
Vitamin C Regulates the Profibrotic Activity of Fibroblasts in In Vitro Replica Settings of Myocardial Infarction
 , , ,
, , ,
Abstract
1. Introduction
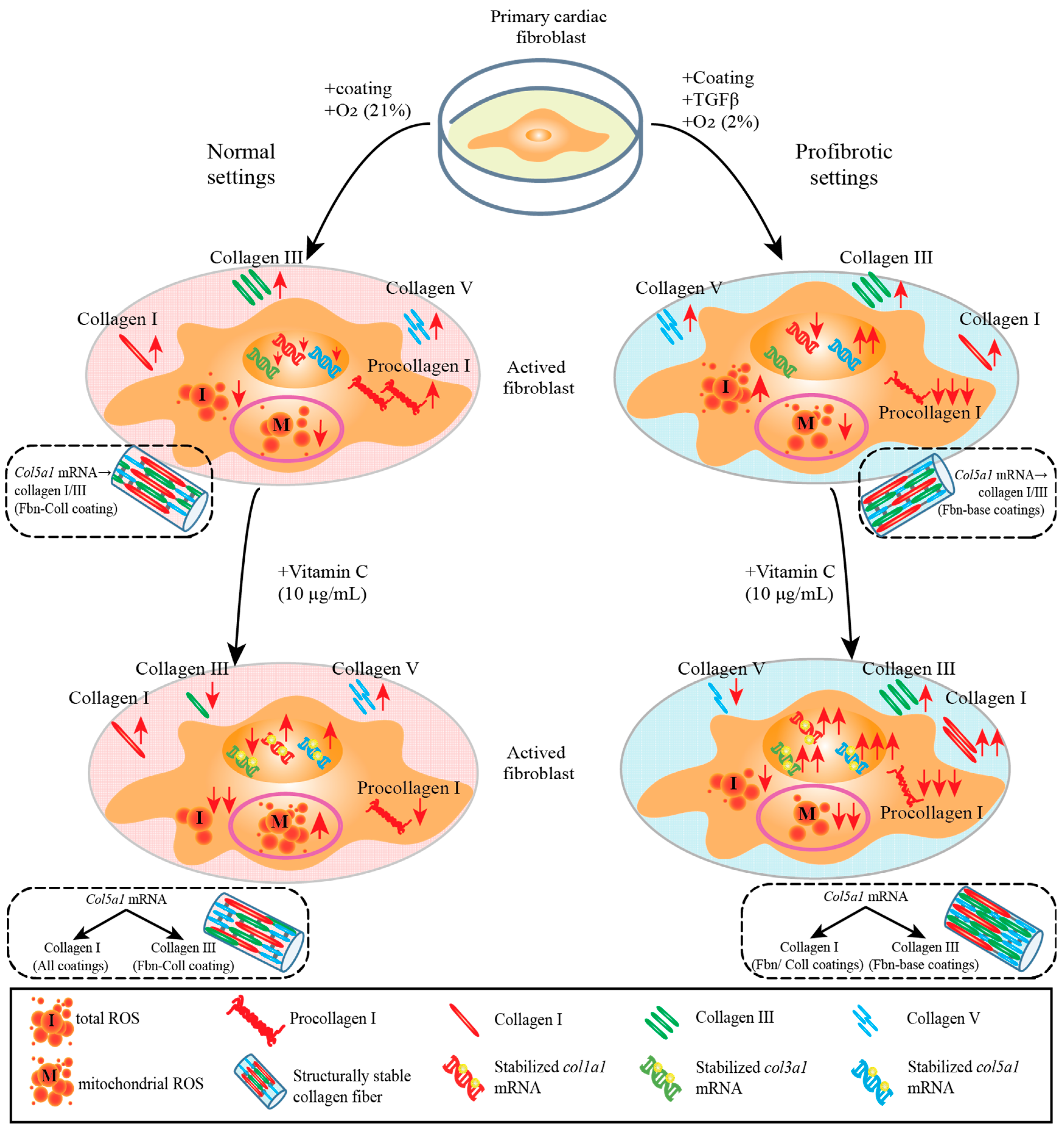
2. Results
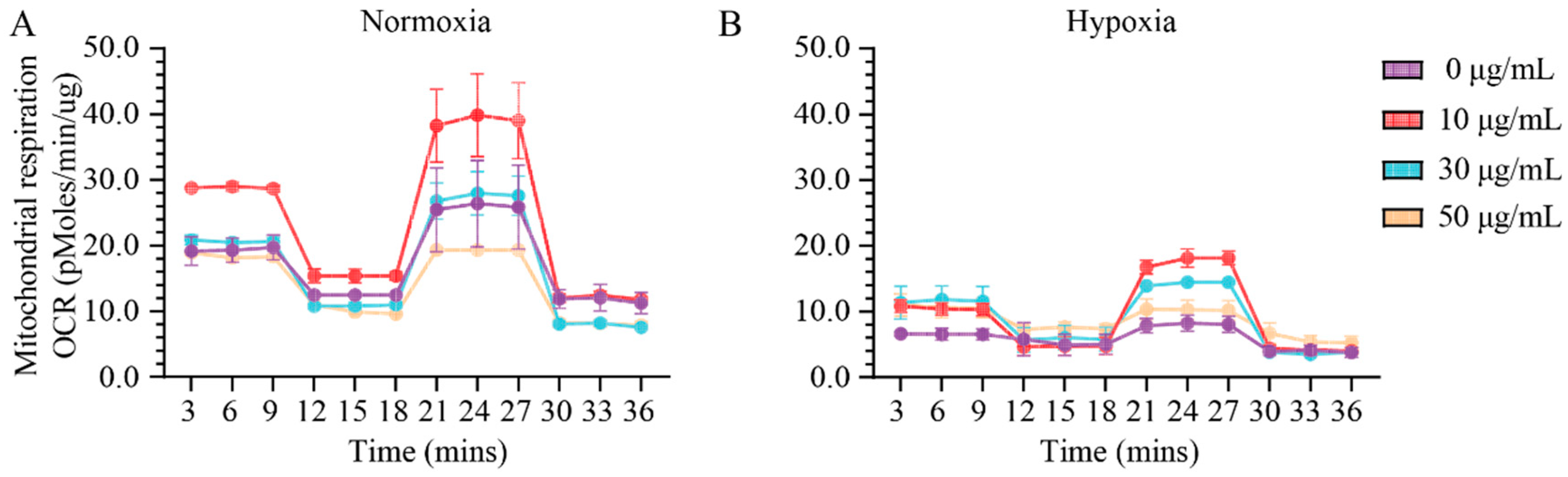
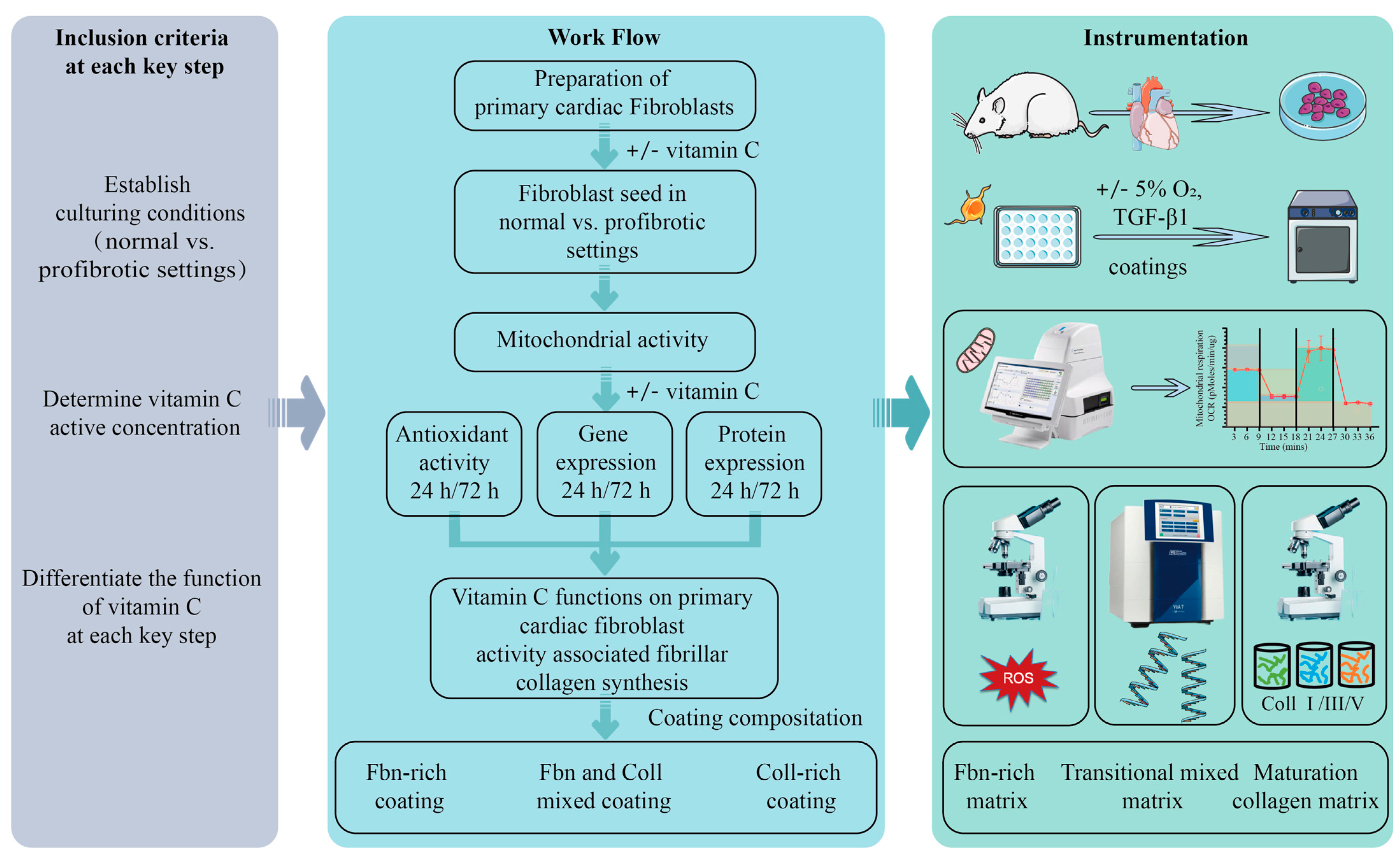
2.1. Assessment of Effect of Vitamin C Activity on Primary Cardiac Fibroblast Metabolism and Treatment Optimization
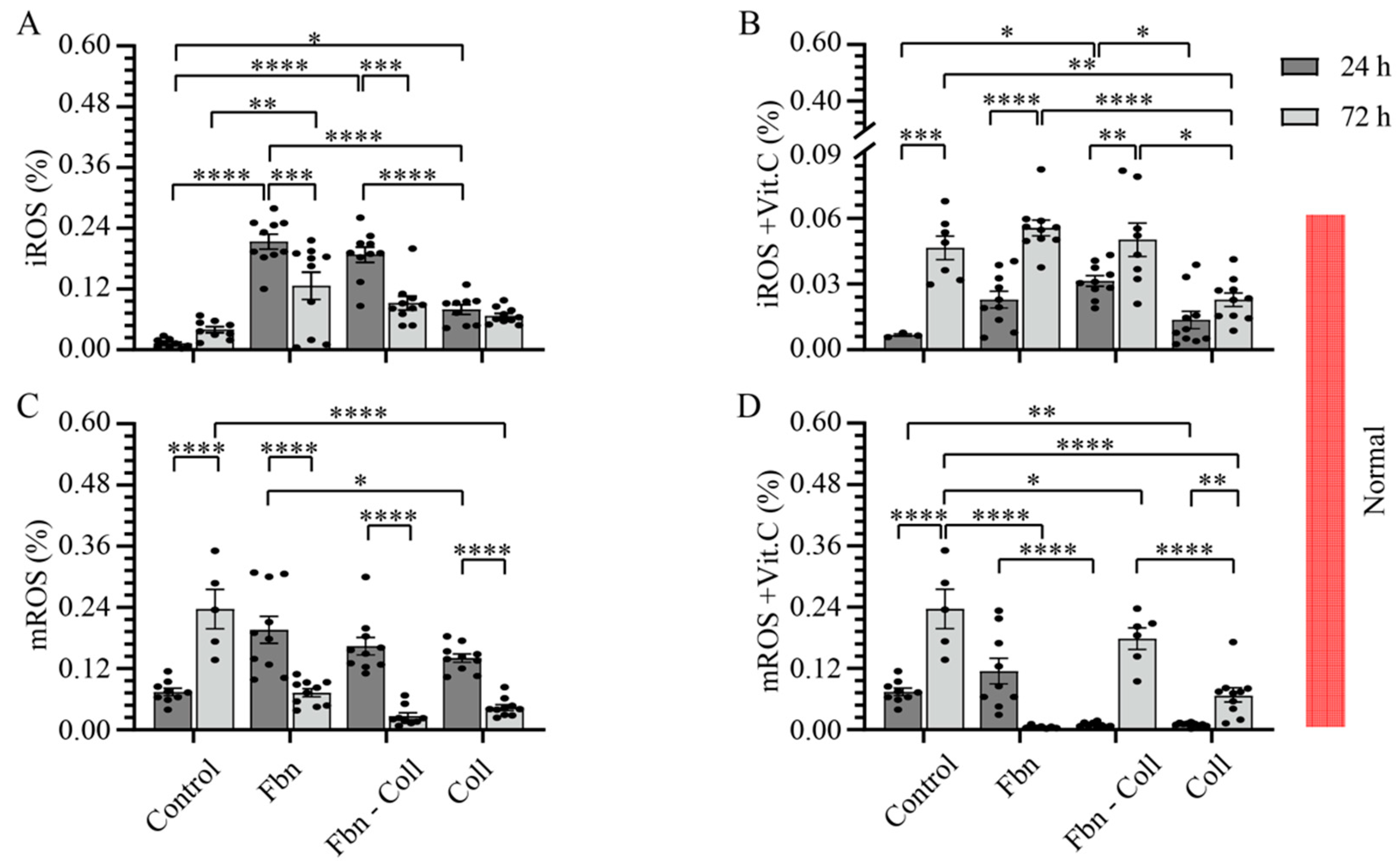
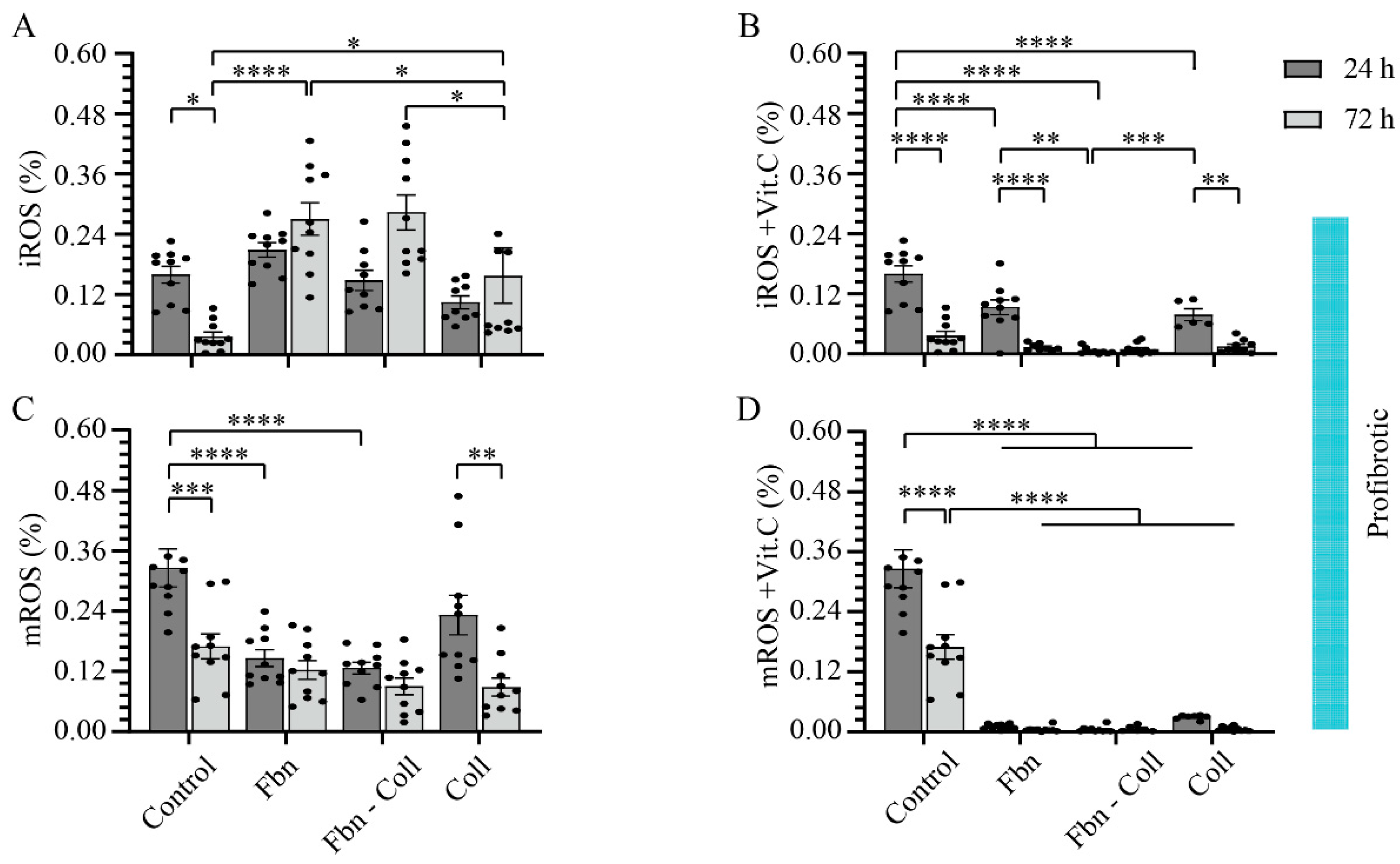
2.2. The Radical Scavenging Role of Vitamin C
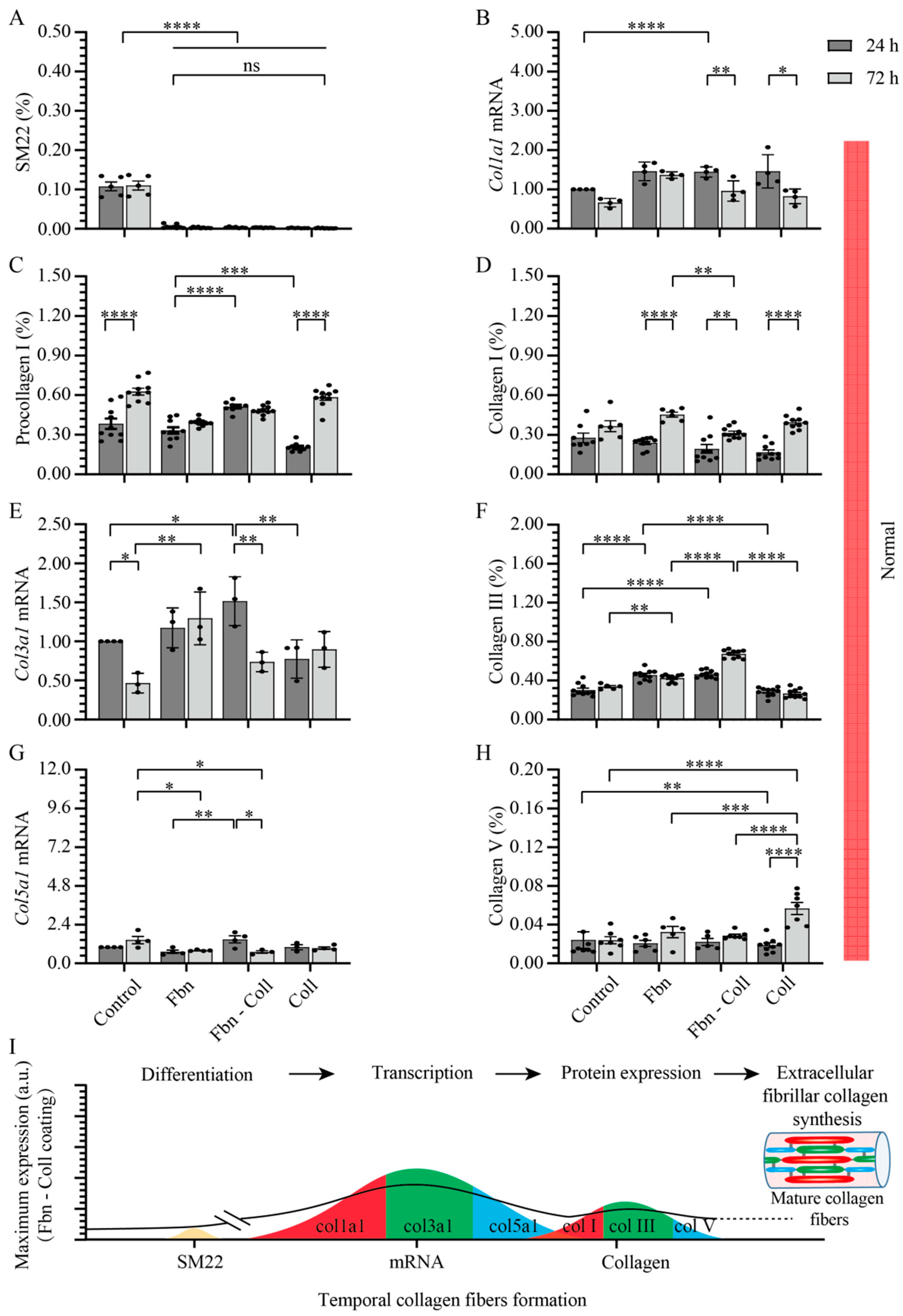
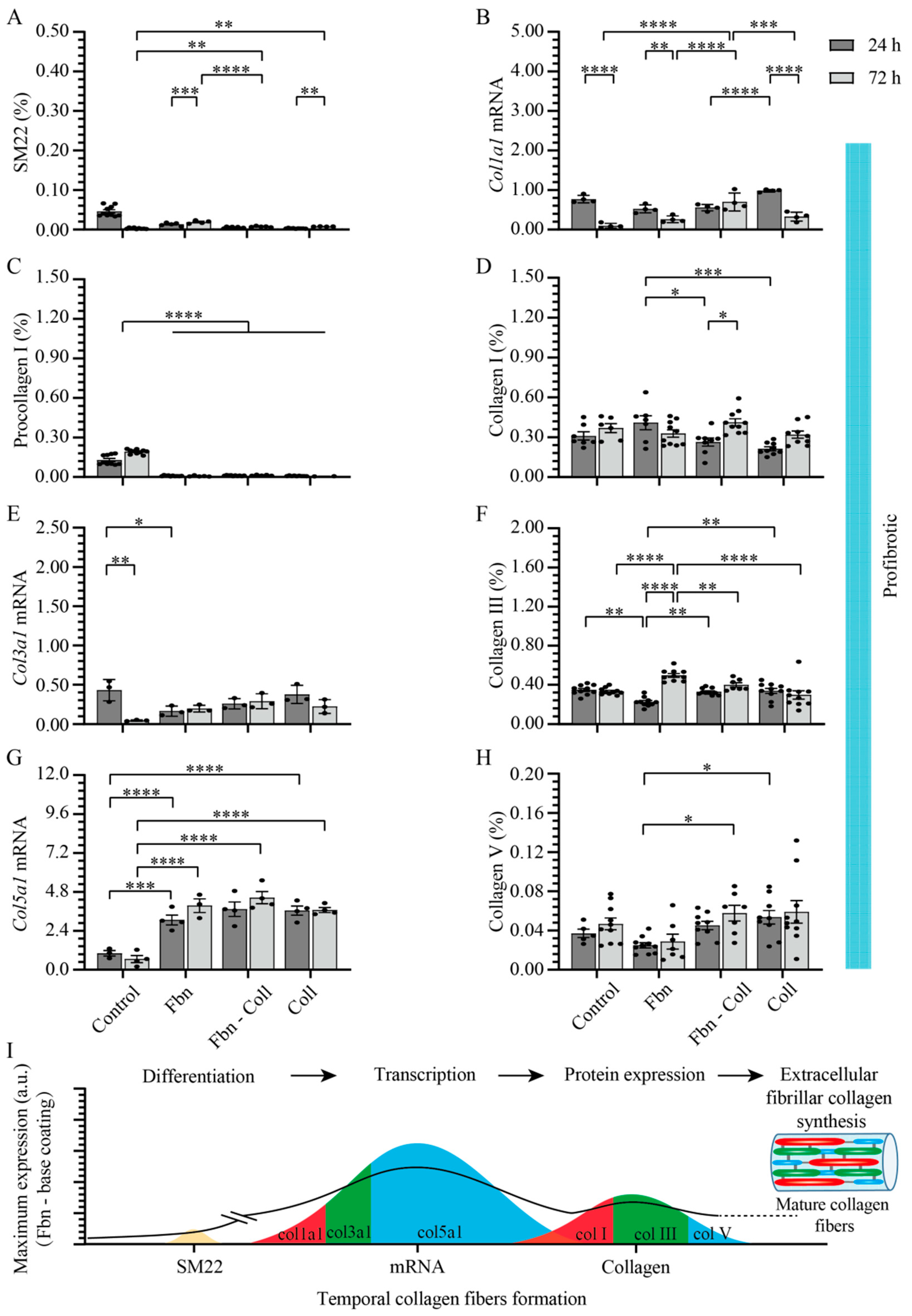
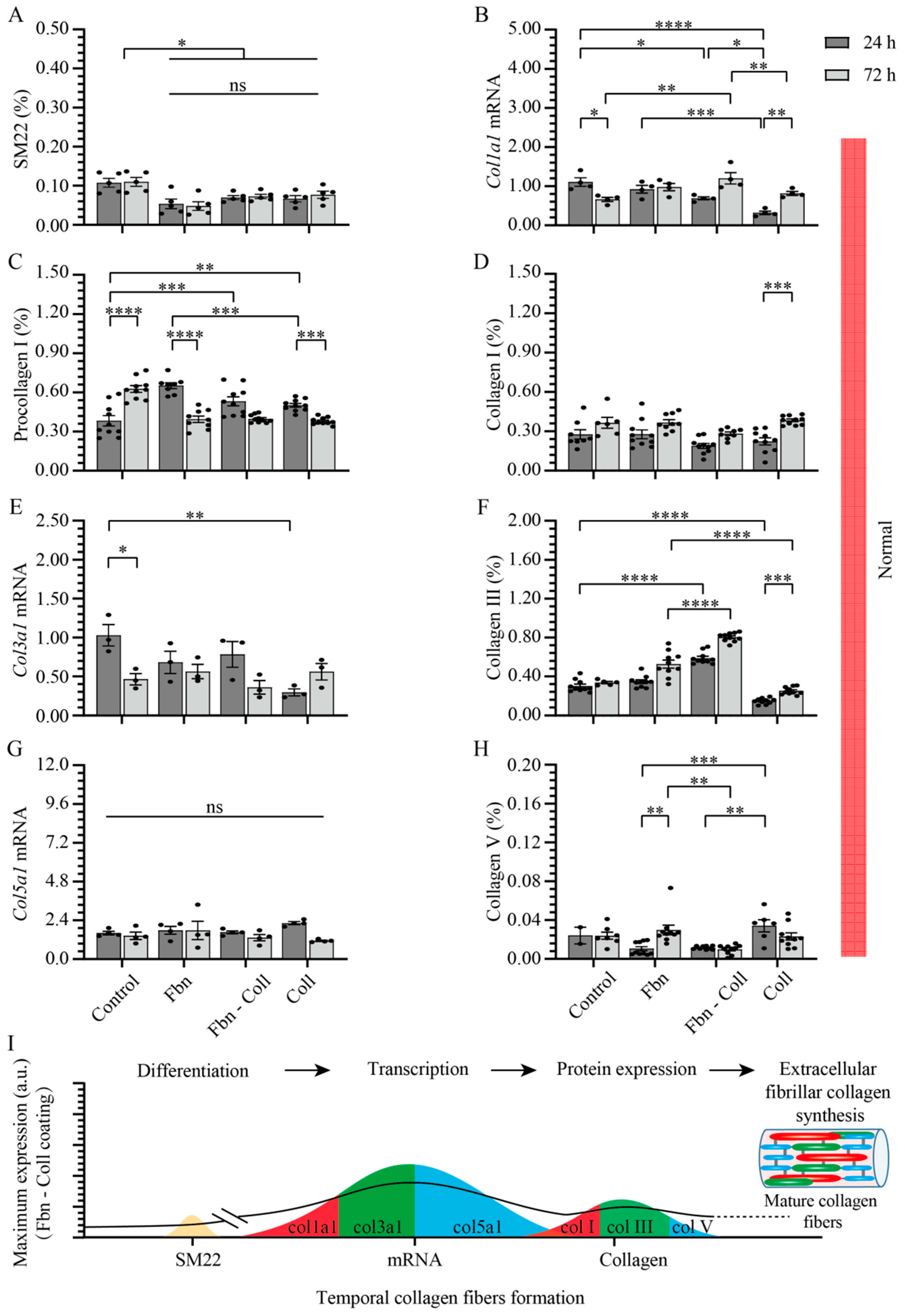
2.3. Substrate Composition Modulate Fibrillar Collagen Synthesis in Normal Settings
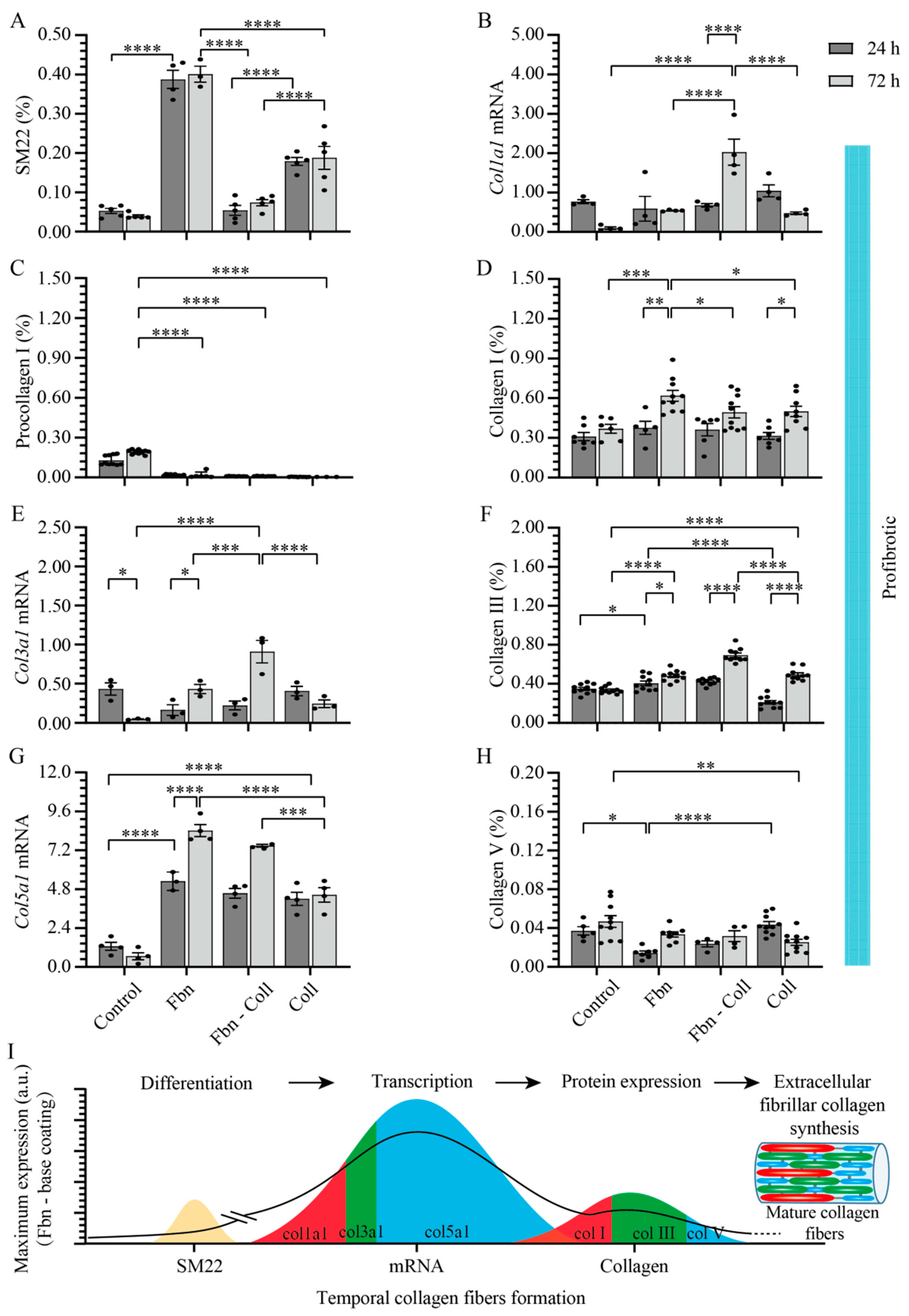
2.4. Active Primary Cardiac Fibroblasts Turned into Profibrotic-Driven col5a1gene Expression
2.5. Vitamin C Regulates Timely the Turnover of De Novo Fibrillar Collagen in Normal Settings
2.6. Vitamin C Effect Associated De Novo Fibrillar Collagen Synthesis Depends on Coating Composition
3. Discussion
3.1. Differentiation
3.2. Antioxidant
3.3. Sustaining mRNA Transcription of ECM Protein
3.4. Modulator of Fibrillar Collagen Synthesis
3.5. Advantages
3.6. Study Limitation
4. Materials and Methods
4.1. Preparation of Coatings for Cell Seeding
4.2. Cell Isolation, Culture, and Treatments
4.3. Analysis of Mitochondrial Activity
4.4. Immunofluorescence
4.5. RNA Isolation and Gene Expression
4.6. Assessment of Mitochondrial and Intracellular ROS
4.7. Data Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ROS | Reactive oxygen species |
| MI | Myocardial infarction |
| ECM | Extracellular matrix |
| iROS | Intracellular total reactive oxygen species |
| mROS | Mitochondrial reactive oxygen species |
| O2•− | Mitochondrial superoxide ions |
| 2D | Two-dimensional |
| DPBS | Dulbecco’s phosphate-buffered saline |
| ATP | Adenosine triphosphate |
| DMEM | Dulbecco′s modified eagle′s medium |
| PAA | Penicillin/Streptomycin |
| TGF-β | Transforming growth factor β |
| FCCP | Carbonyl cyanide-p-trifluoromethoxy phenylhydrazone |
| PFA | Paraformaldehyde |
| DAPI | 4′,6-diamidino-2-phenylindole |
| cDNA | Complementary deoxyribonucleic acid |
| PCR | Polymerase chain reaction |
| mRNA | Messenger ribonucleic acid |
References
- Virani, S.S.; Alonso, A.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics-2020 Update: A Report From the American Heart Association. Circulation 2020, 141, e139–e596. [Google Scholar] [CrossRef]
