
The Ins and Outs of Homeodomain-Leucine Zipper/Hormone Networks in the Regulation of Plant Development


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. HD-ZIP Protein Structure
3. Embryogenesis
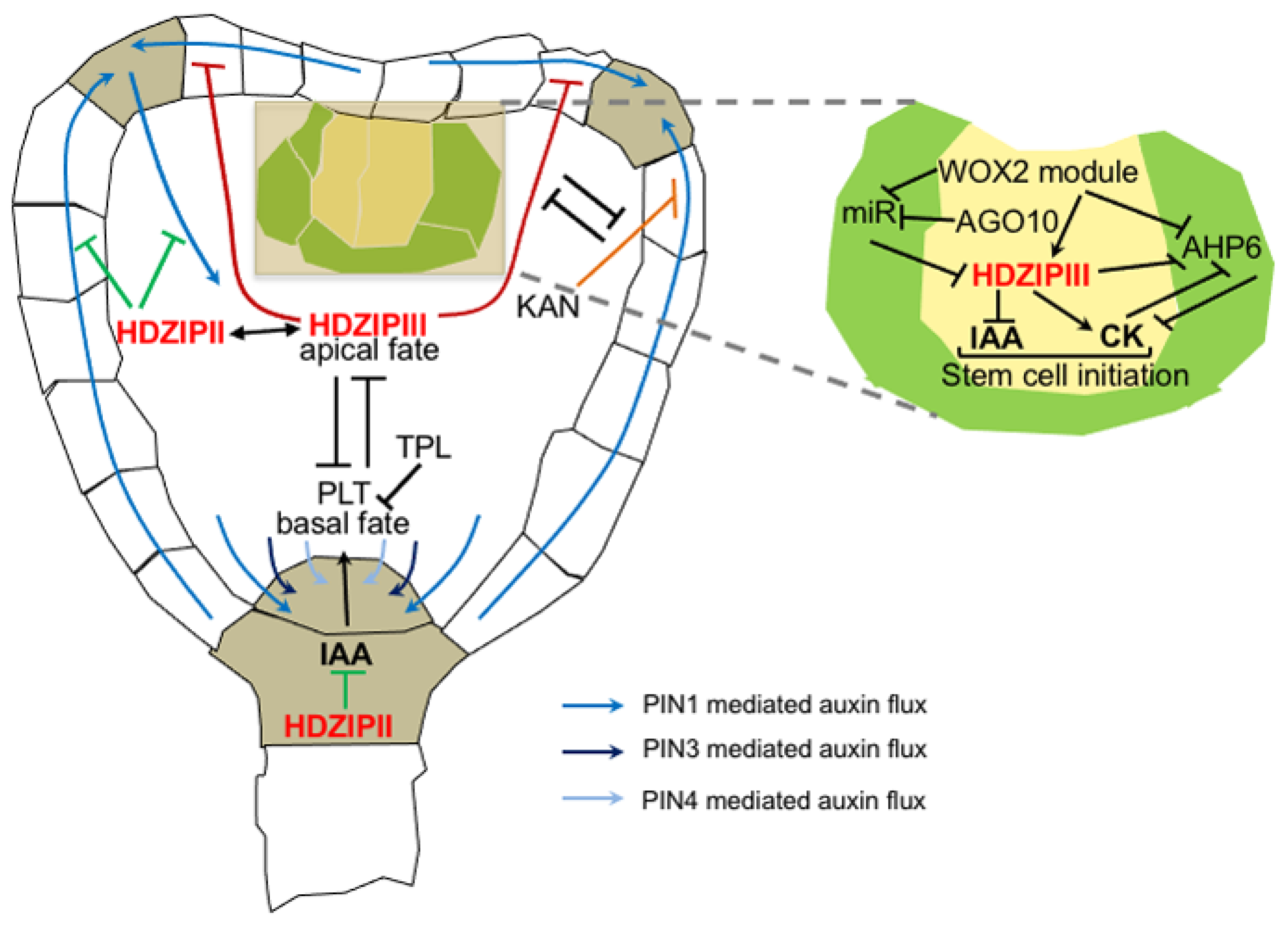
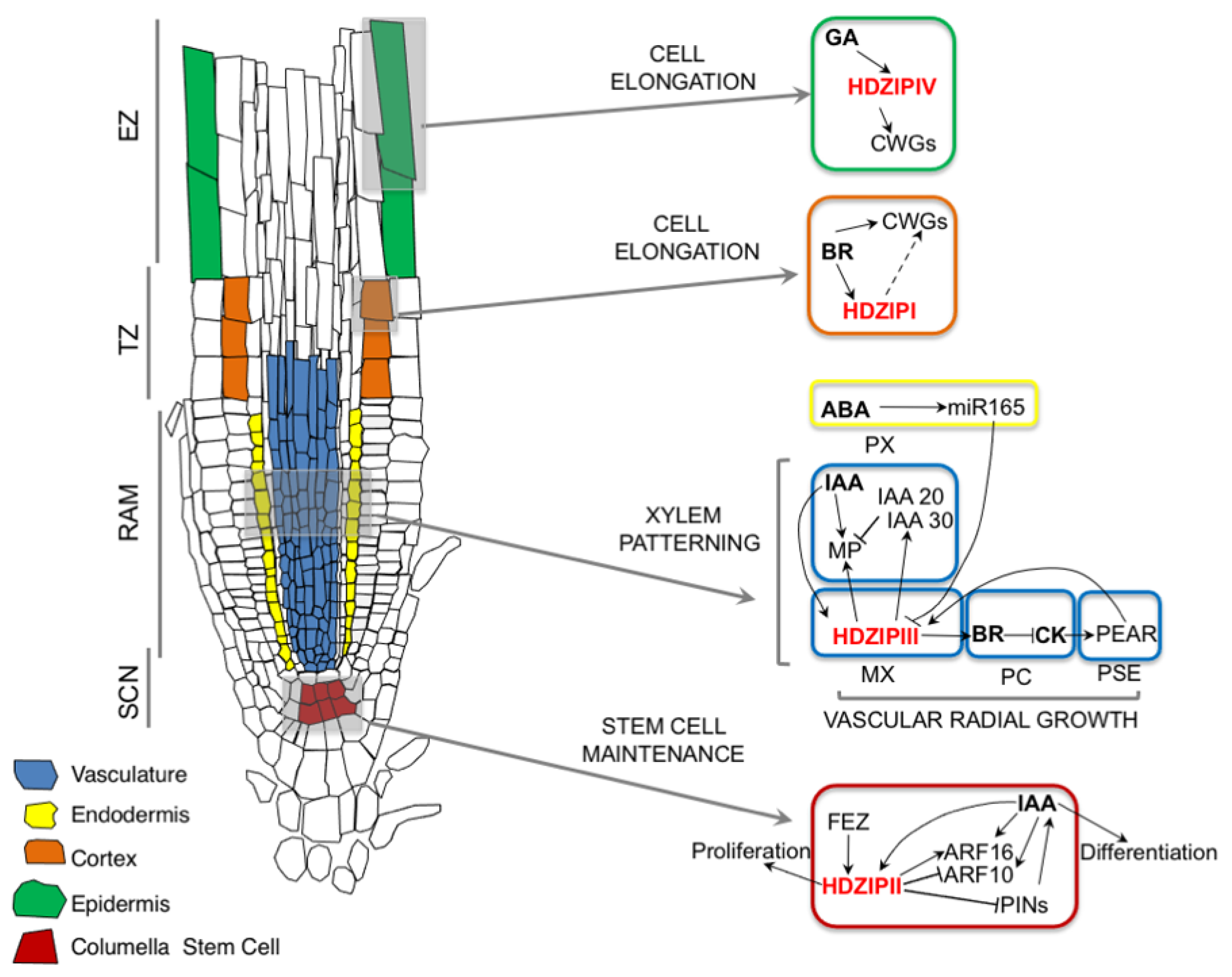
4. Root Development and Patterning
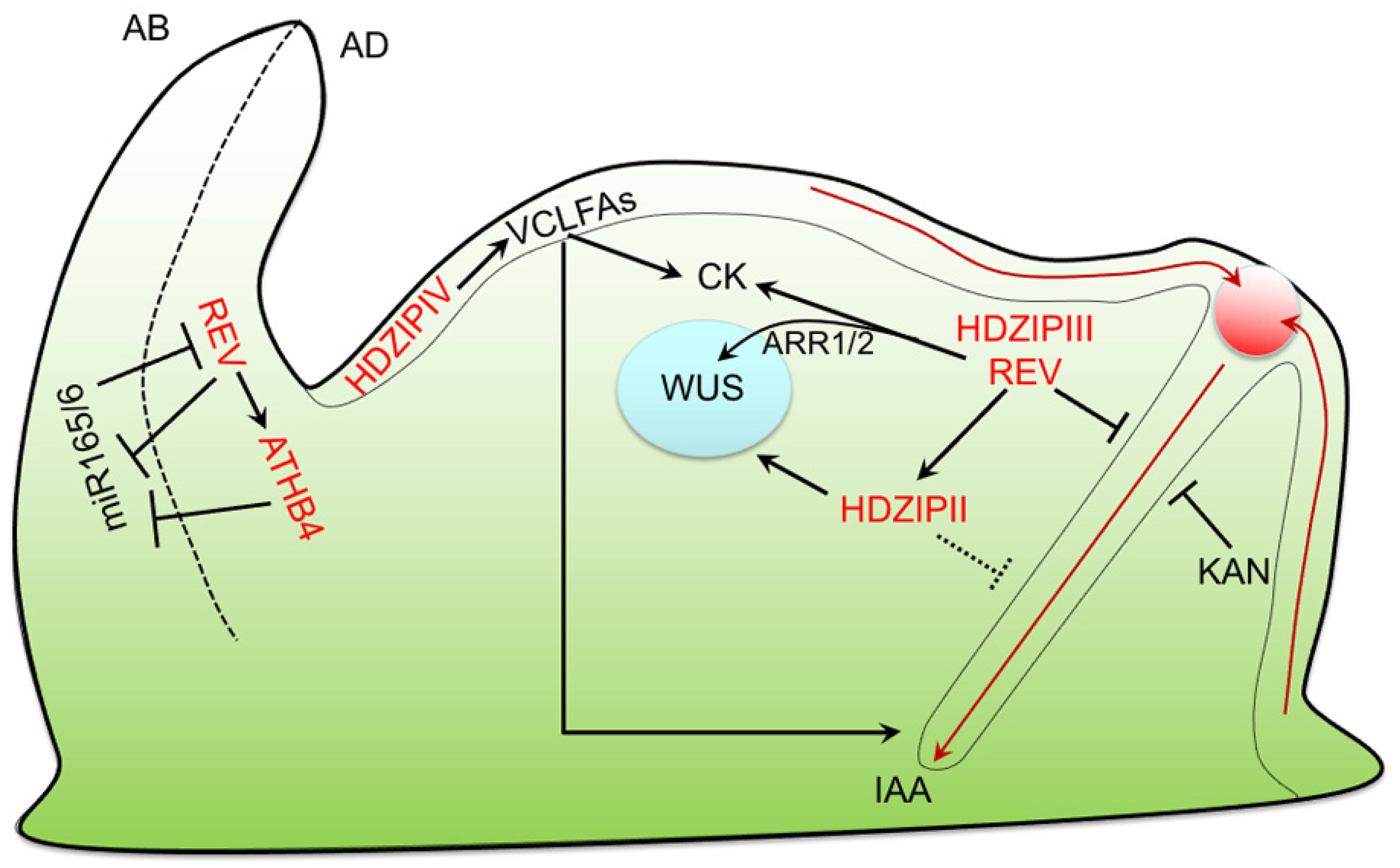
5. SAM Maintenance and Organogenesis
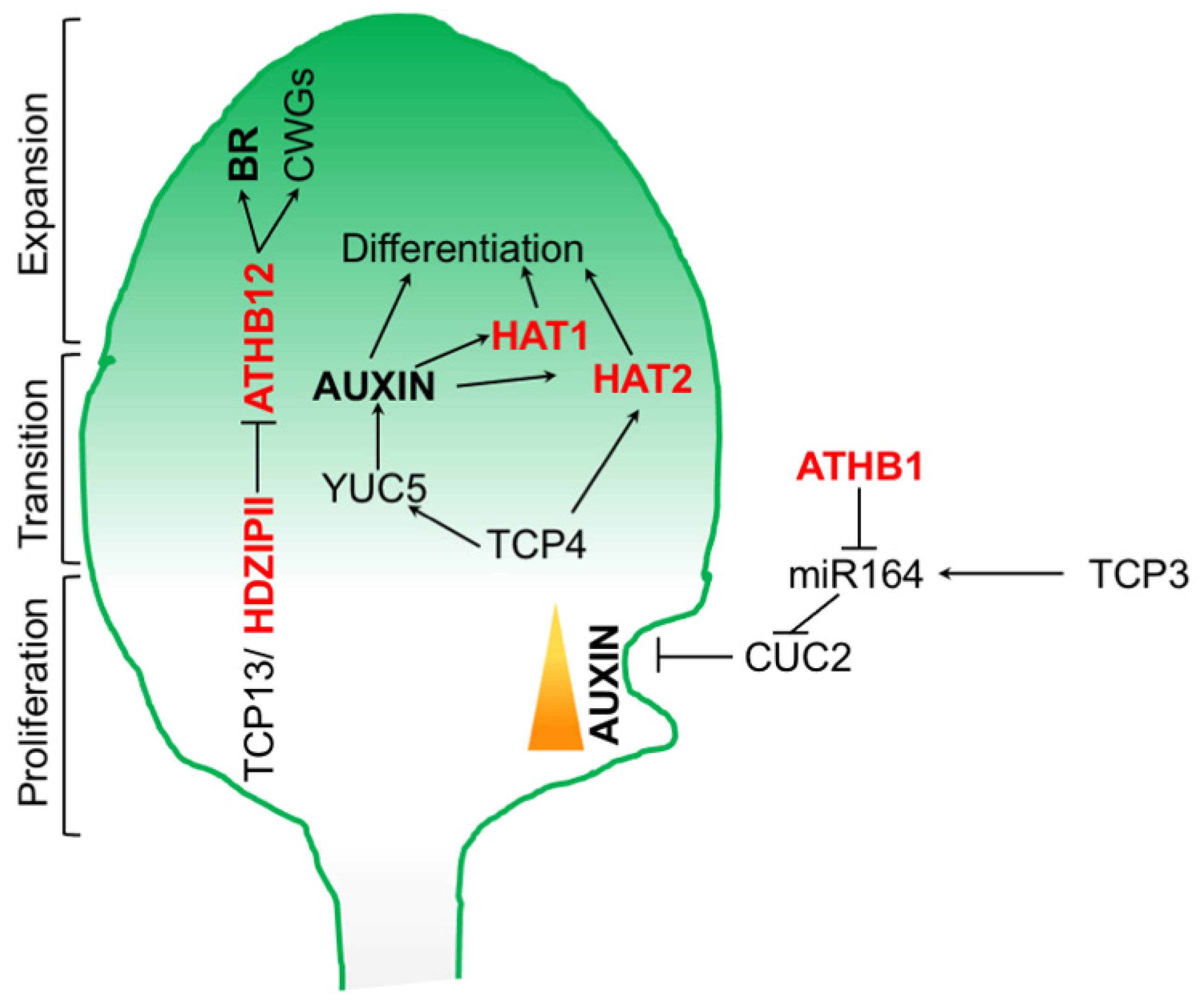
6. Leaf Development
7. Flower and Inflorescence Development
8. Concluding Remarks and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Coen, E.; Rolland-Lagan, A.-G.; Matthews, M.; Bangham, J.A.; Prusinkiewicz, P. The Genetics of Geometry. Proc. Natl. Acad. Sci. USA 2004, 101, 4728–4735. [Google Scholar] [CrossRef] [PubMed]
- Kicheva, A.; Briscoe, J. Control of Tissue Development by Morphogens. Annu. Rev. Cell Dev. Biol. 2023, 39, 91–121. [Google Scholar] [CrossRef] [PubMed]
- Hamant, O.; Saunders, T.E. Shaping Organs: Shared Structural Principles Across Kingdoms. Annu. Rev. Cell Dev. Biol. 2020, 36, 385–410. [Google Scholar] [CrossRef] [PubMed]
- Burda, I.; Martin, A.C.; Roeder, A.H.K.; Collins, M.A. The Dynamics and Biophysics of Shape Formation: Common Themes in Plant and Animal Morphogenesis. Dev. Cell 2023, 58, 2850–2866. [Google Scholar] [CrossRef] [PubMed]
- Pires, N.; Dolan, L. Morphological Evolution in Land Plants: New Designs with Old Genes. Phil Trans. R. Soc. B 2012, 367, 508–518. [Google Scholar] [CrossRef]
- Bowman, J.L.; Briginshaw, L.N.; Fisher, T.J.; Flores-Sandoval, E. Something Ancient and Something Neofunctionalized—Evolution of Land Plant Hormone Signaling Pathways. Curr. Opin. Plant Biol. 2019, 47, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.L. Walkabout on the Long Branches of Plant Evolution. Curr. Opin. Plant Biol. 2013, 16, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Donoghue, P.C.J.; Harrison, C.J.; Paps, J.; Schneider, H. The Evolutionary Emergence of Land Plants. Curr. Biol. 2021, 31, R1281–R1298. [Google Scholar] [CrossRef] [PubMed]
- Romani, F.; Reinheimer, R.; Florent, S.N.; Bowman, J.L.; Moreno, J.E. Evolutionary History of HOMEODOMAIN LEUCINE ZIPPER Transcription Factors during Plant Transition to Land. New Phytol. 2018, 219, 408–421. [Google Scholar] [CrossRef]
- Whitewoods, C.D.; Gonçalves, B.; Cheng, J.; Cui, M.; Kennaway, R.; Lee, K.; Bushell, C.; Yu, M.; Piao, C.; Coen, E. Evolution of Carnivorous Traps from Planar Leaves through Simple Shifts in Gene Expression. Science 2020, 367, 91–96. [Google Scholar] [CrossRef]
- Vlad, D.; Kierzkowski, D.; Rast, M.I.; Vuolo, F.; Dello Ioio, R.; Galinha, C.; Gan, X.; Hajheidari, M.; Hay, A.; Smith, R.S.; et al. Leaf Shape Evolution Through Duplication, Regulatory Diversification, and Loss of a Homeobox Gene. Science 2014, 343, 780–783. [Google Scholar] [CrossRef] [PubMed]
- Reymond, M.C.; Brunoud, G.; Chauvet, A.; Martínez-Garcia, J.F.; Martin-Magniette, M.-L.; Monéger, F.; Scutt, C.P. A Light-Regulated Genetic Module Was Recruited to Carpel Development in Arabidopsis Following a Structural Change to SPATULA. Plant Cell 2012, 24, 2812–2825. [Google Scholar] [CrossRef] [PubMed]
- Perotti, M.F.; Arce, A.L.; Chan, R.L. The Underground Life of Homeodomain-Leucine Zipper Transcription Factors. J. Exp. Bot. 2021, 72, 4005–4021. [Google Scholar] [CrossRef] [PubMed]
- Sessa, G.; Morelli, G.; Ruberti, I. The Athb-1 and -2 HD-Zip Domains Homodimerize Forming Complexes of Different DNA Binding Specificities. EMBO J. 1993, 12, 3507–3517. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, R.C.; Huang, S.C.; Song, L.; Lewsey, M.G.; Bartlett, A.; Nery, J.R.; Galli, M.; Gallavotti, A.; Ecker, J.R. Cistrome and Epicistrome Features Shape the Regulatory DNA Landscape. Cell 2016, 165, 1280–1292. [Google Scholar] [CrossRef]
- Abe, M.; Takahashi, T.; Komeda, Y. Identification of a Cis-Regulatory Element for L1 Layer-Specific Gene Expression, Which Is Targeted by an L1-Specific Homeodomain Protein. Plant J. 2001, 26, 487–494. [Google Scholar] [CrossRef]
- Arce, A.L.; Raineri, J.; Capella, M.; Cabello, J.V.; Chan, R.L. Uncharacterized Conserved Motifs Outside the HD-Zip Domain in HD-Zip Subfamily I Transcription Factors; a Potential Source of Functional Diversity. BMC Plant Biol. 2011, 11, 42. [Google Scholar] [CrossRef]
- Ciarbelli, A.R.; Ciolfi, A.; Salvucci, S.; Ruzza, V.; Possenti, M.; Carabelli, M.; Fruscalzo, A.; Sessa, G.; Morelli, G.; Ruberti, I. The Arabidopsis Homeodomain-Leucine Zipper II Gene Family: Diversity and Redundancy. Plant Mol. Biol. 2008, 68, 465–478. [Google Scholar] [CrossRef]
- Tron, A.E.; Bertoncini, C.W.; Chan, R.L.; Gonzalez, D.H. Redox Regulation of Plant Homeodomain Transcription Factors. J. Biol. Chem. 2002, 277, 34800–34807. [Google Scholar] [CrossRef]
- Schrick, K.; Bruno, M.; Khosla, A.; Cox, P.N.; Marlatt, S.A.; Roque, R.A.; Nguyen, H.C.; He, C.; Snyder, M.P.; Singh, D.; et al. Shared Functions of Plant and Mammalian StAR-Related Lipid Transfer (START) Domains in Modulating Transcription Factor Activity. BMC Biol. 2014, 12, 70. [Google Scholar] [CrossRef]
- Mukherjee, K.; Bürglin, T.R. MEKHLA, a Novel Domain with Similarity to PAS Domains, Is Fused to Plant Homeodomain-Leucine Zipper III Proteins. Plant Physiol. 2006, 140, 1142–1150. [Google Scholar] [CrossRef] [PubMed]
