
Wnt3a Facilitates SARS-CoV-2 Pseudovirus Entry into Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
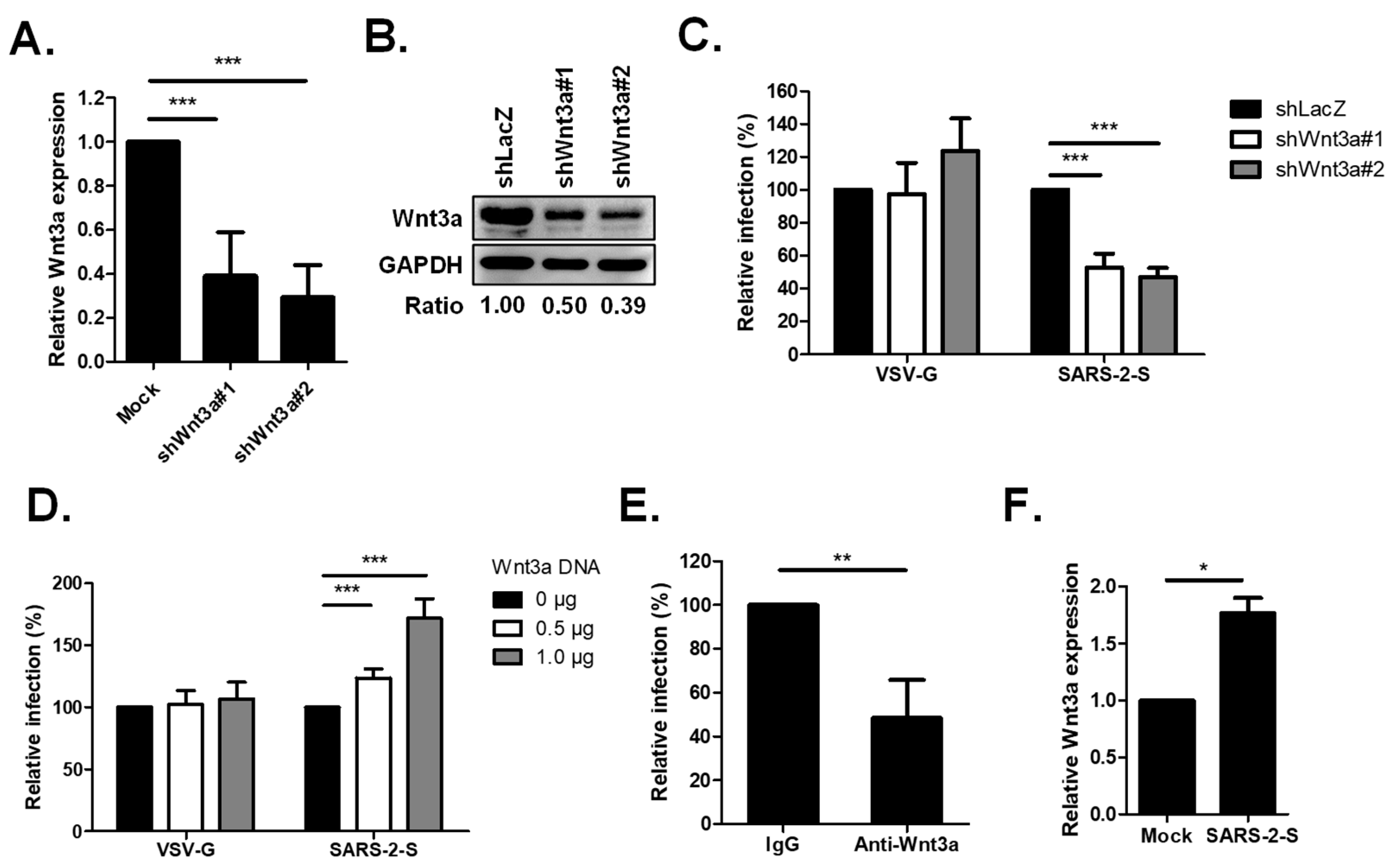
2.1. Wnt3a Is Involved in SARS-CoV-2 Entry
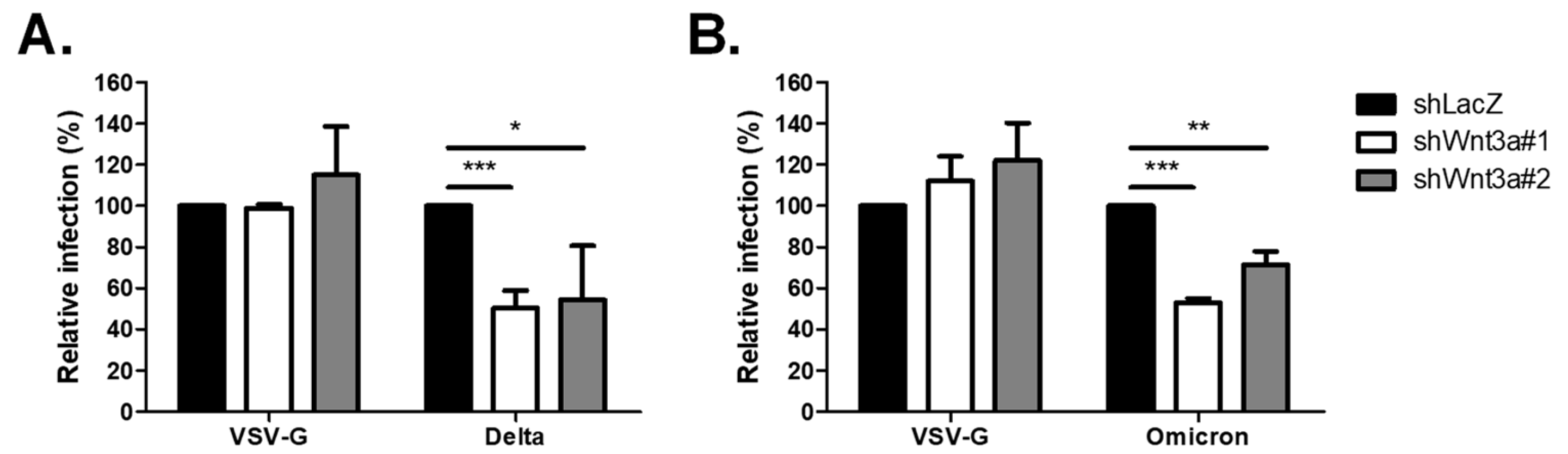
2.2. Loss of Wnt3a-Affected Infectivity of SARS-CoV-2 Variants
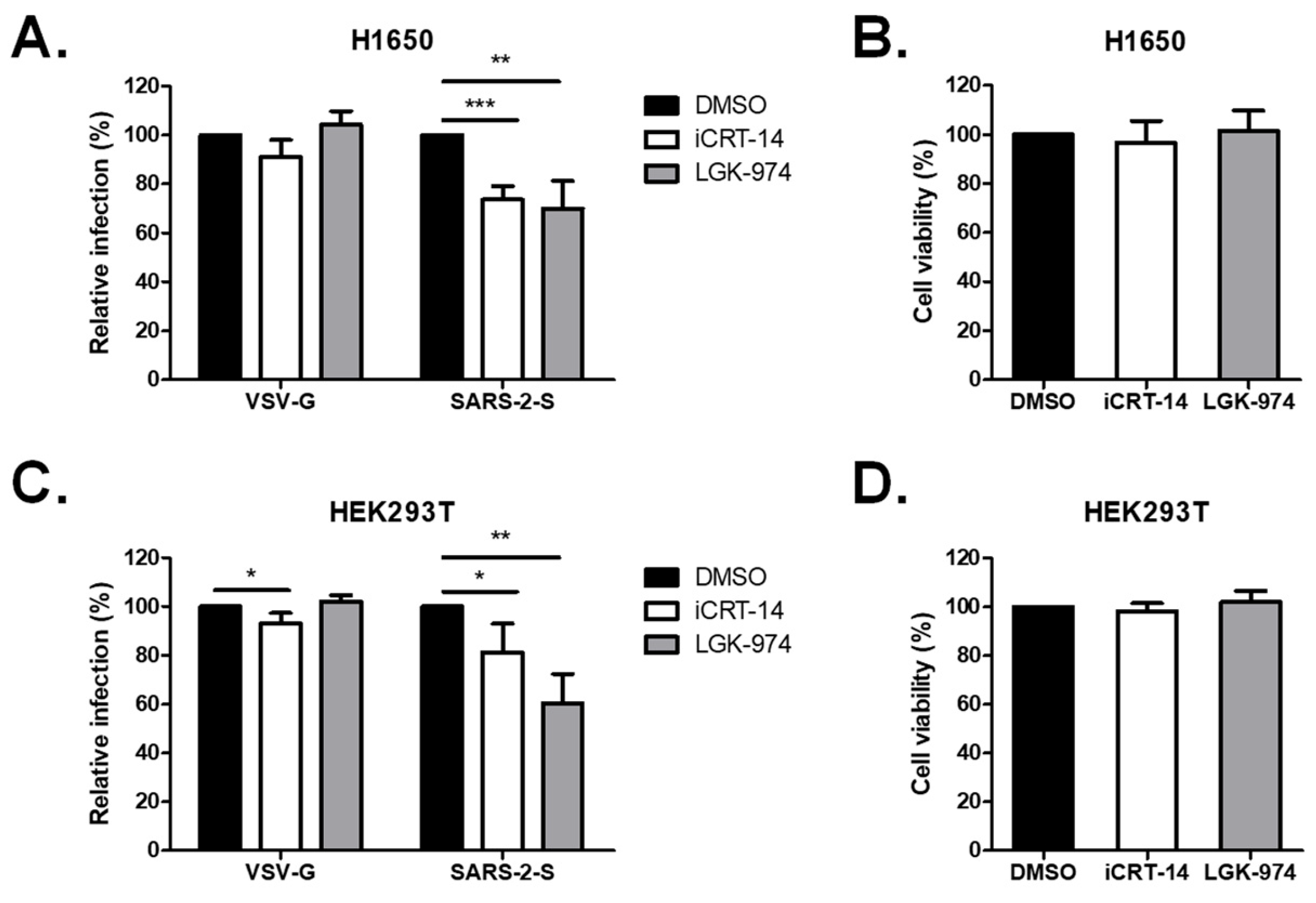
2.3. Wnt3a Inhibitors Attenuated SARS-CoV-2 Vpp Infection
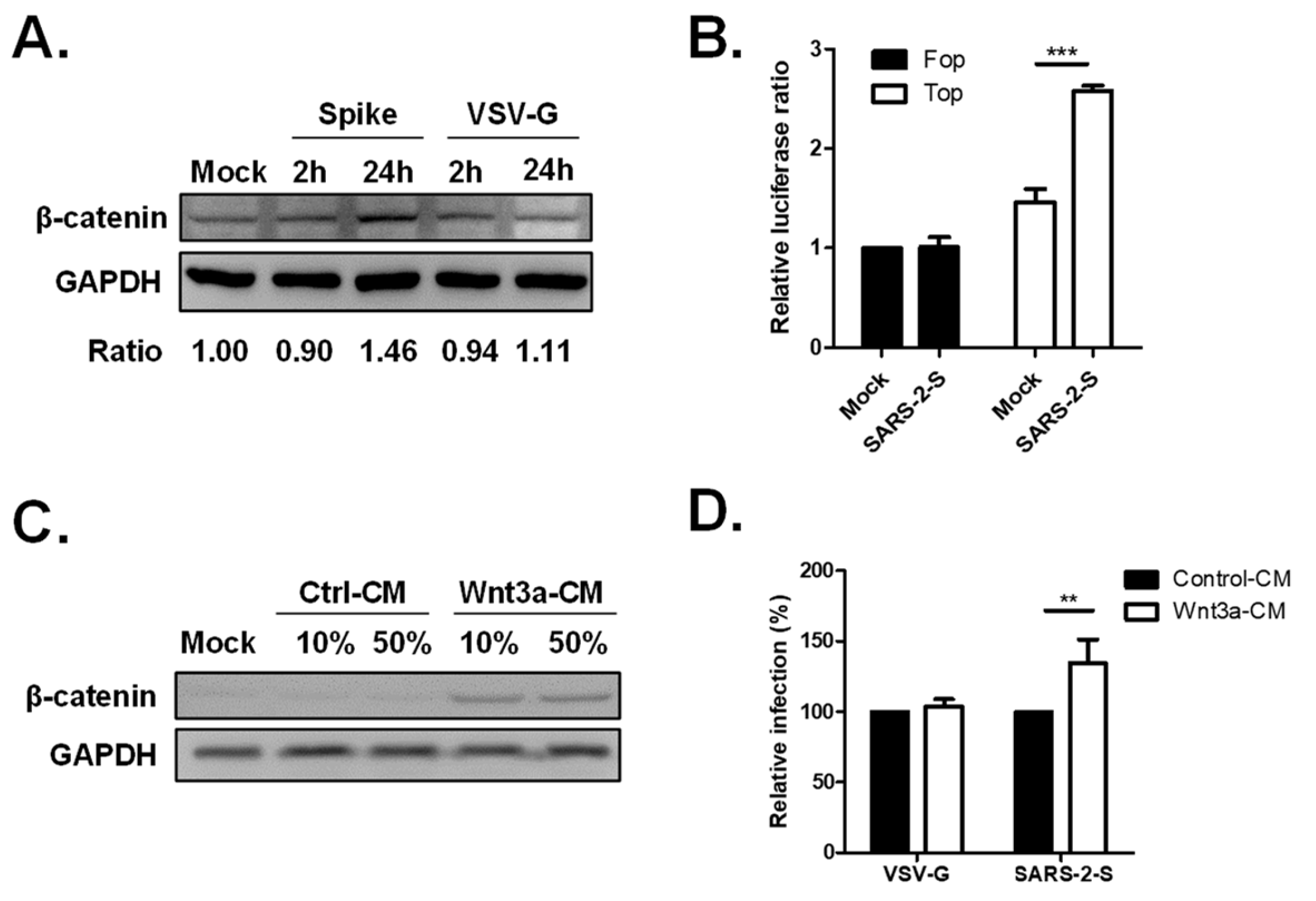
2.4. Activation of the Wnt/β-Catenin Signaling Pathway by SARS-CoV-2 Infection
2.5. The Wnt3a/β-Catenin Pathway Regulates ACE2 Transcription
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Plasmids and Viruses
4.3. Antibodies and Reagents
4.4. RT-qPCR
4.5. Western Blotting
4.6. Luciferase and MTS Assay
4.7. Antibody Competition Assay
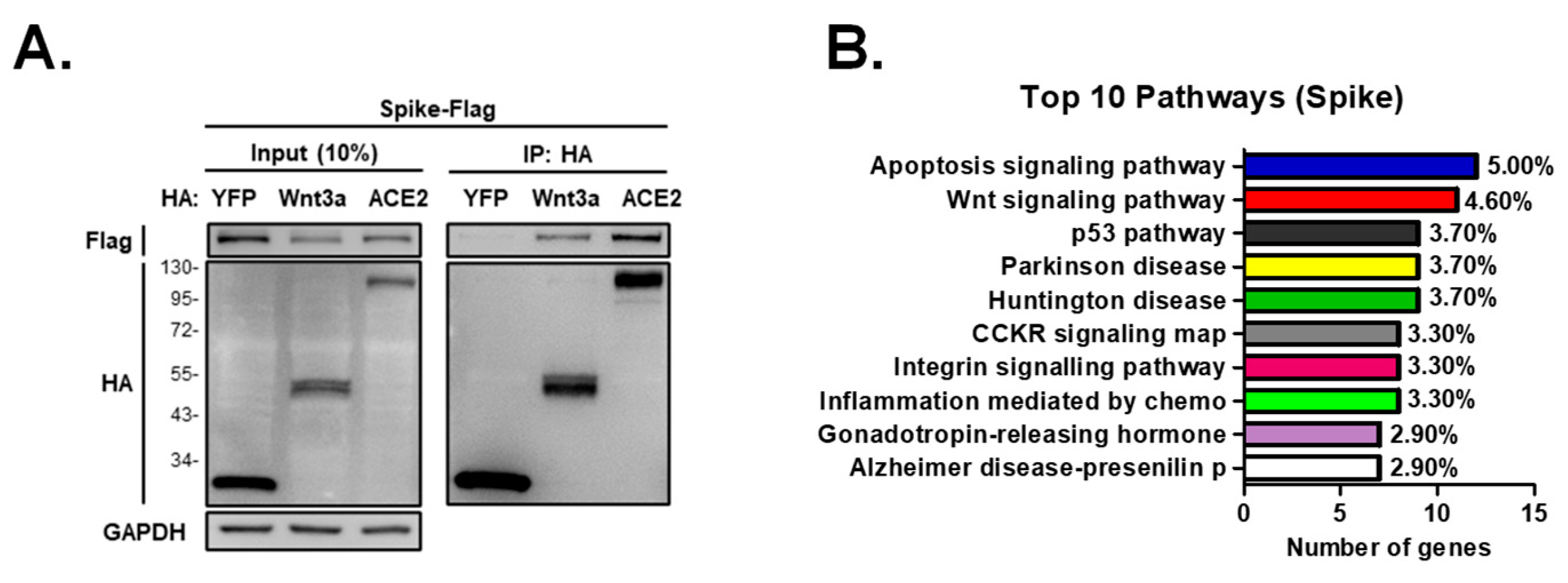
4.8. Immunoprecipitation Assay
4.9. Dual Luciferase Activity Assay
4.10. LC-MS/MS
4.11. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoffmann, M.; Kleine-Weber, H.; Pöhlmann, S. A Multibasic Cleavage Site in the Spike Protein of SARS-CoV-2 Is Essential for Infection of Human Lung Cells. Mol. Cell 2020, 78, 779–784.e5. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.-M.; Yang, W.-L.; Yang, F.-Y.; Zhang, L.; Huang, W.-J.; Hou, W.; Fan, C.-F.; Jin, R.-H.; Feng, Y.-M.; Wang, Y.