- Rusu, M.; Hilse, K.; Schuh, A.; Martin, L.; Slabu, I.; Stoppe, C.; Liehn, E.A. Biomechanical assessment of remote and postinfarction scar remodeling following myocardial infarction. Sci. Rep. 2019, 9, 16744. [Google Scholar] [CrossRef]
- Zhang, W.; Liang, J.; Han, P. Cardiac cell type-specific responses to injury and contributions to heart regeneration. Cell Regen. 2021, 10, 4. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, L.; Cheng, J.; Zhang, H. Risk Prediction Model Based on Biomarkers of Remodeling in Patients with Acute Anterior ST-Segment Elevation Myocardial Infarction. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2021, 27, e927404. [Google Scholar] [CrossRef]
- Silva, A.C.; Pereira, C.; Fonseca, A.; Pinto-do-Ó, P.; Nascimento, D.S. Bearing My Heart: The Role of Extracellular Matrix on Cardiac Development, Homeostasis, and Injury Response. Front. Cell Dev. Biol. 2020, 8, 621644. [Google Scholar] [CrossRef]
- DeLeon-Pennell, K.Y.; Barker, T.H.; Lindsey, M.L. Fibroblasts: The arbiters of extracellular matrix remodeling. Matrix Biol. J. Int. Soc. Matrix Biol. 2020, 91–92, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, S.D.; Frangogiannis, N.G. The Biological Basis for Cardiac Repair After Myocardial Infarction: From Inflammation to Fibrosis. Circ. Res. 2016, 119, 91–112. [Google Scholar] [CrossRef] [PubMed]
- Alberts, B.; Bray, D.; Lewis, J.; Raff, M.; Roberts, K.; Watson, J.J.N.Y.G.S. Molecular Biology of the Cell, 3rd ed.; Garland Publishing: New York, NY, USA, 1994. [Google Scholar] [CrossRef]
- Brodsky, B.; Shah, N.K.J.T.F.J. The triple-lielix motif in proteins. FASEB J. 1995, 9, 1537–1546. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, H.; Hanna, A.; Humeres, C.; Frangogiannis, N.G. Properties and Functions of Fibroblasts and Myofibroblasts in Myocardial Infarction. Cells 2022, 11, 1386. [Google Scholar] [CrossRef]
- Humeres, C.; Shinde, A.V.; Hanna, A.; Alex, L.; Hernández, S.C.; Li, R.; Chen, B.; Conway, S.J.; Frangogiannis, N.G. Smad7 effects on TGF-β and ErbB2 restrain myofibroblast activation and protect from postinfarction heart failure. J. Clin. Investig. 2022, 132, e146926. [Google Scholar] [CrossRef]
- Martinez-Garcia, G.; Rodriguez-Ramos, M.; Santos-Medina, M.; Carrero-Vazquez, A.M.; Chipi-Rodriguez, Y. New model predicts in-hospital complications in myocardial infarction. Discoveries 2022, 10, e142. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Morelli, M.B.; Matarese, A.; Sardu, C.; Santulli, G. Cardiomyocyte-derived exosomal microRNA-92a mediates post-ischemic myofibroblast activation both in vitro and ex vivo. ESC Heart Fail. 2020, 7, 284–288. [Google Scholar] [CrossRef]
