, N. Alcohol dependence-related increase of glial cell density in the anterior cingulate cortex of suicide completers. J. Psychiatry Neurosci. 2009, 34, 281–288. [Google Scholar] [PubMed]
- Smiley, J.F.; Hackett, T.A.; Bleiwas, C.; Petkova, E.; Stankov, A.; Mann, J.J.; Rosoklija, G.; Dwork, A.J. Reduced GABA neuron density in auditory cerebral cortex of subjects with major depressive disorder. J. Chem. Neuroanat. 2016, 76 Pt B, 108–121. [Google Scholar] [CrossRef]
- Underwood, M.D.; Bakalian, M.J.; Escobar, T.; Kassir, S.; Mann, J.J.; Arango, V. Early-Life Adversity, but Not Suicide, Is Associated With Less Prefrontal Cortex Gray Matter in Adulthood. Int. J. Neuropsychopharmacol. 2019, 22, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Boldrini, M.; Galfalvy, H.; Dwork, A.J.; Rosoklija, G.B.; Trencevska-Ivanovska, I.; Pavlovski, G.; Hen, R.; Arango, V.; Mann, J.J. Resilience Is Associated With Larger Dentate Gyrus, While Suicide Decedents With Major Depressive Disorder Have Fewer Granule Neurons. Biol. Psychiatry 2019, 85, 850–862. [Google Scholar] [CrossRef]
- Chen, F.; Bertelsen, A.B.; Holm, I.E.; Nyengaard, J.R.; Rosenberg, R.; Dorph-Petersen, K.A. Hippocampal volume and cell number in depression, schizophrenia, and suicide subjects. Brain Res. 2020, 1727, 146546. [Google Scholar] [CrossRef]
- Rubinow, M.J.; Mahajan, G.; May, W.; Overholser, J.C.; Jurjus, G.J.; Dieter, L.; Herbst, N.; Steffens, D.C.; Miguel-Hidalgo, J.J.; Rajkowska, G.; et al. Basolateral amygdala volume and cell numbers in major depressive disorder: A postmortem stereological study. Brain Struct. Funct. 2016, 221, 171–184. [Google Scholar] [CrossRef]
- Mahar, I.; Labonte, B.; Yogendran, S.; Isingrini, E.; Perret, L.; Davoli, M.A.; Rachalski, A.; Giros, B.; Turecki, G.; Mechawar, N. Disrupted hippocampal neuregulin-1/ErbB3 signaling and dentate gyrus granule cell alterations in suicide. Transl. Psychiatry 2017, 7, e1161. [Google Scholar] [CrossRef]
- Torres-Platas, S.G.; Hercher, C.; Davoli, M.A.; Maussion, G.; Labonté, B.; Turecki, G.; Mechawar, N. Astrocytic hypertrophy in anterior cingulate white matter of depressed suicides. Neuropsychopharmacology 2011, 36, 2650–2658. [Google Scholar] [CrossRef] [PubMed]
- Cobb, J.A.; O’Neill, K.; Milner, J.; Mahajan, G.J.; Lawrence, T.J.; May, W.L.; Miguel-Hidalgo, J.; Rajkowska, G.; Stockmeier, C.A. Density of GFAP-immunoreactive astrocytes is decreased in left hippocampi in major depressive disorder. Neuroscience 2016, 316, 209–220. [Google Scholar] [CrossRef]
- O’Leary, L.A.; Belliveau, C.; Davoli, M.A.; Ma, J.C.; Tanti, A.; Turecki, G.; Mechawar, N. Widespread Decrease of Cerebral Vimentin-Immunoreactive Astrocytes in Depressed Suicides. Front. Psychiatry 2021, 12, 640963. [Google Scholar] [CrossRef]
- Chandley, M.J.; Szebeni, K.; Szebeni, A.; Crawford, J.; Stockmeier, C.A.; Turecki, G.; Miguel-Hidalgo, J.J.; Ordway, G.A. Gene expression deficits in pontine locus coeruleus astrocytes in men with major depressive disorder. J. Psychiatry Neurosci. 2013, 38, 276–284. [Google Scholar] [CrossRef]
