

Unburned Tobacco Smoke Affects Neuroinflammation-Related Pathways in the Rat Mesolimbic System

 ,
,  , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
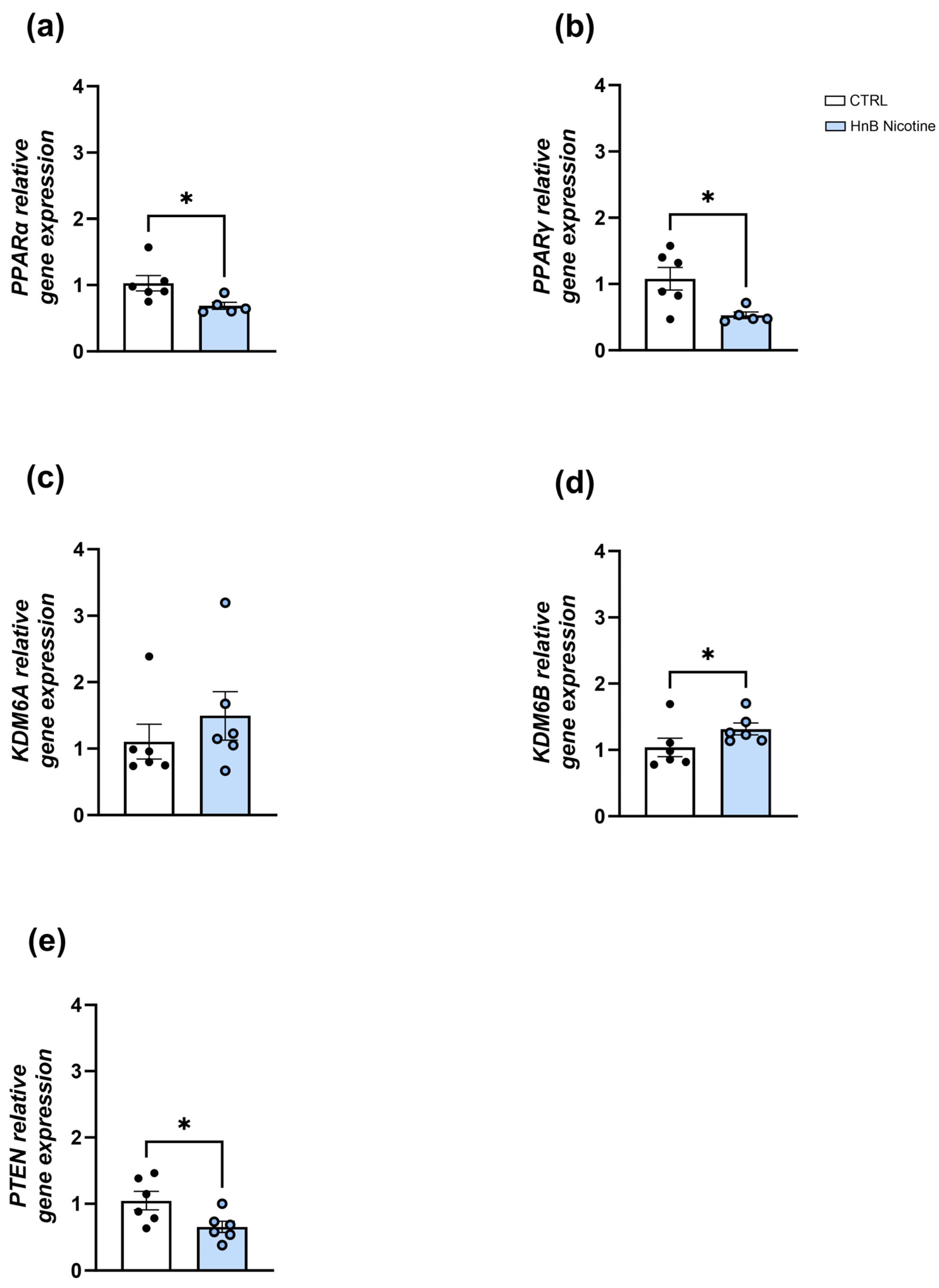
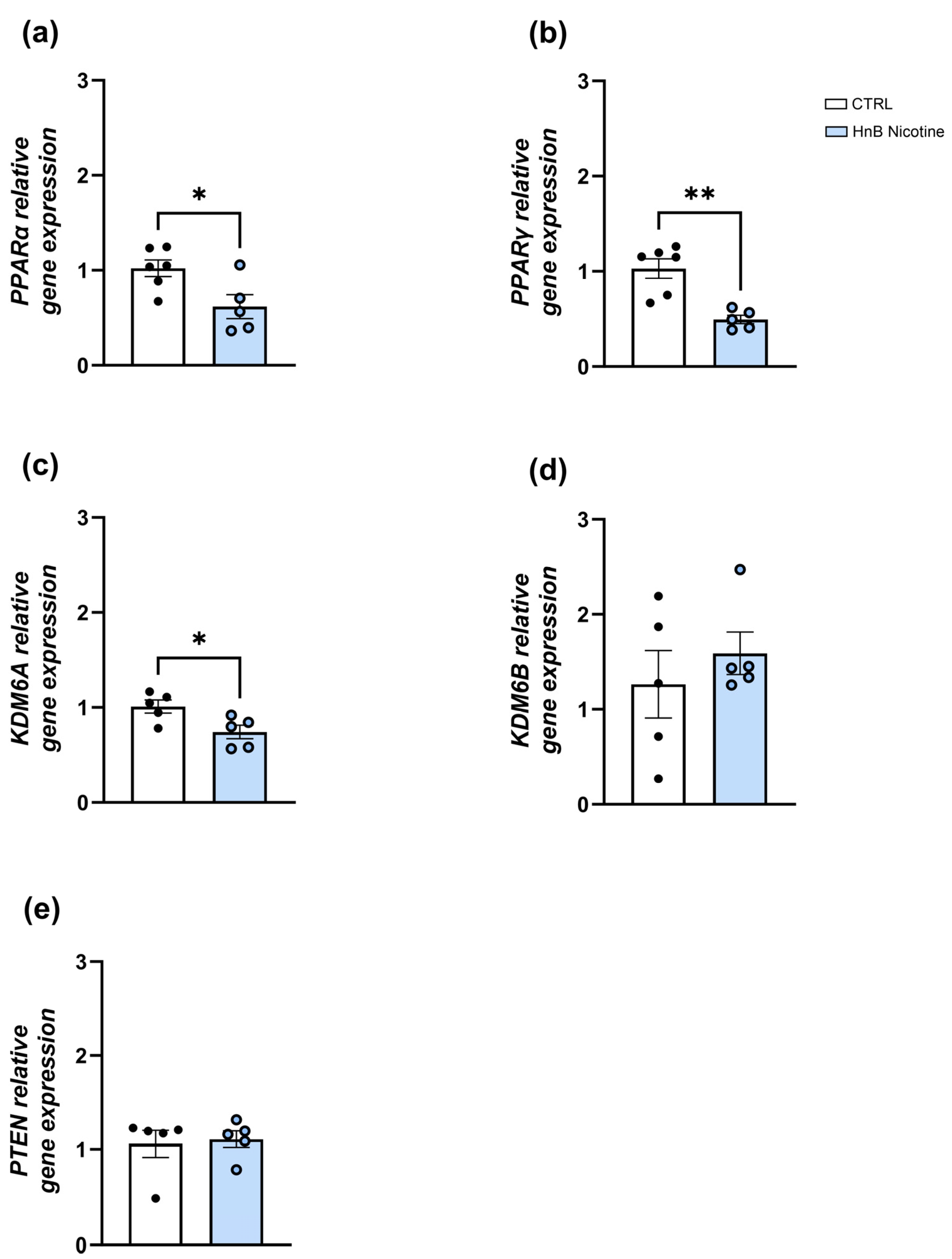
2.1. Gene Expression Analysis
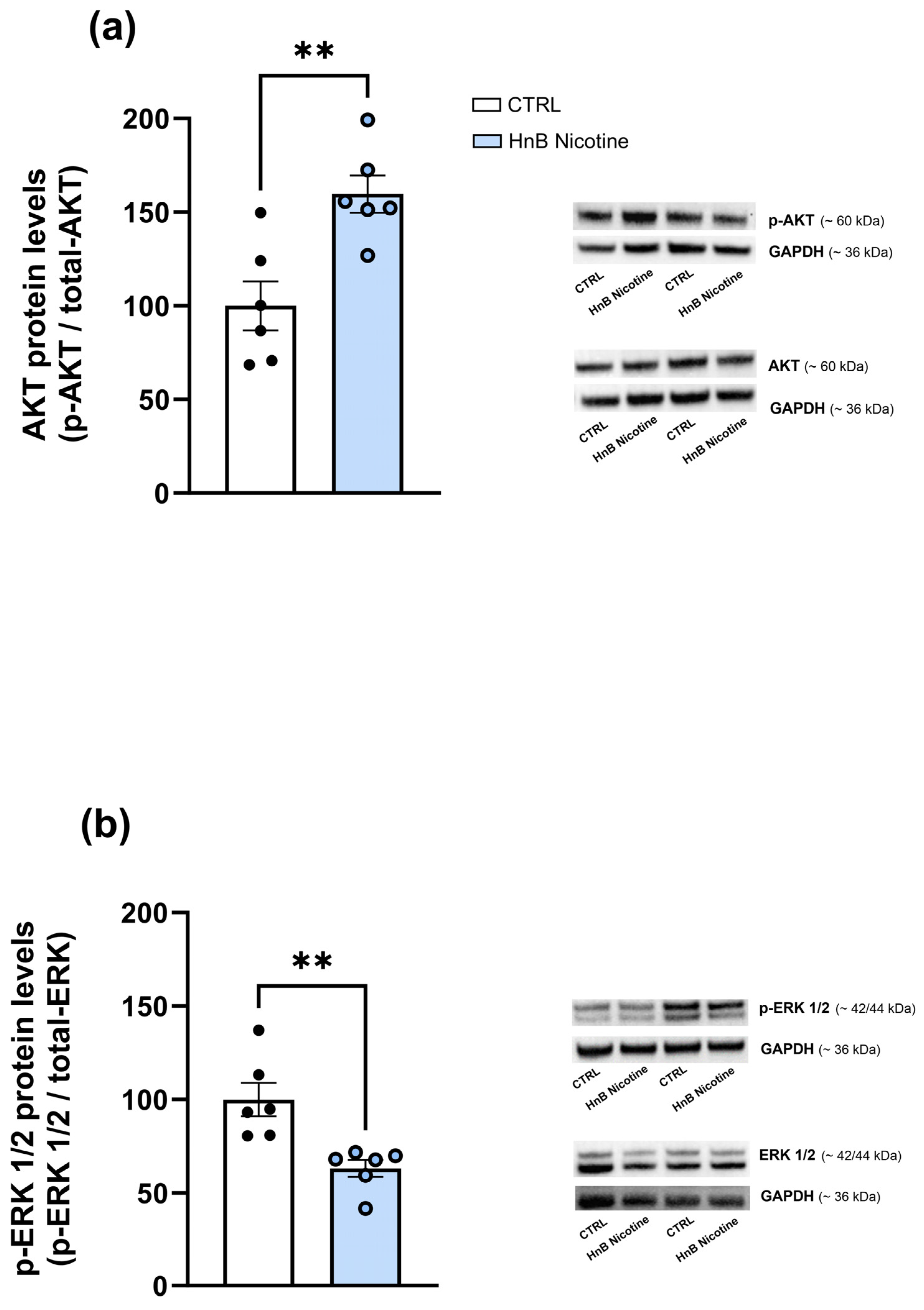
2.2. Protein Analysis
3. Discussion
4. Materials and Methods
4.1. Animal Model and Nicotine Exposure
4.2. RNA Extraction and Gene Expression Analysis Using Real-Time qPCR
- −
- GAPDH Forward 5′-AGACAGCCGCATCTTCTTGT-3′;Reverse 5′-CTTGCCGTGGGTAGAGTCAT-3′;
- −
- KDM6A Forward 5′-TTTGGTCTACTTCCATTACAATGCA-3′;Reverse 5′-AAGCCCAAGTCGTAAATGAATTTC-3′;
- −
- KDM6B Forward 5′-ACCGCCTGCGTGCCTTAC-3′;Reverse 5′-GTGTTGCTGCTGCTGCTACTG-3′;
- −
- PTEN Forward 5′-TGGATTCGACTTAGACTTGACCT-3′;Reverse 5′-GCGGTGTCATAATGTCTCTCAG-3′;
- −
- PPARα Forward 5′-TGGAGTCCACGCATGTGAAG-3′;Reverse 5′-TTGTCGTACGCCAGCTTTAGC-3′;
- −
- PPARγ Forward 5′-CTGTTCGTACGCCAGCTTTAGC-3′;Reverse 5′-GCTCATATCTGTCTCCGTCTTCTT-3′;
4.3. SDS-Page and Immunoblotting
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benowitz, N.L. Nicotine Addiction. N. Engl. J. Med. 2010, 362, 2295–2303. [Google Scholar] [CrossRef]
- Benowitz, N.L. Pharmacology of Nicotine: Addiction, Smoking-Induced Disease, and Therapeutics. Annu. Rev. Pharmacol. Toxicol. 2009, 49, 57–71. [Google Scholar] [CrossRef]
- De Biasi, M.; Dani, J.A. Reward, Addiction, Withdrawal to Nicotine. Annu. Rev. Neurosci. 2011, 34, 105–130. [Google Scholar] [CrossRef]
- Onor, I.O.; Stirling, D.L.; Williams, S.R.; Bediako, D.; Borghol, A.; Harris, M.B.; Darensburg, T.B.; Clay, S.D.; Okpechi, S.C.; Sarpong, D.F. Clinical Effects of Cigarette Smoking: Epidemiologic Impact and Review of Pharmacotherapy Options. Int. J. Environ. Res. Public Health 2017, 14, 1147. [Google Scholar] [CrossRef]
- Rosenthal, H.; Chow, N.; Mehta, S.; Pham, D.; Milanaik, R. Puff Bars: A Dangerous Trend in Adolescent Disposable e-Cigarette Use. Curr. Opin. Pediatr. 2022, 34, 288–294. [Google Scholar] [CrossRef]