- Cadenas, S. ROS and redox signaling in myocardial ischemia-reperfusion injury and cardioprotection. Free. Radic. Biol. Med. 2018, 117, 76–89. [Google Scholar] [CrossRef]
- Chen, C.L.; Zhang, L.; Jin, Z.; Kasumov, T.; Chen, Y.R. Mitochondrial redox regulation and myocardial ischemia-reperfusion injury. Am. J. Physiol. Cell Physiol. 2022, 322, C12–C23. [Google Scholar] [CrossRef]
- Caggiano, L.R.; Lee, J.J.; Holmes, J.W. Surgical reinforcement alters collagen alignment and turnover in healing myocardial infarcts. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H1041–H1050. [Google Scholar] [CrossRef] [PubMed]
- French, B.A.; Holmes, J.W. Implications of scar structure and mechanics for post-infarction cardiac repair and regeneration. Exp. Cell Res. 2019, 376, 98–103. [Google Scholar] [CrossRef]
- Libby, P.; Aikawa, M. Vitamin C, collagen, and cracks in the plaque. Circulation 2002, 105, 1396–1398. [Google Scholar] [CrossRef] [PubMed]
- Cleutjens, J.P.; Verluyten, M.J.; Smiths, J.F.; Daemen, M.J. Collagen remodeling after myocardial infarction in the rat heart. Am. J. Pathol. 1995, 147, 325–338. [Google Scholar]
- Frangogiannis, N.G.J.C.P. Pathophysiology of Myocardial Infarction. Compr. Physiol. 2015, 5, 1841–1875. [Google Scholar] [CrossRef]
- Karimpour, H.; Bahrami, A.; Amini, S.; Rezaei, M.; Amini-Saman, J.; Shahbazi, F.J.J.P.R.I. Effects of a high dose of vitamin C along with thiamine in critically-ill patients with septic shock: A preliminary study. J. Pharm. Res. Int. 2019, 29, 1–7. [Google Scholar] [CrossRef]
- Morelli, M.B.; Gambardella, J.; Castellanos, V.; Trimarco, V.; Santulli, G. Vitamin C and Cardiovascular Disease: An Update. Antioxidants 2020, 9, 1227. [Google Scholar] [CrossRef] [PubMed]
- Traber, M.G.; Leonard, S.W.; Vasu, V.T.; Morrissey, B.M.; Lei, H.J.; Atkinson, J.; Cross, C.E. α-Tocopherol Pharmacokinetics in Adults with Cystic Fibrosis: Benefits of Supplemental Vitamin C Administration. Nutrients 2022, 14, 3717. [Google Scholar] [CrossRef]
- Kouakanou, L.; Peters, C.; Brown, C.E.; Kabelitz, D.; Wang, L.D. Vitamin C, From Supplement to Treatment: A Re-Emerging Adjunct for Cancer Immunotherapy? Front. Immunol. 2021, 12, 765906. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, J.C.; Rouleau, T.; Chessex, P. Interaction between ascorbate and light-exposed riboflavin induces lung remodeling. J. Pharmacol. Exp. Ther. 2004, 311, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Izzo, R.; Trimarco, V.; Mone, P.; Aloè, T.; Capra Marzani, M.; Diana, A.; Fazio, G.; Mallardo, M.; Maniscalco, M.; Marazzi, G.; et al. Combining L-Arginine with vitamin C improves long-COVID symptoms: The LINCOLN Survey. Pharmacol. Res. 2022, 183, 106360. [Google Scholar] [CrossRef]