- Schrick, K.; Ahmad, B.; Nguyen, H.V. HD-Zip IV Transcription Factors: Drivers of Epidermal Cell Fate Integrate Metabolic Signals. Curr. Opin. Plant Biol. 2023, 75, 102417. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, T.; Subedi, B.; Khosla, A.; Begler, E.M.; Stephens, P.M.; Warner, A.L.; Lerma-Reyes, R.; Thompson, K.A.; Gunewardena, S.; Schrick, K. The START Domain Mediates Arabidopsis GLABRA2 Dimerization and Turnover Independently of Homeodomain DNA Binding. Plant Physiol. 2022, 190, 2315–2334. [Google Scholar] [CrossRef] [PubMed]
- Husbands, A.Y.; Feller, A.; Aggarwal, V.; Dresden, C.E.; Holub, A.S.; Ha, T.; Timmermans, M.C.P. The START Domain Potentiates HD-ZIPIII Transcriptional Activity. Plant Cell 2023, 35, 2332–2348. [Google Scholar] [CrossRef] [PubMed]
- Roodbarkelari, F.; Groot, E.P. Regulatory Function of Homeodomain-Leucine Zipper (HD-ZIP) Family Proteins during Embryogenesis. New Phytol. 2017, 213, 95–104. [Google Scholar] [CrossRef]
- Ueda, M.; Aichinger, E.; Gong, W.; Groot, E.; Verstraeten, I.; Vu, L.D.; Smet, I.D.; Higashiyama, T.; Umeda, M.; Laux, T. Transcriptional Integration of Paternal and Maternal Factors in the Arabidopsis Zygote. Genes Dev. 2017, 31, 617–627. [Google Scholar] [CrossRef]
- Ueda, M.; Zhang, Z.; Laux, T. Transcriptional Activation of Arabidopsis Axis Patterning Genes WOX8/9 Links Zygote Polarity to Embryo Development. Dev. Cell 2011, 20, 264–270. [Google Scholar] [CrossRef]
- Breuninger, H.; Rikirsch, E.; Hermann, M.; Ueda, M.; Laux, T. Differential Expression of WOX Genes Mediates Apical-Basal Axis Formation in the Arabidopsis Embryo. Dev. Cell 2008, 14, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Horstman, A.; Fukuoka, H.; Muino, J.M.; Nitsch, L.; Guo, C.; Passarinho, P.; Sanchez-Perez, G.; Immink, R.; Angenent, G.; Boutilier, K. AIL and HDG Proteins Act Antagonistically to Control Cell Proliferation. Development 2015, 142, 454–464. [Google Scholar] [CrossRef]
- Galinha, C.; Hofhuis, H.; Luijten, M.; Willemsen, V.; Blilou, I.; Heidstra, R.; Scheres, B. PLETHORA Proteins as Dose-Dependent Master Regulators of Arabidopsis Root Development. Nature 2007, 449, 1053–1057. [Google Scholar] [CrossRef]
- Aida, M.; Beis, D.; Heidstra, R.; Willemsen, V.; Blilou, I.; Galinha, C.; Nussaume, L.; Noh, Y.-S.; Amasino, R.; Scheres, B. The PLETHORA Genes Mediate Patterning of the Arabidopsis Root Stem Cell Niche. Cell 2004, 119, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, N.; Wu, M.-F.; Winter, C.M.; Berns, M.C.; Nole-Wilson, S.; Yamaguchi, A.; Coupland, G.; Krizek, B.A.; Wagner, D. A Molecular Framework for Auxin-Mediated Initiation of Flower Primordia. Dev. Cell 2013, 24, 271–282. [Google Scholar] [CrossRef]
- Boutilier, K.; Offringa, R.; Sharma, V.K.; Kieft, H.; Ouellet, T.; Zhang, L.; Hattori, J.; Liu, C.-M.; van Lammeren, A.A.M.; Miki, B.L.A.; et al. Ectopic Expression of BABY BOOM Triggers a Conversion from Vegetative to Embryonic Growth. Plant Cell 2002, 14, 1737–1749. [Google Scholar] [CrossRef] [PubMed]
- Abe, M. Regulation of Shoot Epidermal Cell Differentiation by a Pair of Homeodomain Proteins in Arabidopsis. Development 2003, 130, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Takada, S.; Jürgens, G. Transcriptional Regulation of Epidermal Cell Fate in the Arabidopsis Embryo. Development 2007, 134, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- Iida, H.; Yoshida, A.; Takada, S. ATML1 Activity Is Restricted to the Outermost Cells of the Embryo through Post-Transcriptional Repressions. Development 2019, 146, dev169300. [Google Scholar] [CrossRef]
- Nagata, K.; Ishikawa, T.; Kawai-Yamada, M.; Takahashi, T.; Abe, M. Ceramides Mediate Positional Signals in Arabidopsis Thaliana Protoderm Differentiation. Development 2021, 148, dev194969. [Google Scholar] [CrossRef] [PubMed]
- San-Bento, R.; Farcot, E.; Galletti, R.; Creff, A.; Ingram, G. Epidermal Identity Is Maintained by Cell–Cell Communication via a Universally Active Feedback Loop in Arabidopsis Thaliana. Plant J. 2014, 77, 46–58. [Google Scholar] [CrossRef]
- Smith, Z.R.; Long, J.A. Control of Arabidopsis Apical–Basal Embryo Polarity by Antagonistic Transcription Factors. Nature 2010, 464, 423–426. [Google Scholar] [CrossRef]
- Emery, J.F.; Floyd, S.K.; Alvarez, J.; Eshed, Y.; Hawker, N.P.; Izhaki, A.; Baum, S.F.; Bowman, J.L. Radial Patterning of Arabidopsis Shoots by Class III HD-ZIP and KANADI Genes. Curr. Biol. 2003, 13, 1768–1774. [Google Scholar] [CrossRef]
- Floyd, S.K.; Bowman, J.L. Gene Regulation: Ancient microRNA Target Sequences in Plants. Nature 2004, 428, 485–486. [Google Scholar] [CrossRef] [PubMed]
- Miyashima, S.; Honda, M.; Hashimoto, K.; Tatematsu, K.; Hashimoto, T.; Sato-Nara, K.; Okada, K.; Nakajima, K. A Comprehensive Expression Analysis of the Arabidopsis MICRORNA165/6 Gene Family during Embryogenesis Reveals a Conserved Role in Meristem Specification and a Non-Cell-Autonomous Function. Plant Cell Physiol. 2013, 54, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.; Grigg, S.P.; Xie, M.; Christensen, S.; Fletcher, J.C. Regulation of Arabidopsis Shoot Apical Meristem and Lateral Organ Formation by microRNA miR166g and Its AtHD-ZIP Target Genes. Development 2005, 132, 3657–3668. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Honda, M.; Zhu, H.; Zhang, Z.; Guo, X.; Li, T.; Li, Z.; Peng, X.; Nakajima, K.; Duan, L.; et al. Spatiotemporal Sequestration of miR165/166 by Arabidopsis Argonaute10 Promotes Shoot Apical Meristem Maintenance. Cell Rep. 2015, 10, 1819–1827. [Google Scholar] [CrossRef] [PubMed]
- Szemenyei, H.; Hannon, M.; Long, J.A. TOPLESS Mediates Auxin-Dependent Transcriptional Repression During Arabidopsis Embryogenesis. Science 2008, 319, 1380–1384. [Google Scholar] [CrossRef]
- Long, J.; Ohno, C.; Smith, Z.; Meyerowitz, E. TOPLESS Regulates Apical Embryonic Fate in Arabidopsis. Science 2006, 312, 1517–1520. [Google Scholar] [CrossRef] [PubMed]
