
Exploring the Impact of Exosomal Cargos on Osteosarcoma Progression: Insights into Therapeutic Potential


Abstract
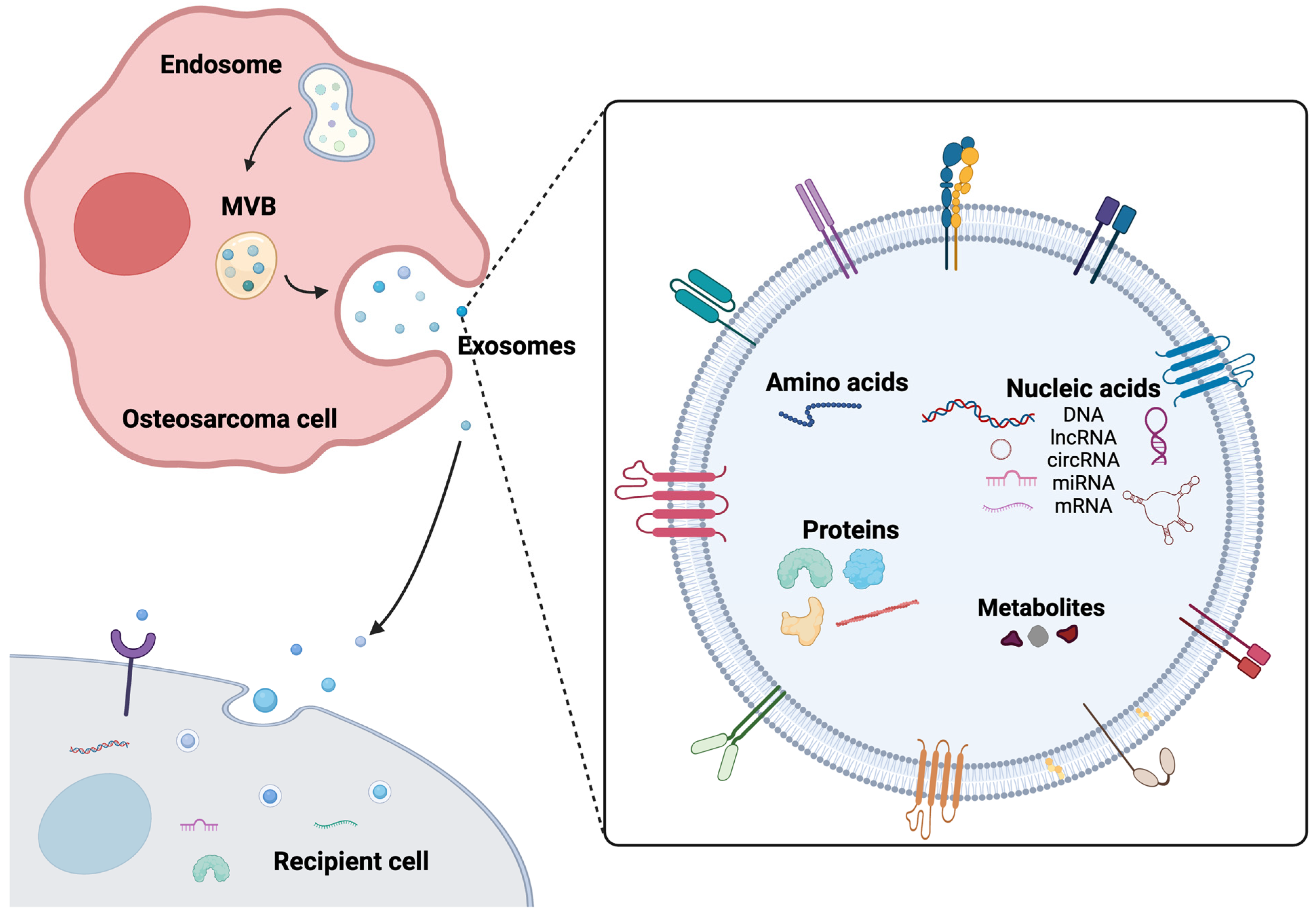
1. Introduction
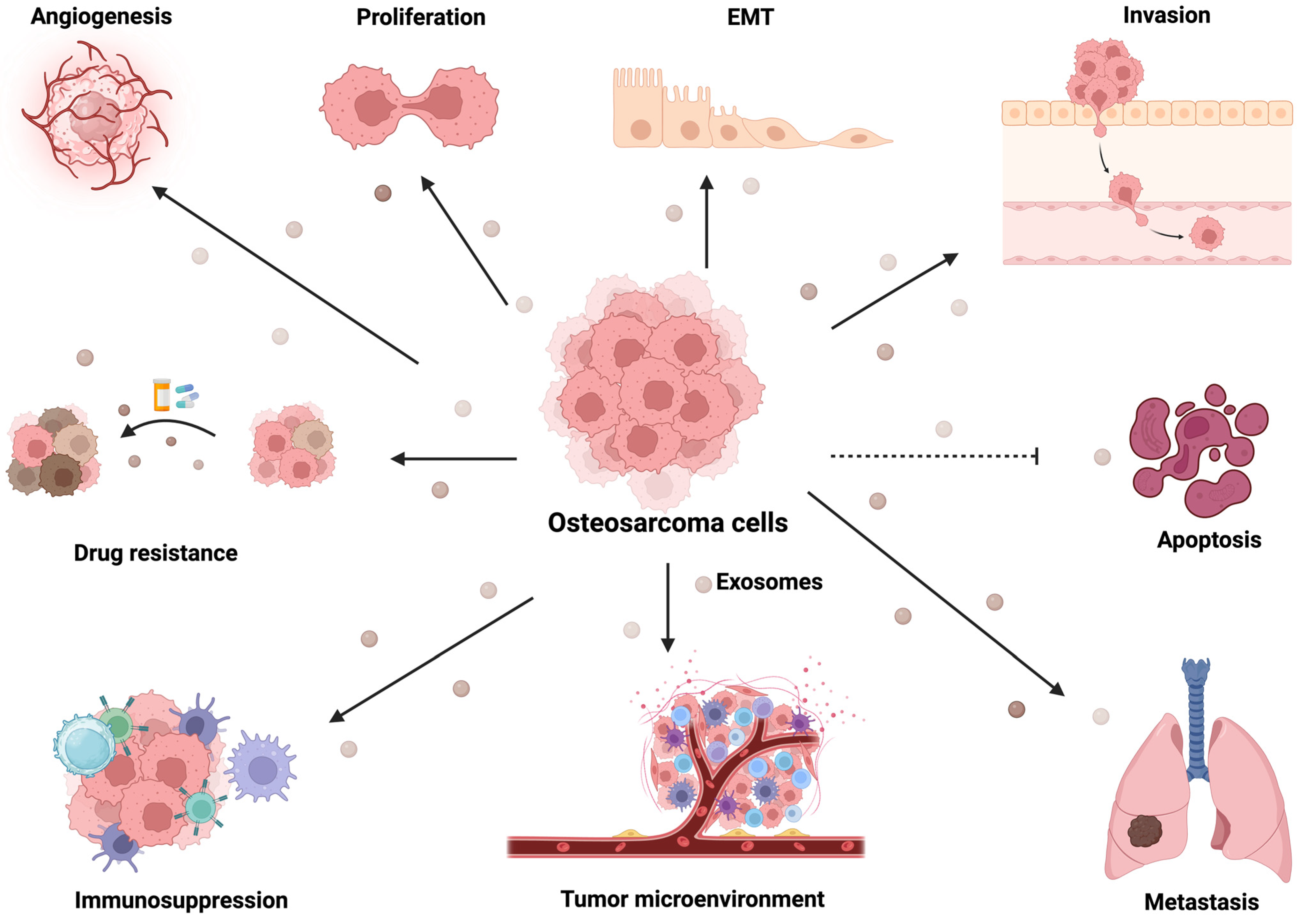
2. Osteosarcoma-Derived Exosomes Mediate Angiogenesis
3. Exosomal Cargos Mediate Metastasis of Osteosarcoma
4. Osteosarcoma-Derived Exosomal Cargos Mediated Immune Regulation
5. Exosomal Cargos Mediate Osteosarcoma Drug Resistance
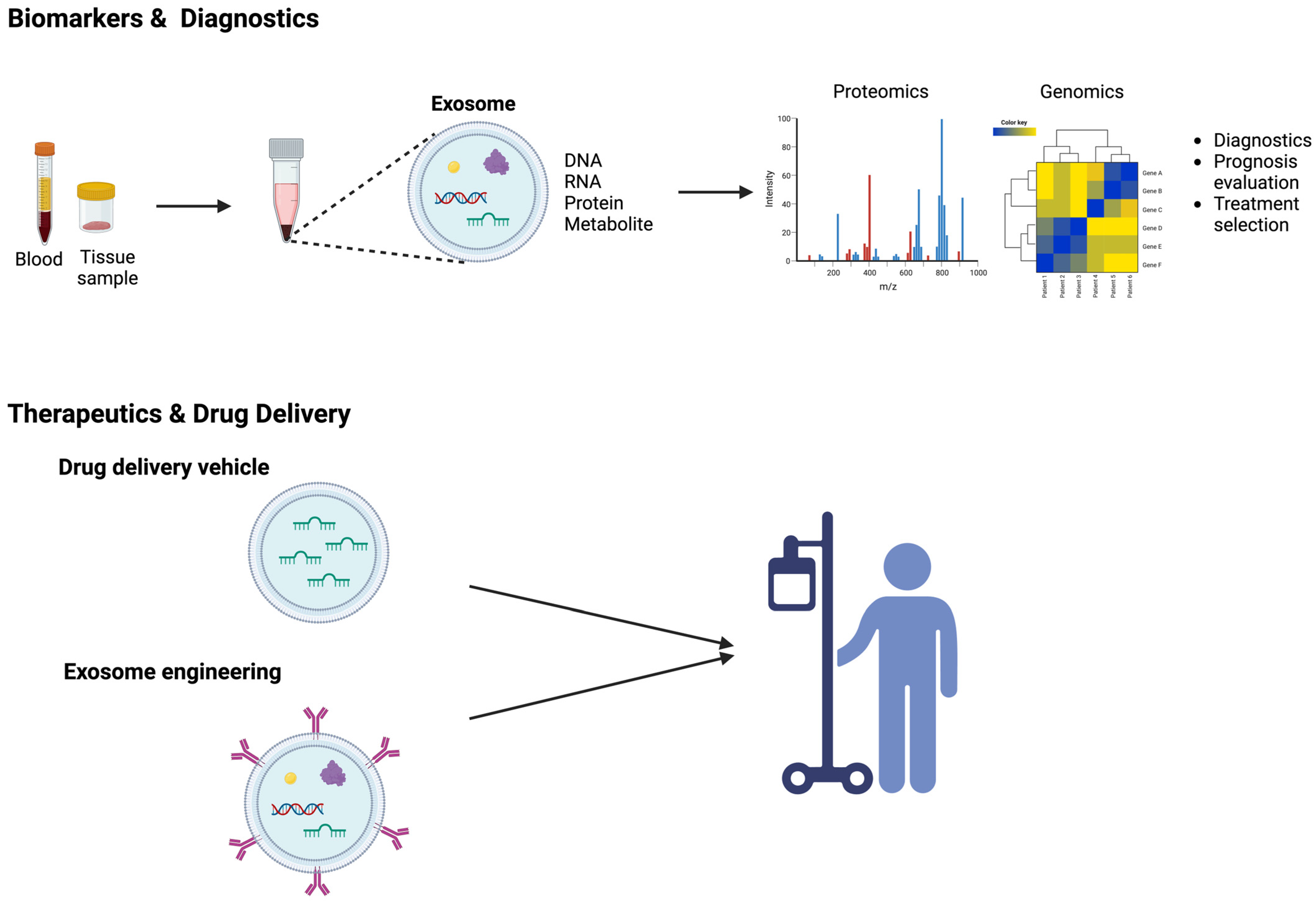
6. Exosomes as Biomarkers in Osteosarcoma
7. Exosomes as a Vehicle for Drug Delivery in Osteosarcoma Therapy
8. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Belayneh, R.; Fourman, M.S.; Bhogal, S.; Weiss, K.R. Update on Osteosarcoma. Curr. Oncol. Rep. 2021, 23, 71. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, G.; Jaffe, N. The Epidemiology of Osteosarcoma. Cancer Treat. Res. 2009, 152, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Bielack, S.S.; Blattmann, C.; Borkhardt, A.; Csóka, M.; Hassenpflug, W.; Kabíčková, E.; Kager, L.; Kessler, T.; Kratz, C.; Kühne, T.; et al. Osteosarcoma and Causes of Death: A Report of 1520 Deceased Patients from the Cooperative Osteosarcoma Study Group (COSS). Eur. J. Cancer 2022, 176, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Meltzer, P.S.; Helman, L.J. New Horizons in the Treatment of Osteosarcoma. N. Engl. J. Med. 2021, 385, 2066–2076. [Google Scholar] [CrossRef] [PubMed]
- Rosen, G.; Marcove, R.C.; Caparros, B.; Nirenberg, A.; Kosloff, C.; Huvos, A.G. Primary Osteogenic Sarcoma. The Rationale for Preoperative Chemotherapy and Delayed Surgery. Cancer 1979, 43, 2163–2177. [Google Scholar] [CrossRef] [PubMed]
- Marina, N.M.; Smeland, S.; Bielack, S.S.; Bernstein, M.; Jovic, G.; Krailo, M.D.; Hook, J.M.; Arndt, C.; van den Berg, H.; Brennan, B.; et al. Comparison of MAPIE versus MAP in Patients with a Poor Response to Preoperative Chemotherapy for Newly Diagnosed High-Grade Osteosarcoma (EURAMOS-1): An Open-Label, International, Randomised Controlled Trial. Lancet Oncol. 2016, 17, 1396–1408. [Google Scholar] [CrossRef]
- Kansara, M.; Teng, M.W.; Smyth, M.J.; Thomas, D.M. Translational Biology of Osteosarcoma. Nat. Rev. Cancer 2014, 14, 722–735. [Google Scholar] [CrossRef]
- Link, M.P.; Goorin, A.M.; Miser, A.W.; Green, A.A.; Pratt, C.B.; Belasco, J.B.; Pritchard, J.; Malpas, J.S.; Baker, A.R.; Kirkpatrick, J.A.; et al. The Effect of Adjuvant Chemotherapy on Relapse-Free Survival in Patients with Osteosarcoma of the Extremity. N. Engl. J. Med. 1986, 314, 1600–1606. [Google Scholar] [CrossRef]
- Théry, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, Biogenesis and Function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef]
- Kowal, J.; Tkach, M.; Théry, C. Biogenesis and Secretion of Exosomes. Curr. Opin. Cell Biol. 2014, 29, 116–125. [Google Scholar] [CrossRef]
- Trajkovic, K.; Hsu, C.; Chiantia, S.; Rajendran, L.; Wenzel, D.; Wieland, F.; Schwille, P.; Brügger, B.; Simons, M. Ceramide Triggers Budding of Exosome Vesicles into Multivesicular Endosomes. Science 2008, 319, 1244–1247. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Boussac, M.; Véron, P.; Ricciardi-Castagnoli, P.; Raposo, G.; Garin, J.; Amigorena, S. Proteomic Analysis of Dendritic Cell-Derived Exosomes: A Secreted Subcellular Compartment Distinct from Apoptotic Vesicles1. J. Immunol. 2001, 166, 7309–7318. [Google Scholar] [CrossRef] [PubMed]
- Dixon, C.L.; Sheller-Miller, S.; Saade, G.R.; Fortunato, S.J.; Lai, A.; Palma, C.; Guanzon, D.; Salomon, C.; Menon, R. Amniotic Fluid Exosome Proteomic Profile Exhibits Unique Pathways of Term and Preterm Labor. Endocrinology 2018, 159, 2229–2240. [Google Scholar] [CrossRef] [PubMed]
- Melo, S.A.; Luecke, L.B.; Kahlert, C.; Fernandez, A.F.; Gammon, S.T.; Kaye, J.; LeBleu, V.S.; Mittendorf, E.A.; Weitz, J.; Rahbari, N.; et al. Glypican-1 Identifies Cancer Exosomes and Detects Early Pancreatic Cancer. Nature 2015, 523, 177–182. [Google Scholar] [CrossRef]
- Saman, S.; Kim, W.; Raya, M.; Visnick, Y.; Miro, S.; Saman, S.; Jackson, B.; McKee, A.C.; Alvarez, V.E.; Lee, N.C.Y.; et al. Exosome-Associated Tau Is Secreted in Tauopathy Models and Is Selectively Phosphorylated in Cerebrospinal Fluid in Early Alzheimer Disease*. J. Biol. Chem. 2012, 287, 3842–3849. [Google Scholar] [CrossRef]
- Lässer, C.; Seyed Alikhani, V.; Ekström, K.; Eldh, M.; Torregrosa Paredes, P.; Bossios, A.; Sjöstrand, M.; Gabrielsson, S.; Lötvall, J.; Valadi, H. Human Saliva, Plasma and Breast Milk Exosomes Contain RNA: Uptake by Macrophages. J. Transl. Med. 2011, 9, 9. [Google Scholar] [CrossRef]
- Keller, S.; Ridinger, J.; Rupp, A.-K.; Janssen, J.W.; Altevogt, P. Body Fluid Derived Exosomes as a Novel Template for Clinical Diagnostics. J. Transl. Med. 2011, 9, 86. [Google Scholar] [CrossRef]
- Masyuk, A.I.; Huang, B.Q.; Ward, C.J.; Gradilone, S.A.; Banales, J.M.; Masyuk, T.V.; Radtke, B.; Splinter, P.L.; LaRusso, N.F. Biliary Exosomes Influence Cholangiocyte Regulatory Mechanisms and Proliferation through Interaction with Primary Cilia. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G990–G999. [Google Scholar] [CrossRef]