- Shah, A.K.; Dhalla, N.S. Effectiveness of Some Vitamins in the Prevention of Cardiovascular Disease: A Narrative Review. Front. Physiol. 2021, 12, 729255. [Google Scholar] [CrossRef]
- Nyyssönen, K.; Parviainen, M.T.; Salonen, R.; Tuomilehto, J.; Salonen, J.T. Vitamin C deficiency and risk of myocardial infarction: Prospective population study of men from eastern Finland. BMJ 1997, 314, 634–638. [Google Scholar] [CrossRef]
- Malik, A.; Bagchi, A.K.; Vinayak, K.; Akolkar, G.; Slezak, J.; Belló-Klein, A.; Jassal, D.S.; Singal, P.K. Vitamin C: Historical perspectives and heart failure. Heart Fail. Rev. 2021, 26, 699–709. [Google Scholar] [CrossRef]
- Rodrigo, R.; Prieto, J.C.; Aguayo, R.; Ramos, C.; Puentes, Á.; Gajardo, A.; Panieri, E.; Rojas-Solé, C.; Lillo-Moya, J.; Saso, L. Joint Cardioprotective Effect of Vitamin C and Other Antioxidants against Reperfusion Injury in Patients with Acute Myocardial Infarction Undergoing Percutaneous Coronary Intervention. Molecules 2021, 26, 5702. [Google Scholar] [CrossRef]
- Shafaei-Bajestani, N.; Talasaz, A.H.; Salarifar, M.; Pourhosseini, H.; Sadri, F.; Jalali, A. Potential Role of Vitamin C Intracoronary Administration in Preventing Cardiac Injury After Primary Percutaneous Coronary Intervention in Patients with ST-Elevation Myocardial Infarction. J. Res. Pharm. Pract. 2019, 8, 75–82. [Google Scholar] [CrossRef]
- DePhillipo, N.N.; Aman, Z.S.; Kennedy, M.I.; Begley, J.P.; Moatshe, G.; LaPrade, R.F. Efficacy of Vitamin C Supplementation on Collagen Synthesis and Oxidative Stress After Musculoskeletal Injuries: A Systematic Review. Orthop. J. Sport. Med. 2018, 6, 2325967118804544. [Google Scholar] [CrossRef]
- Liugan, M.; Carr, A.C. Vitamin C and Neutrophil Function: Findings from Randomized Controlled Trials. Nutrients 2019, 11, 2102. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Qu, Y.; Yu, J.; Song, L.; Chen, S.; Qin, Z.; Gong, J.; Zhan, H.; Gao, Y.; Zhang, J. A chitosan-vitamin C based injectable hydrogel improves cell survival under oxidative stress. Int. J. Biol. Macromol. 2022, 202, 102–111. [Google Scholar] [CrossRef]
- Sacks, B.; Onal, H.; Martorana, R.; Sehgal, A.; Harvey, A.; Wastella, C.; Ahmad, H.; Ross, E.; Pjetergjoka, A.; Prasad, S.; et al. Mitochondrial targeted antioxidants, mitoquinone and SKQ1, not vitamin C, mitigate doxorubicin-induced damage in H9c2 myoblast: Pretreatment vs. co-treatment. BMC Pharm. Toxicol 2021, 22, 49. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Jana, N.R. Vitamin C-Conjugated Nanoparticle Protects Cells from Oxidative Stress at Low Doses but Induces Oxidative Stress and Cell Death at High Doses. ACS Appl. Mater. Interfaces 2017, 9, 41807–41817. [Google Scholar] [CrossRef]
