
Blue Native PAGE–Antibody Shift in Conjunction with Mass Spectrometry to Reveal Protein Subcomplexes: Detection of a Cerebellar α1/α6-Subunits Containing γ-Aminobutyric Acid Type A Receptor Subtype

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Identification of the α1 Subunit in α6-Containing GABAAR Complexes by Affinity Purification
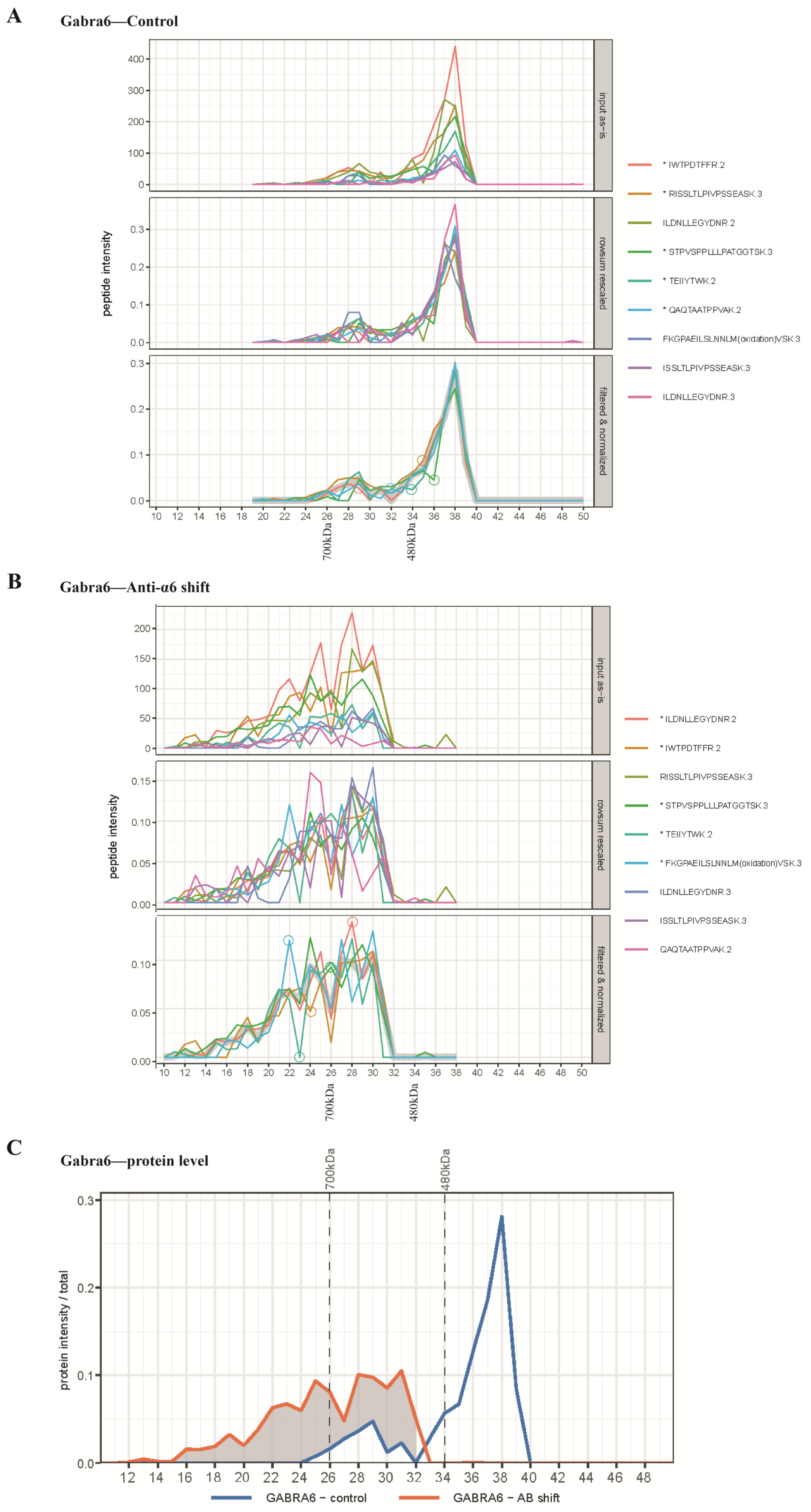
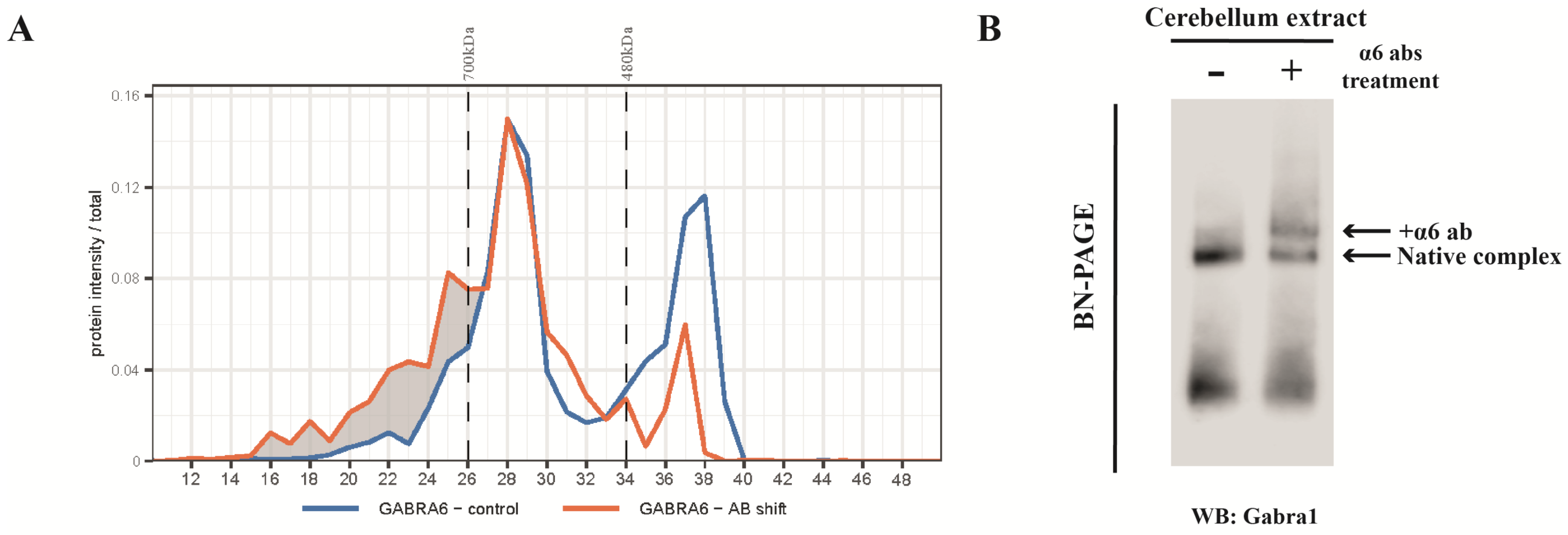
2.2. Antibody Shift BN-PAGE-MS Reveals Multiple Forms of α1- and α6-Containing Complexes
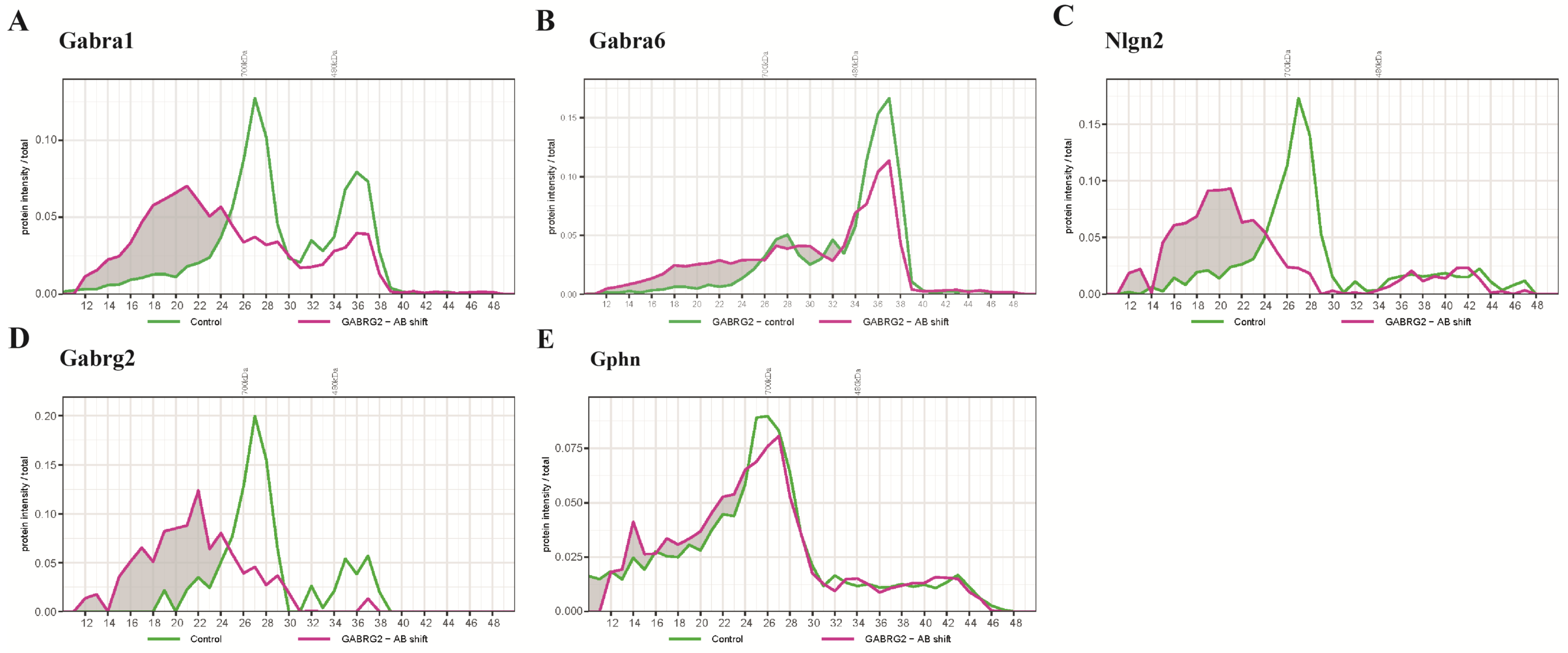
2.3. BN-PAGE-MS Antibody Shift to Reveal the Migration Patterns of Other GABAAR Subunits and Their Associated Proteins
2.4. The Subcellular Localization of the α1- and α6-Subunit-Containing Receptors in Cultured Cerebellar Neurons
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Klaassen, R.V.; Stroeder, J.; Coussen, F.; Hafner, A.-S.; Petersen, J.D.; Renancio, C.; Schmitz, L.J.M.; Normand, E.; Lodder, J.C.; Rotaru, D.C.; et al. Shisa6 Traps AMPA Receptors at Postsynaptic Sites and Prevents Their Desensitization during Synaptic Activity. Nat. Commun. 2016, 7, 10682. [Google Scholar] [CrossRef]
- Straub, C.; Tomita, S. The Regulation of Glutamate Receptor Trafficking and Function by TARPs and Other Transmembrane Auxiliary Subunits. Curr. Opin. Neurobiol. 2012, 22, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Stern, P. Dancing with AMPARs. Sci. Signal. 2010, 3, ec91. [Google Scholar] [CrossRef]