- Plant, A.R.; Larrieu, A.; Causier, B. Repressor for Hire! The Vital Roles of TOPLESS-Mediated Transcriptional Repression in Plants. New Phytol. 2021, 231, 963–973. [Google Scholar] [CrossRef]
- McConnell, J.R.; Emery, J.; Eshed, Y.; Bao, N.; Bowman, J.; Barton, M.K. Role of PHABULOSA and PHAVOLUTA in Determining Radial Patterning in Shoots. Nature 2001, 411, 709–713. [Google Scholar] [CrossRef] [PubMed]
- Prigge, M.J.; Otsuga, D.; Alonso, J.M.; Ecker, J.R.; Drews, G.N.; Clark, S.E. Class III Homeodomain-Leucine Zipper Gene Family Members Have Overlapping, Antagonistic, and Distinct Roles in Arabidopsis Development. Plant Cell 2005, 17, 61–76. [Google Scholar] [CrossRef]
- Izhaki, A.; Bowman, J.L. KANADI and Class III HD-Zip Gene Families Regulate Embryo Patterning and Modulate Auxin Flow during Embryogenesis in Arabidopsis. Plant Cell 2007, 19, 495–508. [Google Scholar] [CrossRef]
- Chandler, J.W.; Cole, M.; Flier, A.; Grewe, B.; Werr, W. The AP2 Transcription Factors DORNRÖSCHEN and DORNRÖSCHEN-LIKE Redundantly Control Arabidopsis Embryo Patterning via Interaction with PHAVOLUTA. Development 2007, 134, 1653–1662. [Google Scholar] [CrossRef] [PubMed]
- Roodbarkelari, F.; Du, F.; Truernit, E.; Laux, T. ZLL/AGO10 Maintains Shoot Meristem Stem Cells during Arabidopsis Embryogenesis by down-Regulating ARF2-Mediated Auxin Response. BMC Biol. 2015, 13, 74. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Hu, F.; Wang, R.; Zhou, X.; Sze, S.-H.; Liou, L.W.; Barefoot, A.; Dickman, M.; Zhang, X. Arabidopsis Argonaute 10 Specifically Sequesters miR166/165 to Regulate Shoot Apical Meristem Development. Cell 2011, 145, 242–256. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tucker, E.; Hermann, M.; Laux, T. A Molecular Framework for the Embryonic Initiation of Shoot Meristem Stem Cells. Dev. Cell 2017, 40, 264–277.e4. [Google Scholar] [CrossRef] [PubMed]
- Turchi, L.; Carabelli, M.; Ruzza, V.; Possenti, M.; Sassi, M.; Peñalosa, A.; Sessa, G.; Salvi, S.; Forte, V.; Morelli, G.; et al. Arabidopsis HD-Zip II Transcription Factors Control Apical Embryo Development and Meristem Function. Development 2013, 140, 2118–2129. [Google Scholar] [CrossRef]
- Turchi, L.; Baima, S.; Morelli, G.; Ruberti, I. Interplay of HD-Zip II and III Transcription Factors in Auxin-Regulated Plant Development. J. Exp. Bot. 2015, 66, 5043–5053. [Google Scholar] [CrossRef]
- Possenti, M.; Sessa, G.; Alfè, A.; Turchi, L.; Ruzza, V.; Sassi, M.; Morelli, G.; Ruberti, I. HD-Zip II Transcription Factors Control Distal Stem Cell Fate in Arabidopsis Roots by Linking Auxin Signaling to the FEZ/SOMBRERO Pathway. Development 2024, 151, dev.202586. [Google Scholar] [CrossRef] [PubMed]
- Zheng, T.; Tan, W.; Yang, H.; Li, T.; Liu, B.; Zhang, D.; Lin, H. Regulation of Anthocyanin Accumulation via MYB75/HAT1/TPL-Mediated Transcriptional Repression. PLoS Genet. 2019, 15, e1007993. [Google Scholar] [CrossRef]
- Gallemí, M.; Molina-Contreras, M.J.; Paulišić, S.; Salla-Martret, M.; Sorin, Ç.; Godoy, M.; Franco-Zorrilla, J.M.; Solano, R.; Martínez-Garçıa, J.F. A Non-DNA-Binding Activity for the ATHB4 Transcription Factor in the Control of Vegetation Proximity. New Phytol. 2017, 216, 798–813. [Google Scholar] [CrossRef]
- De Smet, I.; Lau, S.; Ehrismann, J.S.; Axiotis, I.; Kolb, M.; Kientz, M.; Weijers, D.; Jürgens, G. Transcriptional Repression of BODENLOS by HD-ZIP Transcription Factor HB5 in Arabidopsis Thaliana. J. Exp. Bot. 2013, 64, 3009–3019. [Google Scholar] [CrossRef]
- Weijers, D.; Schlereth, A.; Ehrismann, J.S.; Schwank, G.; Kientz, M.; Jürgens, G. Auxin Triggers Transient Local Signaling for Cell Specification in Arabidopsis Embryogenesis. Dev. Cell 2006, 10, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Carlsbecker, A.; Lee, J.-Y.; Roberts, C.J.; Dettmer, J.; Lehesranta, S.; Zhou, J.; Lindgren, O.; Moreno-Risueno, M.A.; Vatén, A.; Thitamadee, S.; et al. Cell Signalling by microRNA165/6 Directs Gene Dose-Dependent Root Cell Fate. Nature 2010, 465, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Ursache, R.; Miyashima, S.; Chen, Q.; Vatén, A.; Nakajima, K.; Carlsbecker, A.; Zhao, Y.; Helariutta, Y.; Dettmer, J. Tryptophan-Dependent Auxin Biosynthesis Is Required for HD-ZIP III-Mediated Xylem Patterning. Development 2014, 141, 1250–1259. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; de Haan, M.; Mayer, J.; Janacek, D.P.; Hammes, U.Z.; Poppenberger, B.; Sieberer, T. A Novel Chemical Inhibitor of Polar Auxin Transport Promotes Shoot Regeneration by Local Enhancement of HD-ZIP III Transcription. New Phytol. 2022, 235, 1111–1128. [Google Scholar] [CrossRef] [PubMed]
- Brandt, R.; Salla-Martret, M.; Bou-Torrent, J.; Musielak, T.; Stahl, M.; Lanz, C.; Ott, F.; Schmid, M.; Greb, T.; Schwarz, M.; et al. Genome-Wide Binding-Site Analysis of REVOLUTA Reveals a Link between Leaf Patterning and Light-Mediated Growth Responses. Plant J. 2012, 72, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Harrar, Y.; Lin, C.; Reinhart, B.; Newell, N.R.; Talavera-Rauh, F.; Hokin, S.A.; Barton, M.K.; Kerstetter, R.A. Arabidopsis KANADI1 Acts as a Transcriptional Repressor by Interacting with a Specific Cis-Element and Regulates Auxin Biosynthesis, Transport, and Signaling in Opposition to HD-ZIPIII Factors. Plant Cell 2014, 26, 246–262. [Google Scholar] [CrossRef] [PubMed]
- Vatén, A.; Dettmer, J.; Wu, S.; Stierhof, Y.-D.; Miyashima, S.; Yadav, S.R.; Roberts, C.J.; Campilho, A.; Bulone, V.; Lichtenberger, R.; et al. Callose Biosynthesis Regulates Symplastic Trafficking during Root Development. Dev. Cell 2011, 21, 1144–1155. [Google Scholar] [CrossRef] [PubMed]
