
Apigenin: Molecular Mechanisms and Therapeutic Potential against Cancer Spreading




Abstract
1. Introduction
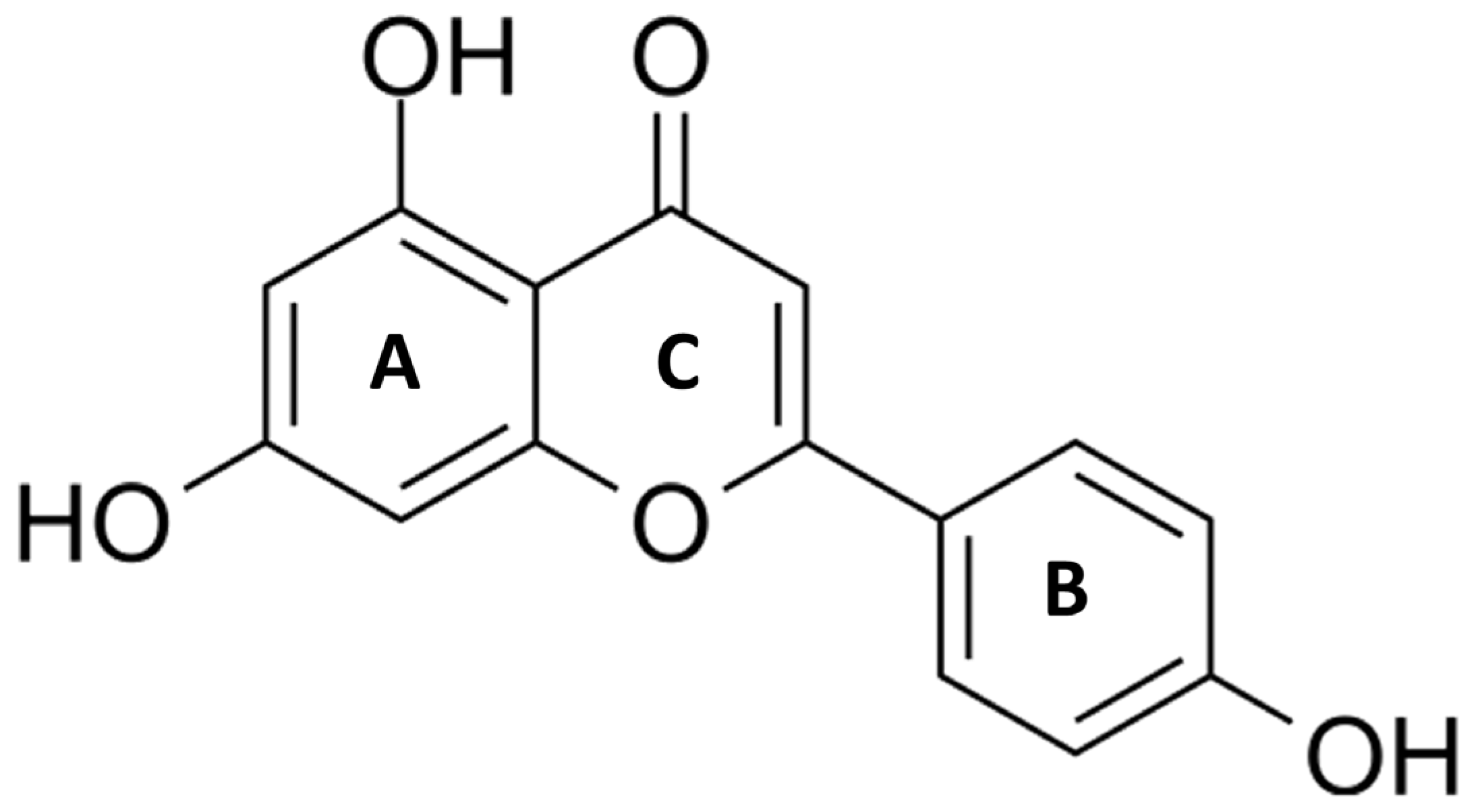
2. Apigenin
3. Apigenin and Angiogenesis
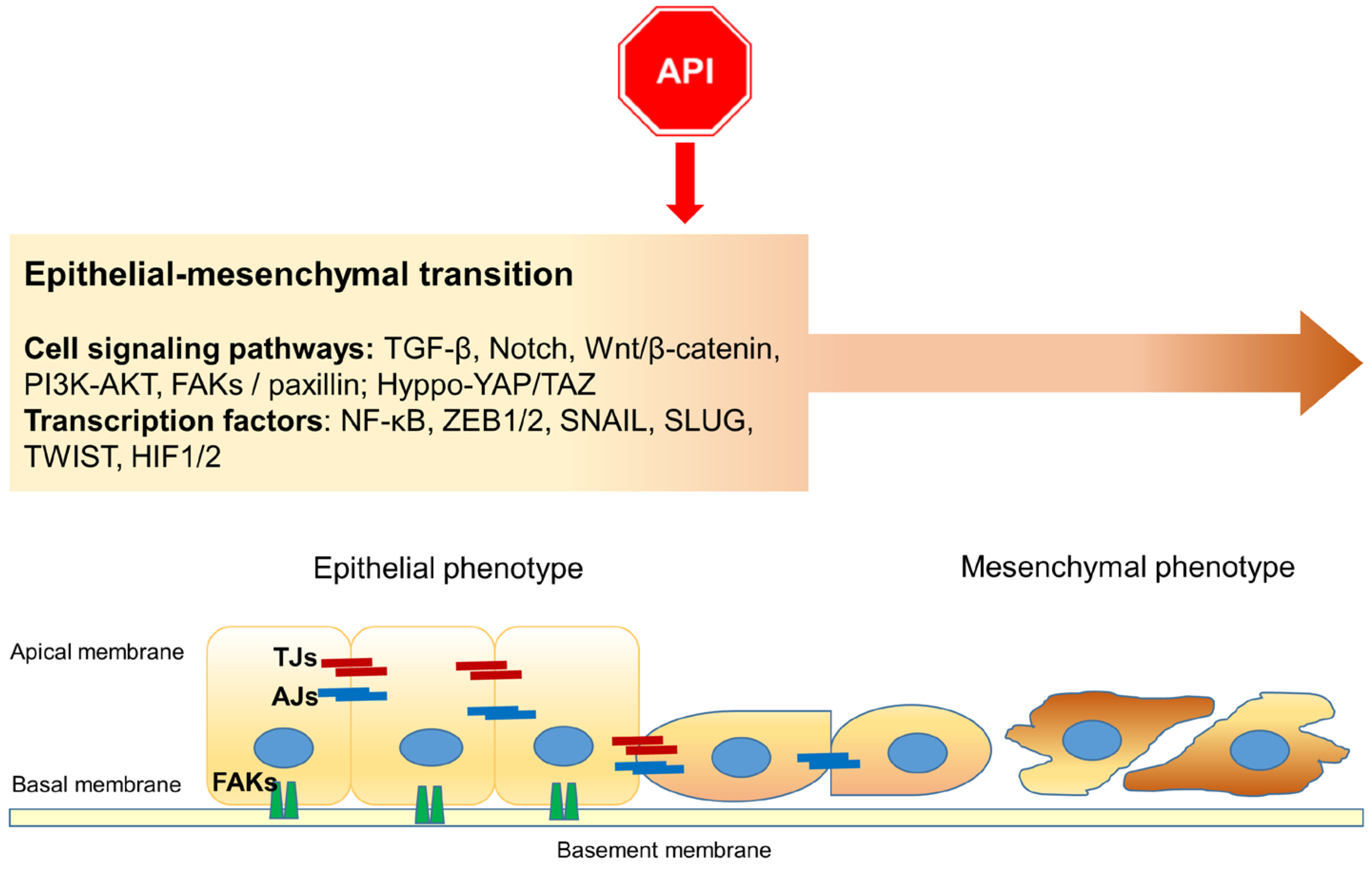
4. Apigenin and Epithelial–Mesenchymal Transition
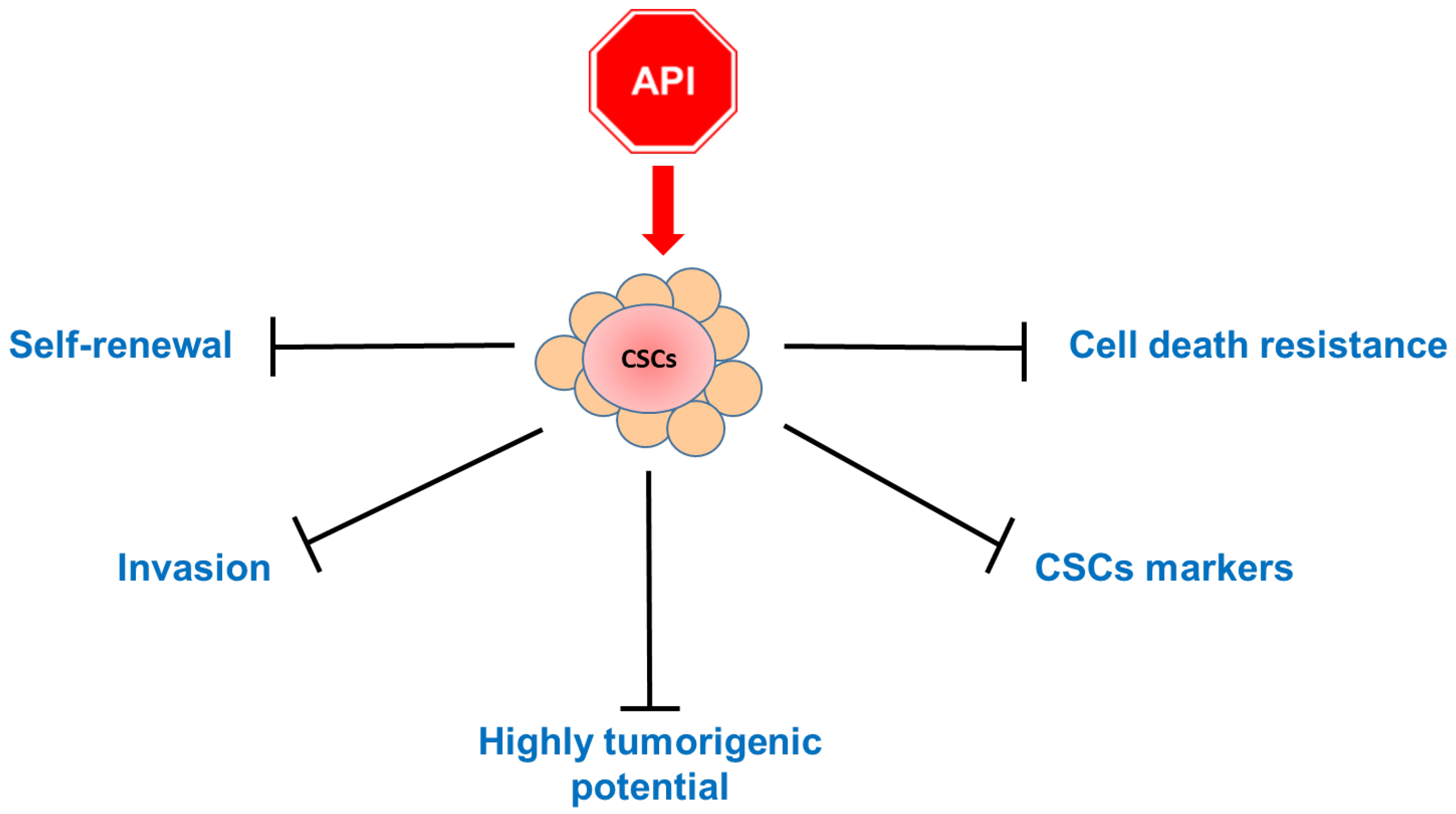
5. Apigenin Inhibits Cancer Stem Cells
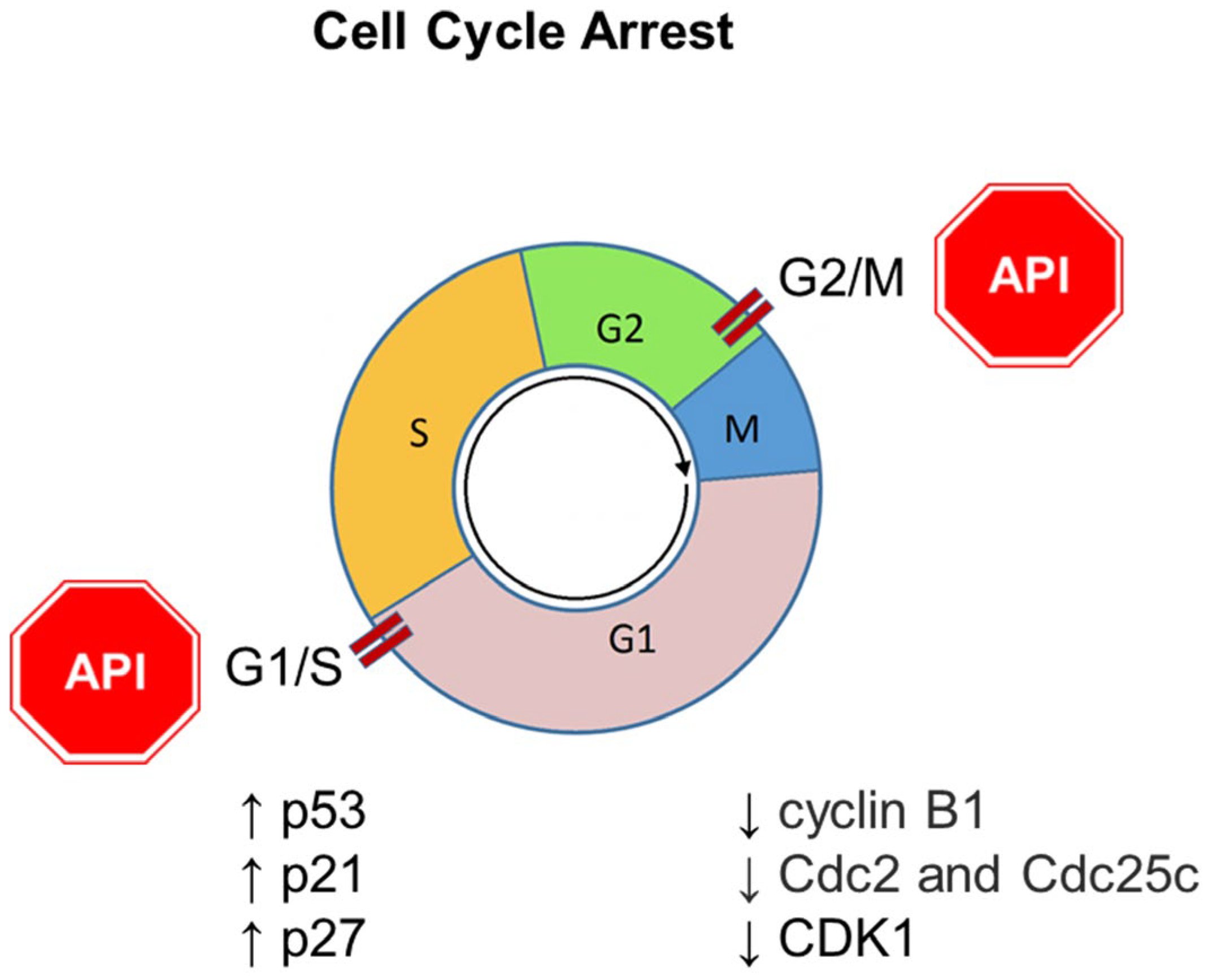
6. Apigenin and Cell Cycle Arrest
7. Apigenin and Programmed Cell Death
7.1. Apoptosis
7.2. Autophagy
7.3. Ferroptosis
7.4. Necroptosis
7.5. Anoikis
8. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABCG2 | ATP-binding cassette subfamily G member 2 |
| Akt | Protein kinase B |
| ALDH1 | Acetaldehyde dehydrogenase 1 |
| AMPK | AMP-activated protein kinase |
| ATF4 | Activating transcription factor 4 |
| ATM | Ataxia-telangiectasia mutated |
| Bad | Bcl-2-associated death promoter |
| Bak | Bcl-2 antagonist/killer |
| Bax | BCL2 associated X, apoptosis regulator |
| Bcl-2 | B-cell lymphoma 2 |
| Bim | Bcl-2 Interacting Mediator of cell death |
| BET | Bromodomain and extraterminal domain |
| BRD4 | Bromodomain containing 4 |
| CC | Colon cancer |
| CD | Cluster of differentiation |
| cdc | Cell division cycle |
| CDK | Cyclin-dependent kinase |
| CHOP | C/EBP homologous protein |
| CK2 | Casein kinase 2 |
| COX | Cyclooxygenase |
| CSCs | Cancer stem cells |
| CTGF | Connective tissue growth factor |
| CYR61 | Cysteine-rich angiogenic inducer 61 |
| DAPK | Death-associated protein kinase |
| DNA | Deoxyribonucleic Acid |
| DR | Death receptors |
| EGFR | Epidermal growth factor receptor |
| ECM | Extracellular matrix |
| EMT | Epithelial–mesenchymal transition |
| ERK | Extracellular signal-related kinase |
| EZH2 | Enhancer of zeste homolog 2 |
| FAKs | Focal adhesion kinases |
| FOX | Forkhead box |
| FTH1 | Ferritin heavy chain 1 |
| 5-FU | 5-fluorouracil |
| Gli1 | GLI Family Zinc Finger 1 |
| GSH | Glutathione |
| GPX4 | Glutathione peroxidase 4 |
| GSK-3β | Glycogen synthase kinase 3-beta |
| HCC | Hepatocellular carcinoma |
| HDAC | Histone deacetylases |
| HDM2 | E3 ligase human double minute 2 |
| HeLa | Human cervical carcinoma cells |
| HGF | Hepatocyte growth factor |
| HIF-1α | Hypoxia-inducible factor 1α |
| HSP | Heat shock protein 90 |
| HUVECs | Human umbilical vein endothelial cells |
| IGF-IR | Type I insulin growth factor receptor |
| IL-6 | Interleukin 6 |
| IKKα | IκB kinase-α |
| IKK | IκB kinase |
| JAK | Janus kinase |
| JNK | Jun N-terminal kinase |
| LC3 | Light chain 3 |
| MAPK | Mitogen-activated protein kinase |
| Mcl-1 | Myeloid cell leukemia-1 |
| MEK | MAP kinase-ERK kinase |
| miRNAs | MicroRNA |
| MLKL | Mixed lineage kinase |
| MMPs | Metalloproteases |
| mRNA | Messenger ribonucleic acid |
| mTOR | Mammalian target of rapamycin |
| MVD | Microvessel density |
| NF-κB | Nuclear factor-kappa B |
| iNOS | Inducible nitric oxide synthase |
| NQO2 | NRH-quinone oxidoreductase 2 |
| Notch | Signal transducer and activator of transcription |
| NSCLC | Non-small cell lung carcinoma |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| PARP | Poly-ADP ribose polymerase |
| PCD | Programmed cell death |
| PCSC | Prostate cancer stem cells |
| PDGF-BB | Platelet-derived growth factor-BB |
| PDGFR-β | Platelet-derived growth factor receptor β |
| PERK | Protein kinase RNA-like endoplasmic reticulum kinase |
| PI3K | Phosphatidylinositol-3-kinase |
| PKB | Protein kinase B |
| PLK-1 | Polo-like kinase 1 |
| PTEN | Phosphatase and tensin homolog |
| PRRs | Pattern recognition receptors |
| p70S6K1 | Ribosomal protein S6 kinase beta-1 |
| RIPK3 | Receptor-interacting serine/threonine kinase 3 |
| ROS | Reactive oxygen species |
| SIRT | Sirtuin |
| Snail | Snail homolog 1/2 of drosophila |
| Sox2 | SEX determining region (SRY) homology box 2 |
| SPOCK1 | Cwcv and kazal-like domains proteoglycan 1 |
| pSTAT3 | Phosphorylated signal transducer and activator of transcription-3 |
| TAZ | PDZ-binding motif |
| TCRs | T cell receptors |
| TEADs | Transcriptional enhanced associate domain |
| hTERT | Human telomerase reverse transcriptase |
| TGF-β | Transforming growth factor-beta |
| TMZ | Temozolomide |
| TNBC | Triple-negative breast cancer |
| TNF | Tumor necrosis factor |
| TNFR | Tumor necrosis factor receptor |
| TRAIL | TNF-related apoptosis-inducing ligand |
| TWIST | Twist family bHLH transcription factor |
| ULK1 | Kinase UNC51-like kinase-1 |
| VEGF | Vascular endothelial growth factor |
| VEGFR2 | Vascular endothelial growth factor receptor-2 |
| YAP | Hippo-Yes-associated protein |
| ZEB1/2 | Zinc finger E-box binding homeobox 1/2 |
| ZO-1 | Zonula Occludens-1 |
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer Statistics for the Year 2020: An Overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Stoletov, K.; Beatty, P.H.; Lewis, J.D. Novel Therapeutic Targets for Cancer Metastasis. Expert. Rev. Anticancer Ther. 2020, 20, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.P.; Massagué, J. Cancer Metastasis: Building a Framework. Cell 2006, 127, 679–695. [Google Scholar] [CrossRef] [PubMed]
- Suhail, Y.; Cain, M.P.; Vanaja, K.; Kurywchak, P.A.; Levchenko, A.; Kalluri, R. Kshitiz Systems Biology of Cancer Metastasis. Cell Syst. 2019, 9, 109–127. [Google Scholar] [CrossRef] [PubMed]
- Babaei, G.; Aziz, S.G.G.; Jaghi, N.Z.Z. EMT, Cancer Stem Cells and Autophagy; The Three Main Axes of Metastasis. Biomed. Pharmacother. 2021, 133, 110909. [Google Scholar] [CrossRef] [PubMed]
- Majidpoor, J.; Mortezaee, K. Steps in Metastasis: An Updated Review. Med. Oncol. 2021, 38, 3. [Google Scholar] [CrossRef] [PubMed]
- Hazafa, A.; Rehman, K.U.; Jahan, N.; Jabeen, Z. The Role of Polyphenol (Flavonoids) Compounds in the Treatment of Cancer Cells. Nutr. Cancer 2020, 72, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, A.H.; Alsahli, M.A.; Almatroudi, A.; Almogbel, M.A.; Khan, A.A.; Anwar, S.; Almatroodi, S.A. The Potential Role of Apigenin in Cancer Prevention and Treatment. Molecules 2022, 27, 6051. [Google Scholar] [CrossRef]
- Nozhat, Z.; Heydarzadeh, S.; Memariani, Z.; Ahmadi, A. Chemoprotective and Chemosensitizing Effects of Apigenin on Cancer Therapy. Cancer Cell Int. 2021, 21, 574. [Google Scholar] [CrossRef]
- Javed, Z.; Sadia, H.; Iqbal, M.J.; Shamas, S.; Malik, K.; Ahmed, R.; Raza, S.; Butnariu, M.; Cruz-Martins, N.; Sharifi-Rad, J. Apigenin Role as Cell-Signaling Pathways Modulator: Implications in Cancer Prevention and Treatment. Cancer Cell Int. 2021, 21, 189. [Google Scholar] [CrossRef] [PubMed]
- Ashrafizadeh, M.; Bakhoda, M.R.; Bahmanpour, Z.; Ilkhani, K.; Zarrabi, A.; Makvandi, P.; Khan, H.; Mazaheri, S.; Darvish, M.; Mirzaei, H. Apigenin as Tumor Suppressor in Cancers: Biotherapeutic Activity, Nanodelivery, and Mechanisms With Emphasis on Pancreatic Cancer. Front. Chem. 2020, 8, 829. [Google Scholar] [CrossRef]
- Sung, B.; Chung, H.Y.; Kim, N.D. Role of Apigenin in Cancer Prevention via the Induction of Apoptosis and Autophagy. J. Cancer Prev. 2016, 21, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Jäger, A.K.; Krydsfeldt, K.; Rasmussen, H.B. Bioassay-Guided Isolation of Apigenin with GABA-Benzodiazepine Activity from Tanacetum Parthenium. Phytother. Res. 2009, 23, 1642–1644. [Google Scholar] [CrossRef]
- Aslam Bhatti, H.; Noor, R. Isolation of Apigenin by Solute-Solvent Extraction from Symphotrichum Novea Anglea. Integr. Food Nutr. Metab. 2019, 6. [Google Scholar] [CrossRef]
- Daneshvar, S.; Zamanian, M.Y.; Ivraghi, M.S.; Golmohammadi, M.; Modanloo, M.; Kamiab, Z.; Pourhosseini, S.M.E.; Heidari, M.; Bazmandegan, G. A Comprehensive View on the Apigenin Impact on Colorectal Cancer: Focusing on Cellular and Molecular Mechanisms. Food Sci. Nutr. 2023, 11, 6789–6801. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Firrman, J.; Liu, L.S.; Yam, K. A Review on Flavonoid Apigenin: Dietary Intake, ADME, Antimicrobial Effects, and Interactions with Human Gut Microbiota. BioMed Res. Int. 2019, 2019, 7010467. [Google Scholar] [CrossRef]
- Thomas, S.D.; Jha, N.K.; Jha, S.K.; Sadek, B.; Ojha, S. Pharmacological and Molecular Insight on the Cardioprotective Role of Apigenin. Nutrients 2023, 15, 385. [Google Scholar] [CrossRef]
- Tang, D.; Chen, K.; Huang, L.; Li, J. Pharmacokinetic Properties and Drug Interactions of Apigenin, a Natural Flavone. Expert. Opin. Drug Metab. Toxicol. 2017, 13, 323–330. [Google Scholar] [CrossRef]
- Gradolatto, A.; Basly, J.P.; Berges, R.; Teyssier, C.; Chagnon, M.C.; Siess, M.H.; Canivenc-Lavier, M.C. Pharmacokinetics and Metabolism of Apigenin in Female and Male Rats after a Single Oral Administration. Drug Metab. Dispos. 2005, 33, 49–54. [Google Scholar] [CrossRef]
