
Cytoophidia Influence Cell Cycle and Size in Schizosaccharomyces pombe



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
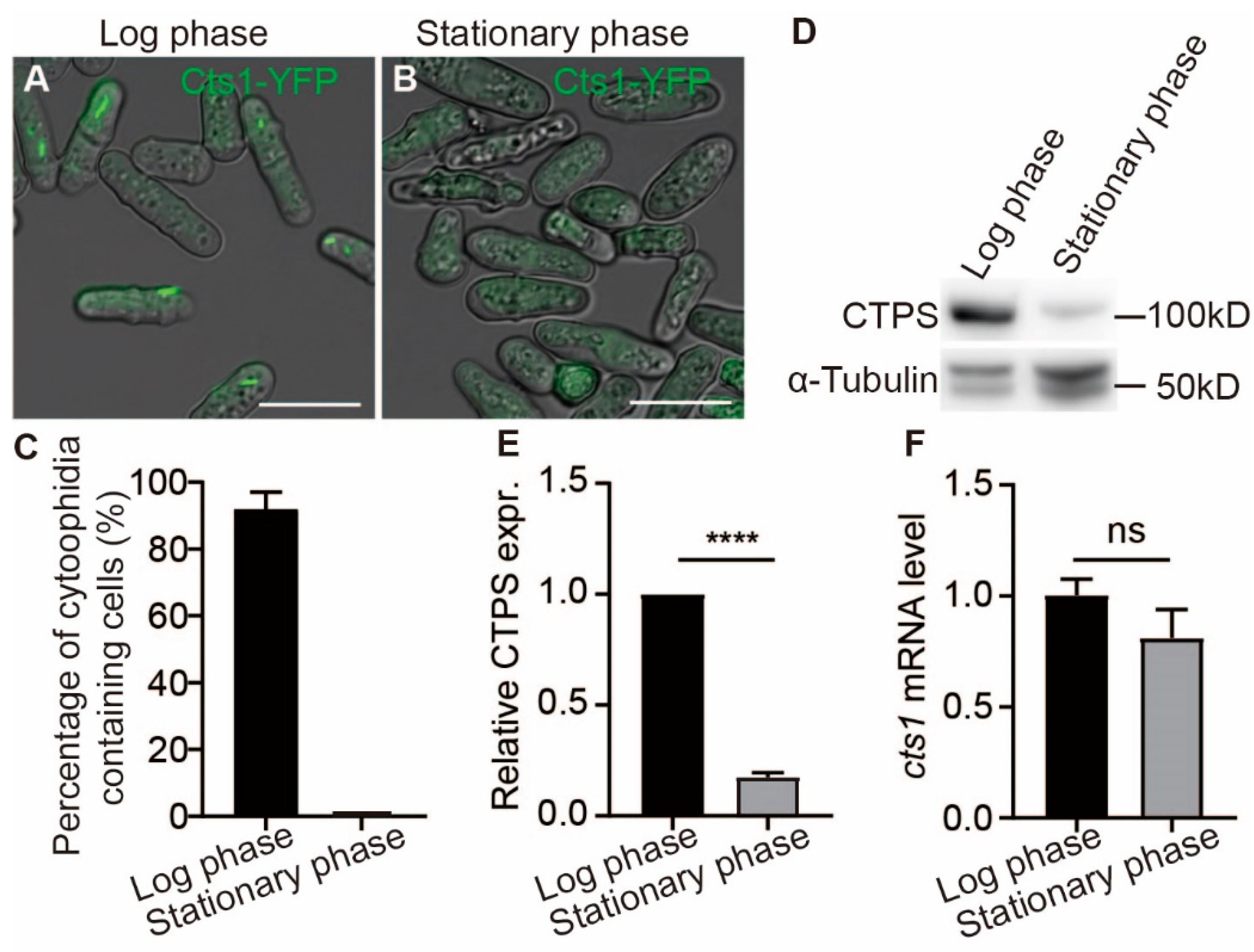
2.1. Cytoophidia Are Observed Exclusively during the Log Phase of S. pombe
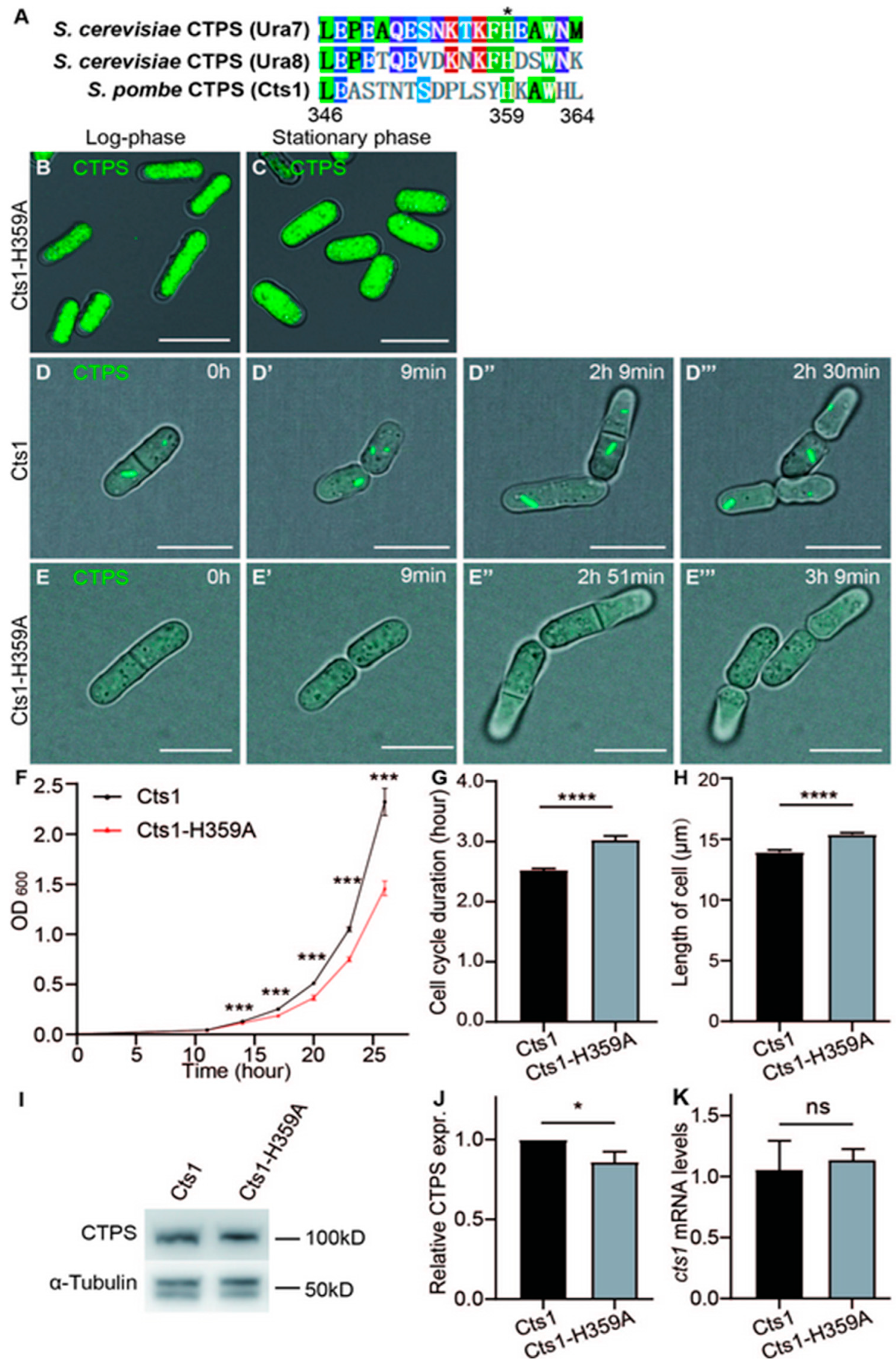
2.2. Loss-Filament Mutation Cts1-H359A Impacts the Cell Cycle and Size of S. pombe
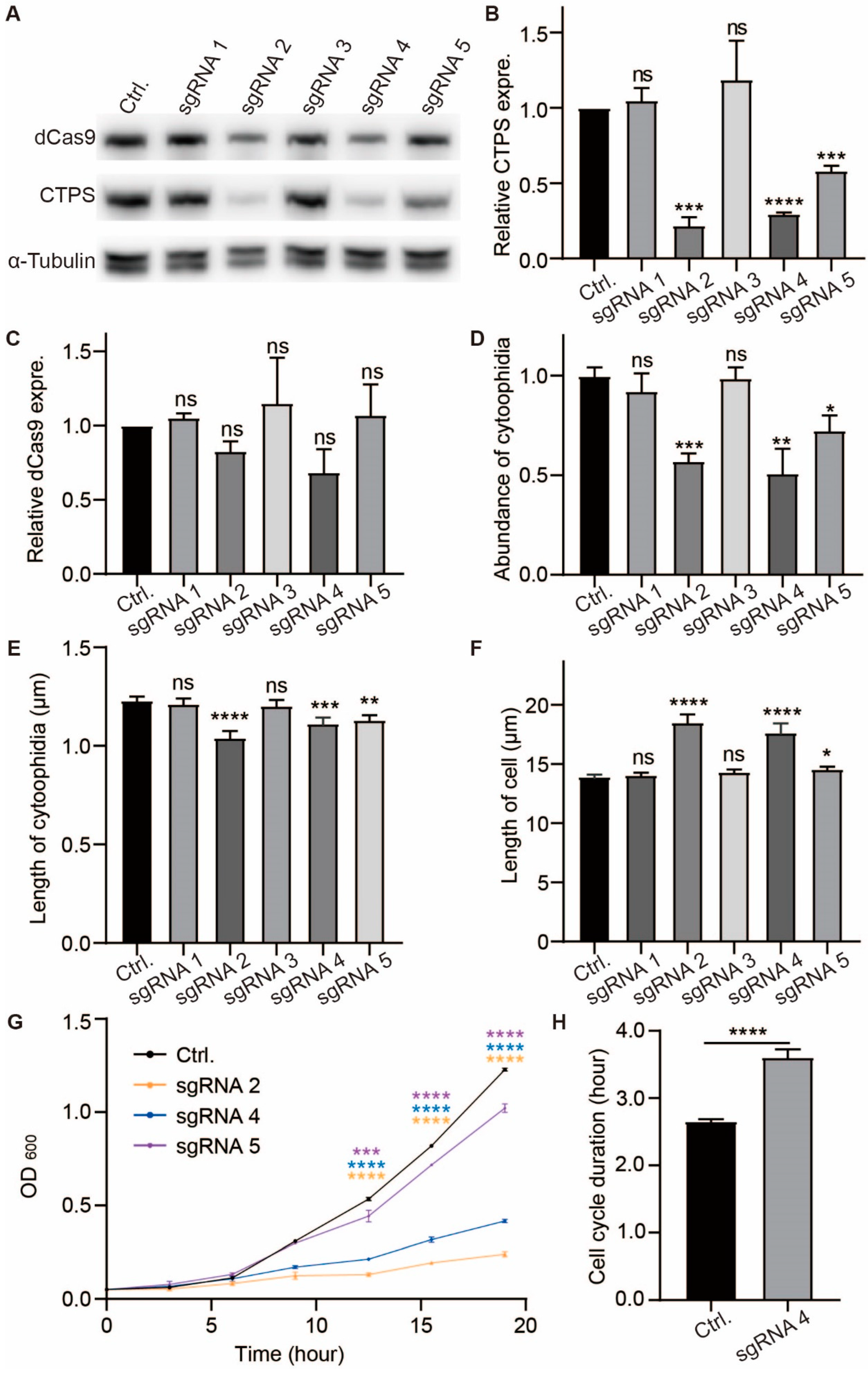
2.3. Both CTPS Protein Levels and Cytoophidia Affect the Cell Cycle and Size of S. pombe
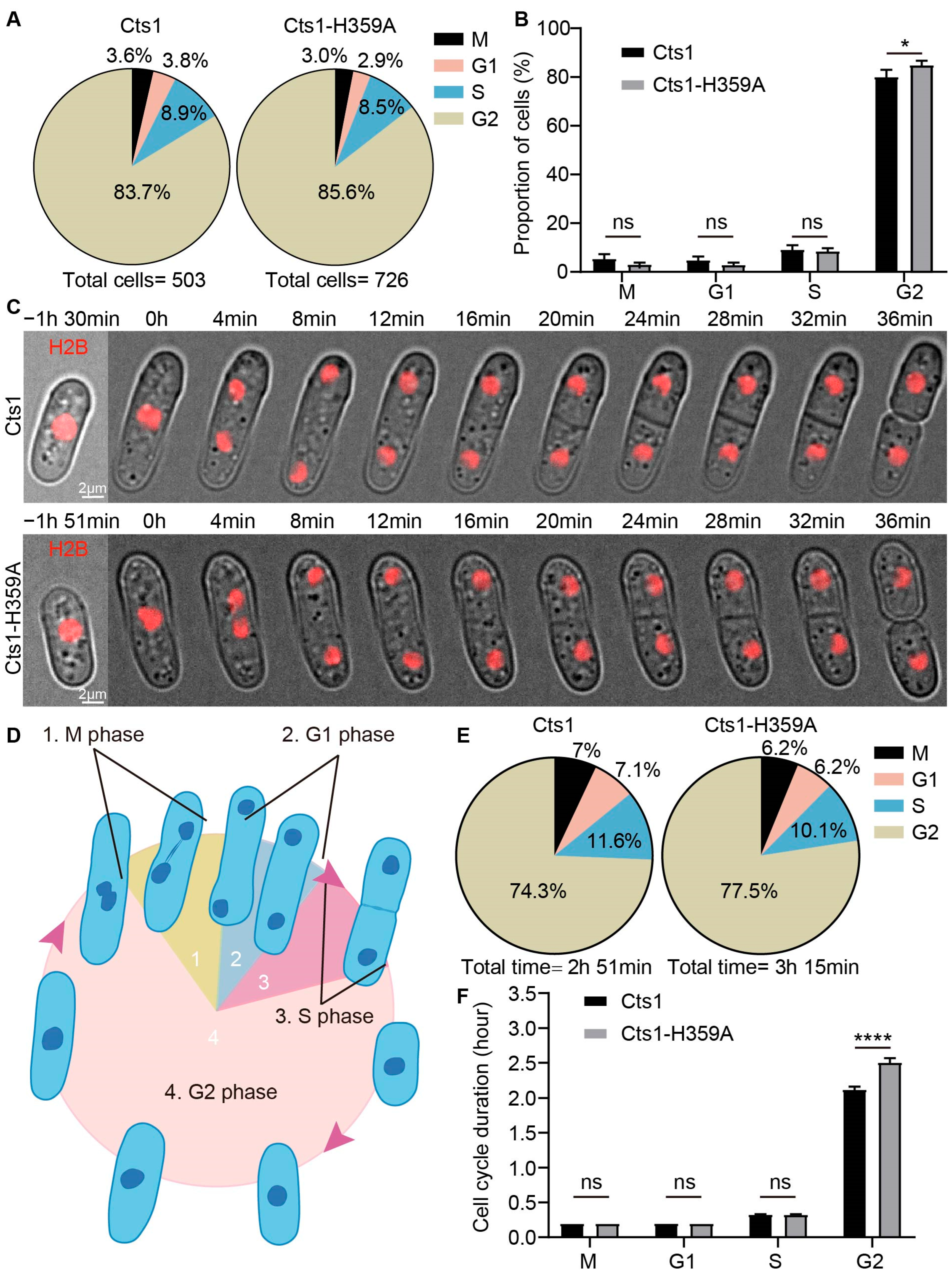
2.4. Cts1-H359A Mutation Extends the Duration of the G2 Phase in S. pombe
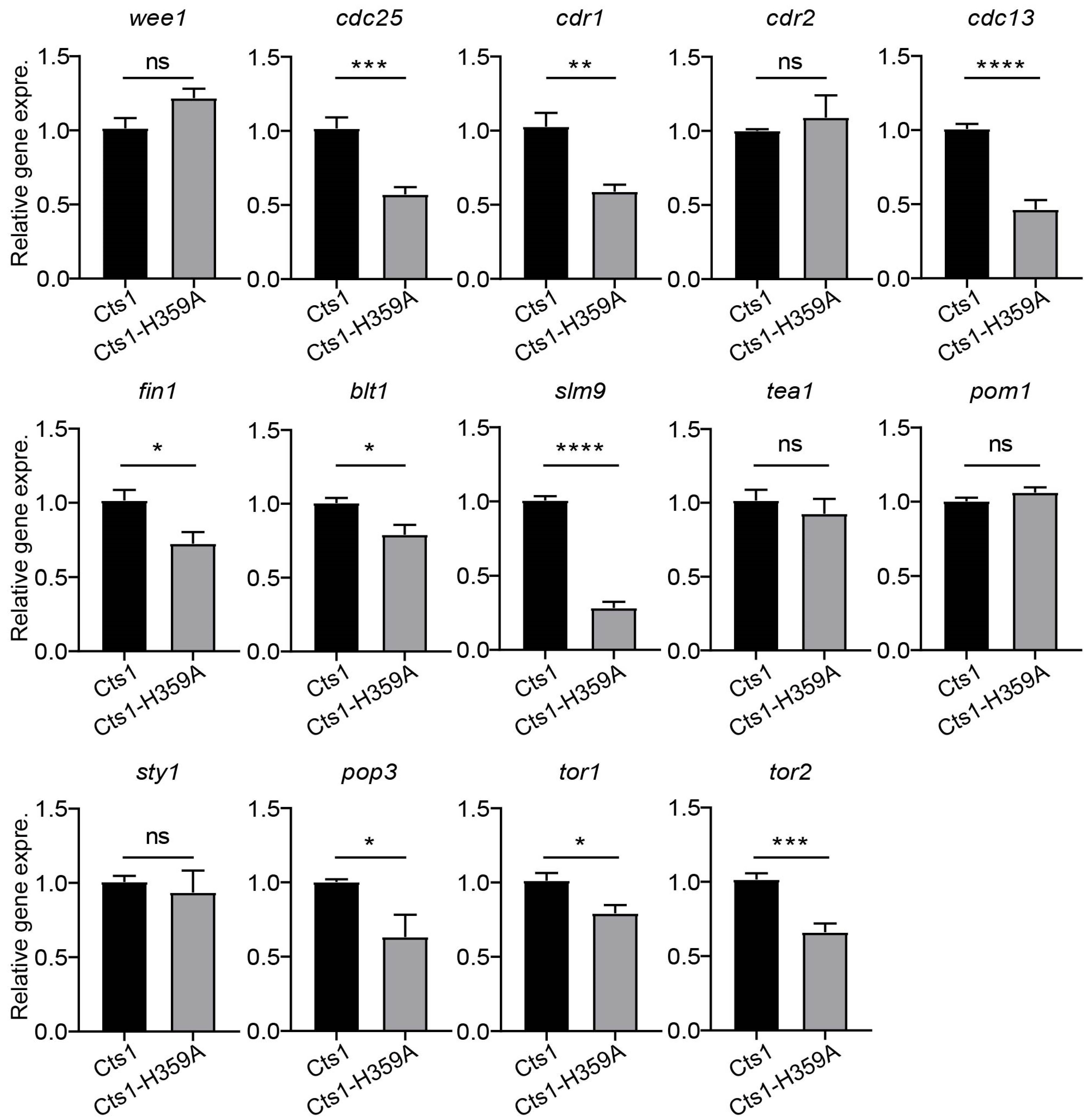
2.5. Cts1-H359A Mutation Perturbs G2/M Transition and the Expression of Cell Growth-Related Genes
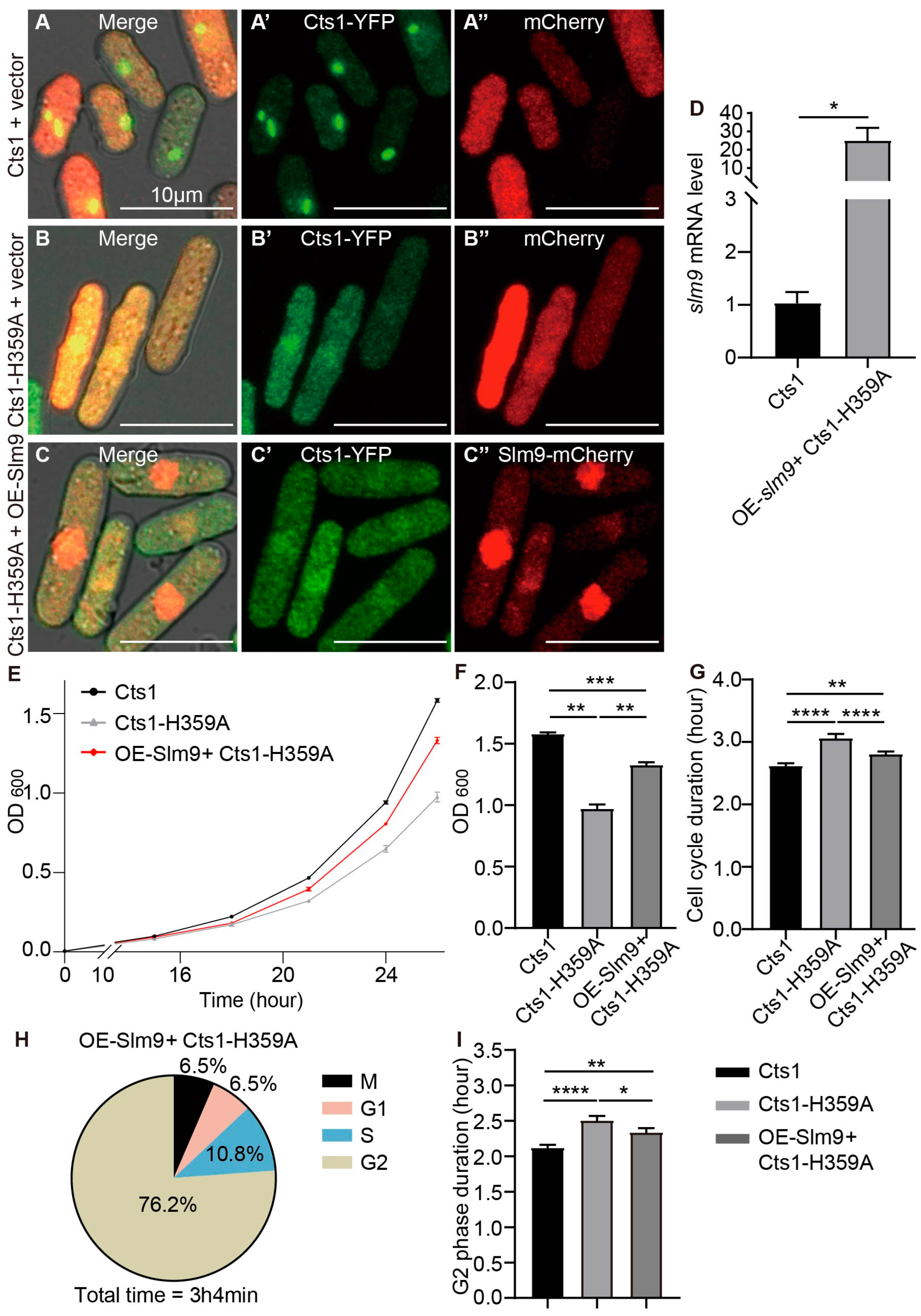
2.6. Overexpression of slm9 Alleviates Phenotypes Caused by Cts1-H359A Mutation
3. Discussion
4. Materials and Methods
4.1. Yeast Strain and Culture Medium
4.2. Plasmid Construction
4.3. Gene Suppression
4.4. Growth Assays
4.5. Cell Fixation, Image Acquisition, and Live-Cell Imaging
4.6. Western Blotting
4.7. RNA Isolation and RT-PCR
4.8. Cell Cycle and Cell Size Analysis
