
Phenotypic Variation in Flower Color and Morphology in the Gerbera (Gerbera hybrida) F1 Hybrid Population and Their Association with EST-SSR Markers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
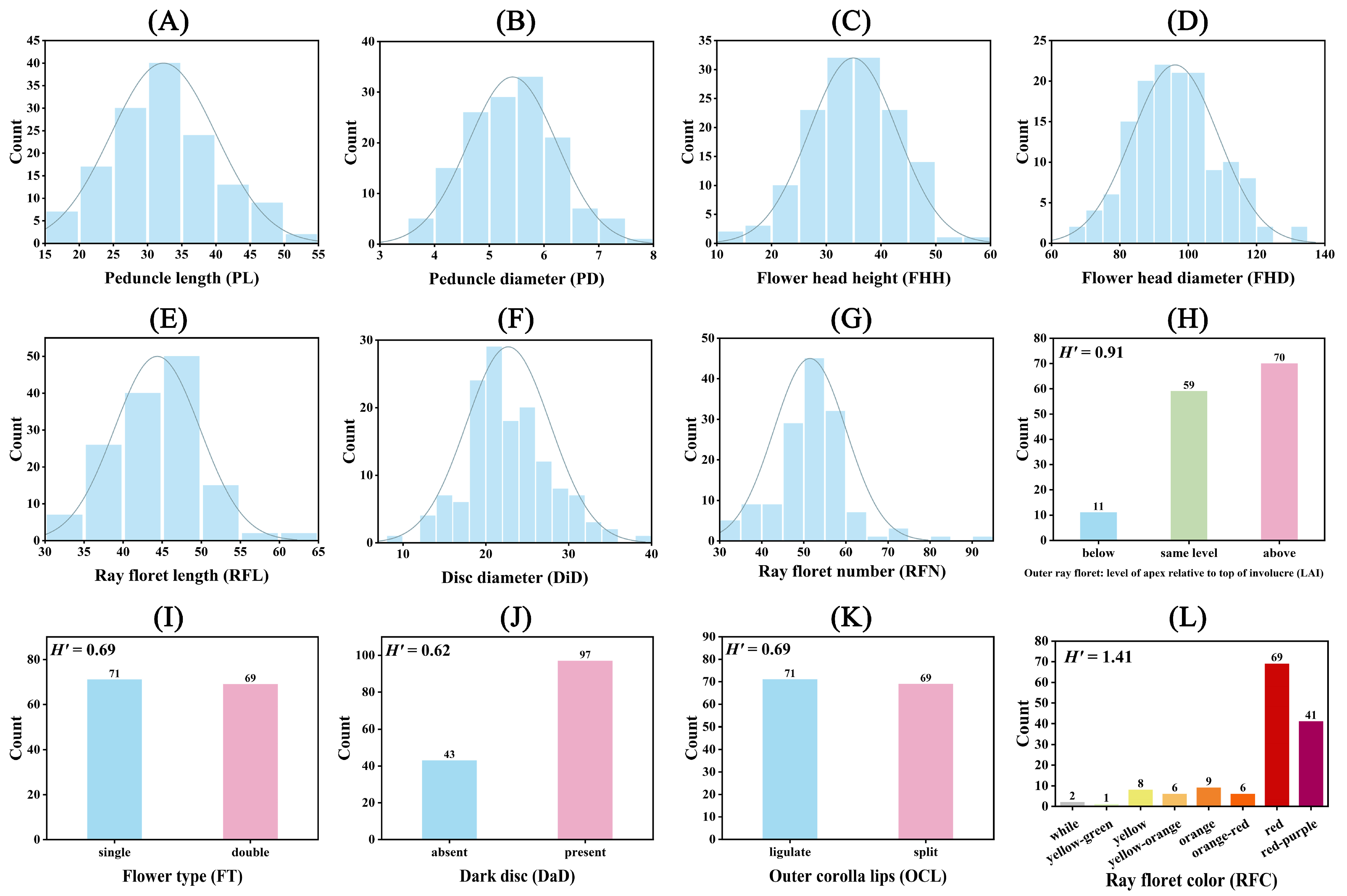
2.1. Genetic Analysis of Morphological Traits and Grouping Colors
2.2. Clustering and Principal Component Analysis Based on 12 Phenotypic Traits
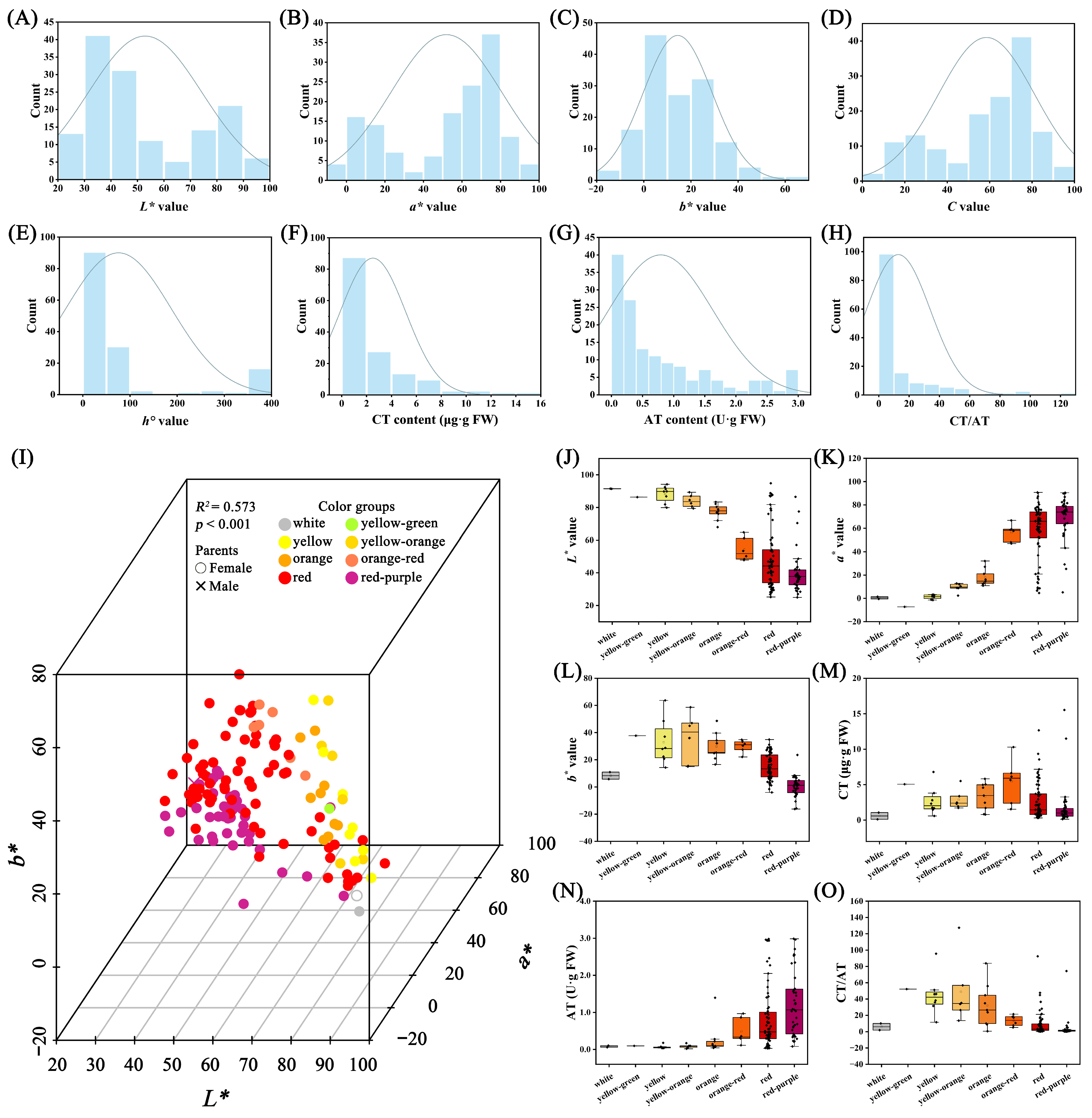
2.3. Colorimeter Determination and Pigment Diversity Analysis
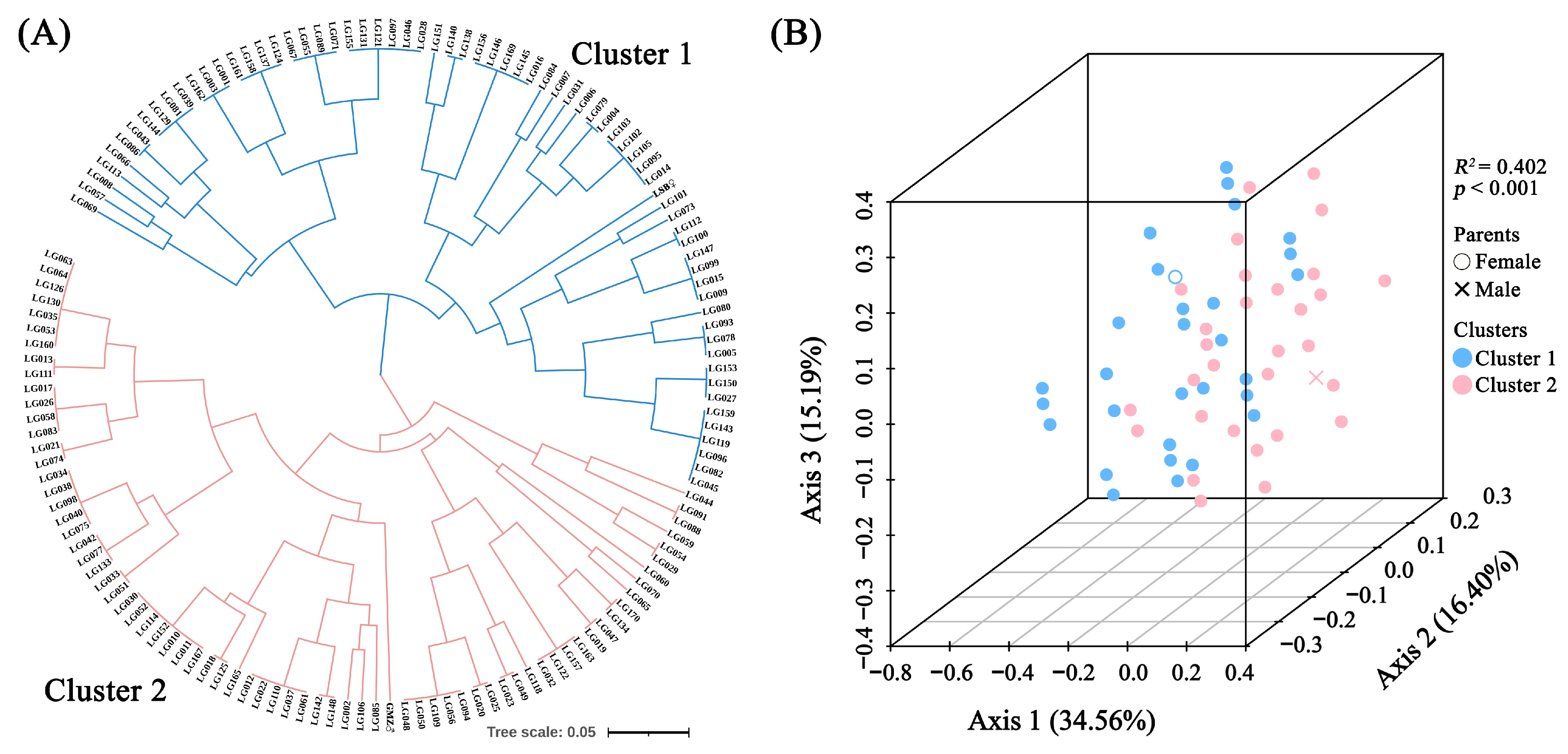
2.4. Clustering and Principal Coordinate Analysis Based on EST-SSR Markers
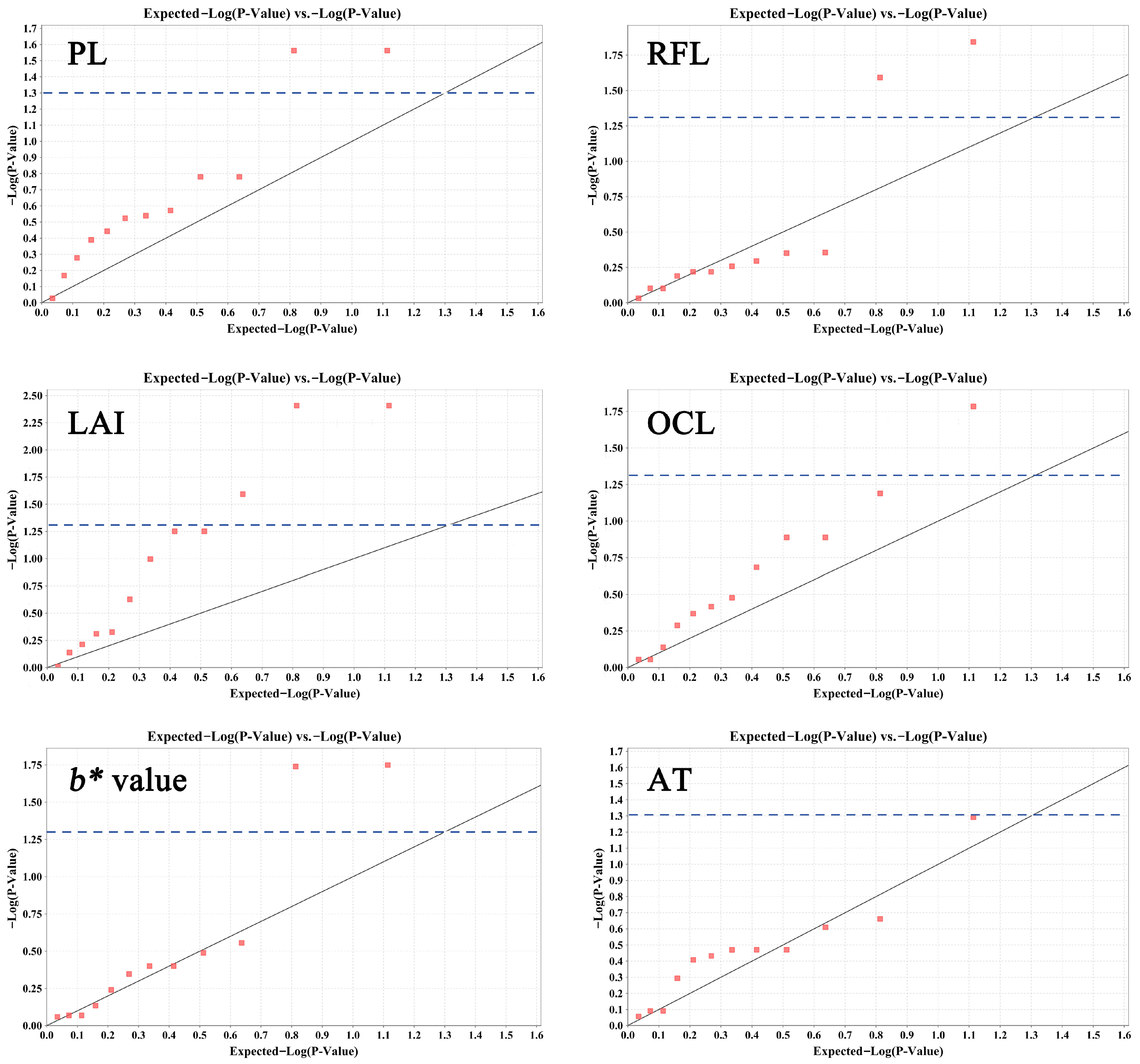
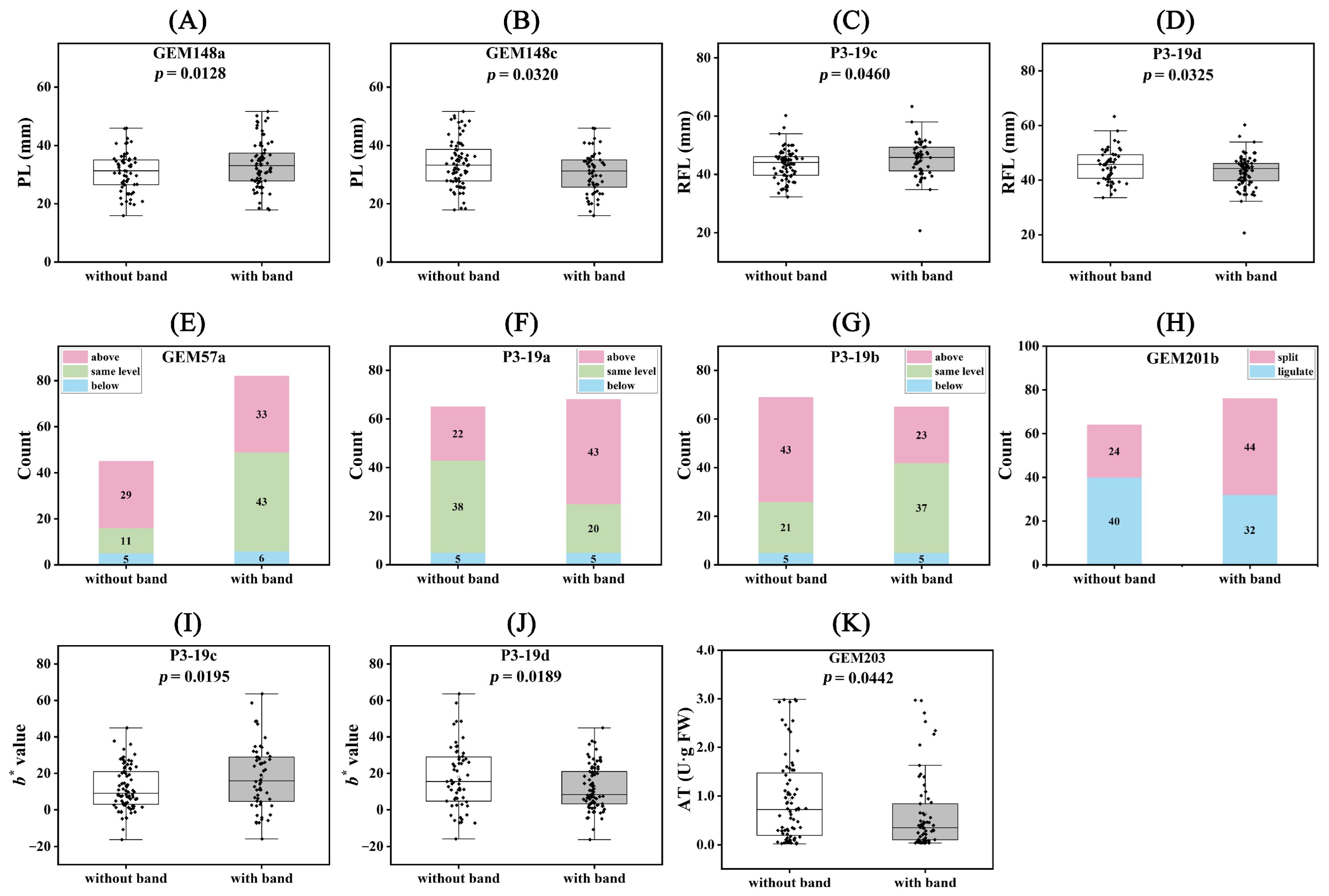
2.5. Association Analysis between Traits and Molecular Markers
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Observation of Morphological Traits
4.3. Determination of Flower Color Traits
4.4. Analysis of Total Carotenoid Content and Anthocyanin Contents
4.5. DNA Extraction and SSR Genotyping
4.6. Statistic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carrodeguas-Gonzalez, A.; Zúñiga-Orozco, A. Aplicación de herramientas moleculares para el mejoramiento genético de pasturas. Pastos Y Forrajes 2021, 44, e121. [Google Scholar]
- Song, X.; Deng, Z. Powdery Mildew Resistance in Gerbera: Mode of Inheritance, Quantitative Trait Locus Identification, and Resistance Responses. J. Am. Soc. Hortic. Sci. 2013, 138, 470–478. [Google Scholar] [CrossRef]
- Teeri, T.H.; Elomaa, P.; Kotilainen, M.; Albert, V.A. Mining plant diversity: Gerbera as a model system for plant developmental and biosynthetic research. Bioessays 2006, 28, 756–767. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Esselink, G.D.; Visser, R.G.F.; van Tuyl, J.M.; Arens, P. Transcriptome Analysis of Gerbera hybrida Including in silico Confirmation of Defense Genes Found. Front. Plant Sci. 2016, 7, 247. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Jin, X.; Yao, W.; Kong, L.; Huang, G.; Tao, Y.; Li, L.; Wang, X.; Wang, Y. A Mini Zinc-Finger Protein (MIF) from Gerbera hybrida Activates the GASA Protein Family Gene, GEG, to Inhibit Ray Petal Elongation. Front. Plant Sci. 2017, 8, 1649. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Han, M.; Jian, L.; Chen, Y.; Sun, S.; Wang, X.; Wang, Y. An ETHYLENE INSENSITIVE3-LIKE1 Protein Directly Targets the GEG Promoter and Mediates Ethylene-Induced Ray Petal Elongation in Gerbera hybrida. Front. Plant Sci. 2020, 10, 1737. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Han, M.; Yao, W.; Wang, Y. Transcriptome analysis reveals the regulation of brassinosteroids on petal growth in Gerbera hybrida. PeerJ 2017, 5, e3382. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Li, L.; Huang, Y.; Wang, Y.; Zhang, W.; Zheng, R.; Zhong, C.; Wang, X. GhWIP2, a WIP zinc finger protein, suppresses cell expansion in Gerbera hybrida by mediating crosstalk between gibberellin, abscisic acid, and auxin. New Phytol. 2018, 219, 728–742. [Google Scholar] [CrossRef]
- Lin, X.; Huang, S.; Huang, G.; Chen, Y.; Wang, X.; Wang, Y. 14-3-3 Proteins Are Involved in BR-Induced Ray Petal Elongation in Gerbera hybrida. Front. Plant Sci. 2021, 12, 718091. [Google Scholar] [CrossRef]
- Li, L.; Zhang, W.; Zhang, L.; Li, N.; Peng, J.; Wang, Y.; Zhong, C.; Yang, Y.; Sun, S.; Liang, S.; et al. Transcriptomic insights into antagonistic effects of gibberellin and abscisic acid on petal growth in Gerbera hybrida. Front. Plant Sci. 2015, 6, 168. [Google Scholar] [CrossRef]