- Fuchs, A.A.; Balne, P.K.; Giuliano, E.A.; Sinha, N.R.; Mohan, R.R. Evaluation of a novel combination of TRAM-34 and ascorbic acid for the treatment of corneal fibrosis in vivo. PLoS ONE 2022, 17, e0262046. [Google Scholar] [CrossRef]
- Piersma, B.; Wouters, O.Y.; de Rond, S.; Boersema, M.; Gjaltema, R.A.F.; Bank, R.A. Ascorbic acid promotes a TGFβ1-induced myofibroblast phenotype switch. Physiol. Rep. 2017, 5, e13324. [Google Scholar] [CrossRef]
- Del Monte-Nieto, G.; Fischer, J.W.; Gorski, D.J.; Harvey, R.P.; Kovacic, J.C. Basic Biology of Extracellular Matrix in the Cardiovascular System, Part 1/4: JACC Focus Seminar. J. Am. Coll. Cardiol. 2020, 75, 2169–2188. [Google Scholar] [CrossRef]
- Valdecantos, M.P.; Pérez-Matute, P.; Quintero, P.; Martínez, J.A. Vitamin C, resveratrol and lipoic acid actions on isolated rat liver mitochondria: All antioxidants but different. Redox Rep. 2010, 15, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Parhizkar, E.; Rashedinia, M.; Karimi, M.; Alipour, S. Design and development of vitamin C-encapsulated proliposome with improved in-vitro and ex-vivo antioxidant efficacy. J. Microencapsul. 2018, 35, 301–311. [Google Scholar] [CrossRef]
- Davison, A.J.; Kettle, A.J.; Fatur, D.J. Mechanism of the inhibition of catalase by ascorbate. Roles of active oxygen species, copper and semidehydroascorbate. J. Biol. Chem. 1986, 261, 1193–1200. [Google Scholar] [CrossRef]
- Gaikwad, S.; Chakraborty, A.; Salwe, S.; Patel, V.; Kulkarni, S.; Banerjee, S. Juglone-ascorbic acid synergy inhibits metastasis and induces apoptotic cell death in poorly differentiated thyroid carcinoma by perturbing SOD and catalase activities. J. Biochem. Mol. Toxicol. 2018, 32, e22176. [Google Scholar] [CrossRef]
- Felipe, K.B.; Benites, J.; Glorieux, C.; Sid, B.; Valenzuela, M.; Kviecinski, M.R.; Pedrosa, R.C.; Valderrama, J.A.; Levêque, P.; Gallez, B.; et al. Antiproliferative effects of phenylaminonaphthoquinones are increased by ascorbate and associated with the appearance of a senescent phenotype in human bladder cancer cells. Biochem. Biophys. Res. Commun. 2013, 433, 573–578. [Google Scholar] [CrossRef]
- Ourique, F.; Kviecinski, M.R.; Zirbel, G.; Castro, L.; Gomes Castro, A.J.; Mena Barreto Silva, F.R.; Valderrama, J.A.; Rios, D.; Benites, J.; Calderon, P.B.; et al. In vivo inhibition of tumor progression by 5 hydroxy-1,4-naphthoquinone (juglone) and 2-(4-hydroxyanilino)-1,4-naphthoquinone (Q7) in combination with ascorbate. Biochem. Biophys. Res. Commun. 2016, 477, 640–646. [Google Scholar] [CrossRef]