- Ernst, C.; Deleva, V.; Deng, X.; Sequeira, A.; Pomarenski, A.; Klempan, T.; Ernst, N.; Quirion, R.; Gratton, A.; Szyf, M.; et al. Alternative splicing, methylation state, and expression profile of tropomyosin-related kinase B in the frontal cortex of suicide completers. Arch. Gen. Psychiatry 2009, 66, 22–32. [Google Scholar] [CrossRef]
- Maussion, G.; Yang, J.; Suderman, M.; Diallo, A.; Nagy, C.; Arnovitz, M.; Mechawar, N.; Turecki, G. Functional DNA methylation in a transcript specific 3’UTR region of TrkB associates with suicide. Epigenetics 2014, 9, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.R.; Kamphuis, W.; Shan, L. Astrocyte Changes in the Prefrontal Cortex From Aged Non-suicidal Depressed Patients. Front. Cell Neurosci. 2019, 13, 503. [Google Scholar] [CrossRef] [PubMed]
- Steiner, J.; Mawrin, C.; Ziegeler, A.; Bielau, H.; Ullrich, O.; Bernstein, H.G.; Bogerts, B. Distribution of HLA-DR-positive microglia in schizophrenia reflects impaired cerebral lateralization. Acta Neuropathol. 2006, 112, 305–316. [Google Scholar] [CrossRef]
- Steiner, J.; Bielau, H.; Brisch, R.; Danos, P.; Ullrich, O.; Mawrin, C.; Bernstein, H.G.; Bogerts, B. Immunological aspects in the neurobiology of suicide: Elevated microglial density in schizophrenia and depression is associated with suicide. J. Psychiatr. Res. 2008, 42, 151–157. [Google Scholar] [CrossRef]
- Petrasch-Parwez, E.; Schöbel, A.; Benali, A.; Moinfar, Z.; Förster, E.; Brüne, M.; Juckel, G. Lateralization of increased density of Iba1-immunopositive microglial cells in the anterior midcingulate cortex of schizophrenia and bipolar disorder. Eur. Arch. Psychiatry Clin. Neurosci. 2020, 270, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Steiner, J.; Walter, M.; Gos, T.; Guillemin, G.J.; Bernstein, H.G.; Sarnyai, Z.; Mawrin, C.; Brisch, R.; Bielau, H.; Meyer zu Schwabedissen, L.; et al. Severe depression is associated with increased microglial quinolinic acid in subregions of the anterior cingulate gyrus: Evidence for an immune-modulated glutamatergic neurotransmission? J. Neuroinflamm. 2011, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Busse, M.; Busse, S.; Myint, A.M.; Gos, T.; Dobrowolny, H.; Müller, U.J.; Bogerts, B.; Bernstein, H.G.; Steiner, J. Decreased quinolinic acid in the hippocampus of depressive patients: Evidence for local anti-inflammatory and neuroprotective responses? Eur. Arch. Psychiatry Clin. Neurosci. 2015, 265, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Naggan, L.; Robinson, E.; Dinur, E.; Goldenberg, H.; Kozela, E.; Yirmiya, R. Suicide in bipolar disorder patients is associated with hippocampal microglia activation and reduction of lymphocytes-activation gene 3 (LAG3) microglial checkpoint expression. Brain Behav. Immun. 2023, 110, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Brisch, R.; Steiner, J.; Mawrin, C.; Krzyżanowska, M.; Jankowski, Z.; Gos, T. Microglia in the dorsal raphe nucleus plays a potential role in both suicide facilitation and prevention in affective disorders. Eur. Arch. Psychiatry Clin. Neurosci. 2017, 267, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Torres-Platas, S.G.; Cruceanu, C.; Chen, G.G.; Turecki, G.; Mechawar, N. Evidence for increased microglial priming and macrophage recruitment in the dorsal anterior cingulate white matter of depressed suicides. Brain Behav. Immun. 2014, 42, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Schnieder, T.P.; Trencevska, I.; Rosoklija, G.; Stankov, A.; Mann, J.J.; Smiley, J.; Dwork, A.J. Microglia of prefrontal white matter in suicide. J. Neuropathol. Exp. Neurol. 2014, 73, 880–890. [Google Scholar] [CrossRef]