- Kloos, W.E.; George, C.G.; Sorge, L.K. Inheritance of the flower types of Gerbera hybrida. J. Am. Soc. Hortic. Sci. 2004, 129, 802–810. [Google Scholar] [CrossRef]
- Kotilainen, M.; Helariutta, Y.; Mehto, M.; Pöllänen, E.; Albert, V.A.; Elomaa, P.; Teeri, T.H. GEG Participates in the Regulation of Cell and Organ Shape during Corolla and Carpel Development in Gerbera hybrida. Plant Cell 1999, 11, 1093–1104. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R. Studies on Genetic Variability in Gerbera (Gerbera jamesonii Bolus Ex. Hooker F.). J. Hortic. Sci. 2013, 8, 111–113. [Google Scholar] [CrossRef]
- Rymbai, H.; Verma, V.K.; Mawleiñ, J.; Hazarika, S. Analysis of genetic divergence, principal component, correlation and path coefficient for quantitative traits of Gerbera (Gerbera jamesonii) in the north eastern region, India. Plant Genet. Resour. Charact. Util. 2023, 21, 229–236. [Google Scholar] [CrossRef]
- Byrne, D. Molecular Marker Use in Tree Fruit and Woody Ornamental Plant Breeding. Hortscience 2006, 41, 985. [Google Scholar] [CrossRef]
- Francia, E.; Tacconi, G.; Crosatti, C.; Barabaschi, D.; Bulgarelli, D.; Dall’Aglio, E.; Vale, G. Marker assisted selection in crop plants. Plant Cell Tissue Organ Cult. 2005, 82, 317–342. [Google Scholar] [CrossRef]
- Prajapati, P.; Singh, A.; Patel, N.L.; Singh, D.; Srivastav, V. Evaluation of genetic diversity in different genotypes of Gerbera jamesonii Bolus using random amplified polymorphic DNA (RAPD) markers. Afr. J. Biotechnol. 2014, 13, 1117–1122. [Google Scholar] [CrossRef]