- Lee, I.G.; Lee, J.; Hong, S.H.; Seo, Y.J. Apigenin’s Therapeutic Potential Against Viral Infection. Front. Biosci. 2023, 28, 237. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zhang, H.; Zhang, X.; Xia, L.; Zhang, J.J. Research Progress on Antisepsis Effect of Apigenin and Its Mechanism of Action. Heliyon 2023, 9, e22290. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, P.; Shikha, D.; Thakur, M.; Aneja, A. Functionality of Apigenin as a Potent Antioxidant with Emphasis on Bioavailability, Metabolism, Action Mechanism and in Vitro and in Vivo Studies: A Review. J. Food Biochem. 2022, 46, e13950. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Qi, M.; Li, P.; Zhan, Y.; Shao, H. Apigenin in Cancer Therapy: Anti-Cancer Effects and Mechanisms of Action. Cell Biosci. 2017, 7, 50. [Google Scholar] [CrossRef] [PubMed]
- Liekens, S.; De Clercq, E.; Neyts, J. Angiogenesis: Regulators and Clinical Applications. Biochem. Pharmacol. 2001, 61, 253–270. [Google Scholar] [CrossRef] [PubMed]
- Unwith, S.; Zhao, H.; Hennah, L.; Ma, D. The Potential Role of HIF on Tumour Progression and Dissemination. Int. J. Cancer 2015, 136, 2491–2503. [Google Scholar] [CrossRef]
- Krock, B.L.; Skuli, N.; Simon, M.C. Hypoxia-Induced Angiogenesis: Good and Evil. Genes Cancer 2011, 2, 1117–1133. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Zeng, W.; Chen, M.; Huang, L.; Li, S.; Li, Z.; Pan, Q.; Lv, S.; Yang, X.; Wang, Y.; et al. Apigenin Suppresses Tumor Angiogenesis and Growth via Inhibiting HIF-1α Expression in Non-Small Cell Lung Carcinoma. Chem. Biol. Interact. 2022, 361, 109966. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Xia, C.; Cao, Z.; Zheng, J.Z.; Reed, E.; Jiang, B.-H. Apigenin Inhibits VEGF and HIF-1 Expression via PI3K/AKT/P70S6K1 and HDM2/P53 Pathways. FASEB J. 2005, 19, 342–353. [Google Scholar] [CrossRef]
- Liu, L.Z.; Fang, J.; Zhou, Q.; Hu, X.; Shi, X.; Jiang, B.H. Apigenin Inhibits Expression of Vascular Endothelial Growth Factor and Angiogenesis in Human Lung Cancer Cells: Implication of Chemoprevention of Lung Cancer. Mol. Pharmacol. 2005, 68, 635–643. [Google Scholar] [CrossRef]
- Choi, H.J.; Eun, J.S.; Kim, B.G.; Kim, S.Y.; Jeon, H.; Soh, Y. Vitexin, an HIF-1alpha Inhibitor, Has Anti-Metastatic Potential in PC12 Cells. Mol. Cells 2006, 22, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Brabletz, S.; Schuhwerk, H.; Brabletz, T.; Stemmler, M.P. Dynamic EMT: A Multi-Tool for Tumor Progression. EMBO J. 2021, 40, e108647. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Hong, W.; Wei, X. The Molecular Mechanisms and Therapeutic Strategies of EMT in Tumor Progression and Metastasis. J. Hematol. Oncol. 2022, 15, 129. [Google Scholar] [CrossRef] [PubMed]
- Akrida, I.; Bravou, V.; Papadaki, H. The Deadly Cross-Talk between Hippo Pathway and Epithelial-Mesenchymal Transition (EMT) in Cancer. Mol. Biol. Rep. 2022, 49, 10065–10076. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Xing, T.; Yang, Z.; Dudek, R.; Lu, Q.; Chen, Y.H. Epithelial Mesenchymal Transition in Embryonic Development, Tissue Repair and Cancer: A Comprehensive Overview. J. Clin. Med. 2017, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, D.M.; Medici, D. Signaling Mechanisms of the Epithelial-Mesenchymal Transition. Sci. Signal 2014, 7, re8. [Google Scholar] [CrossRef] [PubMed]
- Pastushenko, I.; Blanpain, C. EMT Transition States during Tumor Progression and Metastasis. Trends Cell Biol. 2019, 29, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Shim, J.S. Targeting Epithelial-Mesenchymal Transition (EMT) to Overcome Drug Resistance in Cancer. Molecules 2016, 21, 965. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Shen, Y.; Zhang, Z.; Hu, Y.; Zhang, X.; Han, L. Apigenin Inhibits Epithelial-Mesenchymal Transition of Human Colon Cancer Cells through NF-ΚB/Snail Signaling Pathway. Biosci. Rep. 2019, 39, BSR20190452. [Google Scholar] [CrossRef]
- Qin, Y.; Zhao, D.; Zhou, H.G.; Wang, X.H.; Zhong, W.L.; Chen, S.; Gu, W.G.; Wang, W.; Zhang, C.H.; Liu, Y.R.; et al. Apigenin Inhibits NF-ΚB and Snail Signaling, EMT and Metastasis in Human Hepatocellular Carcinoma. Oncotarget 2016, 7, 41421–41431. [Google Scholar] [CrossRef]
- Chien, M.H.; Lin, Y.W.; Wen, Y.C.; Yang, Y.C.; Hsiao, M.; Chang, J.L.; Huang, H.C.; Lee, W.J. Targeting the SPOCK1-Snail/Slug Axis-Mediated Epithelial-to-Mesenchymal Transition by Apigenin Contributes to Repression of Prostate Cancer Metastasis. J. Exp. Clin. Cancer Res. 2019, 38, 246. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.R.; Li, S.Y.; Guo, Q.S.; Zhou, W.; Zhang, H.M. SPOCK1 Involvement in Epithelial-to-Mesenchymal Transition: A New Target in Cancer Therapy? Cancer Manag. Res. 2020, 12, 3561–3569. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Zhou, H.; Liu, G.; Zhao, Y.; Cao, G.; Liu, Q. SPOCK1 Promotes the Invasion and Metastasis of Gastric Cancer through Slug-Induced Epithelial-Mesenchymal Transition. J. Cell Mol. Med. 2018, 22, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yao, Y.T.; Xu, H.; Chen, Y.B.; Gu, M.; Cai, Z.K.; Wang, Z. SPOCK1 Promotes Tumor Growth and Metastasis in Human Prostate Cancer. Drug Des. Devel. Ther. 2016, 10, 2311–2321. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Wu, J.X.; Yang, S.F.; Yang, C.K.; Chen, T.H.; Hsiao, Y.H. Anticancer Effects and Molecular Mechanisms of Apigenin in Cervical Cancer Cells. Cancers 2022, 14, 1824. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhou, H.B.; Liu, J.; Xie, J.; Hu, R. Apigenin Suppresses Proliferation, Invasion, and Epithelial-Mesenchymal Transition of Cervical Carcinoma Cells by Regulation of MiR-152/BRD4 Axis. Kaohsiung J. Med. Sci. 2021, 37, 583–593. [Google Scholar] [CrossRef]
- Chang, J.H.; Cheng, C.W.; Yang, Y.C.; Chen, W.S.; Hung, W.Y.; Chow, J.M.; Chen, P.S.; Hsiao, M.; Lee, W.J.; Chien, M.H. Downregulating CD26/DPPIV by Apigenin Modulates the Interplay between Akt and Snail/Slug Signaling to Restrain Metastasis of Lung Cancer with Multiple EGFR Statuses. J. Exp. Clin. Cancer Res. 2018, 37, 199. [Google Scholar] [CrossRef]
- Lee, H.H.; Jung, J.; Moon, A.; Kang, H.; Cho, H. Antitumor and Anti-Invasive Effect of Apigenin on Human Breast Carcinoma through Suppression of IL-6 Expression. Int. J. Mol. Sci. 2019, 20, 3143. [Google Scholar] [CrossRef]
- Cao, H.H.; Chu, J.H.; Kwan, H.Y.; Su, T.; Yu, H.; Cheng, C.Y.; Fu, X.Q.; Guo, H.; Li, T.; Tse, A.K.W.; et al. Inhibition of the STAT3 Signaling Pathway Contributes to Apigenin-Mediated Anti-Metastatic Effect in Melanoma. Sci. Rep. 2016, 6, 21731. [Google Scholar] [CrossRef]
- García-García, V.A.; Alameda, J.P.; Page, A.; Mérida-García, A.; Navarro, M.; Tejero, A.; Paramio, J.M.; García-Fernández, R.A.; Casanova, M.L. IKKα Induces Epithelial-Mesenchymal Changes in Mouse Skin Carcinoma Cells That Can Be Partially Reversed by Apigenin. Int. J. Mol. Sci. 2022, 23, 1375. [Google Scholar] [CrossRef]
- Ganesan, K.; Xu, B. Molecular Targets of Vitexin and Isovitexin in Cancer Therapy: A Critical Review. Ann. N. Y. Acad. Sci. 2017, 1401, 102–113. [Google Scholar] [CrossRef]
- Zhu, H.; Zhao, N.; Jiang, M. Isovitexin Attenuates Tumor Growth in Human Colon Cancer Cells through the Modulation of Apoptosis and Epithelial-Mesenchymal Transition via PI3K/Akt/MTOR Signaling Pathway. Biochem. Cell Biol. 2021, 99, 741–749. [Google Scholar] [CrossRef]
- Atashzar, M.R.; Baharlou, R.; Karami, J.; Abdollahi, H.; Rezaei, R.; Pourramezan, F.; Zoljalali Moghaddam, S.H. Cancer Stem Cells: A Review from Origin to Therapeutic Implications. J. Cell Physiol. 2020, 235, 790–803. [Google Scholar] [CrossRef]
- Tong, W.W.; Tong, G.H.; Liu, Y. Cancer Stem Cells and Hypoxia-Inducible Factors (Review). Int. J. Oncol. 2018, 53, 469–476. [Google Scholar] [CrossRef]
- Peng, F.; Liao, M.; Qin, R.; Zhu, S.; Peng, C.; Fu, L.; Chen, Y.; Han, B. Regulated Cell Death (RCD) in Cancer: Key Pathways and Targeted Therapies. Signal Transduct. Target. Ther. 2022, 7, 286. [Google Scholar] [CrossRef]
- Walcher, L.; Kistenmacher, A.K.; Suo, H.; Kitte, R.; Dluczek, S.; Strauß, A.; Blaudszun, A.R.; Yevsa, T.; Fricke, S.; Kossatz-Boehlert, U. Cancer Stem Cells-Origins and Biomarkers: Perspectives for Targeted Personalized Therapies. Front. Immunol. 2020, 11, 1280. [Google Scholar] [CrossRef]
- Carnero, A.; Garcia-Mayea, Y.; Mir, C.; Lorente, J.; Rubio, I.T.; LLeonart, M.E. The Cancer Stem-Cell Signaling Network and Resistance to Therapy. Cancer Treat. Rev. 2016, 49, 25–36. [Google Scholar] [CrossRef]
- Yang, L.; Shi, P.; Zhao, G.; Xu, J.; Peng, W.; Zhang, J.; Zhang, G.; Wang, X.; Dong, Z.; Chen, F.; et al. Targeting Cancer Stem Cell Pathways for Cancer Therapy. Signal Transduct. Target. Ther. 2020, 5, 8. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X. Targeting the Wnt/β-Catenin Signaling Pathway in Cancer. J. Hematol. Oncol. 2020, 13, 165. [Google Scholar] [CrossRef]