- Granata, S.; Vivarelli, F.; Morosini, C.; Canistro, D.; Paolini, M.; Fairclough, L.C. Toxicological Aspects Associated with Consumption from Electronic Nicotine Delivery System (ENDS): Focus on Heavy Metals Exposure and Cancer Risk. Int. J. Mol. Sci. 2024, 25, 2737. [Google Scholar] [CrossRef]
- Vivarelli, F.; Granata, S.; Rullo, L.; Mussoni, M.; Candeletti, S.; Romualdi, P.; Fimognari, C.; Cruz-Chamorro, I.; Carrillo-Vico, A.; Paolini, M.; et al. On the Toxicity of E-Cigarettes Consumption: Focus on Pathological Cellular Mechanisms. Pharmacol. Res. 2022, 182, 106315. [Google Scholar] [CrossRef]
- Queloz, S.; Etter, J.-F. A Survey of Users of the IQOS Tobacco Vaporizer: Perceived Dependence and Perceived Effects on Cigarette Withdrawal Symptoms. J. Addict. Dis. 2021, 39, 208–214. [Google Scholar] [CrossRef]
- Adriaens, K.; Gucht, D.V.; Baeyens, F. IQOSTM vs. e-Cigarette vs. Tobacco Cigarette: A Direct Comparison of Short-Term Effects after Overnight-Abstinence. Int. J. Environ. Res. Public Health 2018, 15, 2902. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, M.; Krzystanek, M.; Zejda, J.E.; Majek, P.; Lubanski, J.; Lawson, J.A.; Brozek, G. E-Cigarettes Are More Addictive than Traditional Cigarettes-A Study in Highly Educated Young People. Int. J. Environ. Res. Public Health 2019, 16, 2279. [Google Scholar] [CrossRef] [PubMed]
- Vivarelli, F.; Morosini, C.; Rullo, L.; Losapio, L.M.; Lacorte, A.; Sangiorgi, S.; Ghini, S.; Fagiolino, I.; Franchi, P.; Lucarini, M.; et al. Effects of Unburned Tobacco Smoke on Inflammatory and Oxidative Mediators in the Rat Prefrontal Cortex. Front. Pharmacol. 2024, 15, 1328917. [Google Scholar] [CrossRef]
- Sivandzade, F.; Cucullo, L. Assessing the Protective Effect of Rosiglitazone against Electronic Cigarette/Tobacco Smoke-Induced Blood-Brain Barrier Impairment. BMC Neurosci. 2019, 20, 15. [Google Scholar] [CrossRef]
- Daynes, R.A.; Jones, D.C. Emerging Roles of PPARs in Inflammation and Immunity. Nat. Rev. Immunol. 2002, 2, 748–759. [Google Scholar] [CrossRef]