- Saidi, A.; Hajkazemian, M. Comparative assessment of ISSR, DAMD and RAPD markers for evaluation of genetic diversity of gerbera (Gerbera jamesonii Bolus ex Hooker f.) cultivars. Acta Agric. Slov. 2023, 119, 1–8. [Google Scholar] [CrossRef]
- Gong, L.; Deng, Z. EST-SSR markers for gerbera (Gerbera hybrida). Mol. Breed. 2010, 26, 125–132. [Google Scholar] [CrossRef]
- Gong, L.; Deng, Z. Selection and application of SSR markers for variety discrimination, genetic similarity and relation analysis in gerbera (Gerbera hybrida). Sci. Hortic. 2012, 138, 120–127. [Google Scholar] [CrossRef]
- Zhou, Y.; Yin, M.; Abbas, F.; Sun, Y.; Gao, T.; Yan, F.; Li, X.; Yu, Y.; Yue, Y.; Yu, R.; et al. Classification and Association Analysis of Gerbera (Gerbera hybrida) Flower Color Traits. Front. Plant Sci. 2022, 12, 779288. [Google Scholar] [CrossRef] [PubMed]
- Schlotterer, C. The evolution of molecular markers—Just a matter of fashion? Nat. Rev. Genet. 2004, 5, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; van Silfhout, A.; Shahin, A.; Egberts, R.; Beers, M.; van der Velde, A.; van Houten, A.; van Tuyl, J.M.; Visser, R.G.F.; Arens, P. Erratum to: Genetic mapping and QTL analysis of Botrytis resistance in Gerbera hybrida. Mol. Breed. 2017, 37, 1–2. [Google Scholar] [CrossRef]
- Bhattarai, K.; Sharma, S.; Verma, S.; Peres, N.A.; Xiao, S.; Clark, D.G.; Deng, Z. Construction of a genome-wide genetic linkage map and identification of quantitative trait loci for powdery mildew resistance in Gerbera daisy. Front. Plant Sci. 2023, 13, 1072717. [Google Scholar] [CrossRef] [PubMed]
- Hârţa, M.; Clapa, D.; Cornea-Cipcigan, M.; Borsai, O.; Pop, R.; Cordea, M.I. Multivariate Assessment of Genetic Relationships between Two Streptocarpus Cultivars and Their F1 Progenies Using Morphological Characteristics and SCoT Molecular Markers. Horticulturae 2023, 9, 497. [Google Scholar] [CrossRef]