- Michael, A.; Bajracharya, S.; Yuen, P.; Zhou, H.; Star, R.; Illei, G.; Alevizos, I. Exosomes from Human Saliva as a Source of microRNA Biomarkers. Oral Dis. 2010, 16, 34–38. [Google Scholar] [CrossRef]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and Characterization of Exosomes from Cell Culture Supernatants and Biological Fluids. Curr. Protoc. Cell Biol. 2006, 30, 3.22.1–3.22.29. [Google Scholar] [CrossRef]
- Pisitkun, T.; Shen, R.-F.; Knepper, M.A. Identification and Proteomic Profiling of Exosomes in Human Urine. Proc. Natl. Acad. Sci. USA 2004, 101, 13368–13373. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Kim, H.; Choi, Y.J.; Kim, S.Y.; Lee, J.-E.; Sung, K.J.; Sung, Y.H.; Pack, C.-G.; Jung, M.; Han, B.; et al. Exosomal PD-L1 Promotes Tumor Growth through Immune Escape in Non-Small Cell Lung Cancer. Exp. Mol. Med. 2019, 51, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Thomou, T.; Mori, M.A.; Dreyfuss, J.M.; Konishi, M.; Sakaguchi, M.; Wolfrum, C.; Rao, T.N.; Winnay, J.N.; Garcia-Martin, R.; Grinspoon, S.K.; et al. Adipose-Derived Circulating miRNAs Regulate Gene Expression in Other Tissues. Nature 2017, 542, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, A.; Costa-Silva, B.; Shen, T.-L.; Rodrigues, G.; Hashimoto, A.; Tesic Mark, M.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour Exosome Integrins Determine Organotropic Metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Record, M.; Carayon, K.; Poirot, M.; Silvente-Poirot, S. Exosomes as New Vesicular Lipid Transporters Involved in Cell–Cell Communication and Various Pathophysiologies. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2014, 1841, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Azmi, A.S.; Bao, B.; Sarkar, F.H. Exosomes in Cancer Development, Metastasis, and Drug Resistance: A Comprehensive Review. Cancer Metastasis Rev. 2013, 32, 623–642. [Google Scholar] [CrossRef] [PubMed]
- Ekström, K.; Omar, O.; Granéli, C.; Wang, X.; Vazirisani, F.; Thomsen, P. Monocyte Exosomes Stimulate the Osteogenic Gene Expression of Mesenchymal Stem Cells. PLoS ONE 2013, 8, e75227. [Google Scholar] [CrossRef]
- Umezu, T.; Ohyashiki, K.; Kuroda, M.; Ohyashiki, J.H. Leukemia Cell to Endothelial Cell Communication via Exosomal miRNAs. Oncogene 2013, 32, 2747–2755. [Google Scholar] [CrossRef]
- Camussi, G.; Deregibus, M.-C.; Bruno, S.; Grange, C.; Fonsato, V.; Tetta, C. Exosome/Microvesicle-Mediated Epigenetic Reprogramming of Cells. Am. J. Cancer Res. 2010, 1, 98–110. [Google Scholar]
- Théry, C.; Ostrowski, M.; Segura, E. Membrane Vesicles as Conveyors of Immune Responses. Nat. Rev. Immunol. 2009, 9, 581–593. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-Mediated Transfer of mRNAs and microRNAs Is a Novel Mechanism of Genetic Exchange between Cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Harris, S.L.; Levine, A.J. The Regulation of Exosome Secretion: A Novel Function of the P53 Protein. Cancer Res. 2006, 66, 4795–4801. [Google Scholar] [CrossRef] [PubMed]
- Raimondi, L.; De Luca, A.; Gallo, A.; Costa, V.; Russelli, G.; Cuscino, N.; Manno, M.; Raccosta, S.; Carina, V.; Bellavia, D.; et al. Osteosarcoma Cell-Derived Exosomes Affect Tumor Microenvironment by Specific Packaging of microRNAs. Carcinogenesis 2020, 41, 666–677. [Google Scholar] [CrossRef] [PubMed]
- Wolf-Dennen, K.; Gordon, N.; Kleinerman, E.S. Exosomal Communication by Metastatic Osteosarcoma Cells Modulates Alveolar Macrophages to an M2 Tumor-Promoting Phenotype and Inhibits Tumoricidal Functions. OncoImmunology 2020, 9, 1747677. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Huang, A.C.; Zhang, W.; Zhang, G.; Wu, M.; Xu, W.; Yu, Z.; Yang, J.; Wang, B.; Sun, H.; et al. Exosomal PD-L1 Contributes to Immunosuppression and Is Associated with Anti-PD-1 Response. Nature 2018, 560, 382–386. [Google Scholar] [CrossRef] [PubMed]
- de Nigris, F.; Mancini, F.P.; Schiano, C.; Infante, T.; Zullo, A.; Minucci, P.B.; Al-Omran, M.; Giordano, A.; Napoli, C. Osteosarcoma Cells Induce Endothelial Cell Proliferation during Neo-Angiogenesis. J. Cell. Physiol. 2013, 228, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, M.; Jin, L.; Guo, P.; Zhang, Z.; Zhanghuang, C.; Tan, X.; Mi, T.; Liu, J.; Wu, X.; et al. Exosome Mimetics Derived from Bone Marrow Mesenchymal Stem Cells Deliver Doxorubicin to Osteosarcoma in Vitro and in Vivo. Drug Deliv. 2022, 29, 3291–3303. [Google Scholar] [CrossRef]
- Wei, H.; Chen, J.; Wang, S.; Fu, F.; Zhu, X.; Wu, C.; Liu, Z.; Zhong, G.; Lin, J. A Nanodrug Consisting of Doxorubicin and Exosome Derived from Mesenchymal Stem Cells for Osteosarcoma Treatment In Vitro. Int. J. Nanomed. 2019, 14, 8603–8610. [Google Scholar] [CrossRef]
- Shimbo, K.; Miyaki, S.; Ishitobi, H.; Kato, Y.; Kubo, T.; Shimose, S.; Ochi, M. Exosome-Formed Synthetic microRNA-143 Is Transferred to Osteosarcoma Cells and Inhibits Their Migration. Biochem. Biophys. Res. Commun. 2014, 445, 381–387. [Google Scholar] [CrossRef]