-C.; et al. Cathepsin L plays a key role in SARS-CoV-2 infection in humans and humanized mice and is a promising target for new drug development. Signal Transduct. Target. Ther. 2021, 6, 134. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.; Chaudhary, J.K.; Jain, N.; Chaudhary, P.K.; Khanra, S.; Dhamija, P.; Sharma, A.; Kumar, A.; Handu, S. Role of Structural and Non-Structural Proteins and Therapeutic Targets of SARS-CoV-2 for COVID-19. Cells 2021, 10, 821. [Google Scholar] [CrossRef] [PubMed]
- Qi, F.; Qian, S.; Zhang, S.; Zhang, Z. Single cell RNA sequencing of 13 human tissues identify cell types and receptors of human coronaviruses. Biochem. Biophys. Res. Commun. 2020, 526, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Hikmet, F.; Méar, L.; Edvinsson, Å.; Micke, P.; Uhlén, M.; Lindskog, C. The protein expression profile of ACE2 in human tissues. Mol. Syst. Biol. 2020, 16, e9610. [Google Scholar] [CrossRef] [PubMed]
- Melano, I.; Cheng, W.-C.; Kuo, L.-L.; Liu, Y.-M.; Chou, Y.C.; Hung, M.-C.; Lai, M.M.C.; Sher, Y.-P.; Su, W.-C. A disintegrin and metalloproteinase domain 9 facilitates SARS-CoV-2 entry into cells with low ACE2 expression. Microbiol. Spectr. 2023, 11, e03854-22. [Google Scholar] [CrossRef] [PubMed]