- Honer, W.G.; Falkai, P.; Chen, C.; Arango, V.; Mann, J.J.; Dwork, A.J. Synaptic and plasticity-associated proteins in anterior frontal cortex in severe mental illness. Neuroscience 1999, 91, 1247–1255. [Google Scholar] [CrossRef]
- Tanti, A.; Kim, J.J.; Wakid, M.; Davoli, M.A.; Turecki, G.; Mechawar, N. Child abuse associates with an imbalance of oligodendrocyte-lineage cells in ventromedial prefrontal white matter. Mol. Psychiatry 2018, 23, 2018–2028. [Google Scholar] [CrossRef]
- Chandley, M.J.; Szebeni, A.; Szebeni, K.; Wang-Heaton, H.; Garst, J.; Stockmeier, C.A.; Lewis, N.H.; Ordway, G.A. Markers of elevated oxidative stress in oligodendrocytes captured from the brainstem and occipital cortex in major depressive disorder and suicide. Prog. Neuropsychopharmacol. Biol. Psychiatry 2022, 117, 110559. [Google Scholar] [CrossRef]
- Sha, Q.; Fu, Z.; Escobar Galvis, M.L.; Madaj, Z.; Underwood, M.D.; Steiner, J.A.; Dwork, A.; Simpson, N.; Galfalvy, H.; Rozoklija, G.; et al. Integrative transcriptome- and DNA methylation analysis of brain tissue from the temporal pole in suicide decedents and their controls. Mol. Psychiatry 2023, 29, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xiong, L.; Chen, J.; Wang, Q. Integrative Analyses of scRNA-seq, Bulk mRNA-seq, and DNA Methylation Profiling in Depressed Suicide Brain Tissues. Int. J. Neuropsychopharmacol. 2023, 26, 840–855. [Google Scholar] [CrossRef] [PubMed]
- Nagy, C.; Suderman, M.; Yang, J.; Szyf, M.; Mechawar, N.; Ernst, C.; Turecki, G. Astrocytic abnormalities and global DNA methylation patterns in depression and suicide. Mol. Psychiatry 2015, 20, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Torres-Platas, S.G.; Nagy, C.; Wakid, M.; Turecki, G.; Mechawar, N. Glial fibrillary acidic protein is differentially expressed across cortical and subcortical regions in healthy brains and downregulated in the thalamus and caudate nucleus of depressed suicides. Mol. Psychiatry 2016, 21, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Verwer, R.W.H.; Lucassen, P.J.; Huitinga, I.; Swaab, D.F. Prefrontal cortex alterations in glia gene expression in schizophrenia with and without suicide. J. Psychiatr. Res. 2020, 121, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Schlicht, K.; Büttner, A.; Siedler, F.; Scheffer, B.; Zill, P.; Eisenmenger, W.; Ackenheil, M.; Bondy, B. Comparative proteomic analysis with postmortem prefrontal cortex tissues of suicide victims versus controls. J. Psychiatr. Res. 2007, 41, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Ernst, C.; Nagy, C.; Kim, S.; Yang, J.P.; Deng, X.; Hellstrom, I.C.; Choi, K.H.; Gershenfeld, H.; Meaney, M.J.; Turecki, G. Dysfunction