- Miyashima, S.; Koi, S.; Hashimoto, T.; Nakajima, K. Non-Cell-Autonomous microRNA165 Acts in a Dose-Dependent Manner to Regulate Multiple Differentiation Status in the Arabidopsis Root. Development 2011, 138, 2303–2313. [Google Scholar] [CrossRef] [PubMed]
- Bishopp, A.; Help, H.; El-Showk, S.; Weijers, D.; Scheres, B.; Friml, J.; Benková, E.; Mähönen, A.P.; Helariutta, Y. A Mutually Inhibitory Interaction between Auxin and Cytokinin Specifies Vascular Pattern in Roots. Curr. Biol. 2011, 21, 917–926. [Google Scholar] [CrossRef]
- Bishopp, A.; Lehesranta, S.; Vatén, A.; Help, H.; El-Showk, S.; Scheres, B.; Helariutta, K.; Mähönen, A.P.; Sakakibara, H.; Helariutta, Y. Phloem-Transported Cytokinin Regulates Polar Auxin Transport and Maintains Vascular Pattern in the Root Meristem. Curr. Biol. 2011, 21, 927–932. [Google Scholar] [CrossRef]
- Müller, C.J.; Valdés, A.E.; Wang, G.; Ramachandran, P.; Beste, L.; Uddenberg, D.; Carlsbecker, A. PHABULOSA Mediates an Auxin Signaling Loop to Regulate Vascular Patterning in Arabidopsis. Plant Physiol. 2016, 170, 956–970. [Google Scholar] [CrossRef]
- Baima, S.; Forte, V.; Possenti, M.; Peñalosa, A.; Leoni, G.; Salvi, S.; Felici, B.; Ruberti, I.; Morelli, G. Negative Feedback Regulation of Auxin Signaling by ATHB8/ACL5-BUD2 Transcription Module. Mol. Plant 2014, 7, 1006–1025. [Google Scholar] [CrossRef]
- el-Showk, S.; Help-Rinta-Rahko, H.; Blomster, T.; Siligato, R.; Marée, A.F.M.; Mähönen, A.P.; Grieneisen, V.A. Parsimonious Model of Vascular Patterning Links Transverse Hormone Fluxes to Lateral Root Initiation: Auxin Leads the Way, While Cytokinin Levels Out. PLoS Comput. Biol. 2015, 11, e1004450. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, P.; Wang, G.; Augstein, F.; de Vries, J.; Carlsbecker, A. Continuous Root Xylem Formation and Vascular Acclimation to Water Deficit Involves Endodermal ABA Signalling via miR165. Development 2018, 145, dev159202. [Google Scholar] [CrossRef]
- Bloch, D.; Puli, M.R.; Mosquna, A.; Yalovsky, S. Abiotic Stress Modulates Root Patterning via ABA-Regulated microRNA Expression in the Endodermis Initials. Development 2019, 146, dev177097. [Google Scholar] [CrossRef] [PubMed]
- De Rybel, B.; Adibi, M.; Breda, A.S.; Wendrich, J.R.; Smit, M.E.; Novák, O.; Yamaguchi, N.; Yoshida, S.; Van Isterdael, G.; Palovaara, J.; et al. Integration of Growth and Patterning during Vascular Tissue Formation in Arabidopsis. Science 2014, 345, 1255215. [Google Scholar] [CrossRef]
- De Rybel, B.; Möller, B.; Yoshida, S.; Grabowicz, I.; Barbier de Reuille, P.; Boeren, S.; Smith, R.S.; Borst, J.W.; Weijers, D. A bHLH Complex Controls Embryonic Vascular Tissue Establishment and Indeterminate Growth in Arabidopsis. Dev. Cell 2013, 24, 426–437. [Google Scholar] [CrossRef]
- Muraro, D.; Mellor, N.; Pound, M.P.; Help, H.; Lucas, M.; Chopard, J.; Byrne, H.M.; Godin, C.; Hodgman, T.C.; King, J.R.; et al. Integration of Hormonal Signaling Networks and Mobile microRNAs Is Required for Vascular Patterning in Arabidopsis Roots. Proc. Natl. Acad. Sci. USA 2014, 111, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Dello Ioio, R.; Galinha, C.; Fletcher, A.G.; Grigg, S.P.; Molnar, A.; Willemsen, V.; Scheres, B.; Sabatini, S.; Baulcombe, D.; Maini, P.K.; et al. A PHABULOSA/Cytokinin Feedback Loop Controls Root Growth in Arabidopsis. Curr. Biol. 2012, 22, 1699–1704. [Google Scholar] [CrossRef]
- Ohashi-Ito, K.; Iwamoto, K.; Yamagami, A.; Nakano, T.; Fukuda, H. HD-ZIP III-Dependent Local Promotion of Brassinosteroid Synthesis Suppresses Vascular Cell Division in Arabidopsis Root Apical Meristem. Proc. Natl. Acad. Sci. USA 2023, 120. [Google Scholar] [CrossRef]
- Vukašinović, N.; Wang, Y.; Vanhoutte, I.; Fendrych, M.; Guo, B.; Kvasnica, M.; Jiroutová, P.; Oklestkova, J.; Strnad, M.; Russinova, E. Local Brassinosteroid Biosynthesis Enables Optimal Root Growth. Nat. Plants 2021, 7, 619–632. [Google Scholar] [CrossRef] [PubMed]
- Miyashima, S.; Roszak, P.; Sevilem, I.; Toyokura, K.; Blob, B.; Heo, J.; Mellor, N.; Help-Rinta-Rahko, H.; Otero, S.; Smet, W.; et al. Mobile PEAR Transcription Factors Integrate Positional Cues to Prime Cambial Growth. Nature 2019, 565, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Smetana, O.; Mäkilä, R.; Lyu, M.; Amiryousefi, A.; Sánchez Rodríguez, F.; Wu, M.-F.; Solé-Gil, A.; Leal Gavarrón, M.; Siligato, R.; Miyashima, S.; et al. High Levels of Auxin Signalling Define the Stem-Cell Organizer of the Vascular Cambium. Nature 2019, 565, 485–489. [Google Scholar] [CrossRef]
- He, G.; Liu, P.; Zhao, H.; Sun, J. The HD-ZIP II Transcription Factors Regulate Plant Architecture through the Auxin Pathway. Int. J. Mol. Sci. 2020, 21, 3250. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.T.; Xiang, Z.X.; Li, W.; Gao, X.; Lu, Y.T. Osmotic Stress Represses Root Growth by Modulating the Transcriptional Regulation of PIN-FORMED3. New Phytol. 2021, 232, 1661–1673. [Google Scholar] [CrossRef] [PubMed]
- Sawa, S.; Ohgishi, M.; Goda, H.; Higuchi, K.; Shimada, Y.; Yoshida, S.; Koshiba, T. The HAT2 Gene, a Member of the HD-Zip Gene Family, Isolated as an Auxin Inducible Gene by DNA Microarray Screening, Affects Auxin Response in Arabidopsis. Plant J. 2002, 32, 1011–1022. [Google Scholar] [CrossRef]
- Ruzza, V.; Sessa, G.; Sassi, M.; Morelli, G.; Ruberti, I. Auxin Coordinates Shoot and Root Development During Shade Avoidance Response. In Auxin and Its Role in Plant Development; Zažímalová, E., Petrášek, J., Benková, E., Eds.; Springer: Vienna, Austria, 2014; pp. 389–412. ISBN 978-3-7091-1525-1. [Google Scholar]