- Ljungberg, J.K.; Kling, J.C.; Tran, T.T.; Blumenthal, A. Functions of the WNT Signaling Network in Shaping Host Responses to Infection. Front. Immunol. 2019, 10, 2521. [Google Scholar] [CrossRef]
- Pashirzad, M.; Fiuji, H.; Khazei, M.; Moradi-Binabaj, M.; Ryzhikov, M.; Shabani, M.; Avan, A.; Hassanian, S.M. Role of Wnt3a in the pathogenesis of cancer, current status and prospective. Mol. Biol. Rep. 2019, 46, 5609–5616. [Google Scholar] [CrossRef]
- Wang, C.; Hu, R.; Duan, L.; Hou, Q.; Yang, M.; Wang, T.; Liu, H.; Xiao, S.; Dang, R.; Wang, J.; et al. The canonical Wnt/β-catenin signaling pathway facilitates pseudorabies virus proliferation and enhances virus-induced autophagy. Vet. Microbiol. 2022, 272, 109502. [Google Scholar] [CrossRef]
- Zhu, L.; Jones, C. The canonical Wnt/β-catenin signaling pathway stimulates herpes simplex virus 1 productive infection. Virus Res. 2018, 256, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Harmon, B.; Bird, S.W.; Schudel, B.R.; Hatch, A.V.; Rasley, A.; Negrete, O.A. A Genome-Wide RNA Interference Screen Identifies a Role for Wnt/β-Catenin Signaling during Rift Valley Fever Virus Infection. J. Virol. 2016, 90, 7084–7097. [Google Scholar] [CrossRef] [PubMed]
- Tran, B.M.; Flanagan, D.J.; Ebert, G.; Warner, N.; Tran, H.; Fifis, T.; Kastrappis, G.; Christophi, C.; Pellegrini, M.; Torresi, J.; et al. The Hepatitis B Virus Pre-Core Protein p22 Activates Wnt Signaling. Cancers 2020, 12, 1435. [Google Scholar] [CrossRef] [PubMed]