- Rai, R.; Nguyen, V.Y.; Kim, J.C. Variability analysis and evaluation for major cut flower traits of F1 hybrids in Lilium brownii var. colchesteri. J. Multidiscip. Sci. 2022, 4, 35–41. [Google Scholar] [CrossRef]
- Yang, Y.; Wen, C.; Ma, N.; Zhao, L. Heterosis and genetic analysis of branching in cut-flower chrysanthemums. Euphytica 2015, 205, 915–925. [Google Scholar] [CrossRef]
- Song, X.; Gao, K.; Fan, G.; Zhao, X.; Liu, Z.; Dai, S. Quantitative Classification of the Morphological Traits of Ray Florets in Large-flowered Chrysanthemum. Hortscience 2018, 53, 1258–1265. [Google Scholar] [CrossRef]
- Song, X.; Xu, Y.; Gao, K.; Fan, G.; Zhang, F.; Deng, C.; Dai, S.; Huang, H.; Xin, H.; Li, Y. High-density genetic map construction and identification of loci controlling flower-type traits in Chrysanthemum (Chrysanthemum × morifolium Ramat.). Hortic. Res. 2020, 7, 108. [Google Scholar] [CrossRef]
- Wu, X.; Zhao, X.; Gao, K.; Tian, Y.; Zhang, M.; Anderson, N.O.; Dai, S. Heterosis and Mixed Genetic Analysis of Flowering Traits in Cross Breeding of Day-Neutral Chrysanthemum (Asteraceae). Agronomy 2023, 13, 2107. [Google Scholar] [CrossRef]
- Zhou, Y.; Abbas, F.; He, J.; Yan, F.; Wang, Q.; Yu, Y.; Yu, R.; Fan, Y. Floral volatile chemical diversity in Hedychium F1 hybrid population. Ind. Crop. Prod. 2022, 184, 115032. [Google Scholar] [CrossRef]
- Wei, X.; Zhou, Y.; Abbas, F.; Yan, F.; Zou, X.; Yu, Y.; Gao, T.; He, J.; Wang, Q.; Yu, R.; et al. Distant heteroploid hybridization improved Hedychium floral scent, floral color and morphologcal traits. Ind. Crop. Prod. 2023, 194, 116357. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids: Harnessing plant biomass for biofuels and biomaterials. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Ng, J.; Freitas, L.B.; Smith, S.D. Stepwise evolution of floral pigmentation predicted by biochemical pathway structure. Evolution 2018, 72, 2792–2802. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, Y.; Long, T.; Wang, S.; Yang, J. Regulation Mechanism of Plant Pigments Biosynthesis: Anthocyanins, Carotenoids, and Betalains. Metabolites 2022, 12, 871. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Li, Y.; Wang, J.; Qu, J.; Chen, Y.; Chen, X.; Huang, H.; Dai, S. Flower color classification and correlation between color space values with pigments in potted multiflora chrysanthemum. Sci. Hortic. 2021, 283, 110082. [Google Scholar] [CrossRef]
- Yamamoto, T. DNA Markers and Molecular Breeding in Pear and Other Rosaceae Fruit Trees. Hortic. J. 2021, 90, 1–13. [Google Scholar] [CrossRef]
- Ibrahim, A.K.; Zhang, L.; Niyitanga, S.; Afzal, M.Z.; Xu, Y.; Zhang, L.; Zhang, L.; Qi, J. Principles and approaches of association mapping in plant breeding. Trop. Plant Biol. 2020, 13, 212–224. [Google Scholar] [CrossRef]
- Zhou, Y.; Ye, Y.; Zhu, G.; Xu, Y.; Tan, J.; Liu, J. Diversity, classification, and EST-SSR-based association analysis of caladium ornamental traits. Physiol. Plant. 2023, 175, e13841. [Google Scholar] [CrossRef]
- Shi, Z.; Zhao, W.; Li, Z.; Kang, D.; Ai, P.; Ding, H.; Wang, Z. Development and validation of SSR markers related to flower color based on full-length transcriptome sequencing in Chrysanthemum. Sci. Rep. 2022, 12, 22310. [Google Scholar] [CrossRef]