- Perut, F.; Roncuzzi, L.; Zini, N.; Massa, A.; Baldini, N. Extracellular Nanovesicles Secreted by Human Osteosarcoma Cells Promote Angiogenesis. Cancers 2019, 11, 779. [Google Scholar] [CrossRef]
- Baglio, S.R.; Lagerweij, T.; Pérez-Lanzón, M.; Ho, X.D.; Léveillé, N.; Melo, S.A.; Cleton-Jansen, A.-M.; Jordanova, E.S.; Roncuzzi, L.; Greco, M.; et al. Blocking Tumor-Educated MSC Paracrine Activity Halts Osteosarcoma Progression. Clin. Cancer Res. 2017, 23, 3721–3733. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Z.; Yang, X.; Lu, W.; Chen, Y.; Lin, Y.; Wang, J.; Lin, S.; Yun, J.-P. H3K27 Acetylation Activated-COL6A1 Promotes Osteosarcoma Lung Metastasis by Repressing STAT1 and Activating Pulmonary Cancer-Associated Fibroblasts. Theranostics 2021, 11, 1473–1492. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Liao, D.; Li, J.; Liu, W.; Wang, J.; Zeng, C.; Wang, X.; Cao, Z.; Zhang, R.; Li, M.; et al. Rab22a-NeoF1 Fusion Protein Promotes Osteosarcoma Lung Metastasis through Its Secretion into Exosomes. Signal Transduct. Target. Ther. 2021, 6, 59. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Liu, W.; Zhao, W.; Feng, S.; Duan, A.; Ji, C.; Shen, K.; Liu, W.; Zhou, J.; Jiang, D.; et al. Exosomal Transfer of LCP1 Promotes Osteosarcoma Cell Tumorigenesis and Metastasis by Activating the JAK2/STAT3 Signaling Pathway. Mol. Ther. Nucleic Acids 2020, 21, 900–915. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chu, Y.; Li, K.; Zhang, G.; Guo, Z.; Wu, X.; Qiu, C.; Li, Y.; Wan, X.; Sui, J.; et al. Exosomes Secreted by Adipose-Derived Mesenchymal Stem Cells Foster Metastasis and Osteosarcoma Proliferation by Increasing COLGALT2 Expression. Front. Cell Dev. Biol. 2020, 8, 353. [Google Scholar] [CrossRef] [PubMed]
- Sha, L.; Ma, D.; Chen, C. Exosome-Mediated Hic-5 Regulates Proliferation and Apoptosis of Osteosarcoma via Wnt/β-Catenin Signal Pathway. Aging 2020, 12, 23598–23608. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Liu, W.; He, B.; Wang, L.; Zhang, F.; Shu, H.; Sun, L. Exosomes Derived from Bone Marrow Mesenchymal Stem Cells Promote Osteosarcoma Development by Activating Oncogenic Autophagy. J. Bone Oncol. 2020, 21, 100280. [Google Scholar] [CrossRef]
- Macklin, R.; Wang, H.; Loo, D.; Martin, S.; Cumming, A.; Cai, N.; Lane, R.; Ponce, N.S.; Topkas, E.; Inder, K.; et al. Extracellular Vesicles Secreted by Highly Metastatic Clonal Variants of Osteosarcoma Preferentially Localize to the Lungs and Induce Metastatic Behaviour in Poorly Metastatic Clones. Oncotarget 2016, 7, 43570–43587. [Google Scholar] [CrossRef]
- Endo-Munoz, L.; Cai, N.; Cumming, A.; Macklin, R.; de Long, L.M.; Topkas, E.; Mukhopadhyay, P.; Hill, M.; Saunders, N.A. Progression of Osteosarcoma from a Non-Metastatic to a Metastatic Phenotype Is Causally Associated with Activation of an Autocrine and Paracrine uPA Axis. PLoS ONE 2015, 10, e0133592. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, H.; Sun, X.; Wang, X.; Ren, T.; Huang, Y.; Zhang, R.; Zheng, B.; Guo, W. Exosomal PD-L1 and N-Cadherin Predict Pulmonary Metastasis Progression for Osteosarcoma Patients. J. Nanobiotechnol. 2020, 18, 151. [Google Scholar] [CrossRef]
- Troyer, R.M.; Ruby, C.E.; Goodall, C.P.; Yang, L.; Maier, C.S.; Albarqi, H.A.; Brady, J.V.; Bathke, K.; Taratula, O.; Mourich, D.; et al. Exosomes from Osteosarcoma and Normal Osteoblast Differ in Proteomic Cargo and Immunomodulatory Effects on T Cells. Exp. Cell Res. 2017, 358, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Wang, L.; Wu, C.; Huang, L.; Ruan, Y.; Xue, W. Tumor-Derived Exosomes Induced M2 Macrophage Polarization and Promoted the Metastasis of Osteosarcoma Cells Through Tim-3. Arch. Med. Res. 2021, 52, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xue, L.; Wu, Y.; Wu, Q.; Ren, H.; Song, X. Exosomes Loaded with Programmed Death Ligand-1 Promote Tumor Growth by Immunosuppression in Osteosarcoma. Bioengineered 2021, 12, 9520–9530. [Google Scholar] [CrossRef] [PubMed]
- Torreggiani, E.; Roncuzzi, L.; Perut, F.; Zini, N.; Baldini, N. Multimodal Transfer of MDR by Exosomes in Human Osteosarcoma. Int. J. Oncol. 2016, 49, 189–196. [Google Scholar] [CrossRef]
- Zhan, H.; Xiao, J.; Wang, P.; Mo, F.; Li, K.; Guo, F.; Yu, X.; Liu, X.; Zhang, B.; Dai, M.; et al. Exosomal CTCF Confers Cisplatin Resistance in Osteosarcoma by Promoting Autophagy via the IGF2-AS/miR-579-3p/MSH6 Axis. J. Oncol. 2022, 2022, e9390611. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, A.; Fujiwara, T.; Uotani, K.; Morita, T.; Kiyono, M.; Yokoo, S.; Hasei, J.; Nakata, E.; Kunisada, T.; Ozaki, T. Clinical and Functional Significance of Intracellular and Extracellular microRNA-25-3p in Osteosarcoma. Acta Med. Okayama 2018, 72, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Cao, H.; Gu, G.; Hou, D.; You, Y.; Li, X.; Chen, Y.; Jiao, G. Exosomal MiR-199a-5p Inhibits Tumorigenesis and Angiogenesis by Targeting VEGFA in Osteosarcoma. Front. Oncol. 2022, 12, 884559. [Google Scholar] [CrossRef] [PubMed]