- Lian, I.; Kim, J.; Okazawa, H.; Zhao, J.; Zhao, B.; Yu, J.; Chinnaiyan, A.; Israel, M.A.; Goldstein, L.S.B.; Abujarour, R.; et al. The Role of YAP Transcription Coactivator in Regulating Stem Cell Self-Renewal and Differentiation. Genes Dev. 2010, 24, 1106–1118. [Google Scholar] [CrossRef]
- Ajani, J.A.; Song, S.; Hochster, H.S.; Steinberg, I.B. Cancer Stem Cells: The Promise and the Potential. Semin. Oncol. 2015, 42 (Suppl. 1), S3–S17. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, T.; Ohata, H.; Sato, A.; Yamawaki, K.; Enomoto, T.; Okamoto, K. Tumor-Derived Spheroids: Relevance to Cancer Stem Cells and Clinical Applications. Cancer Sci. 2017, 108, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Yousefnia, S.; Ghaedi, K.; Seyed Forootan, F.; Nasr Esfahani, M.H. Characterization of the Stemness Potency of Mammospheres Isolated from the Breast Cancer Cell Lines. Tumour Biol. 2019, 41, 1010428319869101. [Google Scholar] [CrossRef] [PubMed]
- Dandawate, P.R.; Subramaniam, D.; Jensen, R.A.; Anant, S. Targeting Cancer Stem Cells and Signaling Pathways by Phytochemicals: Novel Approach for Breast Cancer Therapy. Semin. Cancer Biol. 2016, 40–41, 192–208. [Google Scholar] [CrossRef]
- Gu, H.F.; Mao, X.Y.; Du, M. Prevention of Breast Cancer by Dietary Polyphenols-Role of Cancer Stem Cells. Crit. Rev. Food Sci. Nutr. 2020, 60, 810–825. [Google Scholar] [CrossRef]
- Ghanbari-Movahed, M.; Shafiee, S.; Burcher, J.T.; Lagoa, R.; Farzaei, M.H.; Bishayee, A. Anticancer Potential of Apigenin and Isovitexin with Focus on Oncogenic Metabolism in Cancer Stem Cells. Metabolites 2023, 13, 404. [Google Scholar] [CrossRef]
- Kim, B.; Jung, N.; Lee, S.; Sohng, J.K.; Jung, H.J. Apigenin Inhibits Cancer Stem Cell-Like Phenotypes in Human Glioblastoma Cells via Suppression of c-Met Signaling. Phytother. Res. 2016, 30, 1833–1840. [Google Scholar] [CrossRef]
- Hong, W.; Guan, K.L. The YAP and TAZ Transcription Co-Activators: Key Downstream Effectors of the Mammalian Hippo Pathway. Semin. Cell Dev. Biol. 2012, 23, 785–793. [Google Scholar] [CrossRef]
- Li, Y.W.; Xu, J.; Zhu, G.Y.; Huang, Z.J.; Lu, Y.; Li, X.Q.; Wang, N.; Zhang, F.X. Apigenin Suppresses the Stem Cell-like Properties of Triple-Negative Breast Cancer Cells by Inhibiting YAP/TAZ Activity. Cell Death Discov. 2018, 4, 105. [Google Scholar] [CrossRef]
- Sharma, A.; Sinha, S.; Keswani, H.; Shrivastava, N. Kaempferol and Apigenin Suppresses the Stemness Properties of TNBC Cells by Modulating Sirtuins. Mol. Divers. 2022, 26, 3225–3240. [Google Scholar] [CrossRef]
- Sharpe, B.; Beresford, M.; Bowen, R.; Mitchard, J.; Chalmers, A.D. Searching for Prostate Cancer Stem Cells: Markers and Methods. Stem Cell Rev. Rep. 2013, 9, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Erdogan, S.; Doganlar, O.; Doganlar, Z.B.; Serttas, R.; Turkekul, K.; Dibirdik, I.; Bilir, A. The Flavonoid Apigenin Reduces Prostate Cancer CD44(+) Stem Cell Survival and Migration through PI3K/Akt/NF-ΚB Signaling. Life Sci. 2016, 162, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Jiang, Y.; Sun, L. Vitexin Suppresses the Proliferation, Angiogenesis and Stemness of Endometrial Cancer through the PI3K/AKT Pathway. Pharm. Biol. 2023, 61, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Firnau, M.B.; Brieger, A. CK2 and the Hallmarks of Cancer. Biomedicines 2022, 10, 1987. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, Y.; Mao, J.H.; Hsieh, D.; Kim, I.J.; Hu, L.M.; Xu, Z.; Long, H.; Jablons, D.M.; You, L. Inhibition of CK2α Down-Regulates Hedgehog/Gli Signaling Leading to a Reduction of a Stem-like Side Population in Human Lung Cancer Cells. PLoS ONE 2012, 7, e38996. [Google Scholar] [CrossRef] [PubMed]
- Tang, A.Q.; Cao, X.C.; Tian, L.; He, L.; Liu, F. Apigenin Inhibits the Self-Renewal Capacity of Human Ovarian Cancer SKOV3-derived Sphere-Forming Cells. Mol. Med. Rep. 2015, 11, 2221–2226. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cao, X.C.; Xiao, Q.; Quan, M.F. Apigenin Inhibits HeLa Sphere-Forming Cells through Inactivation of Casein Kinase 2α. Mol. Med. Rep. 2015, 11, 665–669. [Google Scholar] [CrossRef]
- Ketkaew, Y.; Osathanon, T.; Pavasant, P.; Sooampon, S. Apigenin Inhibited Hypoxia Induced Stem Cell Marker Expression in a Head and Neck Squamous Cell Carcinoma Cell Line. Arch. Oral. Biol. 2017, 74, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Cao, X.; Cao, X.; Liu, L.; Qiu, Y.; Li, X.; Zhou, L.; Ning, Y.; Ren, K.; Cao, J. Isovitexin Inhibits Stemness and Induces Apoptosis in Hepatocellular Carcinoma SK-Hep-1 Spheroids by Upregulating MiR-34a Expression. Anticancer Agents Med. Chem. 2020, 20, 1654–1663. [Google Scholar] [CrossRef]
- Huang, T.; Alvarez, A.; Hu, B.; Cheng, S.Y. Noncoding RNAs in Cancer and Cancer Stem Cells. Chin. J. Cancer 2013, 32, 582–593. [Google Scholar] [CrossRef]
- Li, Y.; Chen, X.; He, W.; Xia, S.; Jiang, X.; Li, X.; Bai, J.; Li, N.; Chen, L.; Yang, B. Apigenin Enhanced Antitumor Effect of Cisplatin in Lung Cancer via Inhibition of Cancer Stem Cells. Nutr. Cancer 2021, 73, 1489–1497. [Google Scholar] [CrossRef] [PubMed]
- Erdogan, S.; Turkekul, K.; Serttas, R.; Erdogan, Z. The Natural Flavonoid Apigenin Sensitizes Human CD44+ Prostate Cancer Stem Cells to Cisplatin Therapy. Biomed. Pharmacother. 2017, 88, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, W.; Chen, F.; Lu, Z. Combined Effect of Chrysin and Apigenin on Inhibiting the Development and Progression of Colorectal Cancer by Suppressing the Activity of P38-MAPK/AKT Pathway. IUBMB Life 2021, 73, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Farghadani, R.; Naidu, R. The Anticancer Mechanism of Action of Selected Polyphenols in Triple-Negative Breast Cancer (TNBC). Biomed. Pharmacother. 2023, 165, 115170. [Google Scholar] [CrossRef]
- Abotaleb, M.; Samuel, S.M.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Büsselberg, D. Flavonoids in Cancer and Apoptosis. Cancers 2018, 11, 28. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, S.; Geng, B.; Yi, Z. Pelargonidin Induces Antitumor Effects in Human Osteosarcoma Cells via Autophagy Induction, Loss of Mitochondrial Membrane Potential, G2/M Cell Cycle Arrest and Downregulation of PI3K/AKT Signalling Pathway. J. BUON 2018, 23, 735–740. [Google Scholar] [PubMed]
- Dash, B.C.; El-Deiry, W.S. Phosphorylation of P21 in G2/M Promotes Cyclin B-Cdc2 Kinase Activity. Mol. Cell Biol. 2005, 25, 3364–3387. [Google Scholar] [CrossRef] [PubMed]
- Ittiudomrak, T.; Puthong, S.; Roytrakul, S.; Chanchao, C. α-Mangostin and Apigenin Induced Cell Cycle Arrest and Programmed Cell Death in SKOV-3 Ovarian Cancer Cells. Toxicol. Res. 2019, 35, 167–179. [Google Scholar] [CrossRef]
- Lee, Y.; Sung, B.; Kang, Y.J.; Kim, D.H.; Jang, J.Y.; Hwang, S.Y.; Kim, M.; Lim, H.S.; Yoon, J.H.; Chung, H.Y.; et al. Apigenin-Induced Apoptosis Is Enhanced by Inhibition of Autophagy Formation in HCT116 Human Colon Cancer Cells. Int. J. Oncol. 2014, 44, 1599–1606. [Google Scholar] [CrossRef]
- Zhang, L.; Cheng, X.; Gao, Y.; Zheng, J.; Xu, Q.; Sun, Y.; Guan, H.; Yu, H.; Sun, Z. Apigenin Induces Autophagic Cell Death in Human Papillary Thyroid Carcinoma BCPAP Cells. Food Funct. 2015, 6, 3464–3472. [Google Scholar] [CrossRef]
- Ujiki, M.B.; Ding, X.Z.; Salabat, M.R.; Bentrem, D.J.; Golkar, L.; Milam, B.; Talamonti, M.S.; Bell, R.H.; Iwamura, T.; Adrian, T.E. Apigenin Inhibits Pancreatic Cancer Cell Proliferation through G2/M Cell Cycle Arrest. Mol. Cancer 2006, 5, 76. [Google Scholar] [CrossRef]
- Lepley, D.M.; Li, B.; Birt, D.F.; Pelling, J.C. The Chemopreventive Flavonoid Apigenin Induces G2/M Arrest in Keratinocytes. Carcinogenesis 1996, 17, 2367–2375. [Google Scholar] [CrossRef]
- O’Prey, J.; Brown, J.; Fleming, J.; Harrison, P.R. Effects of Dietary Flavonoids on Major Signal Transduction Pathways in Human Epithelial Cells. Biochem. Pharmacol. 2003, 66, 2075–2088. [Google Scholar] [CrossRef]
- Casagrande, F.; Darbon, J.M. Effects of Structurally Related Flavonoids on Cell Cycle Progression of Human Melanoma Cells: Regulation of Cyclin-Dependent Kinases CDK2 and CDK1. Biochem. Pharmacol. 2001, 61, 1205–1215. [Google Scholar] [CrossRef]
- McVean, M.; Weinberg, W.C.; Pelling, J.C. A P21(Waf1)-Independent Pathway for Inhibitory Phosphorylation of Cyclin-Dependent Kinase P34(Cdc2) and Concomitant G(2)/M Arrest by the Chemopreventive Flavonoid Apigenin. Mol. Carcinog. 2002, 33, 36–43. [Google Scholar] [CrossRef]
- Gupta, S.; Afaq, F.; Mukhtar, H. Selective Growth-Inhibitory, Cell-Cycle Deregulatory and Apoptotic Response of Apigenin in Normal versus Human Prostate Carcinoma Cells. Biochem. Biophys. Res. Commun. 2001, 287, 914–920. [Google Scholar] [CrossRef]
- Wang, W.; Heideman, L.; Chung, C.S.; Pelling, J.C.; Koehler, K.J.; Birt, D.F. Cell-Cycle Arrest at G2/M and Growth Inhibition by Apigenin in Human Colon Carcinoma Cell Lines. Mol. Carcinog. 2000, 28, 102–110. [Google Scholar] [CrossRef]