of astrocyte connexins 30 and 43 in dorsal lateral prefrontal cortex of suicide completers. Biol. Psychiatry 2011, 70, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Klempan, T.A.; Sequeira, A.; Canetti, L.; Lalovic, A.; Ernst, C.; ffrench-Mullen, J.; Turecki, G. Altered expression of genes involved in ATP biosynthesis and GABAergic neurotransmission in the ventral prefrontal cortex of suicides with and without major depression. Mol. Psychiatry 2009, 14, 175–189. [Google Scholar] [CrossRef]
- Zhao, J.; Verwer, R.W.; van Wamelen, D.J.; Qi, X.R.; Gao, S.F.; Lucassen, P.J.; Swaab, D.F. Prefrontal changes in the glutamate-glutamine cycle and neuronal/glial glutamate transporters in depression with and without suicide. J. Psychiatr. Res. 2016, 82, 8–15. [Google Scholar] [CrossRef]
- Iacobas, D.A.; Urban-Maldonado, M.; Iacobas, S.; Scemes, E.; Spray, D.C. Array analysis of gene expression in connexin-43 null astrocytes. Physiol. Genom. 2003, 15, 177–190. [Google Scholar] [CrossRef]
- Tanti, A.; Lutz, P.E.; Kim, J.; O’Leary, L.; Théroux, J.F.; Turecki, G.; Mechawar, N. Evidence of decreased gap junction coupling between astrocytes and oligodendrocytes in the anterior cingulate cortex of depressed suicides. Neuropsychopharmacology 2019, 44, 2099–2111. [Google Scholar] [CrossRef] [PubMed]
- Nagy, C.; Torres-Platas, S.G.; Mechawar, N.; Turecki, G. Repression of Astrocytic Connexins in Cortical and Subcortical Brain Regions and Prefrontal Enrichment of H3K9me3 in Depression and Suicide. Int. J. Neuropsychopharmacol. 2017, 20, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Miguel-Hidalgo, J.J.; Overholser, J.C.; Jurjus, G.J.; Meltzer, H.Y.; Dieter, L.; Konick, L.; Stockmeier, C.A.; Rajkowska, G. Vascular and extravascular immunoreactivity for intercellular adhesion molecule 1 in the orbitofrontal cortex of subjects with major depression: Age-dependent changes. J. Affect. Disord. 2011, 132, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Miguel-Hidalgo, J.J.; Wilson, B.A.; Hussain, S.; Meshram, A.; Rajkowska, G.; Stockmeier, C.A. Reduced connexin 43 immunolabeling in the orbitofrontal cortex in alcohol dependence and depression. J. Psychiatr. Res. 2014, 55, 101–109. [Google Scholar] [CrossRef] [PubMed]
- García-Gutiérrez, M.S.; Navarrete, F.; Navarro, G.; Reyes-Resina, I.; Franco, R.; Lanciego, J.L.; Giner, S.; Manzanares, J. Alterations in Gene and Protein Expression of Cannabinoid CB(2) and GPR55 Receptors in the Dorsolateral Prefrontal Cortex of Suicide Victims. Neurotherapeutics 2018, 15, 796–806. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Verwer, R.W.H.; Zhao, J.; Huitinga, I.; Lucassen, P.J.; Swaab, D.F. Changes in glial gene expression in the prefrontal cortex in relation to major depressive disorder, suicide and psychotic features. J. Affect. Disord. 2021, 295, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Punzi, G.; Ursini, G.; Chen, Q.; Radulescu, E.; Tao, R.; Huuki, L.A.; Di Carlo, P.; Collado-Torres, L.; Shin, J.H.; Catanesi, R.; et al. Genetics and Brain Transcriptomics of Completed Suicide. Am. J. Psychiatry 2022, 179, 226–241. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Verwer, R.W.H.; Lucassen, P.J.; Huitinga, I.; Swaab, D.F. Sex difference in glia gene expression in the dorsolateral prefrontal cortex in bipolar disorder: Relation to psychotic features. J. Psychiatr. Res. 2020, 125, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Holmes, S.E.; Hinz, R.; Conen, S.; Gregory, C.J.; Matthews, J.C.; Anton-Rodriguez, J.M.; Gerhard, A.; Talbot, P.S. Elevated Translocator Protein in Anterior Cingulate in Major Depression and a Role for Inflammation in Suicidal Thinking: A Positron Emission Tomography Study. Biol. Psychiatry 2018, 83, 61–69. [Google Scholar] [CrossRef]
- Zeng, D.; He, S.; Ma, C.; Wen, Y.; Song, W.; Xu, Q.; Zhao, N.; Wang, Q.; Yu, Y.; Shen, Y.; et al. Network-based approach to identify molecular signatures in the brains of depressed suicides. Psychiatry Res. 2020, 294, 113513. [Google Scholar] [CrossRef]
- Pantazatos, S.P.; Huang, Y.Y.; Rosoklija, G.B.; Dwork, A.J.; Arango, V.; Mann, J.J. Whole-transcriptome brain expression and exon-usage profiling in major depression and suicide: Evidence for altered glial, endothelial and ATPase activity. Mol. Psychiatry 2017, 22, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Klempan, T.A.; Ernst, C.; Deleva, V.; Labonte, B.; Turecki, G. Characterization of QKI gene expression, genetics, and epigenetics in suicide victims with major depressive disorder. Biol. Psychiatry 2009, 66, 824–831. [Google Scholar] [CrossRef] [PubMed]
- Nagy, C.; Maitra, M.; Tanti, A.; Suderman, M.; Théroux, J.F.; Davoli, M.A.; Perlman, K.; Yerko, V.; Wang, Y.C.; Tripathy, S.J.; et al. Single-nucleus transcriptomics of the prefrontal cortex in major depressive disorder implicates oligodendrocyte precursor cells and excitatory neurons. Nat. Neurosci. 2020, 23, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Sanacora, G.; Banasr, M. From pathophysiology to novel antidepressant drugs: Glial contributions to the pathology and treatment of mood disorders. Biol. Psychiatry 2013, 73, 1172–1179. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.H.; Du, Y.; Chen, L.; Cheng, Y. Glial Cell Abnormalities in Major Psychiatric Diseases: A Systematic Review of Postmortem Brain Studies. Mol. Neurobiol. 2022, 59, 1665–1692. [Google Scholar] [CrossRef] [PubMed]
- Taipale, H.; Lähteenvuo, M.; Tanskanen, A.; Mittendorfer-Rutz, E.; Tiihonen, J. Comparative Effectiveness of Antipsychotics for Risk of Attempted or Completed Suicide Among Persons With Schizophrenia. Schizophr. Bull. 2021, 47, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Reutfors, J.; Andersson, T.M.; Tanskanen, A.; DiBernardo, A.; Li, G.; Brandt, L.; Brenner, P. Risk Factors for Suicide and Suicide Attempts Among Patients With Treatment-Resistant Depression: Nested Case-Control Study. Arch. Suicide Res. 2021, 25, 424–438. [Google Scholar] [CrossRef] [PubMed]
- Selemon, L.D.; Lidow, M.S.; Goldman-Rakic, P.S. Increased volume and glial density in primate prefrontal cortex associated with chronic antipsychotic drug exposure. Biol. Psychiatry 1999, 46, 161–172. [Google Scholar] [CrossRef]