- van der Spek, S.J.F.; Koopmans, F.; Paliukhovich, I.; Ramsden, S.L.; Harvey, K.; Harvey, R.J.; Smit, A.B.; Li, K.W. Glycine Receptor Complex Analysis Using Immunoprecipitation-Blue Native Gel Electrophoresis-Mass Spectrometry. Proteomics 2020, 20, e1900403. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Pandya, N.J.; Koopmans, F.; Castelo-Székelv, V.; van der Schors, R.C.; Smit, A.B.; Li, K.W. Interaction Proteomics Reveals Brain Region-Specific AMPA Receptor Complexes. J. Proteome Res. 2014, 13, 5695–5706. [Google Scholar] [CrossRef] [PubMed]
- Nijtmans, L.G.J.; Henderson, N.S.; Holt, I.J. Blue Native Electrophoresis to Study Mitochondrial and Other Protein Complexes. Methods 2002, 26, 327–334. [Google Scholar] [CrossRef]
- Martenson, J.S.; Yamasaki, T.; Chaudhury, N.H.; Albrecht, D.; Tomita, S. Assembly Rules for GABA(A) Receptor Complexes in the Brain. Elife 2017, 6, e27443. [Google Scholar] [CrossRef] [PubMed]
- Dewson, G. Blue Native PAGE and Antibody Gel Shift to Assess Bak and Bax Conformation Change and Oligomerization. Cold Spring Harb. Protoc. 2015, 2015, 485–489. [Google Scholar] [CrossRef]
- Swamy, M.; Minguet, S.; Siegers, G.M.; Alarcón, B.; Schamel, W.W.A. A Native Antibody-Based Mobility-Shift Technique (NAMOS-Assay) to Determine the Stoichiometry of Multiprotein Complexes. J. Immunol. Methods 2007, 324, 74–83. [Google Scholar] [CrossRef]
- Swamy, M.; Siegers, G.M.; Fiala, G.J.; Molnar, E.; Dopfer, E.P.; Fisch, P.; Schraven, B.; Schamel, W.W.A. Stoichiometry and Intracellular Fate of TRIM-Containing TCR Complexes. Cell Commun. Signal. 2010, 8, 5. [Google Scholar] [CrossRef]
- Wang, F.; Wang, L.; Xu, Z.; Liang, G. Identification and Analysis of Multi-Protein Complexes in Placenta. PLoS ONE 2013, 8, e62988. [Google Scholar] [CrossRef] [PubMed]
- Sieghart, W.; Sperk, G. Subunit Composition, Distribution and Function of GABA(A) Receptor Subtypes. Curr. Top. Med. Chem. 2002, 2, 795–816. [Google Scholar] [CrossRef] [PubMed]