- Zhao, G.; Han, X.; Cheng, W.; Ni, J.; Zhang, Y.; Lin, J.; Song, Z. Apigenin Inhibits Proliferation and Invasion, and Induces Apoptosis and Cell Cycle Arrest in Human Melanoma Cells. Oncol. Rep. 2017, 37, 2277–2285. [Google Scholar] [CrossRef] [PubMed]
- Tseng, T.H.; Chien, M.H.; Lin, W.L.; Wen, Y.C.; Chow, J.M.; Chen, C.K.; Kuo, T.C.; Lee, W.J. Inhibition of MDA-MB-231 Breast Cancer Cell Proliferation and Tumor Growth by Apigenin through Induction of G2/M Arrest and Histone H3 Acetylation-Mediated P21WAF1/CIP1 Expression. Environ. Toxicol. 2017, 32, 434–444. [Google Scholar] [CrossRef]
- Shendge, A.K.; Chaudhuri, D.; Mandal, N. The Natural Flavones, Acacetin and Apigenin, Induce Cdk-Cyclin Mediated G2/M Phase Arrest and Trigger ROS-Mediated Apoptosis in Glioblastoma Cells. Mol. Biol. Rep. 2021, 48, 539–549. [Google Scholar] [CrossRef]
- Der Shi, M.; Shiao, C.K.; Lee, Y.C.; Shih, Y.W. Apigenin, a Dietary Flavonoid, Inhibits Proliferation of Human Bladder Cancer T-24 Cells via Blocking Cell Cycle Progression and Inducing Apoptosis. Cancer Cell Int. 2015, 15, 33. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Mao, Y.; Chen, H.; Lin, Y.; Hu, Z.; Wu, J.; Xu, X.; Xu, X.; Qin, J.; Xie, L. Apigenin Promotes Apoptosis, Inhibits Invasion and Induces Cell Cycle Arrest of T24 Human Bladder Cancer Cells. Cancer Cell Int. 2013, 13, 54. [Google Scholar] [CrossRef]
- Kim, N.W.; Piatyszek, M.A.; Prowse, K.R.; Harley, C.B.; West, M.D.; Ho, P.L.C.; Coviello, G.M.; Wright, W.E.; Weinrich, S.L.; Shay, J.W. Specific Association of Human Telomerase Activity with Immortal Cells and Cancer. Science 1994, 266, 2011–2015. [Google Scholar] [CrossRef]
- Cabuy, E.; De Ridder, L. Telomerase Activity and Expression of Telomerase Reverse Transcriptase Correlated with Cell Proliferation in Meningiomas and Malignant Brain Tumors in Vivo. Virchows Arch. 2001, 439, 176–184. [Google Scholar] [CrossRef]
- Chakrabarti, M.; Banik, N.L.; Ray, S.K. Sequential HTERT Knockdown and Apigenin Treatment Inhibited Invasion and Proliferation and Induced Apoptosis in Human Malignant Neuroblastoma SK-N-DZ and SK-N-BE2 Cells. J. Mol. Neurosci. 2013, 51, 187–198. [Google Scholar] [CrossRef]
- Lepley, D.M.; Pelling, J.C. Induction of P21/WAF1 and G1 Cell-Cycle Arrest by the Chemopreventive Agent Apigenin. Mol. Carcinog. 1997, 19, 75–82. [Google Scholar] [CrossRef]
- Zheng, P.W.; Chiang, L.C.; Lin, C.C. Apigenin Induced Apoptosis through P53-Dependent Pathway in Human Cervical Carcinoma Cells. Life Sci. 2005, 76, 1367–1379. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, Z.; Dai, X.; Zhang, L.; Li, M. Apigenin and Temozolomide Synergistically Inhibit Glioma Growth Through the PI3K/ AKT Pathway. Cancer Biother. Radiopharm. 2024, 39, 125–132. [Google Scholar] [CrossRef]
- Park, J.H.; Pyun, W.Y.; Park, H.W. Cancer Metabolism: Phenotype, Signaling and Therapeutic Targets. Cells 2020, 9, 2308. [Google Scholar] [CrossRef]
- Lee, E.; Song, C.H.; Bae, S.J.; Ha, K.T.; Karki, R. Regulated Cell Death Pathways and Their Roles in Homeostasis, Infection, Inflammation, and Tumorigenesis. Exp. Mol. Med. 2023, 55, 1632–1643. [Google Scholar] [CrossRef]
- He, S.; Huang, Q.; Cheng, J. The Unfolding Story of Dying Tumor Cells during Cancer Treatment. Front. Immunol. 2023, 14, 1073561. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, B.A.; El-Deiry, W.S. Targeting Apoptosis in Cancer Therapy. Nat. Rev. Clin. Oncol. 2020, 17, 395–417. [Google Scholar] [CrossRef] [PubMed]
- Nonnenmacher, L.; Hasslacher, S.; Zimmermann, J.; Karpel-Massler, G.; La Ferla-Brühl, K.; Barry, S.E.; Burster, T.; Siegelin, M.D.; Brühl, O.; Halatsch, M.E.; et al. Cell Death Induction in Cancer Therapy—Past, Present, and Future. Crit. Rev. Oncog. 2016, 21, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Amini, P.; Moazamiyanfar, R.; Dakkali, M.S.; Jafarzadeh, E.; Ganjizadeh, M.; Rastegar-Pouyani, N.; Moloudi, K.; Khodamoradi, E.; Taeb, S.; Najafi, M. Induction of Cancer Cell Death by Apigenin: A Review on Different Cell Death Pathways. Mini Rev. Med. Chem. 2023, 23, 1461–1478. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.Y.; Sung, B.; Kim, N.D. Role of Induced Programmed Cell Death in the Chemopreventive Potential of Apigenin. Int. J. Mol. Sci. 2022, 23, 3757. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Garg, V.K.; Goel, N. Intrinsic and Extrinsic Pathways of Apoptosis: Role in Cancer Development and Prognosis. Adv. Protein Chem. Struct. Biol. 2021, 125, 73–120. [Google Scholar] [CrossRef] [PubMed]
- Kilani-Jaziri, S.; Frachet, V.; Bhouri, W.; Ghedira, K.; Chekir-Ghedira, L.; Ronot, X. Flavones Inhibit the Proliferation of Human Tumor Cancer Cell Lines by Inducing Apoptosis. Drug Chem. Toxicol. 2012, 35, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bahreghani, M.T.; Geraily, G.; Alizadeh, S.; Najafi, M.; Shirazi, A. Apigenin Enhanced Radiation-Induced Apoptosis/Necrosis by Sensitization of LNCaP Prostate Cancer Cells to 6 MV Photon Beams. Cell J. 2021, 23, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.; Chang, C.Y.; Lee, K.R.; Lin, H.J.; Chen, T.H.; Wan, L. Flavones Inhibit Breast Cancer Proliferation through the Akt/FOXO3a Signaling Pathway. BMC Cancer 2015, 15, 958. [Google Scholar] [CrossRef]
- Johnson, J.L.; De Mejia, E.G. Flavonoid Apigenin Modified Gene Expression Associated with Inflammation and Cancer and Induced Apoptosis in Human Pancreatic Cancer Cells through Inhibition of GSK-3β/NF-ΚB Signaling Cascade. Mol. Nutr. Food Res. 2013, 57, 2112–2127. [Google Scholar] [CrossRef]
- Lu, H.F.; Chie, Y.U.J.; Yang, M.S.; Lee, C.S.; Fu, J.J.; Yang, J.S.; Tan, T.W.; Wu, S.H.; Ma, Y.I.S.; Ip, S.W.; et al. Apigenin Induces Caspase-Dependent Apoptosis in Human Lung Cancer A549 Cells through Bax- and Bcl-2-Triggered Mitochondrial Pathway. Int. J. Oncol. 2010, 36, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- Bruno, A.; Siena, L.; Gerbino, S.; Ferraro, M.; Chanez, P.; Giammanco, M.; Gjomarkaj, M.; Pace, E. Apigenin Affects Leptin/Leptin Receptor Pathway and Induces Cell Apoptosis in Lung Adenocarcinoma Cell Line. Eur. J. Cancer 2011, 47, 2042–2051. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.F.; Chie, Y.J.; Yang, M.S.; Lu, K.W.; Fu, J.J.; Yang, J.S.; Chen, H.Y.; Hsia, T.C.; Ma, C.Y.; Ip, S.W.; et al. Apigenin Induces Apoptosis in Human Lung Cancer H460 Cells through Caspase- and Mitochondria-Dependent Pathways. Hum. Exp. Toxicol. 2011, 30, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, J.; Li, Z.; Liu, C.; Yin, L. The Apoptotic Effect of Apigenin on Human Gastric Carcinoma Cells through Mitochondrial Signal Pathway. Tumour Biol. 2014, 35, 7719–7726. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Fu, P.; Gupta, S. Apigenin Induces Apoptosis by Targeting Inhibitor of Apoptosis Proteins and Ku70-Bax Interaction in Prostate Cancer. Apoptosis 2014, 19, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Der Way, T.; Kao, M.C.; Lin, J.K. Degradation of HER2/Neu by Apigenin Induces Apoptosis through Cytochrome c Release and Caspase-3 Activation in HER2/Neu-Overexpressing Breast Cancer Cells. FEBS Lett. 2005, 579, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Liu, B.; Cao, W.; Zhang, W.; Zhang, F.; Zhao, H.; Meng, R.; Zhang, L.; Niu, R.; Hao, X.; et al. Autophagy Inhibition Enhances Apigenin-Induced Apoptosis in Human Breast Cancer Cells. Chin. J. Cancer Res. 2013, 25, 212–222. [Google Scholar] [CrossRef]
- Chen, X.; Xu, H.; Yu, X.; Wang, X.; Zhu, X.; Xu, X. Apigenin Inhibits in Vitro and in Vivo Tumorigenesis in Cisplatin-Resistant Colon Cancer Cells by Inducing Autophagy, Programmed Cell Death and Targeting m-TOR/PI3K/Akt Signalling Pathway. JBUON 2019, 24, 488–493. [Google Scholar]
- Turktekin, M.; Konac, E.; Onen, H.I.; Alp, E.; Yilmaz, A.; Menevse, S. Evaluation of the Effects of the Flavonoid Apigenin on Apoptotic Pathway Gene Expression on the Colon Cancer Cell Line (HT29). J. Med. Food 2011, 14, 1107–1117. [Google Scholar] [CrossRef]
- Borah, S.M.; Kma, L.; Darjee, M.S.; Deka, D.; Lyngdoh, A.; Sharan, R.N.; Baruah, T.J. Apigenin Promotes Cell Death in NCI-H23 Cells by Upregulation of PTEN: Potential Involvement of the Binding of Apigenin with WWP2 Protein. J. Biomol. Struct. Dyn. 2023, 23, 1–15. [Google Scholar] [CrossRef]
- Huang, S.; Yu, M.; Shi, N.; Zhou, Y.; Li, F.; Li, X.; Huang, X.; Jin, J. Apigenin and Abivertinib, a Novel BTK Inhibitor Synergize to Inhibit Diffuse Large B-Cell Lymphoma in Vivo and Vitro. J. Cancer 2020, 11, 2123–2132. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.; Shukla, S.; Gupta, S. Plant Flavonoid Apigenin Inactivates Akt to Trigger Apoptosis in Human Prostate Cancer: An in Vitro and in Vivo Study. Carcinogenesis 2008, 29, 2210–2217. [Google Scholar] [CrossRef] [PubMed]
- Babcook, M.A.; Gupta, S. Apigenin Modulates Insulin-like Growth Factor Axis: Implications for Prevention and Therapy of Prostate Cancer. Curr. Drug Targets 2012.