- Qiao, D.; He, Q.; Cheng, X.; Yao, Y.; Nair, V.; Shao, H.; Qin, A.; Qian, K. Regulation of Avian Leukosis Virus Subgroup J Replication by Wnt/β-Catenin Signaling Pathway. Viruses 2021, 13, 1968. [Google Scholar] [CrossRef]
- More, S.; Yang, X.; Zhu, Z.; Bamunuarachchi, G.; Guo, Y.; Huang, C.; Bailey, K.; Metcalf, J.P.; Liu, L. Regulation of influenza virus replication by Wnt/β-catenin signaling. PLoS ONE 2018, 13, e0191010. [Google Scholar] [CrossRef]
- Maluquer de Motes, C.; Smith, G.L. Vaccinia virus protein A49 activates Wnt signalling by targetting the E3 ligase β-TrCP. J. Gen. Virol. 2017, 98, 3086–3092. [Google Scholar] [CrossRef]
- Chatterjee, S.; Keshry, S.S.; Ghosh, S.; Ray, A.; Chattopadhyay, S. Versatile β-Catenin Is Crucial for SARS-CoV-2 Infection. Microbiol. Spectr. 2022, 10, e01670-22. [Google Scholar] [CrossRef]
- Koval, A.; Xu, J.; Williams, N.; Schmolke, M.; Krause, K.-H.; Katanaev, V.L. Wnt-Independent SARS-CoV-2 Infection in Pulmonary Epithelial Cells. Microbiol. Spectr. 2023, 11, e04827-22. [Google Scholar] [CrossRef]
- Bohan, D.; Van Ert, H.; Ruggio, N.; Rogers, K.J.; Badreddine, M.; Aguilar Briseño, J.A.; Elliff, J.M.; Rojas Chavez, R.A.; Gao, B.; Stokowy, T.; et al. Phosphatidylserine receptors enhance SARS-CoV-2 infection. PLoS Pathog. 2021, 17, e1009743. [Google Scholar] [CrossRef]