- Günther, U.; Benson, J.; Benke, D.; Fritschy, J.M.; Reyes, G.; Knoflach, F.; Crestani, F.; Aguzzi, A.; Arigoni, M.; Lang, Y.; et al. Benzodiazepine-Insensitive Mice Generated by Targeted Disruption of the Gamma 2 Subunit Gene of Gamma-Aminobutyric Acid Type A Receptors. Proc. Natl. Acad. Sci. USA 1995, 92, 7749–7753. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.; Korpi, E.R.; McKernan, R.M.; Pelz, R.; Nusser, Z.; Mäkelä, R.; Mellor, J.R.; Pollard, S.; Bahn, S.; Stephenson, F.A.; et al. Ligand-Gated Ion Channel Subunit Partnerships: GABAA Receptor Alpha6 Subunit Gene Inactivation Inhibits Delta Subunit Expression. J. Neurosci. 1997, 17, 1350–1362. [Google Scholar] [CrossRef] [PubMed]
- Farrant, M.; Nusser, Z. Variations on an Inhibitory Theme: Phasic and Tonic Activation of GABA(A) Receptors. Nat. Rev. Neurosci. 2005, 6, 215–229. [Google Scholar] [CrossRef]
- Jechlinger, M.; Pelz, R.; Tretter, V.; Klausberger, T.; Sieghart, W. Subunit Composition and Quantitative Importance of Hetero-Oligomeric Receptors: GABAA receptors containing α6 subunits. J. Neurosci. 1998, 18, 2449–2457. [Google Scholar] [CrossRef]
- Scholze, P.; Pökl, M.; Längle, S.; Steudle, F.; Fabjan, J.; Ernst, M. Two Distinct Populations of A1α6-Containing GABAA-Receptors in Rat Cerebellum. Front. Synaptic Neurosci. 2020, 12, 43. [Google Scholar] [CrossRef]
- Pirker, S.; Schwarzer, C.; Wieselthaler, A.; Sieghart, W.; Sperk, G. GABA(A) Receptors: Immunocytochemical Distribution of 13 Subunits in the Adult Rat Brain. Neuroscience 2000, 101, 815–850. [Google Scholar] [CrossRef]
- Wu, M.; Tian, H.-L.; Liu, X.; Lai, J.H.C.; Du, S.; Xia, J. Impairment of Inhibitory Synapse Formation and Motor Behavior in Mice Lacking the NL2 Binding Partner LHFPL4/GARLH4. Cell Rep. 2018, 23, 1691–1705. [Google Scholar] [CrossRef]