- Shukla, S.; Gupta, S. Apigenin Suppresses Insulin-like Growth Factor I Receptor Signaling in Human Prostate Cancer: An in Vitro and in Vivo Study. Mol. Carcinog. 2009, 48, 243–252. [Google Scholar] [CrossRef]
- Johnstone, R.W.; Frew, A.J.; Smyth, M.J. The TRAIL Apoptotic Pathway in Cancer Onset, Progression and Therapy. Nat. Rev. Cancer 2008, 8, 782–798. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, J.M.; Zhou, J.Y.; Wu, G.S. The Role of TRAIL in Apoptosis and Immunosurveillance in Cancer. Cancers 2023, 15, 2752. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.H.; Sultana, S. Apigenin Induces Apoptosis in Hep G2 Cells: Possible Role of TNF-Alpha and IFN-Gamma. Toxicology 2006, 217, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, C.; O’Neill, A.; Spengler, B.; Christoffel, V.; Fitzpatrick, J.M.; Watson, R.W.G. Apigenin Drives the Production of Reactive Oxygen Species and Initiates a Mitochondrial Mediated Cell Death Pathway in Prostate Epithelial Cells. Prostate 2005, 63, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Oishi, M.; Iizumi, Y.; Taniguchi, T.; Goi, W.; Miki, T.; Sakai, T. Apigenin Sensitizes Prostate Cancer Cells to Apo2L/TRAIL by Targeting Adenine Nucleotide Translocase-2. PLoS ONE 2013, 8, e55922. [Google Scholar] [CrossRef]
- Chen, M.; Wang, X.; Zha, D.; Cai, F.; Zhang, W.; He, Y.; Huang, Q.; Zhuang, H.; Hua, Z.C. Apigenin Potentiates TRAIL Therapy of Non-Small Cell Lung Cancer via Upregulating DR4/DR5 Expression in a P53-Dependent Manner. Sci. Rep. 2016, 6, 35468. [Google Scholar] [CrossRef]
- Kim, E.Y.; Kim, A.K. Apigenin Sensitizes Huh-7 Human Hepatocellular Carcinoma Cells to TRAIL-Induced Apoptosis. Biomol. Ther. 2012, 20, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.Y.; Yu, J.S.; Yang, M.; Kim, A.K. Sub-Toxic Dose of Apigenin Sensitizes HepG2 Cells to TRAIL through ERK-Dependent up-Regulation of TRAIL Receptor DR5. Mol. Cells 2013, 35, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.H.; Molagoda, I.M.N.; Choi, Y.H.; Park, C.; Moon, D.O.; Kim, G.Y. Apigenin Promotes TRAIL-Mediated Apoptosis Regardless of ROS Generation. Food Chem. Toxicol. 2018, 111, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Masuelli, L.; Marzocchella, L.; Quaranta, A.; Palumbo, C.; Pompa, G.; Izzi, V.; Canini, A.; Modesti, A.; Galvano, F.; Bei, R. Apigenin Induces Apoptosis and Impairs Head and Neck Carcinomas EGFR/ErbB2 Signaling. Front. Biosci. 2011, 16, 1060–1068. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.P.; Chou, T.H.; Ding, H.Y.; Chen, P.R.; Chiang, F.Y.; Kuo, P.L.; Liang, C.H. Apigenin Induces Apoptosis via Tumor Necrosis Factor Receptor- and Bcl-2-Mediated Pathway and Enhances Susceptibility of Head and Neck Squamous Cell Carcinoma to 5-Fluorouracil and Cisplatin. Biochim. Biophys. Acta 2012, 1820, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- Stump, T.A.; Santee, B.N.; Williams, L.P.; Kunze, R.A.; Heinze, C.E.; Huseman, E.D.; Gryka, R.J.; Simpson, D.S.; Amos, S. The Antiproliferative and Apoptotic Effects of Apigenin on Glioblastoma Cells. J. Pharm. Pharmacol. 2017, 69, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Ayele, T.M.; Muche, Z.T.; Teklemariam, A.B.; Kassie, A.B.; Abebe, E.C. Role of JAK2/STAT3 Signaling Pathway in the Tumorigenesis, Chemotherapy Resistance, and Treatment of Solid Tumors: A Systemic Review. J. Inflamm. Res. 2022, 15, 1349–1364. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.S.; Jo, J.K.; Ku, J.M.; Choi, H.S.; Choi, Y.K.; Woo, J.K.; Kim, H.I.; Kang, S.Y.; Lee, K.M.; Nam, K.W.; et al. Induction of Caspase-Dependent Extrinsic Apoptosis by Apigenin through Inhibition of Signal Transducer and Activator of Transcription 3 (STAT3) Signalling in HER2-Overexpressing BT-474 Breast Cancer Cells. Biosci. Rep. 2015, 35, e00276. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.S.; Ku, J.M.; Choi, H.S.; Woo, J.K.; Jang, B.H.; Go, H.; Shin, Y.C.; Ko, S.G. Apigenin Induces Caspase-Dependent Apoptosis by Inhibiting Signal Transducer and Activator of Transcription 3 Signaling in HER2-Overexpressing SKBR3 Breast Cancer Cells. Mol. Med. Rep. 2015, 12, 2977–2984. [Google Scholar] [CrossRef]
- Seo, H.S.; Ku, J.M.; Choi, H.S.; Woo, J.K.; Jang, B.H.; Shin, Y.C.; Ko, S.G. Induction of Caspase-Dependent Apoptosis by Apigenin by Inhibiting STAT3 Signaling in HER2-Overexpressing MDA-MB-453 Breast Cancer Cells. Anticancer Res. 2014, 34, 2869–2882. [Google Scholar]
- Seo, H.S.; Choi, H.S.; Kim, S.R.; Choi, Y.K.; Woo, S.M.; Shin, I.; Woo, J.K.; Park, S.Y.; Shin, Y.C.; Ko, S.K. Apigenin Induces Apoptosis via Extrinsic Pathway, Inducing P53 and Inhibiting STAT3 and NFκB Signaling in HER2-Overexpressing Breast Cancer Cells. Mol. Cell Biochem. 2012, 366, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.J.; Kim, G.H. Apigenin Causes G(2)/M Arrest Associated with the Modulation of P21(Cip1) and Cdc2 and Activates P53-Dependent Apoptosis Pathway in Human Breast Cancer SK-BR-3 Cells. J. Nutr. Biochem. 2009, 20, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Takahashi, H.; Nakai, N.; Yanagita, T.; Ando, N.; Okubo, T.; Saito, K.; Shiga, K.; Hirokawa, T.; Hara, M.; et al. Apigenin Induces Apoptosis by Suppressing Bcl-Xl and Mcl-1 Simultaneously via Signal Transducer and Activator of Transcription 3 Signaling in Colon Cancer. Int. J. Oncol. 2018, 52, 1661–1673. [Google Scholar] [CrossRef]
- Mohan, N.; Ai, W.; Chakrabarti, M.; Banik, N.L.; Ray, S.K. KLF4 Overexpression and Apigenin Treatment down Regulated Anti-Apoptotic Bcl-2 Proteins and Matrix Metalloproteinases to Control Growth of Human Malignant Neuroblastoma SK-N-DZ and IMR-32 Cells. Mol. Oncol. 2013, 7, 464–474. [Google Scholar] [CrossRef]
- Yu, H.; Liu, Z.; Zhou, H.; Dai, W.; Chen, S.; Shu, Y.; Feng, J. JAK-STAT Pathway Modulates the Roles of INOS and COX-2 in the Cytoprotection of Early Phase of Hydrogen Peroxide Preconditioning against Apoptosis Induced by Oxidative Stress. Neurosci. Lett. 2012, 529, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Ruela-De-Sousa, R.R.; Fuhler, G.M.; Blom, N.; Ferreira, C.V.; Aoyama, H.; Peppelenbosch, M.P. Cytotoxicity of Apigenin on Leukemia Cell Lines: Implications for Prevention and Therapy. Cell Death Dis. 2010, 1, e19. [Google Scholar] [CrossRef]
- Budhraja, A.; Gao, N.; Zhang, Z.; Son, Y.O.; Cheng, S.; Wang, X.; Ding, S.; Hitron, A.; Chen, G.; Luo, J.; et al. Apigenin Induces Apoptosis in Human Leukemia Cells and Exhibits Anti-Leukemic Activity in Vivo. Mol. Cancer Ther. 2012, 11, 132–142. [Google Scholar] [CrossRef]
- Adham, A.N.; Abdelfatah, S.; Naqishbandi, A.M.; Mahmoud, N.; Efferth, T. Cytotoxicity of Apigenin toward Multiple Myeloma Cell Lines and Suppression of INOS and COX-2 Expression in STAT1-Transfected HEK293 Cells. Phytomedicine 2021, 80, 153371. [Google Scholar] [CrossRef]
- Arfin, S.; Jha, N.K.; Jha, S.K.; Kesari, K.K.; Ruokolainen, J.; Roychoudhury, S.; Rathi, B.; Kumar, D. Oxidative Stress in Cancer Cell Metabolism. Antioxidants 2021, 10, 642. [Google Scholar] [CrossRef]
- Bai, H.; Jin, H.; Yang, F.; Zhu, H.; Cai, J. Apigenin Induced MCF-7 Cell Apoptosis-Associated Reactive Oxygen Species. Scanning 2014, 36, 622–631. [Google Scholar] [CrossRef]
- Shendge, A.K.; Chaudhuri, D.; Basu, T.; Mandal, N. A Natural Flavonoid, Apigenin Isolated from Clerodendrum Viscosum Leaves, Induces G2/M Phase Cell Cycle Arrest and Apoptosis in MCF-7 Cells through the Regulation of P53 and Caspase-Cascade Pathway. Clin. Transl. Oncol. 2021, 23, 718–730. [Google Scholar] [CrossRef]
- Souza, R.P.; Bonfim-Mendonça, P.D.S.; Gimenes, F.; Ratti, B.A.; Kaplum, V.; Bruschi, M.L.; Nakamura, C.V.; Silva, S.O.; Maria-Engler, S.S.; Consolaro, M.E.L. Oxidative Stress Triggered by Apigenin Induces Apoptosis in a Comprehensive Panel of Human Cervical Cancer-Derived Cell Lines. Oxid. Med. Cell Longev. 2017, 2017, 1512745. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Das, J.; Samadder, A.; Boujedaini, N.; Khuda-Bukhsh, A.R. Apigenin-Induced Apoptosis in A375 and A549 Cells through Selective Action and Dysfunction of Mitochondria. Exp. Biol. Med. 2012, 237, 1433–1448. [Google Scholar] [CrossRef]
- Shukla, S.; Gupta, S. Apigenin-Induced Prostate Cancer Cell Death Is Initiated by Reactive Oxygen Species and P53 Activation. Free Radic. Biol. Med. 2008, 44, 1833–1845. [Google Scholar] [CrossRef] [PubMed]
- Valdameri, G.; Trombetta-Lima, M.; Worfel, P.R.; Pires, A.R.A.; Martinez, G.R.; Noleto, G.R.; Cadena, S.M.S.C.; Sogayar, M.C.; Winnischofer, S.M.B.; Rocha, M.E.M. Involvement of Catalase in the Apoptotic Mechanism Induced by Apigenin in HepG2 Human Hepatoma Cells. Chem. Biol. Interact. 2011, 193, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liao, Y.; Li, T.; Zhong, H.; Shan, L.; Yu, P.; Xia, C.; Xu, L. Apigenin Promotes Apoptosis of 4T1 Cells through PI3K/AKT/Nrf2 Pathway and Improves Tumor Immune Microenvironment in Vivo. Toxicol. Res. 2024, 13, tfae011. [Google Scholar] [CrossRef]
- Juan, C.A.; de la Lastra, J.M.P.; Plou, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef]
- Arango, D.; Parihar, A.; Villamena, F.A.; Wang, L.; Freitas, M.A.; Grotewold, E.; Doseff, A.I. Apigenin Induces DNA Damage through the PKCδ-Dependent Activation of ATM and H2AX Causing down-Regulation of Genes Involved in Cell Cycle Control and DNA Repair. Biochem. Pharmacol. 2012, 84, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Krisanapun, C.; Lee, S.H.; Nualsanit, T.; Sams, C.; Peungvicha, P.; Baek, S.J. Molecular Targets of Apigenin in Colorectal Cancer Cells: Involvement of P21, NAG-1 and P53. Eur. J. Cancer 2010, 46, 3365–3374. [Google Scholar] [CrossRef]
- Vargo, M.A.; Voss, O.H.; Poustka, F.; Cardounel, A.J.; Grotewold, E.; Doseff, A.I. Apigenin-Induced-Apoptosis Is Mediated by the Activation of PKCdelta and Caspases in Leukemia Cells. Biochem. Pharmacol. 2006, 72, 681–692. [Google Scholar] [CrossRef]