- Sassi, M.; Lu, Y.; Zhang, Y.; Wang, J.; Dhonukshe, P.; Blilou, I.; Dai, M.; Li, J.; Gong, X.; Jaillais, Y.; et al. COP1 Mediates the Coordination of Root and Shoot Growth by Light through Modulation of PIN1- and PIN2-Dependent Auxin Transport in Arabidopsis. Development 2012, 139, 3402–3412. [Google Scholar] [CrossRef]
- Steindler, C.; Matteucci, A.; Sessa, G.; Weimar, T.; Ohgishi, M.; Aoyama, T.; Morelli, G.; Ruberti, I. Shade Avoidance Responses Are Mediated by the ATHB-2 HD-Zip Protein, a Negative Regulator of Gene Expression. Development 1999, 126, 4235–4245. [Google Scholar] [CrossRef] [PubMed]
- Merelo, P.; Ram, H.; Pia Caggiano, M.; Ohno, C.; Ott, F.; Straub, D.; Graeff, M.; Cho, S.K.; Yang, S.W.; Wenkel, S.; et al. Regulation of MIR165/166 by Class II and Class III Homeodomain Leucine Zipper Proteins Establishes Leaf Polarity. Proc. Natl. Acad. Sci. USA 2016, 113, 11973–11978. [Google Scholar] [CrossRef]
- Tognacca, R.S.; Carabelli, M.; Morelli, G.; Ruberti, I.; Botto, J.F. ATHB2 Is a Negative Regulator of Germination in Arabidopsis Thaliana Seeds. Sci. Rep. 2021, 11, 9688. [Google Scholar] [CrossRef]
- Sorin, C.; Salla-Martret, M.; Bou-Torrent, J.; Roig-Villanova, I.; Martínez-García, J.F. ATHB4, a Regulator of Shade Avoidance, Modulates Hormone Response in Arabidopsis Seedlings. Plant J. 2009, 59, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Köllmer, I.; Werner, T.; Schmülling, T. Ectopic Expression of Different Cytokinin-Regulated Transcription Factor Genes of Arabidopsis Thaliana Alters Plant Growth and Development. J. Plant Physiol. 2011, 168, 1320–1327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Ye, H.; Guo, H.; Johnson, A.; Zhang, M.; Lin, H.; Yin, Y. Transcription Factor HAT1 Is Phosphorylated by BIN2 Kinase and Mediates Brassinosteroid Repressed Gene Expression in Arabidopsis. Plant J. 2014, 77, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Zhang, D.; Zhou, H.; Zheng, T.; Yin, Y.; Lin, H. Transcription Factor HAT1 Is a Substrate of SnRK2.3 Kinase and Negatively Regulates ABA Synthesis and Signaling in Arabidopsis Responding to Drought. PLoS Genet. 2018, 14, e1007336. [Google Scholar] [CrossRef] [PubMed]
- Nolan, T.M.; Vukašinović, N.; Hsu, C.-W.; Zhang, J.; Vanhoutte, I.; Shahan, R.; Taylor, I.W.; Greenstreet, L.; Heitz, M.; Afanassiev, A.; et al. Brassinosteroid Gene Regulatory Networks at Cellular Resolution in the Arabidopsis Root. Science 2023, 379, eadf4721. [Google Scholar] [CrossRef] [PubMed]
- Hur, Y.-S.; Um, J.-H.; Kim, S.; Kim, K.; Park, H.-J.; Lim, J.-S.; Kim, W.-Y.; Jun, S.E.; Yoon, E.K.; Lim, J.; et al. Arabidopsis Thaliana Homeobox 12 (ATHB12), a Homeodomain-Leucine Zipper Protein, Regulates Leaf Growth by Promoting Cell Expansion and Endoreduplication. New Phytol. 2015, 205, 316–328. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, J.; Guo, S.; Yuan, X.; Zhao, S.; Tian, H.; Dai, S.; Kong, X.; Ding, Z. AtHB7/12 Regulate Root Growth in Response to Aluminum Stress. Int. J. Mol. Sci. 2020, 21, 4080. [Google Scholar] [CrossRef] [PubMed]
- Perotti, M.F.; Ribone, P.A.; Cabello, J.V.; Ariel, F.D.; Chan, R.L. AtHB23 Participates in the Gene Regulatory Network Controlling Root Branching, and Reveals Differences between Secondary and Tertiary Roots. Plant J. 2019, 100, 1224–1236. [Google Scholar] [CrossRef]
- Perotti, M.F.; Arce, A.L.; Ariel, F.D.; Figueroa, C.M.; Chan, R.L. The Transcription Factor AtHB23 Modulates Starch Turnover for Root Development and Plant Survival under Salinity. Environ. Exp. Bot. 2022, 201, 104994. [Google Scholar] [CrossRef]
- Mora, C.C.; Perotti, M.F.; González-Grandío, E.; Ribone, P.A.; Cubas, P.; Chan, R.L. AtHB40 Modulates Primary Root Length and Gravitropism Involving CYCLINB and Auxin Transporters. Plant Sci. 2022, 324, 111421. [Google Scholar] [CrossRef]
- Son, O.; Cho, H.-Y.; Kim, M.-R.; Lee, H.; Lee, M.-S.; Song, E.; Park, J.H.; Nam, K.H.; Chun, J.-Y.; Kim, H.-J.; et al. Induction of a Homeodomain–Leucine Zipper Gene by Auxin Is Inhibited by Cytokinin in Arabidopsis Roots. Biochem. Biophys. Res. Commun. 2004, 326, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.-Q.; Zhao, P.-X.; Mao, J.-L.; Yu, L.-H.; Yuan, Y.; Tang, H.; Liu, Z.-B.; Xiang, C.-B. HOMEOBOX PROTEIN52 Mediates the Crosstalk between Ethylene and Auxin Signaling during Primary Root Elongation by Modulating Auxin Transport-Related Gene Expression. Plant Cell 2018, 30, 2761–2778. [Google Scholar] [CrossRef] [PubMed]
- Masucci, J.D.; Rerie, W.G.; Foreman, D.R.; Zhang, M.; Galway, M.E.; Marks, M.D.; Schiefelbein, J.W. The Homeobox Gene GLABRA 2 Is Required for Position-Dependent Cell Differentiation in the Root Epidermis of Arabidopsis Thaliana. Development 1996, 122, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Di Cristina, M.; Sessa, G.; Dolan, L.; Linstead, P.; Baima, S.; Ruberti, I.; Morelli, G. The Arabidopsis Athb-10 (GLABRA2) Is an HD-Zip Protein Required for Regulation of Root Hair Development. Plant J. 1996, 10, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, Y.; Oka, A.; Rodrigues-Pousada, R.; Possenti, M.; Ruberti, I.; Morelli, G.; Aoyama, T. Modulation of Phospholipid Signaling by GLABRA2 in Root-Hair Pattern Formation. Science 2003, 300, 1427–1430. [Google Scholar] [CrossRef] [PubMed]
- Kuppusamy, K.T.; Chen, A.Y.; Nemhauser, J.L. Steroids Are Required for Epidermal Cell Fate Establishment in Arabidopsis Roots. Proc. Natl. Acad. Sci. USA 2009, 106, 8073–8076. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhu, W.; Chen, Y.; Ito, S.; Asami, T.; Wang, X. Brassinosteroids Control Root Epidermal Cell Fate via Direct Regulation of a MYB-bHLH-WD40 Complex by GSK3-like Kinases. eLife 2014, 3, e02525. [Google Scholar] [CrossRef] [PubMed]