- Li, S.; Ji, F.; Hou, F.; Cui, H.; Shi, Q.; Xing, G.; Weng, Y.; Kang, X. Characterization of Hemerocallis citrina Transcriptome and Development of EST-SSR Markers for Evaluation of Genetic Diversity and Population Structure of Hemerocallis Collection. Front. Plant Sci. 2020, 11, 686. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Cheng, F. Association mapping for yield traits in Paeonia rockii based on SSR markers within transcription factors of comparative transcriptome. BMC Plant Biol. 2020, 20, 245. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; He, R.; Zheng, J.; Hu, Z.; Wu, J.; Leng, P. Development of EST-SSR markers and association mapping with floral traits in Syringa oblata. BMC Plant Biol. 2020, 20, 436. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Warburton, M.; Crouch, J. Association Mapping for Enhancing Maize (Zea mays L.) Genetic Improvement. Crop Sci. 2011, 51, 433–449. [Google Scholar] [CrossRef]
- Sun, X.; Du, Z.; Ren, J.; Amombo, E.; Hu, T.; Fu, J. Association of SSR markers with functional traits from heat stress in diverse tall fescue accessions. BMC Plant Biol. 2015, 15, 116. [Google Scholar] [CrossRef]
- Gonnet, J. Colour effects of co-pigmentation of anthocyanins revisited—1. A colorimetric definition using the CIELAB scale. Food Chem. 1998, 63, 409–415. [Google Scholar] [CrossRef]
- Zhou, Y.; Wei, X.; Abbas, F.; Yu, Y.; Yu, R.; Fan, Y. Genome-wide identification of simple sequence repeats and assessment of genetic diversity in Hedychium. J. Appl. Res. Med. Aromat. Plants 2021, 24, 100312. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 10 September 2023).
- Wei, T.; Simko, V. R Package ‘Corrplot’: Visualization of a Correlation Matrix (Version 0.92). 2021. Available online: https://github.com/taiyun/corrplot (accessed on 10 September 2023).