- Pouwels, K.B.; Pritchard, E.; Matthews, P.C.; Stoesser, N.; Eyre, D.W.; Vihta, K.-D.; House, T.; Hay, J.; Bell, J.I.; Newton, J.N.; et al. Effect of Delta variant on viral burden and vaccine effectiveness against new SARS-CoV-2 infections in the UK. Nat. Med. 2021, 27, 2127–2135. [Google Scholar] [CrossRef]
- Andrews, N.; Stowe, J.; Kirsebom, F.; Toffa, S.; Rickeard, T.; Gallagher, E.; Gower, C.; Kall, M.; Groves, N.; O’Connell, A.-M.; et al. Effectiveness of COVID-19 vaccines against the Omicron (B.1.1.529) variant of concern. medRxiv 2021. medRxiv:2021.12.14.21267615. [Google Scholar]
- Collie, S.; Champion, J.; Moultrie, H.; Bekker, L.-G.; Gray, G. Effectiveness of BNT162b2 Vaccine against Omicron Variant in South Africa. N. Engl. J. Med. 2021, 386, 494–496. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Beltran, W.F.; Lam, E.C.; St Denis, K.; Nitido, A.D.; Garcia, Z.H.; Hauser, B.M.; Feldman, J.; Pavlovic, M.N.; Gregory, D.J.; Poznansky, M.C.; et al. Multiple SARS-CoV-2 variants escape neutralization by vaccine-induced humoral immunity. Cell 2021, 184, 2372–2383.e9. [Google Scholar] [CrossRef] [PubMed]