4.9. Quantification and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, J.L. The Cytoophidium and Its Kind: Filamentation and Compartmentation of Metabolic Enzymes. Annu. Rev. Cell Dev. Biol. 2016, 32, 349–372. [Google Scholar] [CrossRef]
- Noree, C.; Sato, B.K.; Broyer, R.M.; Wilhelm, J.E. Identification of Novel Filament-Forming Proteins in Saccharomyces cerevisiae and Drosophila melanogaster. J. Cell Biol. 2010, 190, 541–551. [Google Scholar] [CrossRef]
- Liu, J.L. Intracellular Compartmentation of CTP Synthase in Drosophila. J. Genet. Genom. 2010, 37, 281–296. [Google Scholar] [CrossRef]
- Ingerson-Mahar, M.; Briegel, A.; Werner, J.N.; Jensen, G.J.; Gitai, Z. The Metabolic Enzyme CTP Synthase Forms Cytoskeletal Filaments. Nat. Cell Biol. 2010, 12, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-C.; Jeng, Y.-M.; Peng, M.; Keppeke, G.D.; Sung, L.-Y.; Liu, J.-L. CTP Synthase Forms the Cytoophidium in Human Hepatocellular Carcinoma. Exp. Cell Res. 2017, 361, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zhang, J.; Tastan, Ö.Y.; Deussen, Z.A.; Siswick, M.Y.Y.; Liu, J.L. Glutamine Analogs Promote Cytoophidium Assembly in Human and Drosophila Cells. J. Genet. Genom. 2011, 38, 391–402. [Google Scholar] [CrossRef]