- Jerez, S.; Araya, H.; Hevia, D.; Irarrázaval, C.E.; Thaler, R.; van Wijnen, A.J.; Galindo, M. Extracellular Vesicles from Osteosarcoma Cell Lines Contain miRNAs Associated with Cell Adhesion and Apoptosis. Gene 2019, 710, 246–257. [Google Scholar] [CrossRef]
- Han, F.; Pu, P.; Wang, C.; Ding, X.; Zhu, Z.; Xiang, W.; Wang, W. Osteosarcoma Cell-Derived Exosomal miR-1307 Promotes Tumorgenesis via Targeting AGAP1. Biomed. Res. Int. 2021, 2021, 7358153. [Google Scholar] [CrossRef]
- Ye, Z.; Zheng, Z.; Peng, L. MicroRNA Profiling of Serum Exosomes in Patients with Osteosarcoma by High-Throughput Sequencing. J. Investig. Med. 2020, 68, 893–901. [Google Scholar] [CrossRef]
- Gong, L.; Bao, Q.; Hu, C.; Wang, J.; Zhou, Q.; Wei, L.; Tong, L.; Zhang, W.; Shen, Y. Exosomal miR-675 from Metastatic Osteosarcoma Promotes Cell Migration and Invasion by Targeting CALN1. Biochem. Biophys. Res. Commun. 2018, 500, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-W.; Wu, X.-F.; Gu, X.-J.; Jiang, X.-H. Exosomal miR-1228 from Cancer-Associated Fibroblasts Promotes Cell Migration and Invasion of Osteosarcoma by Directly Targeting SCAI. Oncol. Res. 2019, 27, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Tang, H.; Zhang, Y.; Zhang, Z.; Huang, P.; Zhu, J. Bone Marrow-Derived Mesenchymal Stem Cell-Derived Exosomal microRNA-208a Promotes Osteosarcoma Cell Proliferation, Migration, and Invasion. J. Cell. Physiol. 2020, 235, 4734–4745. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, B.; Duan, A.; Shen, K.; Zhang, Q.; Tang, X.; Wei, Y.; Tang, J.; Zhang, S. Exosomal Transfer of miR-769-5p Promotes Osteosarcoma Proliferation and Metastasis by Targeting DUSP16. Cancer Cell Int. 2021, 21, 541. [Google Scholar] [CrossRef]
- Zhang, K.; Dong, C.; Chen, M.; Yang, T.; Wang, X.; Gao, Y.; Wang, L.; Wen, Y.; Chen, G.; Wang, X.; et al. Extracellular Vesicle-Mediated Delivery of miR-101 Inhibits Lung Metastasis in Osteosarcoma. Theranostics 2020, 10, 411–425. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, J.; Ren, T.; Huang, Y.; Liang, X.; Yu, Y.; Wang, W.; Niu, J.; Guo, W. Bone Marrow Mesenchymal Stem Cell-Derived Exosomal miR-206 Inhibits Osteosarcoma Progression by Targeting TRA2B. Cancer Lett. 2020, 490, 54–65. [Google Scholar] [CrossRef]
- Zhou, J.; Xu, L.; Yang, P.; Lu, Y.; Lin, S.; Yuan, G. The Exosomal Transfer of Human Bone Marrow Mesenchymal Stem Cell-Derived miR-1913 Inhibits Osteosarcoma Progression by Targeting NRSN2. Am. J. Transl. Res. 2021, 13, 10178–10192. [Google Scholar]
- Xu, Z.; Zhou, X.; Wu, J.; Cui, X.; Wang, M.; Wang, X.; Gao, Z. Mesenchymal Stem Cell-Derived Exosomes Carrying microRNA-150 Suppresses the Proliferation and Migration of Osteosarcoma Cells via Targeting IGF2BP1. Transl. Cancer Res. 2020, 9, 5323–5335. [Google Scholar] [CrossRef]
- Wu, C.; Li, Z.; Feng, G.; Wang, L.; Xie, J.; Jin, Y.; Wang, L.; Liu, S. Tumor Suppressing Role of Serum-Derived Exosomal microRNA-15a in Osteosarcoma Cells through the GATA Binding Protein 2/Murine Double Minute 2 Axis and the P53 Signaling Pathway. Bioengineered 2021, 12, 8378–8395. [Google Scholar] [CrossRef]
- Jiang, M.; Jike, Y.; Liu, K.; Gan, F.; Zhang, K.; Xie, M.; Zhang, J.; Chen, C.; Zou, X.; Jiang, X.; et al. Exosome-Mediated miR-144-3p Promotes Ferroptosis to Inhibit Osteosarcoma Proliferation, Migration, and Invasion through Regulating ZEB1. Mol. Cancer 2023, 22, 113. [Google Scholar] [CrossRef]
- Liu, W.; Long, Q.; Zhang, W.; Zeng, D.; Hu, B.; Liu, S.; Chen, L. miRNA-221-3p Derived from M2-Polarized Tumor-Associated Macrophage Exosomes Aggravates the Growth and Metastasis of Osteosarcoma through SOCS3/JAK2/STAT3 Axis. Aging 2021, 13, 19760–19775. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.-C.; Huang, J.-Y.; Wei, Z.-Y.; Li, Z.-X.; Guo, S.-C. EWSAT1 Acts in Concert with Exosomes in Osteosarcoma Progression and Tumor-Induced Angiogenesis: The “Double Stacking Effect”. Adv. Biosyst. 2020, 4, 2000152. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lin, S.; Xie, X.; Zhu, H.; Fan, T.; Wang, S. Highly Enriched Exosomal lncRNA OIP5-AS1 Regulates Osteosarcoma Tumor Angiogenesis and Autophagy through miR-153 and ATG5. Am. J. Transl. Res. 2021, 13, 4211–4223. [Google Scholar] [PubMed]
- Zhao, W.; Qin, P.; Zhang, D.; Cui, X.; Gao, J.; Yu, Z.; Chai, Y.; Wang, J.; Li, J. Long Non-Coding RNA PVT1 Encapsulated in Bone Marrow Mesenchymal Stem Cell-Derived Exosomes Promotes Osteosarcoma Growth and Metastasis by Stabilizing ERG and Sponging miR-183-5p. Aging 2019, 11, 9581–9596. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, X.; Jiang, N.; Xie, X.; Liu, N.; Liu, J.; Shen, J.; Peng, T. Exosome-Transmitted Linc00852 Associated with Receptor Tyrosine Kinase AXL Dysregulates the Proliferation and Invasion of Osteosarcoma. Cancer Med. 2020, 9, 6354–6366. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, Y.; Wang, J.; Han, Y.; Ren, T.; Huang, Y.; Chen, C.; Huang, Q.; Wang, W.; Niu, J.; et al. Macrophages-Derived Exosomal lncRNA LIFR-AS1 Promotes Osteosarcoma Cell Progression via miR-29a/NFIA Axis. Cancer Cell Int. 2021, 21, 192. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, J.; Ren, T.; Huang, Y.; Yu, Y.; Chen, C.; Huang, Q.; Guo, W. LncRNA CASC15 Is Upregulated in Osteosarcoma Plasma Exosomes and CASC15 Knockdown Inhibits Osteosarcoma Progression by Regulating miR-338-3p/RAB14 Axis. Onco Targets Ther. 2020, 13, 12055–12066. [Google Scholar] [CrossRef]