- Su, X.; Shen, Z.; Yang, Q.; Sui, F.; Pu, J.; Ma, J.; Ma, S.; Yao, D.; Ji, M.; Hou, P. Vitamin C kills thyroid cancer cells through ROS-dependent inhibition of MAPK/ERK and PI3K/AKT pathways via distinct mechanisms. Theranostics 2019, 9, 4461. [Google Scholar] [CrossRef]
- Bogenberger, J.M.; DeLeon, T.T.; Arora, M.; Ahn, D.H.; Borad, M.J. Emerging role of precision medicine in biliary tract cancers. NPJ Precis. Oncol. 2018, 2, 21. [Google Scholar] [CrossRef]
- Szarka, A.; Kapuy, O.; Lőrincz, T.; Bánhegyi, G. Vitamin C and Cell Death. Antioxid. Redox Signal. 2021, 34, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Lőrincz, T.; Holczer, M.; Kapuy, O.; Szarka, A. The Interrelationship of Pharmacologic Ascorbate Induced Cell Death and Ferroptosis. Pathol. Oncol. Res. 2019, 25, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Rouleau, L.; Antony, A.N.; Bisetto, S.; Newberg, A.; Doria, C.; Levine, M.; Monti, D.A.; Hoek, J.B. Synergistic effects of ascorbate and sorafenib in hepatocellular carcinoma: New insights into ascorbate cytotoxicity. Free. Radic. Biol. Med. 2016, 95, 308–322. [Google Scholar] [CrossRef] [PubMed]
- Padayatty, S.J.; Levine, M. Vitamin C: The known and the unknown and Goldilocks. Oral Dis 2016, 22, 463–493. [Google Scholar] [CrossRef] [PubMed]
- Lykkesfeldt, J.; Michels, A.J.; Frei, B. Vitamin C. Adv. Nutr. 2014, 5, 16–18. [Google Scholar] [CrossRef] [PubMed]
- Sauberlich, H.E. Pharmacology of vitamin C. Annu. Rev. Nutr. 1994, 14, 371–391. [Google Scholar] [CrossRef]
- Schumacher, D.; Curaj, A.; Staudt, M.; Simsekyilmaz, S.; Kanzler, I.; Boor, P.; Klinkhammer, B.M.; Li, X.; Bucur, O.; Kaabi, A.; et al. Endogenous Modulation of Extracellular Matrix Collagen during Scar Formation after Myocardial Infarction. Int. J. Mol. Sci. 2022, 23, 14571. [Google Scholar] [CrossRef]
- Cho, S.; Paik, D.T.; Wu, J.C. An extracellular matrix paradox in myocardial scar formation. Signal Transduct. Target. Ther. 2020, 5, 151. [Google Scholar] [CrossRef] [PubMed]
- Saadat, S.; Noureddini, M.; Mahjoubin-Tehran, M.; Nazemi, S.; Shojaie, L.; Aschner, M.; Maleki, B.; Abbasi-Kolli, M.; Rajabi Moghadam, H.; Alani, B.; et al. Pivotal Role of TGF-β/Smad Signaling in Cardiac Fibrosis: Non-coding RNAs as Effectual Players. Front. Cardiovasc. Med. 2020, 7, 588347. [Google Scholar] [CrossRef] [PubMed]
- Johnston, E.F.; Gillis, T.E. Transforming growth factor beta-1 (TGF-β1) stimulates collagen synthesis in cultured rainbow trout cardiac fibroblasts. J. Exp. Biol. 2017, 220, 2645–2653. [Google Scholar] [CrossRef] [PubMed]