- Zhang, J.; Hulme, L.; Liu, J.L. Asymmetric Inheritance of Cytoophidia in Schizosaccharomyces Pombe. Biol. Open 2014, 3, 1092–1097. [Google Scholar] [CrossRef]
- Daumann, M.; Hickl, D.; Zimmer, D.; DeTar, R.A.; Kunz, H.H.; Möhlmann, T. Characterization of Filament-Forming CTP Synthases from Arabidopsis thaliana. Plant J. 2018, 96, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Xiang, H.; Liu, J.L. CTP Synthase Forms Cytoophidia in Archaea. J. Genet. Genom. 2020, 47, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Chang, C.C.; Liu, J.L.; Sung, L.Y. CTPS and IMPDH Form Cytoophidia in Developmental Thymocytes. Exp. Cell Res. 2021, 405, 112662. [Google Scholar] [CrossRef]
- Barry, R.M.; Bitbol, A.-F.; Lorestani, A.; Charles, E.J.; Habrian, C.H.; Hansen, J.M.; Li, H.-J.; Baldwin, E.P.; Wingreen, N.S.; Kollman, J.M.; et al. Large-Scale Filament Formation Inhibits the Activity of CTP Synthetase. eLife 2014, 3, e03638. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Guo, C.J.; Chang, C.C.; Zhong, J.; Hu, H.H.; Lu, G.M.; Liu, J.L. Structural Basis for Ligand Binding Modes of CTP Synthase. Proc. Natl. Acad. Sci. USA 2021, 118, e2026621118. [Google Scholar] [CrossRef]