- Farré, D.; Roset, R.; Huerta, M.; Adsuara, J.E.; Roselló, L.; Albà, M.M.; Messeguer, X. Identification of patterns in biological sequences at the ALGGEN server: PROMO and MALGEN. Nucleic Acids Res. 2003, 31, 3651–3653. [Google Scholar] [CrossRef] [PubMed]
- Messeguer, X.; Escudero, R.; Farré, D.; Núñez, O.; Martínez, J.; Albà, M.M. PROMO: Detection of known transcription regulatory elements using species-tailored searches. Bioinformatics 2002, 18, 333–334. [Google Scholar] [CrossRef] [PubMed]
- Robinson, P.C.; Liew, D.F.L.; Tanner, H.L.; Grainger, J.R.; Dwek, R.A.; Reisler, R.B.; Steinman, L.; Feldmann, M.; Ho, L.-P.; Hussell, T.; et al. COVID-19 therapeutics: Challenges and directions for the future. Proc. Natl. Acad. Sci. USA 2022, 119, e2119893119. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Sharma, S.; Kumar, R.; Tripathi, B.N.; Barua, S.; Ly, H.; Rouse, B.T. Host-Directed Antiviral Therapy. Clin. Microbiol. Rev. 2020, 33, 3. [Google Scholar] [CrossRef]
- Neiheisel, A.; Kaur, M.; Ma, N.; Havard, P.; Shenoy, A.K. Wnt pathway modulators in cancer therapeutics: An update on completed and ongoing clinical trials. Int. J. Cancer 2022, 150, 727–740. [Google Scholar] [CrossRef]
- Rodon, J.; Argilés, G.; Connolly, R.M.; Vaishampayan, U.; de Jonge, M.; Garralda, E.; Giannakis, M.; Smith, D.C.; Dobson, J.R.; McLaughlin, M.E.; et al. Phase 1 study of single-agent WNT974, a first-in-class Porcupine inhibitor, in patients with advanced solid tumours. Br. J. Cancer 2021, 125, 28–37. [Google Scholar] [CrossRef]