- Varoqueaux, F.; Jamain, S.; Brose, N. Neuroligin 2 Is Exclusively Localized to Inhibitory Synapses. Eur. J. Cell Biol. 2004, 83, 449–456. [Google Scholar] [CrossRef]
- Li, K.W.; Gonzalez-Lozano, M.A.; Koopmans, F.; Smit, A.B. Recent Developments in Data Independent Acquisition (DIA) Mass Spectrometry: Application of Quantitative Analysis of the Brain Proteome. Front. Mol. Neurosci. 2020, 13, 564446. [Google Scholar] [CrossRef]
- Kneussel, M.; Helmut Brandstätter, J.; Gasnier, B.; Feng, G.; Sanes, J.R.; Betz, H. Gephyrin-Independent Clustering of Postsynaptic GABAA Receptor Subtypes. Mol. Cell. Neurosci. 2001, 17, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.-O.; Ip, N.Y. Structural Plasticity of Dendritic Spines: The Underlying Mechanisms and Its Dysregulation in Brain Disorders. Biochim. Biophys. Acta 2013, 1832, 2257–2263. [Google Scholar] [CrossRef] [PubMed]
- Li, K.W.; Hornshaw, M.P.; van der Schors, R.C.; Watson, R.; Tate, S.; Casetta, B.; Jimenez, C.R.; Gouwenberg, Y.; Gundelfinger, E.D.; Smalla, K.-H.; et al. Proteomics Analysis of Rat Brain Postsynaptic Density. Implications of the Diverse Protein Functional Groups for the Integration of Synaptic Physiology. J. Biol. Chem. 2004, 279, 987–1002. [Google Scholar] [CrossRef] [PubMed]
- Pandya, N.J.; Klaassen, R.V.; van der Schors, R.C.; Slotman, J.A.; Houtsmuller, A.; Smit, A.B.; Li, K.W. Group 1 Metabotropic Glutamate Receptors 1 and 5 Form a Protein Complex in Mouse Hippocampus and Cortex. Proteomics 2016, 16, 2698–2705. [Google Scholar] [CrossRef] [PubMed]