- Aughey, G.N.; Grice, S.J.; Shen, Q.-J.; Xu, Y.; Chang, C.-C.; Azzam, G.; Wang, P.-Y.; Freeman-Mills, L.; Pai, L.-M.; Sung, L.-Y.; et al. Nucleotide Synthesis Is Regulated by Cytoophidium Formation during Neurodevelopment and Adaptive Metabolism. Biol. Open 2014, 3, 1045–1056. [Google Scholar] [CrossRef] [PubMed]
- Lynch, E.M.; Hicks, D.R.; Shepherd, M.; Endrizzi, J.A.; Maker, A.; Hansen, J.M.; Barry, R.M.; Gitai, Z.; Baldwin, E.P.; Kollman, J.M. Human CTP Synthase Filament Structure Reveals the Active Enzyme Conformation. Nat. Struct. Mol. Biol. 2017, 24, 507–514. [Google Scholar] [CrossRef]
- Tastan, Ö.Y.; Liu, J.L. CTP Synthase Is Required for Optic Lobe Homeostasis in Drosophila. J. Genet. Genom. 2015, 42, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.J.; Liu, J.L. Cytoophidia and filaments: You must unlearn what you have learned. Biochem Soc Trans. 2023, 51, 1245–1256. [Google Scholar] [CrossRef]
- Aughey, G.N.; Grice, S.J.; Liu, J.L. The Interplay between Myc and CTP Synthase in Drosophila. PLoS Genet. 2016, 12, e1005867. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, Y.; Zhou, Y.; Wang, Q.Q.; Ding, K.; Zhao, S.; Lu, P.; Liu, J.L. Cytoophidia Coupling Adipose Architecture and Metabolism. Cell. Mol. Life Sci. 2022, 79, 534. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, Y.; Wang, Q.Q.; Zhou, Y.; Liu, J.L. Fat body-specific reduction of CTPS alleviates HFD-induced obesity. eLife 2023, 12, e85293. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, J.-L. Forming Cytoophidia Prolongs the Half-Life of CTP Synthase. Cell Discov. 2019, 5, 32. [Google Scholar] [CrossRef]
- Chang, C.C.; Peng, M.; Zhong, J.; Zhang, Z.; Keppeke, G.D.; Sung, L.Y.; Liu, J.L. Molecular crowding facilitates bundling of IMPDH polymers and cytoophidium formation. Cell Mol Life Sci. 2022, 79, 420. [Google Scholar] [CrossRef]
- Keppeke, G.D.; Chang, C.C.; Zhang, Z.; Liu, J.L. Effect on cell survival and cytoophidium assembly of the adRP-10-related IMPDH1 missense mutation Asp226Asn. Front Cell Dev Biol. 2023, 11, 1234592. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Liu, J.-L. Cytoophidia Respond to Nutrient Stress in Drosophila. Exp. Cell Res. 2019, 376, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.M.; Horowitz, A.; Lynch, E.M.; Farrell, D.P.; Quispe, J.; DiMaio, F.; Kollman, J.M. Cryo-EM structures of CTP synthase filaments reveal mechanism of pH-sensitive assembly during budding yeast starvation. eLife 2021, 10, e73368. [Google Scholar] [CrossRef]
- Li, Y.L.; Liu, J.L. Hypoosmolality Impedes Cytoophidium Integrity during Nitrogen Starvation. Yeast 2021, 38, 276–289. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.C.; Kizaki, H.; Weber, G.; Morris, H.P. Increased CTP synthetase activity in cancer cells. Nature 1978, 271, 71–73. [Google Scholar] [CrossRef]
- Ellims, P.H.; Gan, T.E.; Medley, G. Cytidine Triphosphate Synthetase Activity in Lymphoproliferative Disorders. Cancer Res. 1983, 43, 1432–1435. [Google Scholar] [PubMed]
- Carcamo, W.C.; Satoh, M.; Kasahara, H.; Terada, N.; Hamazaki, T.; Chan, J.Y.F.; Yao, B.; Tamayo, S.; Covini, G.; von Mühlen, C.A.; et al. Induction of Cytoplasmic Rods and Rings Structures by Inhibition of the CTP and GTP Synthetic Pathway in Mammalian Cells. PLoS ONE 2011, 6, e29690. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, J.L. Temperature-Sensitive Cytoophidium Assembly in Schizosaccharomyces pombe. J. Genet. Genom. 2019, 46, 423–432. [Google Scholar] [CrossRef]
- Ishikawa, K.; Soejima, S.; Masuda, F.; Saitoh, S. Implementation of DCas9-Mediated CRISPRi in the Fission Yeast Schizosaccharomyces pombe. G3 Genes Genomes Genet. 2021, 11, jkab051. [Google Scholar] [CrossRef]
- Forsburg, S.L.; Rhind, N. Basic Methods for Fission Yeast. Yeast 2006, 23, 173–183. [Google Scholar] [CrossRef]