- Wang, H.; Guo, M.; Wei, H.; Chen, Y. Targeting P53 Pathways: Mechanisms, Structures, and Advances in Therapy. Signal Transduct. Target. Ther. 2023, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Cheung, E.C.; Vousden, K.H. The Role of ROS in Tumour Development and Progression. Nat. Rev. Cancer 2022, 22, 280–297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhao, X.H.; Wang, Z.J. Flavones and Flavonols Exert Cytotoxic Effects on a Human Oesophageal Adenocarcinoma Cell Line (OE33) by Causing G2/M Arrest and Inducing Apoptosis. Food Chem. Toxicol. 2008, 46, 2042–2053. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhao, X.H.; Wang, Z.J. Cytotoxicity of Flavones and Flavonols to a Human Esophageal Squamous Cell Carcinoma Cell Line (KYSE-510) by Induction of G2/M Arrest and Apoptosis. Toxicol. In Vitro 2009, 23, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Masuelli, L.; Benvenuto, M.; Mattera, R.; Di Stefano, E.; Zago, E.; Taffera, G.; Tresoldi, I.; Giganti, M.G.; Frajese, G.V.; Berardi, G.; et al. In Vitro and In Vivo Anti-Tumoral Effects of the Flavonoid Apigenin in Malignant Mesothelioma. Front. Pharmacol. 2017, 8, 373. [Google Scholar] [CrossRef] [PubMed]
- Eun, J.C.; Kim, G.H. Apigenin Induces Apoptosis through a Mitochondria/Caspase-Pathway in Human Breast Cancer MDA-MB-453 Cells. J. Clin. Biochem. Nutr. 2009, 44, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Torkin, R.; Lavoie, J.F.; Kaplan, D.R.; Yeger, H. Induction of Caspase-Dependent, P53-Mediated Apoptosis by Apigenin in Human Neuroblastoma. Mol. Cancer Ther. 2005, 4, 1–11. [Google Scholar] [CrossRef]
- Meng, S.; Zhu, Y.; Li, J.F.; Wang, X.; Liang, Z.; Li, S.Q.; Xu, X.; Chen, H.; Liu, B.; Zheng, X.Y.; et al. Apigenin Inhibits Renal Cell Carcinoma Cell Proliferation. Oncotarget 2017, 8, 19834–19842. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.G.; O’Neill, E. PI3K/Akt-Mediated Regulation of P53 in Cancer. Biochem. Soc. Trans. 2014, 42, 798–803. [Google Scholar] [CrossRef]
- Granato, M.; Gilardini Montani, M.S.; Santarelli, R.; D’Orazi, G.; Faggioni, A.; Cirone, M. Apigenin, by Activating P53 and Inhibiting STAT3, Modulates the Balance between pro-Apoptotic and pro-Survival Pathways to Induce PEL Cell Death. J. Exp. Clin. Cancer Res. 2017, 36, 167. [Google Scholar] [CrossRef]
- Kim, S.H.; Kang, J.G.; Kim, C.S.; Ihm, S.H.; Choi, M.G.; Yoo, H.J.; Lee, S.J. Apigenin Induces C-Myc-Mediated Apoptosis in FRO Anaplastic Thyroid Carcinoma Cells. Mol. Cell Endocrinol. 2013, 369, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Capece, D.; Verzella, D.; Flati, I.; Arboretto, P.; Cornice, J.; Franzoso, G. NF-ΚB: Blending Metabolism, Immunity, and Inflammation. Trends Immunol. 2022, 43, 757–775. [Google Scholar] [CrossRef] [PubMed]
- Hoesel, B.; Schmid, J.A. The Complexity of NF-ΚB Signaling in Inflammation and Cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Gupta, S. Suppression of Constitutive and Tumor Necrosis Factor Alpha-Induced Nuclear Factor (NF)-KappaB Activation and Induction of Apoptosis by Apigenin in Human Prostate Carcinoma PC-3 Cells: Correlation with down-Regulation of NF-KappaB-Responsive Genes. Clin. Cancer Res. 2004, 10, 3169–3178. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Kanwal, R.; Shankar, E.; Datt, M.; Chance, M.R.; Fu, P.; MacLennan, G.T.; Gupta, S. Apigenin Blocks IKKα Activation and Suppresses Prostate Cancer Progression. Oncotarget 2015, 6, 31216–31232. [Google Scholar] [CrossRef] [PubMed]
- Hamacher, R.; Saur, D.; Fritsch, R.; Reichert, M.; Schmid, R.M.; Schneider, G. Casein Kinase II Inhibition Induces Apoptosis in Pancreatic Cancer Cells. Oncol. Rep. 2007, 18, 695–701. [Google Scholar] [CrossRef]
- Pandey, M.; Kaur, P.; Shukla, S.; Abbas, A.; Fu, P.; Gupta, S. Plant Flavone Apigenin Inhibits HDAC and Remodels Chromatin to Induce Growth Arrest and Apoptosis in Human Prostate Cancer Cells: In Vitro and in Vivo Study. Mol. Carcinog. 2012, 51, 952–962. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Bhaskaran, N.; Babcook, M.A.; Fu, P.; MacLennan, G.T.; Gupta, S. Apigenin Inhibits Prostate Cancer Progression in TRAMP Mice via Targeting PI3K/Akt/FoxO Pathway. Carcinogenesis 2014, 35, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Ramaiah, M.J.; Tangutur, A.D.; Manyam, R.R. Epigenetic Modulation and Understanding of HDAC Inhibitors in Cancer Therapy. Life Sci. 2021, 277, 119504. [Google Scholar] [CrossRef]
- Chakrabarti, M.; Banik, N.L.; Ray, S.K. MiR-138 Overexpression Is More Powerful than HTERT Knockdown to Potentiate Apigenin for Apoptosis in Neuroblastoma in Vitro and in Vivo. Exp. Cell Res. 2013, 319, 1575–1585. [Google Scholar] [CrossRef]
- Jayasooriya, R.G.P.T.; Kang, S.H.; Kang, C.H.; Choi, Y.H.; Moon, D.O.; Hyun, J.W.; Chang, W.Y.; Kim, G.Y. Apigenin Decreases Cell Viability and Telomerase Activity in Human Leukemia Cell Lines. Food Chem. Toxicol. 2012, 50, 2605–2611. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Sharma, V.; Verma, V.; Pandey, D.; Yadav, S.K.; Maikhuri, J.P.; Gupta, G. Apigenin Manipulates the Ubiquitin-Proteasome System to Rescue Estrogen Receptor-β from Degradation and Induce Apoptosis in Prostate Cancer Cells. Eur. J. Nutr. 2015, 54, 1255–1267. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Landis-Piwowar, K.R.; Chen, M.S.; Dou, Q.P. Inhibition of Proteasome Activity by the Dietary Flavonoid Apigenin Is Associated with Growth Inhibition in Cultured Breast Cancer Cells and Xenografts. Breast Cancer Res. 2007, 9, R80. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.G.; Wang, L.; Liu, W.J.; Wang, J.F.; Zhao, E.J.; Zhou, F.M.; Ji, X.B.; Wang, L.H.; Xia, Z.K.; Wang, W.; et al. Apigenin Inhibits IL-6 Transcription and Suppresses Esophageal Carcinogenesis. Front. Pharmacol. 2019, 10, 1002. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.J.; Kim, B.S.; Chun, S.Y.; Park, Y.K.; Kang, K.S.; Kwon, T.G. Apoptotic Effects of Genistein, Biochanin-A and Apigenin on LNCaP and PC-3 Cells by P21 through Transcriptional Inhibition of Polo-like Kinase-1. J. Korean Med. Sci. 2011, 26, 1489–1494. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xin, Y.; Diao, Y.; Lu, C.; Fu, J.; Luo, L.; Yin, Z. Synergistic Effects of Apigenin and Paclitaxel on Apoptosis of Cancer Cells. PLoS ONE 2011, 6, e29169. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Song, J.; Hwang, S.; Choi, J.; Song, G.; Lim, W. Apigenin Enhances Apoptosis Induction by 5-Fluorouracil through Regulation of Thymidylate Synthase in Colorectal Cancer Cells. Redox Biol. 2021, 47, 102144. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.Y.; Liang, J.Y.; Guo, X.J.; Liu, L.; Guo, Y.B. 5-Fluorouracil Combined with Apigenin Enhances Anticancer Activity through Mitochondrial Membrane Potential (ΔΨm)-Mediated Apoptosis in Hepatocellular Carcinoma. Clin. Exp. Pharmacol. Physiol. 2015, 42, 146–153. [Google Scholar] [CrossRef]
- Shao, H.; Jing, K.; Mahmoud, E.; Huang, H.; Fang, X.; Yu, C. Apigenin Sensitizes Colon Cancer Cells to Antitumor Activity of ABT-263. Mol. Cancer Ther. 2013, 12, 2640–2650. [Google Scholar] [CrossRef]
- Gao, A.M.; Ke, Z.P.; Wang, J.N.; Yang, J.Y.; Chen, S.Y.; Chen, H. Apigenin Sensitizes Doxorubicin-Resistant Hepatocellular Carcinoma BEL-7402/ADM Cells to Doxorubicin via Inhibiting PI3K/Akt/Nrf2 Pathway. Carcinogenesis 2013, 34, 1806–1814. [Google Scholar] [CrossRef]
- Şirin, N.; Elmas, L.; Seçme, M.; Dodurga, Y. Investigation of Possible Effects of Apigenin, Sorafenib and Combined Applications on Apoptosis and Cell Cycle in Hepatocellular Cancer Cells. Gene 2020, 737, 144428. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Khan, M.A.; Mishra, D.; Goel, A.; Ansari, M.A.; Akhtar, K.; Siddique, H.R. Apigenin Enhances Sorafenib Anti-Tumour Efficacy in Hepatocellular Carcinoma. Transl. Oncol. 2024, 43, 101920. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, S.; Davis, K.A.; Choudhury, S.R.; Deeconda, A.; Banik, N.L.; Ray, S.K. Bcl-2 Inhibitor and Apigenin Worked Synergistically in Human Malignant Neuroblastoma Cell Lines and Increased Apoptosis with Activation of Extrinsic and Intrinsic Pathways. Biochem. Biophys. Res. Commun. 2009, 388, 705–710. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, T.; Shi, Z.; Hu, C.; Li, Q.; Sun, C. Synergism Antiproliferative Effects of Apigenin and Naringenin in NSCLC Cells. Molecules 2023, 28, 4947. [Google Scholar] [CrossRef] [PubMed]
- Smiljkovic, M.; Stanisavljevic, D.; Stojkovic, D.; Petrovic, I.; Vicentic, J.M.; Popovic, J.; Golic Grdadolnik, S.; Markovic, D.; Sanković-Babić, S.; Glamoclija, J.; et al. Apigenin-7-O-Glucoside versus Apigenin: Insight into the Modes of Anticandidal and Cytotoxic Actions. EXCLI J. 2017, 16, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Bhosale, P.B.; Abusaliya, A.; Kim, H.H.; Ha, S.E.; Park, M.Y.; Jeong, S.H.; Vetrivel, P.; Heo, J.D.; Kim, J.A.; Won, C.K.; et al. Apigetrin Promotes TNFα-Induced Apoptosis, Necroptosis, G2/M Phase Cell Cycle Arrest, and ROS Generation through Inhibition of NF-ΚB Pathway in Hep3B Liver Cancer Cells. Cells 2022, 11, 2734. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Lu, N.N.; Feng, L. Apigetrin Inhibits Gastric Cancer Progression through Inducing Apoptosis and Regulating ROS-Modulated STAT3/JAK2 Pathway. Biochem. Biophys. Res. Commun. 2018, 498, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.M.; Ma, R.H.; Ni, Z.J.; Thakur, K.; Cespedes-Acuña, C.L.; Jiang, L.; Wei, Z.J. Apigenin 7-O-Glucoside Promotes Cell Apoptosis through the PTEN/PI3K/AKT Pathway and Inhibits Cell Migration in Cervical Cancer HeLa Cells. Food Chem. Toxicol. 2020, 146, 111843. [Google Scholar] [CrossRef]
- Najafipour, R.; Momeni, A.M.; Mirmazloomi, Y.; Moghbelinejad, S. Vitexin Induces Apoptosis in MCF-7 Breast Cancer Cells through the Regulation of Specific MiRNAs Expression. Int. J. Mol. Cell Med. 2022, 11, 197–206. [Google Scholar] [CrossRef]