- Borassi, C.; Gloazzo Dorosz, J.; Ricardi, M.M.; Carignani Sardoy, M.; Pol Fachin, L.; Marzol, E.; Mangano, S.; Rodríguez Garcia, D.R.; Martínez Pacheco, J.; Rondón Guerrero, Y.d.C.; et al. A Cell Surface Arabinogalactan-Peptide Influences Root Hair Cell Fate. New Phytol. 2020, 227, 732–743. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Tao, R.; Feng, Y.; Xiao, Z.; Zhang, D.; Peng, Y.; Wen, X.; Wang, Y.; Guo, H. EIN3 and RSL4 Interfere with an MYB–bHLH–WD40 Complex to Mediate Ethylene-Induced Ectopic Root Hair Formation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2021, 118, e2110004118. [Google Scholar] [CrossRef]
- Fridman, Y.; Elkouby, L.; Holland, N.; Vragović, K.; Elbaum, R.; Savaldi-Goldstein, S. Root Growth Is Modulated by Differential Hormonal Sensitivity in Neighboring Cells. Genes Dev. 2014, 28, 912–920. [Google Scholar] [CrossRef]
- Cai, X.-T.; Xu, P.; Wang, Y.; Xiang, C.-B. Activated Expression of AtEDT1/HDG11 Promotes Lateral Root Formation in Arabidopsis Mutant Edt1 by Upregulating Jasmonate Biosynthesis. J. Integr. Plant Biol. 2015, 57, 1017–1030. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Cai, X.-T.; Wang, Y.; Xing, L.; Chen, Q.; Xiang, C.-B. HDG11 Upregulates Cell-Wall-Loosening Protein Genes to Promote Root Elongation in Arabidopsis. J. Exp. Bot. 2014, 65, 4285–4295. [Google Scholar] [CrossRef] [PubMed]
- Surina, S.; Yamagami, A.; Miyaji, T.; Chagan, Z.; Chung, K.M.; Mitsuda, N.; Nishida, K.; Tachibana, R.; Zhu, Z.; Miyakawa, T.; et al. BIL9 Promotes Both Plant Growth via BR Signaling and Drought Stress Resistance by Binding with the Transcription Factor HDG11. Plant Cell Physiol. 2024, pcae009. [Google Scholar] [CrossRef] [PubMed]
- Rombolá-Caldentey, B.; Rueda-Romero, P.; Iglesias-Fernández, R.; Carbonero, P.; Oñate-Sánchez, L. Arabidopsis DELLA and Two HD-ZIP Transcription Factors Regulate GA Signaling in the Epidermis through the L1 Box Cis -Element. Plant Cell 2014, 26, 2905–2919. [Google Scholar] [CrossRef] [PubMed]
- Green, K.A.; Prigge, M.J.; Katzman, R.B.; Clark, S.E. CORONA, a Member of the Class III Homeodomain Leucine Zipper Gene Family in Arabidopsis, Regulates Stem Cell Specification and Organogenesis. Plant Cell 2005, 17, 691–704. [Google Scholar] [CrossRef] [PubMed]
- Mandel, T.; Candela, H.; Landau, U.; Asis, L.; Zelinger, E.; Carles, C.C.; Williams, L.E. Differential Regulation of Meristem Size, Morphology and Organization by the ERECTA, CLAVATA and Class III HD-ZIP Pathways. Development 2016, 143, 1612–1622. [Google Scholar] [CrossRef] [PubMed]
- Grigg, S.P.; Canales, C.; Hay, A.; Tsiantis, M. SERRATE Coordinates Shoot Meristem Function and Leaf Axial Patterning in Arabidopsis. Nature 2005, 437, 1022–1026. [Google Scholar] [CrossRef] [PubMed]
- Sassi, M.; Vernoux, T. Auxin and Self-Organization at the Shoot Apical Meristem. J. Exp. Bot. 2013, 64, 2579–2592. [Google Scholar] [CrossRef]
- Heisler, M.G. Integration of Core Mechanisms Underlying Plant Aerial Architecture. Front. Plant Sci. 2021, 12, 786338. [Google Scholar] [CrossRef]
- Reinhardt, D.; Pesce, E.-R.; Stieger, P.; Mandel, T.; Baltensperger, K.; Bennett, M.; Traas, J.; Friml, J.; Kuhlemeier, C. Regulation of Phyllotaxis by Polar Auxin Transport. Nature 2003, 426, 255–260. [Google Scholar] [CrossRef]
- Heisler, M.G.; Ohno, C.; Das, P.; Sieber, P.; Reddy, G.V.; Long, J.A.; Meyerowitz, E.M. Patterns of Auxin Transport and Gene Expression during Primordium Development Revealed by Live Imaging of the Arabidopsis Inflorescence Meristem. Curr. Biol. 2005, 15, 1899–1911. [Google Scholar] [CrossRef] [PubMed]
- Sassi, M.; Ali, O.; Boudon, F.; Cloarec, G.; Abad, U.; Cellier, C.; Chen, X.; Gilles, B.; Milani, P.; Friml, J.; et al. An Auxin-Mediated Shift toward Growth Isotropy Promotes Organ Formation at the Shoot Meristem in Arabidopsis. Curr. Biol. 2014, 24, 2335–2342. [Google Scholar] [CrossRef]
- Caggiano, M.P.; Yu, X.; Bhatia, N.; Larsson, A.; Ram, H.; Ohno, C.K.; Sappl, P.; Meyerowitz, E.M.; Jönsson, H.; Heisler, M.G. Cell Type Boundaries Organize Plant Development. eLife 2017, 6, e27421. [Google Scholar] [CrossRef] [PubMed]
- Ram, H.; Sahadevan, S.; Gale, N.; Caggiano, M.P.; Yu, X.; Ohno, C.; Heisler, M.G. An Integrated Analysis of Cell-Type Specific Gene Expression Reveals Genes Regulated by REVOLUTA and KANADI1 in the Arabidopsis Shoot Apical Meristem. PLoS Genet. 2020, 16, e1008661. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.-Q.; Lian, H.; Zhou, C.-M.; Xu, L.; Jiao, Y.; Wang, J.-W. A Two-Step Model for de Novo Activation of WUSCHEL during Plant Shoot Regeneration. Plant Cell 2017, 29, 1073–1087. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.; Dai, X.; Wang, R.; Wang, J.; Liu, Z.; Xiang, F. ARGONAUTE10 Inhibits In Vitro Shoot Regeneration Via Repression of miR165/166 in Arabidopsis Thaliana. Plant Cell Physiol. 2017, 58, 1789–1800. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Sasaki, Y.; Hashimoto, K.; Nakajima, K.; Gasser, C.S. CORONA, PHABULOSA and PHAVOLUTA Collaborate with BELL1 to Confine WUSCHEL Expression to the Nucellus in Arabidopsis Ovules. Development 2016, 143, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Abad Vivero, U.C. Morphogenesis at the Shoot Apical Meristem. Ph.D. Thesis, Université de Lyon, Lyon, France, 2017. Available online: https://theses.hal.science/tel-01948883 (accessed on 22 May 2024).