- Kuivaniemi, H.; Tromp, G. Type III collagen (COL3A1): Gene and protein structure, tissue distribution, and associated diseases. Gene 2019, 707, 151–171. [Google Scholar] [CrossRef]
- Saunders, J.T.; Schwarzbauer, J.E. Fibronectin matrix as a scaffold for procollagen proteinase binding and collagen processing. Mol. Biol. Cell 2019, 30, 2218–2226. [Google Scholar] [CrossRef]
- Carr, A.C.; Maggini, S. Vitamin C and Immune Function. Nutrients 2017, 9, 1211. [Google Scholar] [CrossRef]
- Palano, G.; Jansson, M.; Backmark, A.; Martinsson, S.; Sabirsh, A.; Hultenby, K.; Åkerblad, P.; Granberg, K.L.; Jennbacken, K.; Müllers, E.; et al. A high-content, in vitro cardiac fibrosis assay for high-throughput, phenotypic identification of compounds with anti-fibrotic activity. J. Mol. Cell. Cardiol. 2020, 142, 105–117. [Google Scholar] [CrossRef]
- Paten, J.A.; Martin, C.L.; Wanis, J.T.; Siadat, S.M.; Figueroa-Navedo, A.M.; Ruberti, J.W.; Deravi, L.F. Molecular Interactions between Collagen and Fibronectin: A Reciprocal Relationship that Regulates De Novo Fibrillogenesis. Chem 2019, 5, 2126–2145. [Google Scholar] [CrossRef]
- Pinkas, D.M.; Ding, S.; Raines, R.T.; Barron, A.E. Tunable, Post-translational Hydroxylation of Collagen Domains in Escherichia coli. ACS Chem. Biol. 2011, 6, 320–324. [Google Scholar] [CrossRef]
- Li, L.; Ma, Y.; He, G.; Ma, S.; Wang, Y.; Sun, Y. Pilose antler extract restores type I and III collagen to accelerate wound healing. Biomed. Pharmacother. 2023, 161, 114510. [Google Scholar] [CrossRef]
- Raziyeva, K.; Kim, Y.; Zharkinbekov, Z.; Temirkhanova, K.; Saparov, A. Novel Therapies for the Treatment of Cardiac Fibrosis Following Myocardial Infarction. Biomedicines 2022, 10, 2178. [Google Scholar] [CrossRef] [PubMed]
- Mircea, A.A. Post-myocardial infarction treatment with resiniferatoxin modulates the expression of important genes involved in inflammation, plaque stability and angiogenesis. Discoveries 2023, 11, e163. [Google Scholar] [CrossRef]
- Curaj, A.; Schumacher, D.; Rusu, M.; Staudt, M.; Li, X.; Simsekyilmaz, S.; Jankowski, V.; Jankowski, J.; Dumitrascu, A.R.; Hausenloy, D.J.; et al. Neutrophils Modulate Fibroblast Function and Promote Healing and Scar Formation after Murine Myocardial Infarction. Int. J. Mol. Sci. 2020, 21, 3685. [Google Scholar] [CrossRef] [PubMed]
- Doppler, S.A.; Carvalho, C.; Lahm, H.; Deutsch, M.A.; Dreßen, M.; Puluca, N.; Lange, R.; Krane, M. Cardiac fibroblasts: More than mechanical support. J. Thorac. Dis. 2017, 9, S36–S51. [Google Scholar] [CrossRef]