- Czemplik, M.; Mierziak, J.; Szopa, J.; Kulma, A. Flavonoid C-Glucosides Derived from Flax Straw Extracts Reduce Human Breast Cancer Cell Growth In Vitro and Induce Apoptosis. Front. Pharmacol. 2016, 7, 282. [Google Scholar] [CrossRef]
- Bhardwaj, M.; Cho, H.J.; Paul, S.; Jakhar, R.; Khan, I.; Lee, S.J.; Kim, B.Y.; Krishnan, M.; Khaket, T.P.; Lee, H.G.; et al. Vitexin Induces Apoptosis by Suppressing Autophagy in Multi-Drug Resistant Colorectal Cancer Cells. Oncotarget 2017, 9, 3278–3291. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Li, D.; Chen, H.; Zhang, J.; Jin, X. Vitexin Induces G2/M-phase Arrest and Apoptosis via Akt/MTOR Signaling Pathway in Human Glioblastoma Cells. Mol. Med. Rep. 2018, 17, 4599–4604. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Jiang, Q.; Liu, H.; Luo, S. Vitexin Induces Apoptosis through Mitochondrial Pathway and PI3K/Akt/MTOR Signaling in Human Non-Small Cell Lung Cancer A549 Cells. Biol. Res. 2019, 52, 7. [Google Scholar] [CrossRef]
- An, F.; Wang, S.; Tian, Q.; Zhu, D. Effects of Orientin and Vitexin from Trollius Chinensis on the Growth and Apoptosis of Esophageal Cancer EC-109 Cells. Oncol. Lett. 2015, 10, 2627–2633. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Chien, Y.S.; Chiu, T.H.; Huang, W.W.; Lu, C.C.; Chiang, J.H.; Yang, J.S. Apoptosis Triggered by Vitexin in U937 Human Leukemia Cells via a Mitochondrial Signaling Pathway. Oncol. Rep. 2012, 28, 1883–1888. [Google Scholar] [CrossRef]
- Huang, J.; Zhou, Y.; Zhong, X.; Su, F.; Xu, L. Effects of Vitexin, a Natural Flavonoid Glycoside, on the Proliferation, Invasion, and Apoptosis of Human U251 Glioblastoma Cells. Oxid. Med. Cell Longev. 2022, 2022, 3129155. [Google Scholar] [CrossRef] [PubMed]
- He, J.D.; Wang, Z.; Li, S.P.; Xu, Y.J.; Yu, Y.; Ding, Y.J.; Yu, W.L.; Zhang, R.X.; Zhang, H.M.; Du, H.Y. Vitexin Suppresses Autophagy to Induce Apoptosis in Hepatocellular Carcinoma via Activation of the JNK Signaling Pathway. Oncotarget 2016, 7, 84520–84532. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Guan, X.; Hou, R.; Zhang, X.; Guo, F.; Zhang, Z.; Hua, C. Vitexin Attenuates Epithelial Ovarian Cancer Cell Viability and Motility in Vitro and Carcinogenesis in Vivo via P38 and ERK1/2 Pathways Related VEGFA. Ann. Transl. Med. 2020, 8, 1139. [Google Scholar] [CrossRef]
- Wang, L.; Klionsky, D.J.; Shen, H.M. The Emerging Mechanisms and Functions of Microautophagy. Nat. Rev. Mol. Cell Biol. 2023, 24, 186–203. [Google Scholar] [CrossRef]
- Kitada, M.; Koya, D. Autophagy in Metabolic Disease and Ageing. Nat. Rev. Endocrinol. 2021, 17, 647–661. [Google Scholar] [CrossRef]
- Deleyto-Seldas, N.; Efeyan, A. The MTOR-Autophagy Axis and the Control of Metabolism. Front. Cell Dev. Biol. 2021, 9, 655731. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Dominguez, R.; Perez-Medina, M.; Lopez-Gonzalez, J.S.; Galicia-Velasco, M.; Aguilar-Cazares, D. The Double-Edge Sword of Autophagy in Cancer: From Tumor Suppression to Pro-Tumor Activity. Front. Oncol. 2020, 10, 578418. [Google Scholar] [CrossRef] [PubMed]
- Naponelli, V.; Modernelli, A.; Bettuzzi, S.; Rizzi, F. Roles of Autophagy Induced by Natural Compounds in Prostate Cancer. BioMed Res. Int. 2015, 2015, 121826. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, S.; Ma, B. Autophagy and Autophagy-Related Proteins in Cancer. Mol. Cancer 2020, 19, 12. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.S.; Tian, D.; Liao, X.; Zhang, Y.; Xiao, J.; Chen, W.; Liu, Q.; Chen, Y.; Li, D.; Zhu, L.; et al. Apigenin Combined With Gefitinib Blocks Autophagy Flux and Induces Apoptotic Cell Death Through Inhibition of HIF-1α, c-Myc, p-EGFR, and Glucose Metabolism in EGFR L858R+T790M-Mutated H1975 Cells. Front. Pharmacol. 2019, 10, 260. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Pi, C.; Wang, G. Inhibition of PI3K/Akt/MTOR Pathway by Apigenin Induces Apoptosis and Autophagy in Hepatocellular Carcinoma Cells. Biomed. Pharmacother. 2018, 103, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.M.; Chen, H.H.; Lin, C.A.; Wu, H.C.; Sheu, J.J.C.; Chen, H.J. Apigenin-Induced Lysosomal Degradation of β-Catenin in Wnt/β-Catenin Signaling. Sci. Rep. 2017, 7, 372. [Google Scholar] [CrossRef]
- Kim, T.W.; Lee, H.G. Apigenin Induces Autophagy and Cell Death by Targeting EZH2 under Hypoxia Conditions in Gastric Cancer Cells. Int. J. Mol. Sci. 2021, 22, 13455. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Meng, Z.; Chen, Y.; Yu, L.; Gao, B.; Zheng, Y.; Guan, S. Apigenin Induced Autophagy and Stimulated Autophagic Lipid Degradation. Food Funct. 2020, 11, 9208–9215. [Google Scholar] [CrossRef]
- Zeng, J.; Xie, H.; Zhang, Z.L.; Li, Z.X.; Shi, L.; Wu, K.Y.; Zhou, Y.; Tian, Z.; Zhang, Y.; Zhou, W.; et al. Apigenin Regulates the Migration, Invasion, and Autophagy of Hepatocellular Carcinoma Cells by Downregulating YAP. Neoplasma 2022, 69, 292–302. [Google Scholar] [CrossRef]
- Janda, E.; Martino, C.; Riillo, C.; Parafati, M.; Lascala, A.; Mollace, V.; Boutin, J.A. Apigenin and Luteolin Regulate Autophagy by Targeting NRH-Quinone Oxidoreductase 2 in Liver Cells. Antioxidants 2021, 10, 776. [Google Scholar] [CrossRef] [PubMed]
- Gilardini Montani, M.S.; Cecere, N.; Granato, M.; Romeo, M.A.; Falcinelli, L.; Ciciarelli, U.; D’orazi, G.; Faggioni, A.; Cirone, M. Mutant P53, Stabilized by Its Interplay with HSP90, Activates a Positive Feed-Back Loop Between NRF2 and P62 That Induces Chemo-Resistance to Apigenin in Pancreatic Cancer Cells. Cancers 2019, 11, 703. [Google Scholar] [CrossRef]
- Lascala, A.; Martino, C.; Parafati, M.; Salerno, R.; Oliverio, M.; Pellegrino, D.; Mollace, V.; Janda, E. Analysis of Proautophagic Activities of Citrus Flavonoids in Liver Cells Reveals the Superiority of a Natural Polyphenol Mixture over Pure Flavones. J. Nutr. Biochem. 2018, 58, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Kayacan, S.; Yilancioglu, K.; Akdemir, A.S.; Kaya-Dagistanli, F.; Melikoglu, G.; Ozturk, M. Synergistic Effect of Apigenin and Curcumin on Apoptosis, Paraptosis and Autophagy-Related Cell Death in HeLa Cells. Anticancer Res. 2021, 41, 1271–1282. [Google Scholar] [CrossRef]
- Ghazy, E.; Taghi, H.S. The Autophagy-Inducing Mechanisms of Vitexin, Cinobufacini, and Physalis Alkekengi Hydroalcoholic Extract against Breast Cancer in Vitro and in Vivo. J. Gastrointest. Cancer 2022, 53, 592–596. [Google Scholar] [CrossRef]
- Battaglia, A.M.; Chirillo, R.; Aversa, I.; Sacco, A.; Costanzo, F.; Biamonte, F. Ferroptosis and Cancer: Mitochondria Meet the “Iron Maiden” Cell Death. Cells 2020, 9, 1505. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, C.; Zhao, Y.; Gao, G. Mitochondria Regulation in Ferroptosis. Eur. J. Cell Biol. 2020, 99, 151058. [Google Scholar] [CrossRef]
- Stepanić, V.; Kučerová-Chlupáčová, M. Review and Chemoinformatic Analysis of Ferroptosis Modulators with a Focus on Natural Plant Products. Molecules 2023, 28, 475. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Schorpp, K.; Jin, J.; Yozwiak, C.E.; Hoffstrom, B.G.; Decker, A.M.; Rajbhandari, P.; Stokes, M.E.; Bender, H.G.; Csuka, J.M.; et al. Transferrin Receptor Is a Specific Ferroptosis Marker. Cell Rep. 2020, 30, 3411–3423. [Google Scholar] [CrossRef]
- Li, D.; Li, Y. The Interaction between Ferroptosis and Lipid Metabolism in Cancer. Signal Transduct. Target. Ther. 2020, 5, 108. [Google Scholar] [CrossRef]
- Mou, Y.; Wang, J.; Wu, J.; He, D.; Zhang, C.; Duan, C.; Li, B. Ferroptosis, a New Form of Cell Death: Opportunities and Challenges in Cancer. J. Hematol. Oncol. 2019, 12, 34. [Google Scholar] [CrossRef] [PubMed]
- Adham, A.N.; Hegazy, M.E.F.; Naqishbandi, A.M.; Efferth, T. Induction of Apoptosis, Autophagy and Ferroptosis by Thymus Vulgaris and Arctium Lappa Extract in Leukemia and Multiple Myeloma Cell Lines. Molecules 2020, 25, 5016. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Rong, G.; Liu, Y.; Huang, W.; He, D.; Lu, R. Delivery of Apigenin-Loaded Magnetic Fe2O3/Fe3O4@mSiO2 Nanocomposites to A549 Cells and Their Antitumor Mechanism. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 120, 111719. [Google Scholar] [CrossRef] [PubMed]
- Ketelut-Carneiro, N.; Fitzgerald, K.A. Apoptosis, Pyroptosis, and Necroptosis-Oh My! The Many Ways a Cell Can Die. J. Mol. Biol. 2022, 434, 167378. [Google Scholar] [CrossRef] [PubMed]
- Ai, Y.; Meng, Y.; Yan, B.; Zhou, Q.; Wang, X. The Biochemical Pathways of Apoptotic, Necroptotic, Pyroptotic, and Ferroptotic Cell Death. Mol. Cell 2024, 84, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Fan, Z.; Luo, G.; Yang, C.; Huang, Q.; Fan, K.; Cheng, H.; Jin, K.; Ni, Q.; Yu, X.; et al. The Role of Necroptosis in Cancer Biology and Therapy. Mol. Cancer 2019, 18, 100. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wan, P.; Choksi, S.; Liu, Z.G. Necroptosis and Tumor Progression. Trends Cancer 2022, 8, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Park, K.S.; Nam, H.S.; Cho, M.K.; Lee, S.H. Apigenin Causes Necroptosis by Inducing ROS Accumulation, Mitochondrial Dysfunction, and ATP Depletion in Malignant Mesothelioma Cells. Korean J. Physiol. Pharmacol. 2020, 24, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Warkad, M.S.; Kim, C.H.; Kang, B.G.; Park, S.H.; Jung, J.S.; Feng, J.H.; Inci, G.; Kim, S.C.; Suh, H.W.; Lim, S.S.; et al. Metformin-Induced ROS Upregulation as Amplified by Apigenin Causes Profound Anticancer Activity While Sparing Normal Cells. Sci. Rep. 2021, 11, 14002. [Google Scholar] [CrossRef]
- Hu, G.; Li, J.; Zeng, Y.; Liu, L.; Yu, Z.; Qi, X.; Liu, K.; Yao, H. The Anoikis-Related Gene Signature Predicts Survival Accurately in Colon Adenocarcinoma. Sci. Rep. 2023, 13, 13919. [Google Scholar] [CrossRef]