- Li, F.; Chen, X.; Shang, C.; Ying, Q.; Zhou, X.; Zhu, R.; Lu, H.; Hao, X.; Dong, Q.; Jiang, Z. Bone Marrow Mesenchymal Stem Cells-Derived Extracellular Vesicles Promote Proliferation, Invasion and Migration of Osteosarcoma Cells via the lncRNA MALAT1/miR-143/NRSN2/Wnt/β-Catenin Axis. Onco Targets Ther. 2021, 14, 737–749. [Google Scholar] [CrossRef]
- Zhu, G.; Xia, Y.; Zhao, Z.; Li, A.; Li, H.; Xiao, T. LncRNA XIST from the Bone Marrow Mesenchymal Stem Cell Derived Exosome Promotes Osteosarcoma Growth and Metastasis through miR-655/ACLY Signal. Cancer Cell Int. 2022, 22, 330. [Google Scholar] [CrossRef]
- Wang, B.; Wang, X.; Li, P.; Niu, X.; Liang, X.; Liu, G.; Liu, Z.; Ge, H. Osteosarcoma Cell-Derived Exosomal ELFN1-AS1 Mediates Macrophage M2 Polarization via Sponging miR-138-5p and miR-1291 to Promote the Tumorgenesis of Osteosarcoma. Front. Oncol. 2022, 12, 881022. [Google Scholar] [CrossRef]
- Hu, X.; Wen, Y.; Tan, L.; Wang, J.; Tang, F.; Wang, Y.; Zheng, C.; Zhang, Y.; Gong, T.; Min, L. Exosomal Long Non-Coding RNA ANCR Mediates Drug Resistance in Osteosarcoma. Front. Oncol. 2022, 11, 735254. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Pei, Y.; Wang, W.; Liu, F.; Zheng, K.; Zhang, X. Extracellular Nanovesicles-transmitted Circular RNA Has_circ_0000190 Suppresses Osteosarcoma Progression. J. Cell. Mol. Med. 2020, 24, 2202–2214. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Lin, Y.; Mi, C. Cisplatin-Resistant Osteosarcoma Cell-Derived Exosomes Confer Cisplatin Resistance to Recipient Cells in an Exosomal Circ_103801-Dependent Manner. Cell Biol. Int. 2021, 45, 858–868. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-W.; Liu, S.-C.; Sun, H.-L.; Huang, T.-Y.; Chan, C.-H.; Yang, C.-Y.; Yeh, H.-I.; Huang, Y.-L.; Chou, W.-Y.; Lin, Y.-M.; et al. CCL5/CCR5 Axis Induces Vascular Endothelial Growth Factor-Mediated Tumor Angiogenesis in Human Osteosarcoma Microenvironment. Carcinogenesis 2015, 36, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Klotzsche-von Ameln, A.; Prade, I.; Grosser, M.; Kettelhake, A.; Rezaei, M.; Chavakis, T.; Flamme, I.; Wielockx, B.; Breier, G. PHD4 Stimulates Tumor Angiogenesis in Osteosarcoma Cells via TGF-α. Mol. Cancer Res. 2013, 11, 1337–1348. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, H.F.; Brown, L.F.; Detmar, M.; Dvorak, A.M. Vascular Permeability Factor/Vascular Endothelial Growth Factor, Microvascular Hyperpermeability, and Angiogenesis. Am. J. Pathol. 1995, 146, 1029–1039. [Google Scholar] [PubMed]
- Folkman, J. What Is the Evidence That Tumors Are Angiogenesis Dependent? JNCI J. Natl. Cancer Inst. 1990, 82, 4–7. [Google Scholar] [CrossRef]
- Medina, P.P.; Nolde, M.; Slack, F.J. OncomiR Addiction in an in Vivo Model of microRNA-21-Induced Pre-B-Cell Lymphoma. Nature 2010, 467, 86–90. [Google Scholar] [CrossRef]
- Si, M.-L.; Zhu, S.; Wu, H.; Lu, Z.; Wu, F.; Mo, Y.-Y. miR-21-Mediated Tumor Growth. Oncogene 2007, 26, 2799–2803. [Google Scholar] [CrossRef]
- Chan, J.A.; Krichevsky, A.M.; Kosik, K.S. MicroRNA-21 Is an Antiapoptotic Factor in Human Glioblastoma Cells. Cancer Res. 2005, 65, 6029–6033. [Google Scholar] [CrossRef]
- Qi, J.; Zhang, R.; Wang, Y. Exosomal miR-21-5p Derived from Bone Marrow Mesenchymal Stem Cells Promote Osteosarcoma Cell Proliferation and Invasion by Targeting PIK3R1. J. Cell. Mol. Med. 2021, 25, 11016–11030. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xu, B.; Miu, X.; Deng, Z.; Liao, H.; Hao, L. Inhibition of miRNA-21 Attenuates the Proliferation and Metastasis of Human Osteosarcoma by Upregulating PTEN. Exp. Ther. Med. 2018, 15, 1036–1040. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Li, L.; Lu, Y.; Yu, X.; Chen, H.; Yin, Q.; Zhang, Y. miRNA-21 Inhibition Inhibits Osteosarcoma Cell Proliferation by Targeting PTEN and Regulating the TGF-Β1 Signaling Pathway. Oncol. Lett. 2018, 16, 4337–4342. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhang, J.; Quan, H.; Wang, J.; Liang, Y. MicroRNA-143 Expression Inhibits the Growth and the Invasion of Osteosarcoma. J. Orthop. Surg. Res. 2022, 17, 236. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Lv, B.; Li, Y.; Cheng, Q.; Su, C.; Yin, G. MiR-143 Regulates the Proliferation and Migration of Osteosarcoma Cells through Targeting MAPK7. Arch. Biochem. Biophys. 2017, 630, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wu, H.; Li, Y.; Pan, H.; Meng, T.; Wang, X. MicroRNA-143 Promotes Apoptosis of Osteosarcoma Cells by Caspase-3 Activation via Targeting Bcl-2. Biomed. Pharmacother. 2016, 80, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Osaki, M.; Takeshita, F.; Sugimoto, Y.; Kosaka, N.; Yamamoto, Y.; Yoshioka, Y.; Kobayashi, E.; Yamada, T.; Kawai, A.; Inoue, T.; et al. MicroRNA-143 Regulates Human Osteosarcoma Metastasis by Regulating Matrix Metalloprotease-13 Expression. Mol. Ther. 2011, 19, 1123–1130. [Google Scholar] [CrossRef] [PubMed]