- Colato, A.; Fontanari, J.F. Fission Yeast Handbook. Phys. Rev. Lett. 2001, 87, 238102. [Google Scholar] [CrossRef]
- Kanoh, J.; Russell, P. Slm9, a Novel Nuclear Protein Involved in Mitotic Control in Fission Yeast. Genetics 2000, 155, 623–631. [Google Scholar] [CrossRef]
- Anderson, H.E.; Kagansky, A.; Wardle, J.; Rappsilber, J.; Allshire, R.C.; Whitehall, S.K. Silencing Mediated by the Schizosaccharomyces pombe HIRA Complex Is Dependent upon the Hpc2-like Protein, Hip4. PLoS ONE 2010, 5, 2–9. [Google Scholar] [CrossRef]
- Feng, H.C.; Andreadis, C.; Liu, J.L. Histone Transcription Regulator Slm9 Is Required for Cytoophidium Biogenesis. Exp. Cell Res. 2021, 403, 112582. [Google Scholar] [CrossRef]
- Zhou, X.; Guo, C.J.; Hu, H.H.; Zhong, J.; Sun, Q.; Liu, D.; Zhou, S.; Chang, C.C.; Liu, J.L. Drosophila CTP Synthase Can Form Distinct Substrate- and Product-Bound Filaments. J. Genet. Genom. 2019, 46, 537–545. [Google Scholar] [CrossRef]
- Bähler, J.; Wu, J.Q.; Longtine, M.S.; Shah, N.G.; McKenzie, A.; Steever, A.B.; Wach, A.; Philippsen, P.; Pringle, J.R. Heterologous Modules for Efficient and Versatile PCR-Based Gene Targeting in Schizosaccharomyces pombe. Yeast 1998, 14, 943–951. [Google Scholar] [CrossRef]