- Nobusawa, T.; Okushima, Y.; Nagata, N.; Kojima, M.; Sakakibara, H.; Umeda, M. Synthesis of Very-Long-Chain Fatty Acids in the Epidermis Controls Plant Organ Growth by Restricting Cell Proliferation. PLoS Biol. 2013, 11, e1001531. [Google Scholar] [CrossRef] [PubMed]
- Roudier, F.; Gissot, L.; Beaudoin, F.; Haslam, R.; Michaelson, L.; Marion, J.; Molino, D.; Lima, A.; Bach, L.; Morin, H.; et al. Very-Long-Chain Fatty Acids Are Involved in Polar Auxin Transport and Developmental Patterning in Arabidopsis. Plant Cell 2010, 22, 364–375. [Google Scholar] [CrossRef]
- Wang, H.; Lu, Z.; Xu, Y.; Zhang, J.; Han, L.; Chai, M.; Wang, Z.-Y.; Yang, X.; Lu, S.; Tong, J.; et al. Roles of Very Long-Chain Fatty Acids in Compound Leaf Patterning in Medicago Truncatula. Plant Physiol. 2023, 191, 1751–1770. [Google Scholar] [CrossRef]
- Reinhardt, D.; Frenz, M.; Mandel, T.; Kuhlemeier, C. Microsurgical and Laser Ablation Analysis of Interactions between the Zones and Layers of the Tomato Shoot Apical Meristem. Development 2003, 130, 4073–4083. [Google Scholar] [CrossRef] [PubMed]
- Savaldi-Goldstein, S.; Peto, C.; Chory, J. The Epidermis Both Drives and Restricts Plant Shoot Growth. Nature 2007, 446, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Heisler, M.G.; Byrne, M.E. Progress in Understanding the Role of Auxin in Lateral Organ Development in Plants. Curr. Opin. Plant Biol. 2020, 53, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Mallory, A.C.; Reinhart, B.J.; Jones-Rhoades, M.W.; Tang, G.; Zamore, P.D.; Barton, M.K.; Bartel, D.P. MicroRNA Control of PHABULOSA in Leaf Development: Importance of Pairing to the microRNA 5′ Region. EMBO J. 2004, 23, 3356–3364. [Google Scholar] [CrossRef]
- Garcia, D.; Collier, S.A.; Byrne, M.E.; Martienssen, R.A. Specification of Leaf Polarity in Arabidopsis via the Trans-Acting siRNA Pathway. Curr. Biol. 2006, 16, 933–938. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.R.; Arreola, A.; Gallagher, T.L.; Gasser, C.S. ETTIN (ARF3) Physically Interacts with KANADI Proteins to Form a Functional Complex Essential for Integument Development and Polarity Determination in Arabidopsis. Development 2012, 139, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Chitwood, D.H.; Nogueira, F.T.S.; Howell, M.D.; Montgomery, T.A.; Carrington, J.C.; Timmermans, M.C.P. Pattern Formation via Small RNA Mobility. Genes Dev. 2009, 23, 549–554. [Google Scholar] [CrossRef]
- Reinhart, B.J.; Liu, T.; Newell, N.R.; Magnani, E.; Huang, T.; Kerstetter, R.; Michaels, S.; Barton, M.K. Establishing a Framework for the Ad/Abaxial Regulatory Network of Arabidopsis: Ascertaining Targets of Class III Homeodomain Leucine Zipper and KANADI Regulation. Plant Cell 2013, 25, 3228–3249. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zhao, C.; Zhou, J.; Yang, Y.; Wang, P.; Zhu, X.; Tang, G.; Bressan, R.A.; Zhu, J.-K. The miR165/166 Mediated Regulatory Module Plays Critical Roles in ABA Homeostasis and Response in Arabidopsis Thaliana. PLoS Genet. 2016, 12, e1006416. [Google Scholar] [CrossRef]
- Liu, T.; Longhurst, A.D.; Talavera-Rauh, F.; Hokin, S.A.; Barton, M.K. The Arabidopsis Transcription Factor ABIG1 Relays ABA Signaled Growth Inhibition and Drought Induced Senescence. eLife 2016, 5, e13768. [Google Scholar] [CrossRef]
- Preciado, J.; Begcy, K.; Liu, T. The Arabidopsis HDZIP Class II Transcription Factor ABA INSENSITIVE TO GROWTH 1 Functions in Leaf Development. J. Exp. Bot. 2022, 73, 1978–1991. [Google Scholar] [CrossRef]
- Hasegawa, R.; Arakawa, T.; Fujita, K.; Tanaka, Y.; Ookawa, Z.; Sakamoto, S.; Takasaki, H.; Ikeda, M.; Yamagami, A.; Mitsuda, N.; et al. Arabidopsis Homeobox-Leucine Zipper Transcription Factor BRASSINOSTEROID-RELATED HOMEOBOX 3 Regulates Leaf Greenness by Suppressing BR Signaling. Plant Biotechnol. 2022, 39, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Han, Q.; Li, Y.; Yang, F.; Li, J.; Li, P.; Xu, X.; Lin, H.; Zhang, D. A HAT1-DELLA Signaling Module Regulates Trichome Initiation and Leaf Growth by Achieving Gibberellin Homeostasis. New Phytol. 2021, 231, 1220–1235. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; An, L.; Sun, L.; Zhu, S.; Xi, W.; Broun, P.; Yu, H.; Gan, Y. Zinc Finger Protein5 Is Required for the Control of Trichome Initiation by Acting Upstream of Zinc Finger Protein8 in Arabidopsis. Plant Physiol. 2011, 157, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Sun, L.; Zhao, Y.; An, L.; Yan, A.; Meng, X.; Gan, Y. Zinc Finger Protein 6 (ZFP6) Regulates Trichome Initiation by Integrating Gibberellin and Cytokinin Signaling in Arabidopsis Thaliana. New Phytol. 2013, 198, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Donner, T.J.; Sherr, I.; Scarpella, E. Regulation of Preprocambial Cell State Acquisition by Auxin Signaling in Arabidopsis Leaves. Development 2009, 136, 3235–3246. [Google Scholar] [CrossRef] [PubMed]
- Krishna, A.; Gardiner, J.; Donner, T.J.; Scarpella, E. Control of Vein-Forming, Striped Gene Expression by Auxin Signaling. BMC Biol. 2021, 19, 213. [Google Scholar] [CrossRef] [PubMed]
- Baima, S.; Nobili, F.; Sessa, G.; Lucchetti, S.; Ruberti, I.; Morelli, G. The Expression of the Athb-8 Homeobox Gene Is Restricted to Provascular Cells in Arabidopsis Thaliana. Development 1995, 121, 4171–4182. [Google Scholar] [CrossRef] [PubMed]
- Carabelli, M.; Possenti, M.; Sessa, G.; Ruzza, V.; Morelli, G.; Ruberti, I. Arabidopsis HD-Zip II Proteins Regulate the Exit from Proliferation during Leaf Development in Canopy Shade. J. Exp. Bot. 2018, 69, 5419–5431. [Google Scholar] [CrossRef]
- Challa, K.R.; Rath, M.; Nath, U. The CIN-TCP Transcription Factors Promote Commitment to Differentiation in Arabidopsis Leaf Pavement Cells via Both Auxin-Dependent and Independent Pathways. PLoS Genet. 2019, 15, e1007988. [Google Scholar] [CrossRef]
- Hur, Y.-S.; Kim, J.; Kim, S.; Son, O.; Kim, W.-Y.; Kim, G.-T.; Ohme-Takagi, M.; Cheon, C.-I. Identification of TCP13 as an Upstream Regulator of ATHB12 during Leaf Development. Genes 2019, 10, 644. [Google Scholar] [CrossRef] [PubMed]
- Son, O.; Hur, Y.-S.; Kim, Y.-K.; Lee, H.-J.; Kim, S.; Kim, M.-R.; Nam, K.H.; Lee, M.-S.; Kim, B.-Y.; Park, J.; et al. ATHB12, an ABA-Inducible Homeodomain-Leucine Zipper (HD-Zip) Protein of Arabidopsis, Negatively Regulates the Growth of the Inflorescence Stem by Decreasing the Expression of a Gibberellin 20-Oxidase Gene. Plant Cell Physiol. 2010, 51, 1537–1547. [Google Scholar] [CrossRef] [PubMed]
- Vuolo, F.; Kierzkowski, D.; Runions, A.; Hajheidari, M.; Mentink, R.A.; Gupta, M.D.; Zhang, Z.; Vlad, D.; Wang, Y.; Pecinka, A.; et al. LMI1 Homeodomain Protein Regulates Organ Proportions by Spatial Modulation of Endoreduplication. Genes Dev. 2018, 32, 1361–1366. [Google Scholar] [CrossRef]
- Miguel, V.N.; Manavella, P.A.; Chan, R.L.; Capella, M.A. The AtHB1 Transcription Factor Controls the miR164-CUC2 Regulatory Node to Modulate Leaf Development. Plant Cell Physiol. 2020, 61, 659–670. [Google Scholar] [CrossRef]
- Bilsborough, G.D.; Runions, A.; Barkoulas, M.; Jenkins, H.W.; Hasson, A.; Galinha, C.; Laufs, P.; Hay, A.; Prusinkiewicz, P.; Tsiantis, M. Model for the Regulation of Arabidopsis Thaliana Leaf Margin Development. Proc. Natl. Acad. Sci. USA 2011, 108, 3424–3429. [Google Scholar] [CrossRef] [PubMed]