- Cotter, D.R.; Pariante, C.M.; Everall, I.P. Glial cell abnormalities in major psychiatric disorders: The evidence and implications. Brain Res. Bull. 2001, 55, 585–595. [Google Scholar] [CrossRef]
- Orsolini, L.; Latini, R.; Pompili, M.; Serafini, G.; Volpe, U.; Vellante, F.; Fornaro, M.; Valchera, A.; Tomasetti, C.; Fraticelli, S.; et al. Understanding the Complex of Suicide in Depression: From Research to Clinics. Psychiatry Investig. 2020, 17, 207–221. [Google Scholar] [CrossRef]
- Anestis, M.D.; Soberay, K.A.; Gutierrez, P.M.; Hernández, T.D.; Joiner, T.E. Reconsidering the link between impulsivity and suicidal behavior. Pers. Soc. Psychol. Rev. 2014, 18, 366–386. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.; Klein, K.M.; Buchman-Schmitt, J.M.; Hom, M.A.; Hagan, C.R.; Joiner, T.E. Routinized Assessment of Suicide Risk in Clinical Practice: An Empirically Informed Update. J. Clin. Psychol. 2015, 71, 1186–1200. [Google Scholar] [CrossRef] [PubMed]
- Janelidze, S.; Mattei, D.; Westrin, Å.; Träskman-Bendz, L.; Brundin, L. Cytokine levels in the blood may distinguish suicide attempters from depressed patients. Brain Behav. Immun. 2011, 25, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Ducasse, D.; Olié, E.; Guillaume, S.; Artéro, S.; Courtet, P. A meta-analysis of cytokines in suicidal behavior. Brain Behav. Immun. 2015, 46, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Snijders, G.; Sneeboer, M.A.M.; Fernández-Andreu, A.; Udine, E.; Boks, M.P.; Ormel, P.R.; van Berlekom, A.B.; van Mierlo, H.C.; Böttcher, C.; Priller, J.; et al. Distinct non-inflammatory signature of microglia in post-mortem brain tissue of patients with major depressive disorder. Mol. Psychiatry 2021, 26, 3336–3349. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.A.; Reis-de-Oliveira, G.; Pierone, B.C.; Martins-de-Souza, D.; Kaster, M.P. Depicting the molecular features of suicidal behavior: A review from an “omics” perspective. Psychiatry Res. 2024, 332, 115682. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.R.; Liu, J.C.; Bao, J.S.; Bai, Q.Q.; Wang, G.Q. Interaction of Microglia and Astrocytes in the Neurovascular Unit. Front. Immunol. 2020, 11, 1024. [Google Scholar] [CrossRef] [PubMed]
- Haruwaka, K.; Ikegami, A.; Tachibana, Y.; Ohno, N.; Konishi, H.; Hashimoto, A.; Matsumoto, M.; Kato, D.; Ono, R.; Kiyama, H.; et al. Dual microglia effects on blood brain barrier permeability induced by systemic inflammation. Nat. Commun. 2019, 10, 5816. [Google Scholar] [CrossRef] [PubMed]
- Kealy, J.; Greene, C.; Campbell, M. Blood-brain barrier regulation in psychiatric disorders. Neurosci. Lett. 2020, 726, 133664. [Google Scholar] [CrossRef]
- Wu, H.; Wang, J.; Teng, T.; Yin, B.; He, Y.; Jiang, Y.; Liu, X.; Yu, Y.; Li, X.; Zhou, X. Biomarkers of intestinal permeability and blood-brain barrier permeability in adolescents with major depressive disorder. J. Affect. Disord. 2023, 323, 659–666. [Google Scholar] [CrossRef]