- Holenstein, C.N.; Horvath, A.; Schär, B.; Schoenenberger, A.D.; Bollhalder, M.; Goedecke, N.; Bartalena, G.; Otto, O.; Herbig, M.; Guck, J.; et al. The Relationship between Metastatic Potential and in Vitro Mechanical Properties of Osteosarcoma Cells. MBoC 2019, 30, 887–898. [Google Scholar] [CrossRef]
- Wu, S.C.; Kim, A.; Gu, Y.; Martinez, D.I.; Zocchi, L.; Chen, C.C.; Lopez, J.; Salcido, K.; Singh, S.; Wu, J.; et al. UHRF1 Overexpression Promotes Osteosarcoma Metastasis through Altered Exosome Production and AMPK/SEMA3E Suppression. Oncogenesis 2022, 11, 1–12. [Google Scholar] [CrossRef]
- Deng, C.; Xu, Y.; Fu, J.; Zhu, X.; Chen, H.; Xu, H.; Wang, G.; Song, Y.; Song, G.; Lu, J.; et al. Reprograming the Tumor Immunologic Microenvironment Using Neoadjuvant Chemotherapy in Osteosarcoma. Cancer Sci. 2020, 111, 1899–1909. [Google Scholar] [CrossRef]
- Dumars, C.; Ngyuen, J.-M.; Gaultier, A.; Lanel, R.; Corradini, N.; Gouin, F.; Heymann, D.; Heymann, M.-F. Dysregulation of Macrophage Polarization Is Associated with the Metastatic Process in Osteosarcoma. Oncotarget 2016, 7, 78343–78354. [Google Scholar] [CrossRef] [PubMed]
- Qian, B.-Z.; Pollard, J.W. Macrophage Diversity Enhances Tumor Progression and Metastasis. Cell 2010, 141, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the Full Spectrum of Macrophage Activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Wieckowski, E.; Taylor, D.D.; Reichert, T.E.; Watkins, S.; Whiteside, T.L. Fas Ligand–Positive Membranous Vesicles Isolated from Sera of Patients with Oral Cancer Induce Apoptosis of Activated T Lymphocytes. Clin. Cancer Res. 2005, 11, 1010–1020. [Google Scholar] [CrossRef] [PubMed]
- Dorayappan, K.D.P.; Wanner, R.; Wallbillich, J.J.; Saini, U.; Zingarelli, R.; Suarez, A.A.; Cohn, D.E.; Selvendiran, K. Hypoxia-Induced Exosomes Contribute to a More Aggressive and Chemoresistant Ovarian Cancer Phenotype: A Novel Mechanism Linking STAT3/Rab Proteins. Oncogene 2018, 37, 3806–3821. [Google Scholar] [CrossRef] [PubMed]
- Lobb, R.J.; van Amerongen, R.; Wiegmans, A.; Ham, S.; Larsen, J.E.; Möller, A. Exosomes Derived from Mesenchymal Non-Small Cell Lung Cancer Cells Promote Chemoresistance. Int. J. Cancer 2017, 141, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Yan, C.; Mu, L.; Huang, K.; Li, X.; Tao, D.; Wu, Y.; Qin, J. Fibroblast-Derived Exosomes Contribute to Chemoresistance through Priming Cancer Stem Cells in Colorectal Cancer. PLoS ONE 2015, 10, e0125625. [Google Scholar] [CrossRef]
- Ji, R.; Zhang, B.; Zhang, X.; Xue, J.; Yuan, X.; Yan, Y.; Wang, M.; Zhu, W.; Qian, H.; Xu, W. Exosomes Derived from Human Mesenchymal Stem Cells Confer Drug Resistance in Gastric Cancer. Cell Cycle 2015, 14, 2473–2483. [Google Scholar] [CrossRef]
- Shao, H.; Chung, J.; Lee, K.; Balaj, L.; Min, C.; Carter, B.S.; Hochberg, F.H.; Breakefield, X.O.; Lee, H.; Weissleder, R. Chip-Based Analysis of Exosomal mRNA Mediating Drug Resistance in Glioblastoma. Nat. Commun. 2015, 6, 6999. [Google Scholar] [CrossRef]
- Chen, W.; Liu, X.; Lv, M.; Chen, L.; Zhao, J.; Zhong, S.; Ji, M.; Hu, Q.; Luo, Z.; Wu, J.; et al. Exosomes from Drug-Resistant Breast Cancer Cells Transmit Chemoresistance by a Horizontal Transfer of microRNAs. PLoS ONE 2014, 9, e95240. [Google Scholar] [CrossRef]
- Yati, S.; Silathapanasakul, A.; Thakaeng, C.; Chanasakulniyom, M.; Songtawee, N.; Porntadavity, S.; Pothacharoen, P.; Pruksakorn, D.; Kongtawelert, P.; Yenchitsomanus, P.; et al. Extracellular Vesicle-Mediated IL-1 Signaling in Response to Doxorubicin Activates PD-L1 Expression in Osteosarcoma Models. Cells 2022, 11, 1042. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Quan, L.; Zhang, C.; Liu, A.; Tong, D.; Wang, J. Over-Activated PD-1/PD-L1 Axis Facilitates the Chemoresistance of Diffuse Large B-Cell Lymphoma Cells to the CHOP Regimen. Oncol. Lett. 2018, 15, 3321–3328. [Google Scholar] [CrossRef] [PubMed]
- Black, M.; Barsoum, I.B.; Truesdell, P.; Cotechini, T.; Macdonald-Goodfellow, S.K.; Petroff, M.; Siemens, D.R.; Koti, M.; Craig, A.W.B.; Graham, C.H. Activation of the PD-1/PD-L1 Immune Checkpoint Confers Tumor Cell Chemoresistance Associated with Increased Metastasis. Oncotarget 2016, 7, 10557–10567. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Pang, J.; Peng, Y.; Molina, J.R.; Yang, P.; Liu, S. Elevated Cellular PD1/PD-L1 Expression Confers Acquired Resistance to Cisplatin in Small Cell Lung Cancer Cells. PLoS ONE 2016, 11, e0162925. [Google Scholar] [CrossRef]
- Bao, Q.; Gong, L.; Wang, J.; Wen, J.; Shen, Y.; Zhang, W. Extracellular Vesicle RNA Sequencing Reveals Dramatic Transcriptomic Alterations Between Metastatic and Primary Osteosarcoma in a Liquid Biopsy Approach. Ann. Surg. Oncol. 2018, 25, 2642–2651. [Google Scholar] [CrossRef]