- Torii, K.U. Stomatal Development in the Context of Epidermal Tissues. Ann. Bot. 2021, 128, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Peterson, K.M.; Shyu, C.; Burr, C.A.; Horst, R.J.; Kanaoka, M.M.; Omae, M.; Sato, Y.; Torii, K.U. Arabidopsis Homeodomain-Leucine Zipper IV Proteins Promote Stomatal Development and Ectopically Induce Stomata beyond the Epidermis. Development 2013, 140, 1924–1935. [Google Scholar] [CrossRef] [PubMed]
- Le, J.; Liu, X.-G.; Yang, K.-Z.; Chen, X.-L.; Zou, J.-J.; Wang, H.-Z.; Wang, M.; Vanneste, S.; Morita, M.; Tasaka, M.; et al. Auxin Transport and Activity Regulate Stomatal Patterning and Development. Nat. Commun. 2014, 5, 3090. [Google Scholar] [CrossRef]
- de Marcos, A.; Houbaert, A.; Triviño, M.; Delgado, D.; Martín-Trillo, M.; Russinova, E.; Fenoll, C.; Mena, M. A Mutation in the bHLH Domain of the SPCH Transcription Factor Uncovers a BR-Dependent Mechanism for Stomatal Development. Plant Physiol. 2017, 174, 823–842. [Google Scholar] [CrossRef]
- Mohamed, D.; Vonapartis, E.; Corcega, D.Y.; Gazzarrini, S. ABA Guides Stomatal Proliferation and Patterning through the EPF-SPCH Signaling Pathway in Arabidopsis Thaliana. Development 2023, 150, dev201258. [Google Scholar] [CrossRef]
- Abad, U.; Sassi, M.; Traas, J. Flower Development: From Morphodynamics to Morphomechanics. Phil Trans. R. Soc. B 2017, 372, 20150545. [Google Scholar] [CrossRef] [PubMed]
- Denay, G.; Chahtane, H.; Tichtinsky, G.; Parcy, F. A Flower Is Born: An Update on Arabidopsis Floral Meristem Formation. Curr. Opin. Plant Biol. 2017, 35, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Moubayidin, L.; Østergaard, L. Dynamic Control of Auxin Distribution Imposes a Bilateral-to-Radial Symmetry Switch during Gynoecium Development. Curr. Biol. 2014, 24, 2743–2748. [Google Scholar] [CrossRef] [PubMed]
- Schuster, C.; Gaillochet, C.; Lohmann, J.U. Arabidopsis HECATE Genes Function in Phytohormone Control during Gynoecium Development. Development 2015, 142, 3343–3350. [Google Scholar] [CrossRef] [PubMed]
- Carabelli, M.; Turchi, L.; Morelli, G.; Østergaard, L.; Ruberti, I.; Moubayidin, L. Coordination of Biradial-to-Radial Symmetry and Tissue Polarity by HD-ZIP II Proteins. Nat. Commun. 2021, 12, 4321. [Google Scholar] [CrossRef] [PubMed]
- Gaillochet, C.; Stiehl, T.; Wenzl, C.; Ripoll, J.-J.; Bailey-Steinitz, L.J.; Li, L.; Pfeiffer, A.; Miotk, A.; Hakenjos, J.P.; Forner, J.; et al. Control of Plant Cell Fate Transitions by Transcriptional and Hormonal Signals. eLife 2017, 6, e30135. [Google Scholar] [CrossRef] [PubMed]
- Zúñiga-Mayo, V.M.; Marsch-Martínez, N.; de Folter, S. JAIBA, a Class-II HD-ZIP Transcription Factor Involved in the Regulation of Meristematic Activity, and Important for Correct Gynoecium and Fruit Development in Arabidopsis. Plant J. 2012, 71, 314–326. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Ubaldo, H.; Campos, S.E.; López-Gómez, P.; Luna-García, V.; Zúñiga-Mayo, V.M.; Armas-Caballero, G.E.; González-Aguilera, K.L.; DeLuna, A.; Marsch-Martínez, N.; Espinosa-Soto, C.; et al. The Protein–Protein Interaction Landscape of Transcription Factors during Gynoecium Development in Arabidopsis. Mol. Plant 2023, 16, 260–278. [Google Scholar] [CrossRef] [PubMed]
- Nole-Wilson, S.; Azhakanandam, S.; Franks, R.G. Polar Auxin Transport Together with AINTEGUMENTA and REVOLUTA Coordinate Early Arabidopsis Gynoecium Development. Dev. Biol. 2010, 346, 181–195. [Google Scholar] [CrossRef]
- Denay, G.; Gabrielle, T.; Marie, L.M.; Hicham, C.; Sylvie, H.; Irene, L.-V.; Christian, W.; Manuel, F.-Z.J.; Rüdiger, S.; Lohmann, J.U.; et al. Control of Stem-Cell Niche Establishment in Arabidopsis Flowers by REVOLUTA and the LEAFY-RAX1 Module. BioRxiv 2018. [Google Scholar] [CrossRef]
- Hashimoto, K.; Miyashima, S.; Sato-Nara, K.; Yamada, T.; Nakajima, K. Functionally Diversified Members of the MIR165/6 Gene Family Regulate Ovule Morphogenesis in Arabidopsis Thaliana. Plant Cell Physiol. 2018, 59, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, J.; Wenkel, S.; Chandler, J.W.; Werr, W.; Jiao, Y. Spatiotemporal Control of Axillary Meristem Formation by Interacting Transcriptional Regulators. Development 2018, 145, dev158352. [Google Scholar] [CrossRef] [PubMed]
- Kamata, N.; Okada, H.; Komeda, Y.; Takahashi, T. Mutations in Epidermis-Specific HD-ZIP IV Genes Affect Floral Organ Identity in Arabidopsis Thaliana. Plant J. 2013, 75, 430–440. [Google Scholar] [CrossRef]
- Vadde, B.V.L.; Russell, N.J.; Bagde, S.R.; Askey, B.; Saint-Antoine, M.; Brownfield, B.; Mughal, S.; Apprill, L.E.; Khosla, A.; Clark, F.K.; et al. The Transcription Factor ATML1 Maintains Giant Cell Identity by Inducing Synthesis of Its Own Long-Chain Fatty Acid-Containing Ligands. BioRxiv 2024. [Google Scholar] [CrossRef]
- González-Grandío, E.; Pajoro, A.; Franco-Zorrilla, J.M.; Tarancón, C.; Immink, R.G.H.; Cubas, P. Abscisic Acid Signaling Is Controlled by a BRANCHED1/HD-ZIP I Cascade in Arabidopsis Axillary Buds. Proc. Natl. Acad. Sci. USA 2017, 114, E245–E254. [Google Scholar] [CrossRef]
- Dong, S.; Tarkowska, D.; Sedaghatmehr, M.; Welsch, M.; Gupta, S.; Mueller-Roeber, B.; Balazadeh, S. The HB40-JUB1 Transcriptional Regulatory Network Controls Gibberellin Homeostasis in Arabidopsis. Mol. Plant 2022, 15, 322–339. [Google Scholar] [CrossRef]
- Saddic, L.A.; Huvermann, B.; Bezhani, S.; Su, Y.; Winter, C.M.; Kwon, C.S.; Collum, R.P.; Wagner, D. The LEAFY Target LMI1 Is a Meristem Identity Regulator and Acts Together with LEAFY to Regulate Expression of CAULIFLOWER. Development 2006, 133, 1673–1682. [Google Scholar] [CrossRef]
- Takatsuka, H.; Umeda, M. Hormonal Control of Cell Division and Elongation along Differentiation Trajectories in Roots. J. Exp. Bot. 2014, 65, 2633–2643. [Google Scholar] [CrossRef]
- Ruberti, I.; Sessa, G.; Ciolfi, A.; Possenti, M.; Carabelli, M.; Morelli, G. Plant Adaptation to Dynamically Changing Environment: The Shade Avoidance Response. Biotechnol. Adv. 2012, 30, 1047–1058. [Google Scholar] [CrossRef]
- Perotti, M.F.; Ribone, P.A.; Chan, R.L. Plant Transcription Factors from the Homeodomain-Leucine Zipper Family I. Role in Development and Stress Responses. IUBMB Life 2017, 69, 280–289. [Google Scholar] [CrossRef]
- Wei, H.; Song, Z.; Xie, Y.; Cheng, H.; Yan, H.; Sun, F.; Liu, H.; Shen, J.; Li, L.; He, X.; et al. High Temperature Inhibits Vascular Development via the PIF4-miR166-HB15 Module in Arabidopsis. Curr. Biol. 2023, 33, 3203–3214.e4. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sessa, G.; Carabelli, M.; Sassi, M. The Ins and Outs of Homeodomain-Leucine Zipper/Hormone Networks in the Regulation of Plant Development. Int. J. Mol. Sci. 2024, 25, 5657. https://doi.org/10.3390/ijms25115657
Sessa G, Carabelli M, Sassi M. The Ins and Outs of Homeodomain-Leucine Zipper/Hormone Networks in the Regulation of Plant Development. International Journal of Molecular Sciences. 2024; 25(11):5657. https://doi.org/10.3390/ijms25115657
Chicago/Turabian StyleSessa, Giovanna, Monica Carabelli, and Massimiliano Sassi. 2024. "The Ins and Outs of Homeodomain-Leucine Zipper/Hormone Networks in the Regulation of Plant Development" International Journal of Molecular Sciences 25, no. 11: 5657. https://doi.org/10.3390/ijms25115657
APA StyleSessa, G., Carabelli, M., & Sassi, M. (2024). The Ins and Outs of Homeodomain-Leucine Zipper/Hormone Networks in the Regulation of Plant Development. International Journal of Molecular Sciences, 25(11), 5657. https://doi.org/10.3390/ijms25115657