- Ventorp, F.; Barzilay, R.; Erhardt, S.; Samuelsson, M.; Träskman-Bendz, L.; Janelidze, S.; Weizman, A.; Offen, D.; Brundin, L. The CD44 ligand hyaluronic acid is elevated in the cerebrospinal fluid of suicide attempters and is associated with increased blood-brain barrier permeability. J. Affect. Disord. 2016, 193, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Ueno, K.; Funayama, R.; Sakai, M.; Nariai, N.; Kojima, K.; Kikuchi, Y.; Li, X.; Ono, C.; Kanatani, J.; et al. Sex-Specific Differences in the Transcriptome of the Human Dorsolateral Prefrontal Cortex in Schizophrenia. Mol. Neurobiol. 2023, 60, 1083–1098. [Google Scholar] [CrossRef] [PubMed]
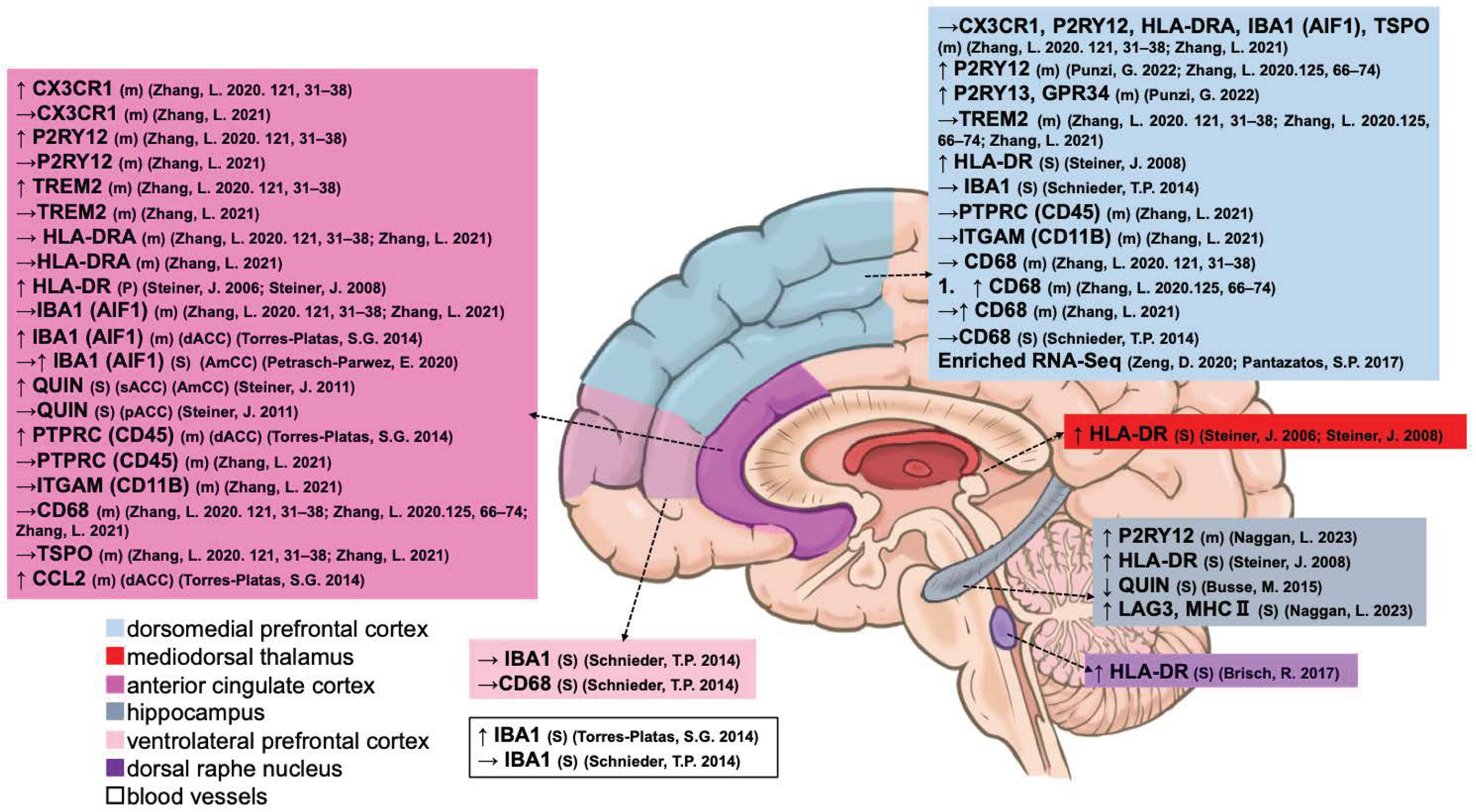
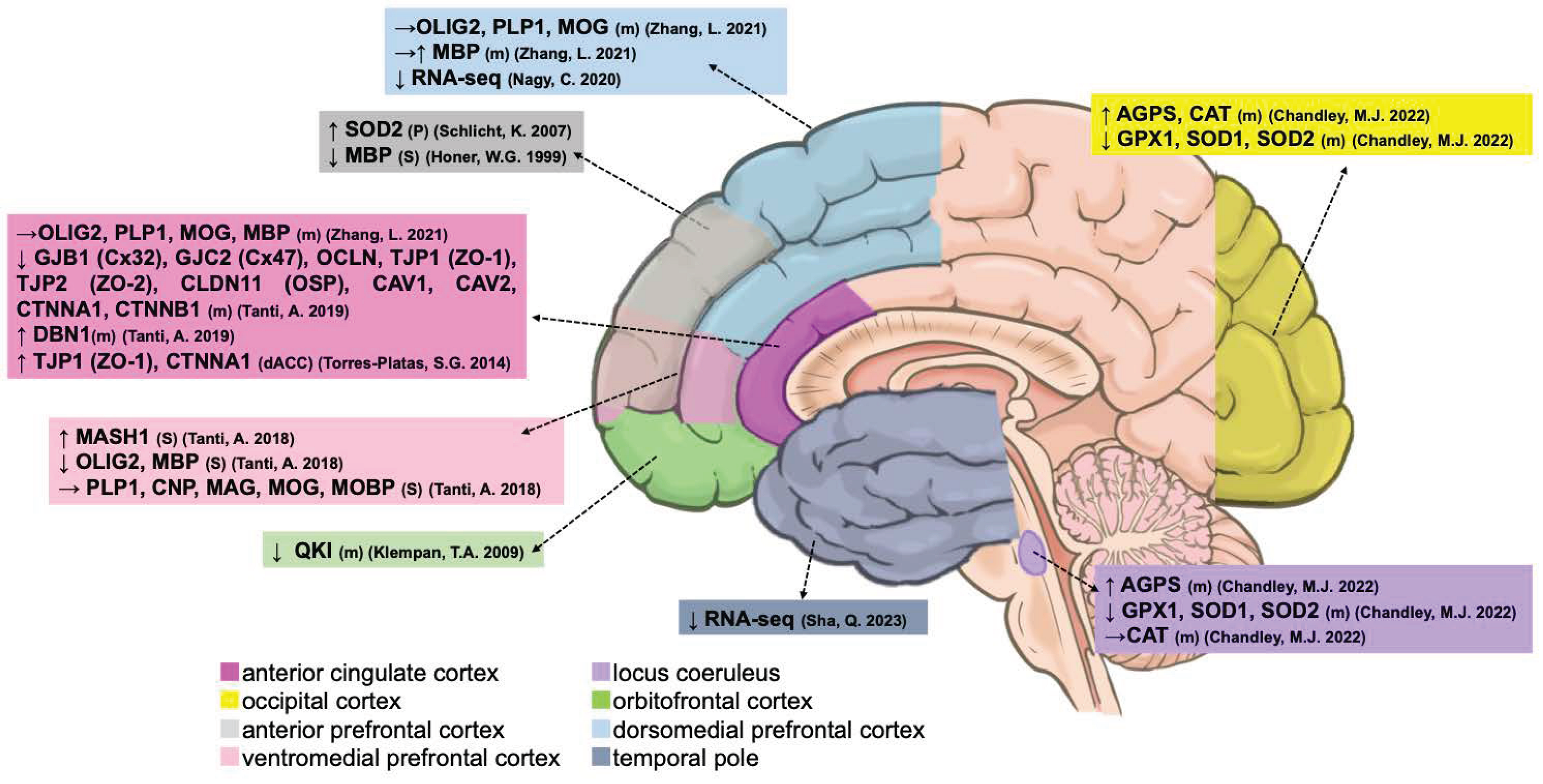
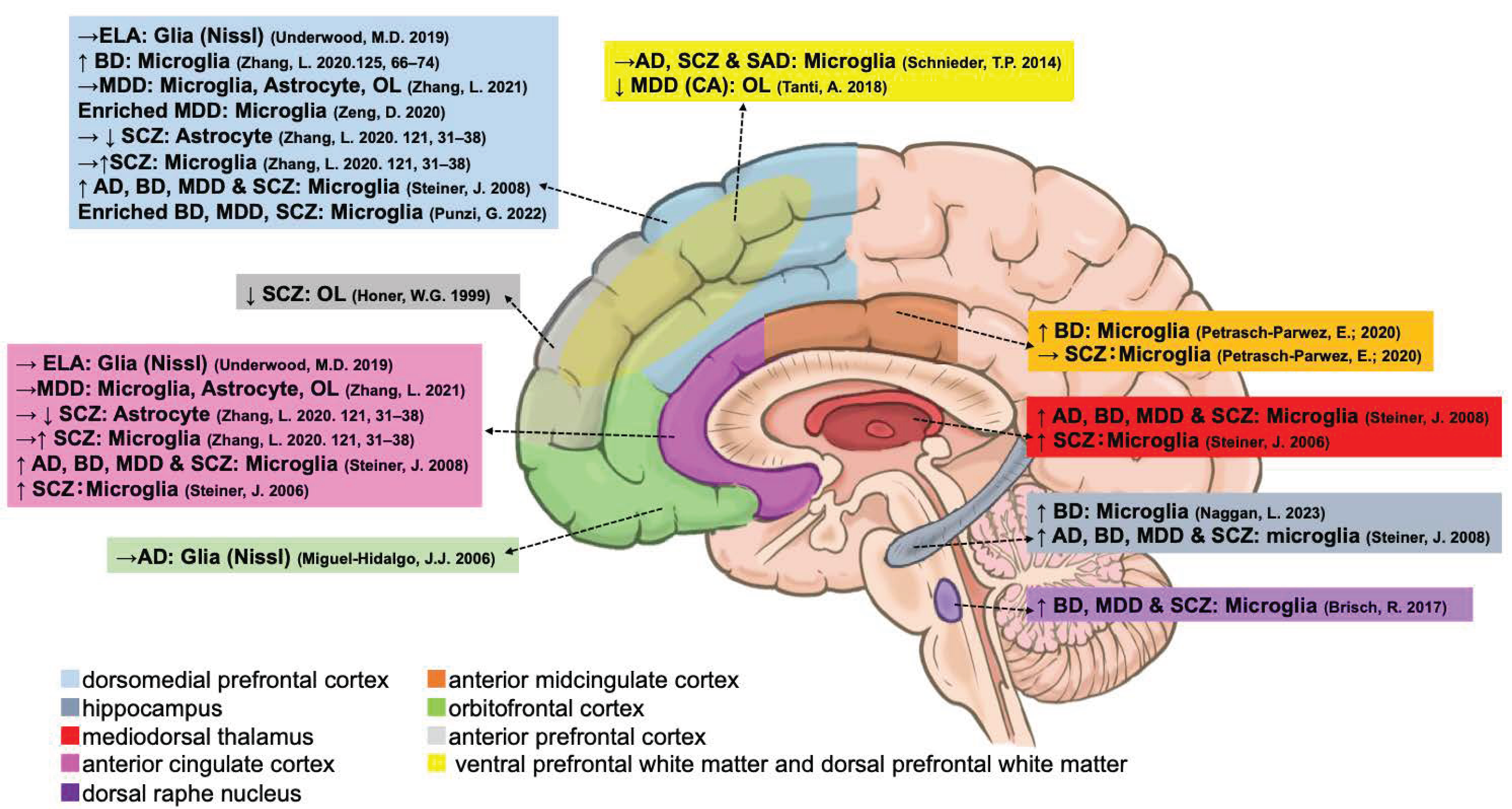
| Microglia | Type | ACC | dACC | sACC | pACC | DLPFC | VLPFC | MD | AmCC | HIPP | DRN | Blood Vessels |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CX3CR1 | mRNA | ↑ [65] → [76] | — | — | — | → [65,76] | — | — | — | — | — | — |
| P2RY12 | mRNA | ↑ [65] → [76] | — | — | — | ↑ [77,78] → [65,76] | — | — | — | ↑ [54] | — | — |
| P2RY13 | mRNA | — | — | — | — | ↑ [77] | — | — | — | — | — | — |
| GPR34 | mRNA | — | — | — | — | ↑ [77] | — | — | — | — | — | — |
| TREM2 | mRNA | ↑ [65] → [76] | — | — | — | → [65,76,78] | — | — | — | — | — | — |
| HLA-DRA | mRNA | → [65,76] | — | — | — | → [65,76] | — | — | — | — | — | — |
| HLA-DR | Stain | ↑ [49] #, [50] $ | — | — | — | ↑ [50] $ | — | ↑ [49] #, [50] $ | — | ↑ [50] $ | ↑ [55] | — |
| IBA1 (AIF1) | mRNA | → [65,76] | ↑ [56] | — | — | → [65,76] | — | — | — | — | — | — |
| IBA1 | Stain | — | — | — | — | → [57] | → [57] | — | →↑ [51] | — | — | ↑ [56] → [57] |
| QUIN | Stain | — | — | ↑ [52] | → [52] | — | — | — | ↑ [52] | ↓ [53] | — | — |
| PTPRC (CD45) | mRNA | → [76] | ↑ [56] | — | — | → [76] | — | — | — | — | — | — |
| ITGAM (CD11B) | mRNA | → [76] | — | — | — | → [76] | — | — | — | — | — | — |
| CD68 | mRNA | → [65,76,78] | — | — | — | → [65] ↑ [78] →↑ [76] | — | — | — | — | — | — |
| CD68 | Stain | — | — | — | — | → [57] | → [57] | — | — | — | — | — |
| TSPO | mRNA | → [65,76] | — | — | — | → [65,76] | — | — | — | — | — | — |
| LAG3 | Stain | — | — | — | — | — | — | — | — | ↑ [54] | — | — |
| MHC II | Stain | — | — | — | — | — | — | — | — | ↑ [54] | — | — |
| CCL2 | mRNA | — | ↑ [56] | — | — | — | — | — | — | — | — | — |
| RNA-Seq | mRNA | — | — | — | — | Enriched [80] | — | — | — | — | — | — |
| RNA-Seq | mRNA | — | — | — | — | Enriched [81] | — | — | — | — | — | — |
| Oligodendrocyte | Type | ACC | dACC | aPFC | DLPFC | VMPFC | LC | OCC | OFC | TP |
|---|---|---|---|---|---|---|---|---|---|---|
| AGPS | mRNA | — | — | — | — | — | ↑ [60] | ↑ [60] | — | — |
| SOD1 | mRNA | — | — | — | — | — | ↓ [60] | ↓ [60] | — | — |
| SOD2 | mRNA | — | — | — | — | — | ↓ [60] | ↓ [60] | — | — |
| SOD2 | Protein | — | — | ↑ [66] | — | — | — | — | — | — |
| GPX1 | mRNA | — | — | — | — | — | ↓ [60] | ↓ [60] | — | — |
| CAT | mRNA | — | — | — | — | — | → [60] | ↑ [60] | — | — |
| OLIG2 | mRNA | → [76] | — | — | → [76] | — | — | — | — | — |
| OLIG2 | Stain | — | — | — | — | ↓ [59] | — | — | — | — |
| PLP1 | mRNA | → [76] | — | — | → [76] | — | — | — | — | — |
| PLP1 | Stain | — | — | — | — | → [59] | — | — | — | — |
| CNP | Stain | — | — | — | — | → [59] | — | — | — | — |