- Burgermeister, E.; Seger, R. PPARgamma and MEK Interactions in Cancer. PPAR Res. 2008, 2008, 309469. [Google Scholar] [CrossRef]
- Lucas, R.M.; Luo, L.; Stow, J.L. ERK1/2 in Immune Signalling. Biochem. Soc. Trans. 2022, 50, 1341–1352. [Google Scholar] [CrossRef]
- Kitagishi, Y.; Matsuda, S. Diets Involved in PPAR and PI3K/AKT/PTEN Pathway May Contribute to Neuroprotection in a Traumatic Brain Injury. Alzheimers Res. Ther. 2013, 5, 42. [Google Scholar] [CrossRef]
- Rullo, L.; Franchi, S.; Amodeo, G.; Caputi, F.F.; Verduci, B.; Losapio, L.M.; Sacerdote, P.; Romualdi, P.; Candeletti, S. Interplay between Prokineticins and Histone Demethylase KDM6A in a Murine Model of Bortezomib-Induced Neuropathy. Int. J. Mol. Sci. 2021, 22, 11913. [Google Scholar] [CrossRef]
- Kohno, M.; Link, J.; Dennis, L.E.; McCready, H.; Huckans, M.; Hoffman, W.F.; Loftis, J.M. Neuroinflammation in Addiction: A Review of Neuroimaging Studies and Potential Immunotherapies. Pharmacol. Biochem. Behav. 2019, 179, 34–42. [Google Scholar] [CrossRef]
- Johnstone, A.L.; Andrade, N.S.; Barbier, E.; Khomtchouk, B.B.; Rienas, C.A.; Lowe, K.; Van Booven, D.J.; Domi, E.; Esanov, R.; Vilca, S.; et al. Dysregulation of the Histone Demethylase KDM6B in Alcohol Dependence Is Associated with Epigenetic Regulation of Inflammatory Signaling Pathways. Addict. Biol. 2021, 26, e12816. [Google Scholar] [CrossRef]
- Zhang, Y.-X.; Akumuo, R.C.; España, R.A.; Yan, C.-X.; Gao, W.-J.; Li, Y.-C. The Histone Demethylase KDM6B in the Medial Prefrontal Cortex Epigenetically Regulates Cocaine Reward Memory. Neuropharmacology 2018, 141, 113–125. [Google Scholar] [CrossRef]
- Qureshi, R.; Kindo, M.; Boulberdaa, M.; von Hunolstein, J.-J.; Steenman, M.; Nebigil, C.G. A Prokineticin-Driven Epigenetic Switch Regulates Human Epicardial Cell Stemness and Fate. Stem Cells 2018, 36, 1589–1602. [Google Scholar] [CrossRef] [PubMed]
- Garza, A.P.; Morton, L.; Pállinger, É.; Buzás, E.I.; Schreiber, S.; Schott, B.H.; Dunay, I.R. Initial and Ongoing Tobacco Smoking Elicits Vascular Damage and Distinct Inflammatory Response Linked to Neurodegeneration. Brain Behav. Immun Health 2023, 28, 100597. [Google Scholar] [CrossRef] [PubMed]