- Gheware, A.; Ray, A.; Rana, D.; Bajpai, P.; Nambirajan, A.; Arulselvi, S.; Mathur, P.; Trikha, A.; Arava, S.; Das, P.; et al. ACE2 protein expression in lung tissues of severe COVID-19 infection. Sci. Rep. 2022, 12, 4058. [Google Scholar] [CrossRef]
- Mesta, F.; Coll, A.M.; Ramírez M, Á.; Delgado-Roche, L. Predictors of Mortality in Hospitalized COVID-19 Patients: A Mexican Population-Based Cohort Study. Biomedicine 2021, 11, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.Y.; Park, H.H.; Kim, H.; Kim, H.N.; Kim, I.; Jeon, S.; Kim, W.; Bae, J.-S.; Lee, W. Wnt5a and Wnt11 as acute respiratory distress syndrome biomarkers for severe acute respiratory syndrome coronavirus 2 patients. Eur. Respir. J. 2020, 56, 2001531. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [PubMed]
- Melano, I.; Kuo, L.L.; Lo, Y.C.; Sung, P.W.; Tien, N.; Su, W.C. Effects of Basic Amino Acids and Their Derivatives on SARS-CoV-2 and Influenza-A Virus Infection. Viruses 2021, 13, 1301. [Google Scholar] [CrossRef]
- Chen, H.-J.; Hsu, L.-S.; Shia, Y.-T.; Lin, M.-W.; Lin, C.-M. The β-catenin/TCF complex as a novel target of resveratrol in the Wnt/β-catenin signaling pathway. Biochem. Pharmacol. 2012, 84, 1143–1153. [Google Scholar] [CrossRef]