- Shalem, O.; Sanjana, N.E.; Hartenian, E.; Shi, X.; Scott, D.A.; Mikkelson, T.; Heckl, D.; Ebert, B.L.; Root, D.E.; Doench, J.G.; et al. Genome-scale CRISPR-Cas9 knockout screening in human cells. Science 2014, 343, 84–87. [Google Scholar] [CrossRef]
- Matsuo, Y.; Asakawa, K.; Toda, T.; Katayama, S. A Rapid Method for Protein Extraction from Fission Yeast. Biosci. Biotechnol. Biochem. 2006, 70, 1992–1994. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, R.; Li, Y.-L.; Liu, J.-L. Cytoophidia Influence Cell Cycle and Size in Schizosaccharomyces pombe. Int. J. Mol. Sci. 2024, 25, 608. https://doi.org/10.3390/ijms25010608
Deng R, Li Y-L, Liu J-L. Cytoophidia Influence Cell Cycle and Size in Schizosaccharomyces pombe. International Journal of Molecular Sciences. 2024; 25(1):608. https://doi.org/10.3390/ijms25010608
Chicago/Turabian StyleDeng, Ruolan, Yi-Lan Li, and Ji-Long Liu. 2024. "Cytoophidia Influence Cell Cycle and Size in Schizosaccharomyces pombe" International Journal of Molecular Sciences 25, no. 1: 608. https://doi.org/10.3390/ijms25010608
APA StyleDeng, R., Li, Y.-L., & Liu, J.-L. (2024). Cytoophidia Influence Cell Cycle and Size in Schizosaccharomyces pombe. International Journal of Molecular Sciences, 25(1), 608. https://doi.org/10.3390/ijms25010608