- Peakall, R.; Smouse, P.E. genalex 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.B.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology Package. R Package Version 2.6-4. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 10 September 2023).
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Zou, X.; Yan, F.; He, J.; Zeng, S.; Yu, Y.; Tang, X.; Liang, X.; Cai, X.; Yu, R.; et al. Phenotypic Variation in Flower Color and Morphology in the Gerbera (Gerbera hybrida) F1 Hybrid Population and Their Association with EST-SSR Markers. Int. J. Mol. Sci. 2024, 25, 203. https://doi.org/10.3390/ijms25010203
Zhou Y, Zou X, Yan F, He J, Zeng S, Yu Y, Tang X, Liang X, Cai X, Yu R, et al. Phenotypic Variation in Flower Color and Morphology in the Gerbera (Gerbera hybrida) F1 Hybrid Population and Their Association with EST-SSR Markers. International Journal of Molecular Sciences. 2024; 25(1):203. https://doi.org/10.3390/ijms25010203
Chicago/Turabian StyleZhou, Yiwei, Xinru Zou, Fulong Yan, Jingjuan He, Sixian Zeng, Yunyi Yu, Xiaoshuang Tang, Xuanguo Liang, Xiuping Cai, Rangcai Yu, and et al. 2024. "Phenotypic Variation in Flower Color and Morphology in the Gerbera (Gerbera hybrida) F1 Hybrid Population and Their Association with EST-SSR Markers" International Journal of Molecular Sciences 25, no. 1: 203. https://doi.org/10.3390/ijms25010203
APA StyleZhou, Y., Zou, X., Yan, F., He, J., Zeng, S., Yu, Y., Tang, X., Liang, X., Cai, X., Yu, R., & Fan, Y. (2024). Phenotypic Variation in Flower Color and Morphology in the Gerbera (Gerbera hybrida) F1 Hybrid Population and Their Association with EST-SSR Markers. International Journal of Molecular Sciences, 25(1), 203. https://doi.org/10.3390/ijms25010203