- Matheson, J.; Le Foll, B. Therapeutic Potential of Peroxisome Proliferator-Activated Receptor (PPAR) Agonists in Substance Use Disorders: A Synthesis of Preclinical and Human Evidence. Cells 2020, 9, 1196. [Google Scholar] [CrossRef] [PubMed]
- Domi, E.; Caputi, F.F.; Romualdi, P.; Domi, A.; Scuppa, G.; Candeletti, S.; Atkins, A.; Heilig, M.; Demopulos, G.; Gaitanaris, G.; et al. Activation of PPARγ Attenuates the Expression of Physical and Affective Nicotine Withdrawal Symptoms through Mechanisms Involving Amygdala and Hippocampus Neurotransmission. J. Neurosci. 2019, 39, 9864–9875. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.B.; Most, D.; Blednov, Y.A.; Harris, R.A. PPAR Agonists Regulate Brain Gene Expression: Relationship to Their Effects on Ethanol Consumption. Neuropharmacology 2014, 86, 397–407. [Google Scholar] [CrossRef] [PubMed]
- de Guglielmo, G.; Kallupi, M.; Scuppa, G.; Demopulos, G.; Gaitanaris, G.; Ciccocioppo, R. Pioglitazone Attenuates the Opioid Withdrawal and Vulnerability to Relapse to Heroin Seeking in Rodents. Psychopharmacology 2017, 234, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.-X.; Hempel, B.; Crissman, M.; Pari, S.; Klein, B.; Bi, G.-H.; Alton, H. PPARα and PPARγ Are Expressed in Midbrain Dopamine Neurons and Modulate Dopamine- and Cannabinoid-Mediated Behavior in Mice. Mol. Psychiatry 2023, 28, 4203–4214. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, G.; La Rana, G.; Russo, R.; Sasso, O.; Iacono, A.; Esposito, E.; Mattace Raso, G.; Cuzzocrea, S.; Loverme, J.; Piomelli, D.; et al. Central Administration of Palmitoylethanolamide Reduces Hyperalgesia in Mice via Inhibition of NF-kappaB Nuclear Signalling in Dorsal Root Ganglia. Eur. J. Pharmacol. 2009, 613, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Stravodimou, A.; Mazzoccoli, G.; Voutsadakis, I.A. Peroxisome Proliferator-Activated Receptor Gamma and Regulations by the Ubiquitin-Proteasome System in Pancreatic Cancer. PPAR Res. 2012, 2012, 367450. [Google Scholar] [CrossRef]