- Fujiwara, T.; Uotani, K.; Yoshida, A.; Morita, T.; Nezu, Y.; Kobayashi, E.; Yoshida, A.; Uehara, T.; Omori, T.; Sugiu, K.; et al. Clinical Significance of Circulating miR-25-3p as a Novel Diagnostic and Prognostic Biomarker in Osteosarcoma. Oncotarget 2017, 8, 33375–33392. [Google Scholar] [CrossRef]
- Xu, J.-F.; Wang, Y.-P.; Zhang, S.-J.; Chen, Y.; Gu, H.-F.; Dou, X.-F.; Xia, B.; Bi, Q.; Fan, S.-W. Exosomes Containing Differential Expression of microRNA and mRNA in Osteosarcoma That Can Predict Response to Chemotherapy. Oncotarget 2017, 8, 75968–75978. [Google Scholar] [CrossRef]
- Cuscino, N.; Raimondi, L.; De Luca, A.; Carcione, C.; Russelli, G.; Conti, L.; Baldi, J.; Conaldi, P.G.; Giavaresi, G.; Gallo, A. Gathering Novel Circulating Exosomal microRNA in Osteosarcoma Cell Lines and Possible Implications for the Disease. Cancers 2019, 11, 1924. [Google Scholar] [CrossRef]
- Huo, S.; Dou, D. Circ_0056285 Regulates Proliferation, Apoptosis and Glycolysis of Osteosarcoma Cells via miR-1244/TRIM44 Axis. Cancer Manag. Res. 2021, 13, 1257–1270. [Google Scholar] [CrossRef]
- Wang, L.; Wu, J.; Song, S.; Chen, H.; Hu, Y.; Xu, B.; Liu, J. Plasma Exosome-Derived Sentrin SUMO-Specific Protease 1: A Prognostic Biomarker in Patients with Osteosarcoma. Front. Oncol. 2021, 11, 625109. [Google Scholar] [CrossRef]
- Han, Z.; Peng, C.; Yi, J.; Wang, Y.; Liu, Q.; Yang, Y.; Long, S.; Qiao, L.; Shen, Y. Matrix-Assisted Laser Desorption Ionization Mass Spectrometry Profiling of Plasma Exosomes Evaluates Osteosarcoma Metastasis. iScience 2021, 24, 102906. [Google Scholar] [CrossRef] [PubMed]
- Cambier, L.; Stachelek, K.; Triska, M.; Jubran, R.; Huang, M.; Li, W.; Zhang, J.; Li, J.; Cobrinik, D. Extracellular Vesicle-Associated Repetitive Element DNAs as Candidate Osteosarcoma Biomarkers. Sci. Rep. 2021, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Yi, J.; Yang, Y.; Li, D.; Peng, C.; Long, S.; Peng, X.; Shen, Y.; Liu, B.; Qiao, L. SERS and MALDI-TOF MS Based Plasma Exosome Profiling for Rapid Detection of Osteosarcoma. Analyst 2021, 146, 6496–6505. [Google Scholar] [CrossRef] [PubMed]
- Thakur, B.K.; Zhang, H.; Becker, A.; Matei, I.; Huang, Y.; Costa-Silva, B.; Zheng, Y.; Hoshino, A.; Brazier, H.; Xiang, J.; et al. Double-Stranded DNA in Exosomes: A Novel Biomarker in Cancer Detection. Cell Res. 2014, 24, 766–769. [Google Scholar] [CrossRef] [PubMed]
- Elliott, R.O.; He, M. Unlocking the Power of Exosomes for Crossing Biological Barriers in Drug Delivery. Pharmaceutics 2021, 13, 122. [Google Scholar] [CrossRef] [PubMed]
- Yong, T.; Zhang, X.; Bie, N.; Zhang, H.; Zhang, X.; Li, F.; Hakeem, A.; Hu, J.; Gan, L.; Santos, H.A.; et al. Tumor Exosome-Based Nanoparticles Are Efficient Drug Carriers for Chemotherapy. Nat. Commun. 2019, 10, 3838. [Google Scholar] [CrossRef]
- Zhu, X.; Badawi, M.; Pomeroy, S.; Sutaria, D.S.; Xie, Z.; Baek, A.; Jiang, J.; Elgamal, O.A.; Mo, X.; Perle, K.L.; et al. Comprehensive Toxicity and Immunogenicity Studies Reveal Minimal Effects in Mice Following Sustained Dosing of Extracellular Vesicles Derived from HEK293T Cells. J. Extracell. Vesicles 2017, 6, 1324730. [Google Scholar] [CrossRef]
- Chen, C.C.; Liu, L.; Ma, F.; Wong, C.W.; Guo, X.E.; Chacko, J.V.; Farhoodi, H.P.; Zhang, S.X.; Zimak, J.; Ségaliny, A.; et al. Elucidation of Exosome Migration Across the Blood–Brain Barrier Model In Vitro. Cell. Mol. Bioeng. 2016, 9, 509–529. [Google Scholar] [CrossRef]
- Segal, M.; Biscans, A.; Gilles, M.-E.; Anastasiadou, E.; Luca, R.D.; Lim, J.; Khvorova, A.; Slack, F.J. Hydrophobically Modified Let-7b miRNA Enhances Biodistribution to NSCLC and Downregulates HMGA2 In Vivo. Mol. Ther. Nucleic Acids 2020, 19, 267–277. [Google Scholar] [CrossRef]
- Abello, J.; Nguyen, T.D.T.; Marasini, R.; Aryal, S.; Weiss, M.L. Biodistribution of Gadolinium- and near Infrared-Labeled Human Umbilical Cord Mesenchymal Stromal Cell-Derived Exosomes in Tumor Bearing Mice. Theranostics 2019, 9, 2325–2345. [Google Scholar] [CrossRef]
- Notaro, A.; Emanuele, S.; Geraci, F.; D’Anneo, A.; Lauricella, M.; Calvaruso, G.; Giuliano, M. WIN55,212-2-Induced Expression of Mir-29b1 Favours the Suppression of Osteosarcoma Cell Migration in a SPARC-Independent Manner. Int. J. Mol. Sci. 2019, 20, 5235. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wu, W.; Jing, D.; Yang, L.; Guo, H.; Wang, L.; Zhang, W.; Pu, F.; Shao, Z. Engineered Exosome as Targeted lncRNA MEG3 Delivery Vehicles for Osteosarcoma Therapy. J. Control. Release 2022, 343, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Ruijin Hospital. Construction and Clinical Application of Microfluidic Exosome Chip for Early Diagnosis of Pulmonary Metastasis of Osteosarcoma; Ruijin Hospital: Shanghai, China, 2022. Available online: https://clinicaltrials.gov/study/NCT05101655 (accessed on 21 August 2023).
- Shen, Y. A Pilot Study of Circulating Exosome RNA as Diagnostic and Prognostic Markers in Lung Metastases of Primary High-Grade Osteosarcoma; Ruijin Hospital: Shanghai, China, 2023. Available online: https://clinicaltrials.gov/study/NCT03108677 (accessed on 21 August 2023).