| MAG | Stain | — | — | — | — | → [59] | — | — | — | — |
| MASH1 | Stain | — | — | — | — | ↑ [59] | — | — | — | — |
| MOG | mRNA | → [76] | — | — | → [76] | — | — | — | — | — |
| MOG | Stain | — | — | — | — | → [59] | — | — | — | — |
| MOBP | Stain | — | — | — | — | → [59] | — | — | — | — |
| MBP | mRNA | → [76] | — | — | →↑ [76] | — | — | — | — | |
| MBP | Stain | — | — | ↓ [58] | — | ↓ [59] | — | — | — | — |
| GJB1 (Cx32) | mRNA | ↓ [71] | — | — | — | — | — | — | — | — |
| GJC2 (Cx47) | mRNA | ↓ [71] | — | — | — | — | — | — | — | — |
| OCLN | mRNA | ↓ [71] | — | — | — | — | — | — | — | — |
| TJP1 (ZO-1) | mRNA | ↓ [71] | ↑ [56] | — | — | — | — | — | — | — |
| TJP2 (ZO-2) | mRNA | ↓ [71] | — | — | — | — | — | — | — | |
| CLDN11 (OSP) | mRNA | ↓ [71] | — | — | — | — | — | — | — | — |
| CAV1 | mRNA | ↓ [71] | — | — | — | — | — | — | — | — |
| CAV2 | mRNA | ↓ [71] | — | — | — | — | — | — | — | — |
| CTNNA1 | mRNA | ↓ [71] | ↑ [56] | — | — | — | — | — | — | — |
| CTNNB1 | mRNA | ↓ [71] | — | — | — | — | — | — | — | — |
| DBN1 | mRNA | ↑ [71] | — | — | — | — | — | — | — | — |
| QKI | mRNA | — | — | — | — | — | — | — | ↓ [82] | — |
| RNA-Seq | mRNA | — | — | — | ↓ [83] | — | — | — | — | ↓ [61] |
| Diagnoses | Cell Type | Changes | Region |
|---|---|---|---|
| Ad | Glia (Nissl) | → [34] | OFC |
| ELA | Glia (Nissl) | → [37] | ACC, DLPFC |
| BD | Microglia | ↑ [54] | HIPP |
| BD | Microglia | ↑ [78] | DLPFC |
| BD | Microglia | ↑ [51] | AmCC |
| MDD | Astrocyte | → [76] | ACC, DLPFC |
| MDD | Microglia | → [76] | ACC, DLPFC |
| MDD | OL | → [76] | ACC, DLPFC |
| MDD | Microglia | enriched [80] | DLPFC |
| SCZ | Astrocyte | →↓ [65] | ACC, DLPFC |
| SCZ | Microglia | →↑ [65] | ACC, DLPFC |
| AD, BD, MDD, and SCZ | Microglia | ↑ [50] $ | ACC, DLPFC, HIPP, MD |
| SCZ | Microglia | ↑ [49] # | ACC, MD |
| AD, SCZ, and SAD | Microglia | → [57] | VD |
| SCZ | Microglia | → [51] | AmCC |
| SCZ | OL | ↓ [58] | aPFC |
| BD MDD SCZ | Microglia | ↑ [55] | DRN |
| MDD (CA) | OL | ↓ [59] | VD |
| BD MDD SCZ | Microglia | enriched [77] | DLPFC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamamoto, M.; Sakai, M.; Yu, Z.; Nakanishi, M.; Yoshii, H. Glial Markers of Suicidal Behavior in the Human Brain—A Systematic Review of Postmortem Studies. Int. J. Mol. Sci. 2024, 25, 5750. https://doi.org/10.3390/ijms25115750
Yamamoto M, Sakai M, Yu Z, Nakanishi M, Yoshii H. Glial Markers of Suicidal Behavior in the Human Brain—A Systematic Review of Postmortem Studies. International Journal of Molecular Sciences. 2024; 25(11):5750. https://doi.org/10.3390/ijms25115750
Chicago/Turabian StyleYamamoto, Mana, Mai Sakai, Zhiqian Yu, Miharu Nakanishi, and Hatsumi Yoshii. 2024. "Glial Markers of Suicidal Behavior in the Human Brain—A Systematic Review of Postmortem Studies" International Journal of Molecular Sciences 25, no. 11: 5750. https://doi.org/10.3390/ijms25115750
APA StyleYamamoto, M., Sakai, M., Yu, Z., Nakanishi, M., & Yoshii, H. (2024). Glial Markers of Suicidal Behavior in the Human Brain—A Systematic Review of Postmortem Studies. International Journal of Molecular Sciences, 25(11), 5750. https://doi.org/10.3390/ijms25115750