- Kim, T.; Yang, Q. Peroxisome-Proliferator-Activated Receptors Regulate Redox Signaling in the Cardiovascular System. World J. Cardiol. 2013, 5, 164–174. [Google Scholar] [CrossRef]
- Collo, G.; Cavalleri, L.; Spano, P. Structural Plasticity in Mesencephalic Dopaminergic Neurons Produced by Drugs of Abuse: Critical Role of BDNF and Dopamine. Front. Pharmacol. 2014, 5, 259. [Google Scholar] [CrossRef] [PubMed]
- Skelton, P.D.; Frazel, P.W.; Lee, D.; Suh, H.; Luikart, B.W. Pten Loss Results in Inappropriate Excitatory Connectivity. Mol. Psychiatry 2019, 24, 1627–1640. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, M.S.; Markou, A. The “Stop” and “Go” of Nicotine Dependence: Role of GABA and Glutamate. Cold Spring Harb. Perspect. Med. 2013, 3, a012146. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.M.; Hake, P.W.; Denenberg, A.; Nowell, M.; Piraino, G.; Zingarelli, B. Phosphorylation of Extracellular Signal-Regulated Kinase (ERK)-1/2 Is Associated with the Downregulation of Peroxisome Proliferator-Activated Receptor (PPAR)-γ during Polymicrobial Sepsis. Mol. Med. 2010, 16, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, K.H.; Sodhi, A.; Chakraborty, R. Peptidoglycan Induced Expression of Peroxisome Proliferator-Activated Receptor γ in Mouse Peritoneal Macrophages: Role of ERK and JNK MAP Kinases. Cytokine 2012, 60, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Kawahata, I.; Cheng, A.; Fukunaga, K. The Role of CaMKII and ERK Signaling in Addiction. Int. J. Mol. Sci. 2021, 22, 3189. [Google Scholar] [CrossRef] [PubMed]
- Gould, T.J. Epigenetic and Long-Term Effects of Nicotine on Biology, Behavior, and Health. Pharmacol. Res. 2023, 192, 106741. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.J.; Ramesha, S.; Weinstock, L.D.; Gao, T.; Ping, L.; Xiao, H.; Dammer, E.B.; Duong, D.D.; Levey, A.I.; Lah, J.J.; et al. Extracellular Signal-Regulated Kinase Regulates Microglial Immune Responses in Alzheimer’s Disease. J. Neurosci. Res. 2021, 99, 1704–1721. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, R.; Doi, T.; Hayakawa, K.; Morioka, K.; Imamura, O.; Takishima, K.; Hamanoue, M.; Sawada, Y.; Nagao, M.; Tanaka, S.; et al. The Crucial Role of Erk2 in Demyelinating Inflammation in the Central Nervous System. J. Neuroinflamm. 2016, 13, 235. [Google Scholar] [CrossRef]
- Morella, I.; Pohořalá, V.; Calpe-López, C.; Brambilla, R.; Spanagel, R.; Bernardi, R.E. Nicotine Self-Administration and ERK Signaling Are Altered in RasGRF2 Knockout Mice. Front. Pharmacol. 2022, 13, 986566. [Google Scholar] [CrossRef]
- Yang, J.H.; Sohn, S.; Kim, S.; Kim, J.; Oh, J.H.; Ryu, I.S.; Go, B.S.; Choe, E.S. Repeated Nicotine Exposure Increases the Intracellular Interaction between ERK-mGluR5 in the Nucleus Accumbens More in Adult than Adolescent Rats. Addict. Biol. 2021, 26, e12913. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.-L.; Quizon, P.M.; Zhu, J. Molecular Mechanism: ERK Signaling, Drug Addiction, and Behavioral Effects. Prog. Mol. Biol. Transl. Sci. 2016, 137, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Vivarelli, F.; Canistro, D.; Cirillo, S.; Elias, R.J.; Granata, S.; Mussoni, M.; Burattini, S.; Falcieri, E.; Turrini, E.; Fimognari, C.; et al. Unburned Tobacco Cigarette Smoke Alters Rat Ultrastructural Lung Airways and DNA. Nicotine Tob. Res. 2021, 23, 2127–2134. [Google Scholar] [CrossRef] [PubMed]
- Granata, S.; Canistro, D.; Vivarelli, F.; Morosini, C.; Rullo, L.; Mercatante, D.; Rodriguez-Estrada, M.T.; Baracca, A.; Sgarbi, G.; Solaini, G.; et al. Potential Harm of IQOS Smoke to Rat Liver. Int. J. Mol. Sci. 2023, 24, 12462. [Google Scholar] [CrossRef] [PubMed]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE Guidelines 2.0: Updated Guidelines for Reporting Animal Research. PLoS Biol. 2020, 18, e3000410. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 7th ed.; Academic Press: San Diego, CA, USA, 2013. [Google Scholar]
- Chomczynski, P.; Sacchi, N. The Single-Step Method of RNA Isolation by Acid Guanidinium Thiocyanate-Phenol-Chloroform Extraction: Twenty-Something Years On. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef]