- Nim, H.T.; Furtado, M.B.; Costa, M.W.; Kitano, H.; Rosenthal, N.A.; Boyd, S.E. CARFMAP: A Curated Pathway Map of Cardiac Fibroblasts. PLoS ONE 2015, 10, e0143274. [Google Scholar] [CrossRef]
- Ivey, M.J.; Tallquist, M.D. Defining the Cardiac Fibroblast. Circ. J. Off. J. Jpn. Circ. Soc. 2016, 80, 2269–2276. [Google Scholar] [CrossRef] [PubMed]
- Duband, J.L.; Gimona, M.; Scatena, M.; Sartore, S.; Small, J.V. Calponin and SM 22 as differentiation markers of smooth muscle: Spatiotemporal distribution during avian embryonic development. Differentiation 1993, 55, 1–11. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Faber, J.; Fonseca, L.M. How sample size influences research outcomes. Dent. Press J. Orthod. 2014, 19, 29. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequences | Product Length | Annealing Temp. (°C) | GenBank Numbers |
|---|---|---|---|---|
| Col1a1 | 5′-GCTCCTCTTAGGGGCCACT (Forward) | 103 bp | 60 °C | NM_ 007742.4 |
| 5′-CCACGTCTCACCATTGGGG (Reverse) | ||||
| Col3a1 | 5′-TCTGAGCTGCTTCTTCCTCTCT (Forward) | 98 bp | 60 °C | NM_ 009930.2 |
| 5′-GAAGAAACCAGGTTCCACTTTG(Reverse) | ||||
| Col5a1 | 5′-GCTACTCCTGTTCCTGCTGC (Forward) | 100 bp | 60 °C | NM_ 006497644.5 |
| 5′-TGAGGGCAAATTGTGAAAATC (Reverse) | ||||
| β-actin | 5′-AGCCATGTACGTAGCCATCC (Forward) | 228 bp | 60 °C | NM_ 007393.5 |
| 5′-CTCTCAGCTGTGGTGGTGAA (Reverse) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Zheng, H.; Nilcham, P.; Bucur, O.; Vogt, F.; Slabu, I.; Liehn, E.A.; Rusu, M. Vitamin C Regulates the Profibrotic Activity of Fibroblasts in In Vitro Replica Settings of Myocardial Infarction. Int. J. Mol. Sci. 2023, 24, 8379. https://doi.org/10.3390/ijms24098379
Xu Y, Zheng H, Nilcham P, Bucur O, Vogt F, Slabu I, Liehn EA, Rusu M. Vitamin C Regulates the Profibrotic Activity of Fibroblasts in In Vitro Replica Settings of Myocardial Infarction. International Journal of Molecular Sciences. 2023; 24(9):8379. https://doi.org/10.3390/ijms24098379
Chicago/Turabian StyleXu, Yichen, Huabo Zheng, Pakhwan Nilcham, Octavian Bucur, Felix Vogt, Ioana Slabu, Elisa Anamaria Liehn, and Mihaela Rusu. 2023. "Vitamin C Regulates the Profibrotic Activity of Fibroblasts in In Vitro Replica Settings of Myocardial Infarction" International Journal of Molecular Sciences 24, no. 9: 8379. https://doi.org/10.3390/ijms24098379
APA StyleXu, Y., Zheng, H., Nilcham, P., Bucur, O., Vogt, F., Slabu, I., Liehn, E. A., & Rusu, M. (2023). Vitamin C Regulates the Profibrotic Activity of Fibroblasts in In Vitro Replica Settings of Myocardial Infarction. International Journal of Molecular Sciences, 24(9), 8379. https://doi.org/10.3390/ijms24098379