{kind=link}
{kind=link}
{kind=link}
| Cargo Type | Parent Cell | Target Cells | Exosomal Cargo | Biological Function | Reference |
|---|---|---|---|---|---|
| protein | OS cells | HUVEC | VEGF-A, IL-6, IL-8 | Promote angiogenesis | [33] |
| OS cells | Macrophages | IL-10, TGFβ2, and CCL2 | Facilitate immunosuppressive TME | [34] | |
| OS cells | HUVEC | VEGF, PDGF-AA, CD26, CD105, endostatin, ET-1, PAI-1, THBS1, TIMP-1, PEDF, uPA, ANG-2, TF3, PTX3, and HB-EGF | Promote angiogenesis | [40] | |
| OS cells | MSC | TGF-β | Induce secretion of proinflammatory cytokine IL-6 | [41] | |
| OS cells | CAF | COL6A1 | Promote OS migration, invasion, and lung metastasis | [42] | |
| OS cells | Macrophages | Rab22a-NeoF1 | Promote lung metastasis | [43] | |
| BMSC | OS cells | LCP1 | Promote OS proliferation, migration, invasion, and metastasis | [44] | |
| Adipose MSC | OS cells | COLGALT2 | Promote OS proliferation, migration, and invasion | [45] | |
| OS cells | OS cells | Hic-5 | Promote OS proliferation and inhibit apoptosis | [46] | |
| BMSCs | OS cells | ATG5 | Promote OS proliferation, migration, invasion, and lung metastasis | [47] | |
| OS cells | OS cells | NPM1, CCT2, CCT4, CCT6A, CCT8, VIM, CLTC, COL6A2, HNRNPC, PKM, ACTN4, MYH10, PAICS, VCP, ANXA1, and ACLY. | Promote OS migration and invasion | [48] | |
| OS cells | OS cells | uPA | Promote OS migration | [49] | |
| OS cells | OS cells | PD-L1, N-cadherin | Promote migration, invasion, and pulmonary metastasis | [50] | |
| OS cells | PBMC | TGF-β | Reduce T cell proliferation and T cell activity | [51] | |
| OS cells | Macrophages | TIM-3 | Promote OS migration, invasion, EMT, M2 macrophage polarization, and lung metastasis | [52] | |
| OS cells | CD3 + T cells | PD-L1 | Promote tumor growth | [53] | |
| OS cells | OS cells | MDR-1 P-gp | Induce a doxorubicin-resistant phenotype | [54] | |
| OS cells | OS cells | CTCF | Enhance CDDP resistance of OS cells | [55] | |
| miRNA | OS cells | OS cells | miR-21-5p and miR-148a-3p | Promote angiogenesis | [33] |
| OS cells | HUVEC | miR-10b-5p, miR-106a-5p, miR-125b-5p, miR-143-3p, miR-145-5p, miR-146a-5p, miR-150-5p, miR-21-5p, miR-26a-5p, miR-27a-3p, miR-382-5p, miR-92a-3p, and miR-93-5p | Promote angiogenesis | [40] | |
| OS cells | HUVEC | miR-25-3p | Promoted invasion and angiogenesis | [56] | |
| OS cells | HUVEC | miR-199-5p | Inhibit HUVEC proliferation, migration, and neovascularization | [57] | |
| OS cells | NA | miR-21-5p, miR-143-3p, miR-148a-3p, and 181a-5p | Regulate cell adhesion and apoptosis | [58] | |
| OS cells | OS cells | miR-1307 | Promoted OS proliferation, migration, and invasion | [59] | |
| OS cells | OS cells | miR-195-3p | Promote OS proliferation and invasion | [60] | |
| OS cells | Osteoblasts | miR-675 | Promote OS migration and invasion | [61] | |
| CAF | OS cells | miR-1228 | Promote OS migration and invasion | [62] | |
| BMSC | OS cells | miR-208a | Promote OS viability, clonogenicity, and migration | [63] | |
| BMSC | OS cells | miR-769-5p | Promote OS proliferation and metastasis | [64] | |
| AD-MSC | OS cells | miR-101 | Inhibit OS migration and invasion | [65] | |
| BMSC | OS cells | miR-206 | Inhibit OS proliferation, migration, and invasion; induce apoptosis; suppresses tumor growth and metastasis | [66] | |
| BMSC | OS cells | miR-1913 | Inhibit OS proliferation, migration, and invasion | [67] | |
| BMSC | OS cells | miR-150 | Inhibit OS cell proliferation, migration, and invasion; induce apoptosis and suppress tumor growth | [68] | |
| Serum | OS cells | miR-15a | Inhibit OS cell viability; promotes apoptosis and cell cycle arrests | [69] | |
| OS cells | OS cells | miR-144-3p | inhibits OS proliferation, invasion and migration | [70] | |
| OS cells | Macrophages | miR-221-3p | Enhance M2 polarization of TAMs; promote OS cell viability, migration, invasion; reduce apoptosis | [71] | |
| lncRNA | OS cells | human microvascular endothelial cell line | lncRNA EWSAT-1 | Promote angiogenesis | [72] |
| OS cells | HUVEC | lncRNA OIP5-AS1 | Promote angiogenesis | [73] | |
| BMSC | OS cells | lncRNA PVT1 | Promote tumor growth and metastasis | [74] | |
| OS cells | OS cells | lncRNA LINC00852 | Promote OS proliferation, migration, invasion, and metastasis | [75] | |
| Macrophages | OS cells | lncRNA LIFR-AS1 | Promote OS proliferation, migration, invasion, and metastasis | [76] | |
| Plasma | OS cells | lncRNA CASC15 | Promote tumor growth and metastasis | [77] | |
| BMSCs | OS cells | lncRNA MALAT1 | Promote OS proliferation, migration, invasion, and tumor growth | [78] | |
| BMSCs | OS cells | lncRNA XIST | Promote OS proliferation, migration, and invasion | [79] | |
| OS cells | Macrophages | lncRNA ELFN1-AS1 | Promote proliferation, migration, and invasion; promote macrophage M2 polarization and tumor growth | [80] | |
| OS cells | OS cells | lncRNA ANCR | Induce DOX chemoresistance | [81] | |
| circRNA | OS cells | OS cells | circ_0000190 | Inhibit OS proliferation, migration, and invasion | [82] |
| OS cells | OS cells | circ_103801 | Enhance chemoresistance to CDDP | [83] |
| Exosome Source | Exosomal Cargo | Biomarker | Number of Patients | Reference |
|---|---|---|---|---|
| Serum | miR-25-3p | Diagnosis and prognosis | 45 | [56] |
| Plasma | miR-92a-3p, miR-130a-3p, miR-195-3p, miR-335-5p, let-7i-3p | Diagnosis | 25 | [60] |
| Serum | miR-675 | Diagnosis | 8 | [61] |
| Plasma | miR-101 | Diagnosis | 41 | [65] |
| Tissue | miR-101 | Diagnosis | 129 | [65] |
| Serum | lncRNA CASC15 | Diagnosis | 5 | [77] |
| Tissue | lncRNA CASC15 | Diagnosis | 30 | [77] |
| Plasma | circ_0000190 | Diagnosis | 60 | [82] |
| Serum | TGFβ | Prognosis | 10 | [41] |
| Plasma | PD-L1, N-cadherin | Prognosis | 70 | [50] |
| Serum | circ_103801 | Prognosis | 43 | [83] |
| Serum | Annexin2, CDC5L, Smad2, P27, CIP4, MTAP, PEDF, WWOX | Prognosis | 93 | [117] |
| Serum | miR-135b, miR-148a, miR-27a, miR-9, miR-124, miR-133a, miR-199a-3p, miR-385 | Prognosis | 93 | [117] |
| Serum | circ_0056285 | Diagnosis | 35 | [119] |
| Plasma | SENP1 | Prognosis | 146 | [120] |
| Plasma | IGLV2-23, IGLV4-3, IGLV1-51, IGKV3-15, IGHV4-4, IGLV4-60, HBA1 | Prognosis | 40 | [121] |
| Serum | HSATI, HSATII, LINE1-P1, Charlie 3 | Diagnosis | 12 | [122] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.C.; Benavente, C.A. Exploring the Impact of Exosomal Cargos on Osteosarcoma Progression: Insights into Therapeutic Potential. Int. J. Mol. Sci. 2024, 25, 568. https://doi.org/10.3390/ijms25010568
Chen CC, Benavente CA. Exploring the Impact of Exosomal Cargos on Osteosarcoma Progression: Insights into Therapeutic Potential. International Journal of Molecular Sciences. 2024; 25(1):568. https://doi.org/10.3390/ijms25010568
Chicago/Turabian StyleChen, Claire C., and Claudia A. Benavente. 2024. "Exploring the Impact of Exosomal Cargos on Osteosarcoma Progression: Insights into Therapeutic Potential" International Journal of Molecular Sciences 25, no. 1: 568. https://doi.org/10.3390/ijms25010568
APA StyleChen, C. C., & Benavente, C. A. (2024). Exploring the Impact of Exosomal Cargos on Osteosarcoma Progression: Insights into Therapeutic Potential. International Journal of Molecular Sciences, 25(1), 568. https://doi.org/10.3390/ijms25010568