- Hsu, Y.-H.; Chao, C.-N.; Huang, H.-Y.; Zhao, P.-W.; Hsu, P.-H.; Shen, C.-H.; Chen, S.-Y.; Fang, C.-Y. Histone deacetylase III interactions with BK polyomavirus large tumor antigen may affect protein stability. Virol. J. 2023, 20, 155. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melano, I.; Chen, H.-J.; Ngwira, L.; Hsu, P.-H.; Kuo, L.-L.; Noriega, L.; Su, W.-C. Wnt3a Facilitates SARS-CoV-2 Pseudovirus Entry into Cells. Int. J. Mol. Sci. 2024, 25, 217. https://doi.org/10.3390/ijms25010217
Melano I, Chen H-J, Ngwira L, Hsu P-H, Kuo L-L, Noriega L, Su W-C. Wnt3a Facilitates SARS-CoV-2 Pseudovirus Entry into Cells. International Journal of Molecular Sciences. 2024; 25(1):217. https://doi.org/10.3390/ijms25010217
Chicago/Turabian StyleMelano, Ivonne, Hui-Jye Chen, Loveness Ngwira, Pang-Hung Hsu, Li-Lan Kuo, Lloyd Noriega, and Wen-Chi Su. 2024. "Wnt3a Facilitates SARS-CoV-2 Pseudovirus Entry into Cells" International Journal of Molecular Sciences 25, no. 1: 217. https://doi.org/10.3390/ijms25010217
APA StyleMelano, I., Chen, H.-J., Ngwira, L., Hsu, P.-H., Kuo, L.-L., Noriega, L., & Su, W.-C. (2024). Wnt3a Facilitates SARS-CoV-2 Pseudovirus Entry into Cells. International Journal of Molecular Sciences, 25(1), 217. https://doi.org/10.3390/ijms25010217