- Caputi, F.F.; Di Cesare Mannelli, L.; Rullo, L.; Micheli, L.; Stamatakos, S.; Posa, L.; Ghelardini, C.; Romualdi, P.; Candeletti, S. The Active Second-Generation Proteasome Inhibitor Oprozomib Reverts the Oxaliplatin-Induced Neuropathy Symptoms. Biochem. Pharmacol. 2020, 182, 114255. [Google Scholar] [CrossRef]
- Caputi, F.F.; Rullo, L.; Acquas, E.; Ciccocioppo, R.; Candeletti, S.; Romualdi, P. Evidence of a PPARγ-Mediated Mechanism in the Ability of Withania Somnifera to Attenuate Tolerance to the Antinociceptive Effects of Morphine. Pharmacol. Res. 2019, 139, 422–430. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morosini, C.; Vivarelli, F.; Rullo, L.; Volino, E.; Losapio, L.M.; Paolini, M.; Romualdi, P.; Canistro, D.; Candeletti, S. Unburned Tobacco Smoke Affects Neuroinflammation-Related Pathways in the Rat Mesolimbic System. Int. J. Mol. Sci. 2024, 25, 5259. https://doi.org/10.3390/ijms25105259
Morosini C, Vivarelli F, Rullo L, Volino E, Losapio LM, Paolini M, Romualdi P, Canistro D, Candeletti S. Unburned Tobacco Smoke Affects Neuroinflammation-Related Pathways in the Rat Mesolimbic System. International Journal of Molecular Sciences. 2024; 25(10):5259. https://doi.org/10.3390/ijms25105259
Chicago/Turabian StyleMorosini, Camilla, Fabio Vivarelli, Laura Rullo, Emilia Volino, Loredana Maria Losapio, Moreno Paolini, Patrizia Romualdi, Donatella Canistro, and Sanzio Candeletti. 2024. "Unburned Tobacco Smoke Affects Neuroinflammation-Related Pathways in the Rat Mesolimbic System" International Journal of Molecular Sciences 25, no. 10: 5259. https://doi.org/10.3390/ijms25105259
APA StyleMorosini, C., Vivarelli, F., Rullo, L., Volino, E., Losapio, L. M., Paolini, M., Romualdi, P., Canistro, D., & Candeletti, S. (2024). Unburned Tobacco Smoke Affects Neuroinflammation-Related Pathways in the Rat Mesolimbic System. International Journal of Molecular Sciences, 25(10), 5259. https://doi.org/10.3390/ijms25105259