- Taddei, M.L.; Giannoni, E.; Fiaschi, T.; Chiarugi, P. Anoikis: An Emerging Hallmark in Health and Diseases. J. Pathol. 2012, 226, 380–393. [Google Scholar] [CrossRef] [PubMed]
- Sattari Fard, F.; Jalilzadeh, N.; Mehdizadeh, A.; Sajjadian, F.; Velaei, K. Understanding and Targeting Anoikis in Metastasis for Cancer Therapies. Cell Biol. Int. 2023, 47, 683–698. [Google Scholar] [CrossRef] [PubMed]
- Adeshakin, F.O.; Adeshakin, A.O.; Afolabi, L.O.; Yan, D.; Zhang, G.; Wan, X. Mechanisms for Modulating Anoikis Resistance in Cancer and the Relevance of Metabolic Reprogramming. Front. Oncol. 2021, 11, 626577. [Google Scholar] [CrossRef] [PubMed]
- Hasnat, M.A.; Pervin, M.; Lim, J.H.; Lim, B.O. Apigenin Attenuates Melanoma Cell Migration by Inducing Anoikis through Integrin and Focal Adhesion Kinase Inhibition. Molecules 2015, 20, 21157–21166. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.M.; Meng, D.; Fang, J. Apigenin Inhibited Migration and Invasion of Human Ovarian Cancer A2780 Cells through Focal Adhesion Kinase. Carcinogenesis 2008, 29, 2369–2376. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Chen, W.K.; Wang, C.J.; Lin, W.L.; Tseng, T.H. Apigenin Inhibits HGF-Promoted Invasive Growth and Metastasis Involving Blocking PI3K/Akt Pathway and Beta 4 Integrin Function in MDA-MB-231 Breast Cancer Cells. Toxicol. Appl. Pharmacol. 2008, 226, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Ma, J.; Zhu, H.Y.; Zhang, X.H.; Du, Z.Y.; Xu, Y.J.; Yu, X.D. Apigenin Inhibits Proliferation and Induces Apoptosis in Human Multiple Myeloma Cells through Targeting the Trinity of CK2, Cdc37 and Hsp90. Mol. Cancer 2011, 10. [Google Scholar] [CrossRef]
- Tavsan, Z.; Kayali, H.A. Flavonoids Showed Anticancer Effects on the Ovarian Cancer Cells: In-volvement of Reactive Oxygen Species, Apoptosis, Cell Cycle and Invasion. Biomed. Pharmacother. 2019, 116. [Google Scholar] [CrossRef]
- Abid, R.; Ghazanfar, S.; Farid, A.; Sulaman, S.M.; Idrees, M.; Amen, R.A.; Muzammal, M.; Shahzad, M.K.; Mohamed, M.O.; Khaled, A.A.; et al. Pharmacological Properties of 4′,5,7-Trihydroxyflavone (Apigenin) and Its Impact on Cell Signaling Pathways. Molecules 2022, 27, 4304. [Google Scholar] [CrossRef]
- Zhou, Y.; Yu, Y.; Lv, H.; Zhang, H.; Liang, T.; Zhou, G.; Huang, L.; Tian, Y.; Liang, W. Apigenin in Cancer Therapy: From Mechanism of Action to Nano-Therapeutic Agent. Food Chem. Toxicol. 2022, 168, 113385. [Google Scholar] [CrossRef]
- Gates, M.A.; Tworoger, S.S.; Hecht, J.L.; De Vivo, I.; Rosner, B.; Hankinson, S.E. A Prospective Study of Dietary Flavonoid Intake and Incidence of Epithelial Ovarian Cancer. Int. J. Cancer 2007, 121, 2225–2232. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lee, I.M.; Zhang, S.M.; Blumberg, J.B.; Buring, J.E.; Sesso, H.D. Dietary Intake of Selected Flavonols, Flavones, and Flavonoid-Rich Foods and Risk of Cancer in Middle-Aged and Older Women. Am. J. Clin. Nutr. 2009, 89, 905–912. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Concentration (µg/g) |
|---|---|
| Dried parsley | 45,035 |
| Dried chamomille flower | 3000–5000 |
| Parsley | 2154.6 |
| Celery seed | 786.5 |
| Vinespinach | 622 |
| Chinese celery | 240.2 |
| Kumquats | 218.7 |
| Celery | 191 |
| Dried oregano | 177.1 |
| Artichoke | 74.8 |
| Juniper berries | 72.6 |
| Peppermint | 53.9 |
| Types of Cancer | Cells/Animal Model | Molecular Pathway/Protein | Ref. | |
|---|---|---|---|---|
| Apoptosis | bladder | T-24 | ↑Bax/Bcl-2; ↑Bad; ↑Bak; ↑caspase-9, ↑caspase-3; ↑caspase-7; ↑c-PARP | [101,102] |
| RT112 | ↑c-PARP; ↑fragmented DNA | [117] | ||
| breast | SK-BR-3 | ↑p53; ↑Bax; ↑Cyt c; ↑LDH | [152] | |
| BT-474, MDA-MB-453, MCF-7, T47-D, HBL-100 | ↑Cyt c; ↑caspase-3; ↑fragmentated DNA | [126] | ||
| T47D, MDA-MB-231 | ↓PI3K/Akt/mTOR; ↑Bax/Bcl2 | [127] | ||
| MDA-MB-231/xenograft | ↓proteasome; ↑Bax; ↑c-PARP | [193] | ||
| MCF-7, Hs578T, MDA-MB-231 | ↓PI3K/Akt; ↑FOXO3a; | [119] | ||
| MCF-7 | ↑ROS | [160] | ||
| 4T1/xenografts | ↓PI3K/Akt/Nrf2 | [166] | ||
| MCF-7 | ↑ROS; ↑p53; ↑Bax/Bcl-2; ↑c-PARP; ↑caspases | [161] | ||
| SKBR3; BT-474; MDA-MB-453 | ↓JAK2/STAT3/VEGF; ↑c-PARP; ↑caspase-8; ↑caspase-3 | [148,149,150,176] | ||
| MDA-MB-453 | ↓HER2; ↓JAK1; ↓STAT3; ↑p53 | [150,176] | ||
| cervical | HeLa, SiHa, CaSki, C33A | ↑ROS | [162] | |
| HeLa | ↑Fas/APO-1; ↑caspase-3; ↓Bcl-2 | [107] | ||
| HeLa, C33A | ↑Bax/Bcl-2 | [45] | ||
| colon | HCT116 | ↑caspase-8; ↑caspase-9; ↑caspase-3; ↑c-PARP | [89] | |
| HT-29/xenografts | ↓mTOR/PI3K/Akt; ↑Bax/Bcl-2; ↓CCND1 | [128,129] | ||
| HT29, DLD-1, COLO320 and HCT116 | ↓Mcl-1; ↓Bcl-xL; ↓STAT3 | [153] | ||
| HCT-116, SW480, HT-29, LoVo/xenografts | ↑NAG-1; ↑p53; ↑PKCδ; ↑ATM | [169] | ||
| esophageal | OE33, KYSE-510 | ↑caspase-9; ↑caspase-3; ↑p53 | [173,174] | |
| Eca-109, Kyse-30 | ↓IL-6; ↑c-PARP; ↑caspase-8 | [194] | ||
| gastric | HGC-27, SGC-7901 | ↑Bax/Bcl-2; ↓MMP; ↑caspase-3 | [124] | |
| head and neck | CAL-27, Scc-15 and FaDu | ↓Akt; ↓Erk1/2; ↓EGFR; ↓ErbB2 | [144] | |
| SCC25 | ↑TNF-R; ↑TRAIL-R; ↓Bcl-2; ↑caspase-3 ↑ROS; ↓GSH | [145] | ||
| leukemia | K562 | ↑c-PARP; ↑ DNA fragmentation | [117] | |
| HL60 | ↓JAK/STAT; ↓PI3K/PKB; ↑caspase-8; ↑caspase-7 | [156,157] | ||
| THP-1 | ↑DNA damage; ↑ATM; ↑H2AX; ↑p38; ↑PKCδ | [168] | ||
| THP-1, U937, HL60, Jurkat, K562 | ↑caspase-3; ↑PKCδ | [170] | ||
| U937, THP-1 and HL60 | ↓telomerase; ↑caspase | [191] | ||
| liver | HepG2 | ↑ROS; ↓catalase; ↓GSH | [165] | |
| HepG2 | ↓PI3K/Akt/mTOR; ↑caspase-3; ↑caspase-9; ↑c-PARP; ↑Bax/Bcl-2 | [226] | ||
| BEL-7402/xenografts | ↓PI3K/Akt/Nrf2 | [200] | ||
| lung | H460, A549 | ↑c-PARP; ↑Bax/Bcl-2; ↑caspase 3 | [121,122,123] | |
| A549 | ↑ROS; ↑DNA fragmentation; ↑Cyt c; ↑AIF; ↑Endo G ↑Bax/Bcl-2, ↑caspase 3, ↑caspase-9, ↑PARP | [163] | ||
| A549, H1299 | ↑Bad; ↑Bax/Bcl-2, ↑Bcl-xL; ↑DR4; ↑DR5; ↑TRAIL | [140] | ||
| NCI-H23 | ↑PTEN; ↑Bax/Bcl-2; ↑caspase 3 | [130] | ||
| lymphoma | U2932, OCI-LY10 | ↓PI3K/Akt/mTOR; ↓p-mTOR, ↓p-AKT, p-↓IKK, ↓p-p65 | [131] | |
| BC3, BCBL-1 | ↓Akt/mTOR; ↓ROS; ↑p53; ↑catalase; ↑c-PARP | [183] | ||
| melanoma | A375 | ↑ ROS; ↑Cyt c; ↑Bax/Bcl-2, ↑caspase 3, ↑caspase 9, ↑PARP; ↓MMP; ↑DNA fragmentation | [163] | |
| A375, C8161 | ↑caspase-3 ↑c-PARP ↓(p)-ERK1/2, ↓Akt; ↓mTOR | [132] | ||
| mesothelioma | MM-F1, MM-B1, H-Meso-1/xenografts | ↑Bax/Bcl-2; ↑p53, ↑caspase 8; ↑caspase-9, ↑c-PARP; ↓AKT, ↓c-Jun; ↓NF-κB | [177] | |
| multiple myeloma | U266, RPMI 8226 | ↓Mcl-1, ↓Bcl-2, ↓Bcl-xL, ↓XIAP, ↓Survivin; ↓CK2; ↓Hsp90/Cdc37/Cdk4 | [257] | |
| NCI-H929 | ↓STAT1/COX-2/iNOS; ↑ROS; ↓MMP | [244] | ||
| neuroblastoma | NUB-7, LAN-5/xenografts | ↑p53; ↑Bax; ↑caspase-3 | [180] | |
| ovarian | A2780, OVCAR-3, SKOV-3 | ↑ROS; ↓ROS; ↑caspase | [258] | |
| SKOV | ↑caspase-9; ↑caspase-3; ↑Bcl-2 | [85] | ||
| prostate | 22Rv1/xenografts | ↑p53; ↑ROS; ↑Cyt c; ↑caspase-3, ↑Bax/Bcl-2 | [164] | |
| PWR-1E, LNCaP, PC-3, DU145 | ↑ROS; ↑caspase 3; ↑caspase-7; ↑caspase-8; ↑caspase-9; ↓c-IAP2; ↓MMP: ↑Cyt c | [140] | ||
| LNCaP | ↑Bax/Bcl-2 | [119] | ||
| PC-3, DU145 | ↓XIAP; ↓c-IAP1; ↓c-IAP2; survivin; ↑Bax/Bcl-2; ↑Cyt c; ↓HDAC1 | [125] | ||
| PC-3, 22Rv1/xenografts | ↓HDAC1; ↓HDAC3; ↑Bax/Bcl-2 | [187] | ||
| LNCaP, PC-3 | ↑p21; ↓PLK-1 | [195] | ||
| DU-145, PC-3/xenografts | ↓proteasome; ↑ER-β; ↑caspase-8; ↑caspase-3 | [192] | ||
| DU-145, PC-3 | ↓IFG-IR/PI3K/MAPK; ↓IGF-IR; ↓Akt; ↓GSK-3β; ↓cyclin D1; ↑p27/kip1 | [133,134] | ||
| PC3 | ↓Akt; Bad | [132] | ||
| PC-3, 22Rv1/TRAMP mice | ↓NF-ĸB; ↓IKKβ | [184,185] | ||
| DU145 | ↓ANT2 | [139] | ||
| pancreas | MiaPaCa2, DanG | ↓NF-ĸB; ↓CK2; ↓p-Bid; ↑caspase-8 | [186] | |
| BxPC-3, PANC-1 | ↑Cyt c; ↑caspase 9; ↑caspase 3 | [120] | ||
| renal | ACHN/xenografts | ↑p53; ↑Bax; ↑caspase-9; ↑caspase-3 | [178] | |
| thyroid | FRO | ↑c-Myc; ↑p38; ↑p53 | [181] | |
| Autophagy | colon | HCT-116, SW480 | ↓β-catenin; ↓Akt/mTOR | [227] |
| gastric | AGS, SNU-638 | ↑PERK-ATF4-CHOP; ↑GRP78; ↓HIF-1α ↓Ezh2; ↑Atg5; ↑LC3; ↑AMPK and ↑ULK1; ↓p-mTOR; ↓p62 | [228] | |
| liver | SMMC-7721, SK-HEP1 | ↑LC3-II; ↑ULK1; ↓SQSTM1/p62; ↓YAP | [230] | |
| HepG2 | ↑LC3-II/LC3-I; ↑AMPK; ↑NQO2 | [231] | ||
| pancreas | AsPC-1 | ↑AIF; ↑p62; ↑LC3-II | [232] | |
| thyroid | BCPAP | ↑LC3-II; ↑beclin-1; AVO accumulation | [90] | |
| Ferroptosis | myeloma | NCI-H929 | ↓MMP; ↑LC3-II; ↑beclin-1; ↑ROS | [242] |
| HEK293 | ↓MMP; ↑caspase-3; ↑caspase-9; p38; ↑LC3-II; ↑beclin-1; ↑ROS; ↓Akt; ↓COX-2; ↓iNOS | [158] | ||
| lung | A549 | ↑ROS; ↓GPX4; ↓SOD; ↑Bax; ↑caspase-3; ↑caspase-8 | [243] | |
| Necroptosis | mesothelioma | MSTO-211H, H-2452 | ↑p-MLKL; ↑p-RIP3 | [248] |
| pancreas | AsPC-1 | ↑MLKL; ↑p-MLKL; ↑RIP3; ↑p-RIP3 | [249] | |
| Anoikis | breast | MDA-MB-231/xenografts | ↓PI3K/Akt; ↓integrin β4 | [256] |
| cervical | C33A | ↓PI3K/Akt; ↓FAK | [45] | |
| melanoma | A2058, A375 | ↑caspase-3; ↑c-PARP; ↓FAK; ↓ERK1/2; ↓Integrin | [254] | |
| ovarian | A2780/xenografts | ↓FAK | [255] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naponelli, V.; Rocchetti, M.T.; Mangieri, D. Apigenin: Molecular Mechanisms and Therapeutic Potential against Cancer Spreading. Int. J. Mol. Sci. 2024, 25, 5569. https://doi.org/10.3390/ijms25105569
Naponelli V, Rocchetti MT, Mangieri D. Apigenin: Molecular Mechanisms and Therapeutic Potential against Cancer Spreading. International Journal of Molecular Sciences. 2024; 25(10):5569. https://doi.org/10.3390/ijms25105569
Chicago/Turabian StyleNaponelli, Valeria, Maria Teresa Rocchetti, and Domenica Mangieri. 2024. "Apigenin: Molecular Mechanisms and Therapeutic Potential against Cancer Spreading" International Journal of Molecular Sciences 25, no. 10: 5569. https://doi.org/10.3390/ijms25105569
APA StyleNaponelli, V., Rocchetti, M. T., & Mangieri, D. (2024). Apigenin: Molecular Mechanisms and Therapeutic Potential against Cancer Spreading. International Journal of Molecular Sciences, 25(10), 5569. https://doi.org/10.3390/ijms25105569