- Uezu, A.; Kanak, D.J.; Bradshaw, T.W.A.; Soderblom, E.J.; Catavero, C.M.; Burette, A.C.; Weinberg, R.J.; Soderling, S.H. Identification of an Elaborate Complex Mediating Postsynaptic Inhibition. Science 2016, 353, 1123–1129. [Google Scholar] [CrossRef] [PubMed]
- Music, M.; Soosaipillai, A.; Batruch, I.; Prassas, I.; Bogdanos, D.P.; Diamandis, E.P. A Proteome-Wide Immuno-Mass Spectrometric Identification of Serum Autoantibodies. Clin. Proteom. 2019, 16, 25. [Google Scholar] [CrossRef]
- Bernard, V.; Somogyi, P.; Bolam, J.P. Cellular, Subcellular, and Subsynaptic Distribution of AMPA-Type Glutamate Receptor Subunits in the Neostriatum of the Rat. J. Neurosci. Soc. Neurosci. 1997, 17, 819–833. [Google Scholar] [CrossRef]
- Herrold, A.A.; Persons, A.L.; Napier, T.C. Cellular Distribution of AMPA Receptor Subunits and MGlu5 Following Acute and Repeated Administration of Morphine or Methamphetamine. J. Neurochem. 2013, 126, 503–517. [Google Scholar] [CrossRef]
- Wittig, I.; Braun, H.-P.; Schägger, H. Blue Native PAGE. Nat. Protoc. 2006, 1, 418–428. [Google Scholar] [CrossRef]
- Vincis Pereira Sanglard, L.; Colas des Francs-Small, C. High-Throughput BN-PAGE for Mitochondrial Respiratory Complexes. Methods Mol. Biol. 2022, 2363, 111–119. [Google Scholar] [CrossRef]
- van der Spek, S.J.F.; Pandya, N.J.; Koopmans, F.; Paliukhovich, I.; van der Schors, R.C.; Otten, M.; Smit, A.B.; Li, K.W. Expression and Interaction Proteomics of GluA1- and GluA3-Subunit-Containing AMPARs Reveal Distinct Protein Composition. Cells 2022, 11, 3648. [Google Scholar] [CrossRef] [PubMed]
- Nusser, Z.; Ahmad, Z.; Tretter, V.; Fuchs, K.; Wisden, W.; Sieghart, W.; Somogyi, P. Alterations in the Expression of GABAA Receptor Subunits in Cerebellar Granule Cells after the Disruption of the Alpha6 Subunit Gene. Eur. J. Neurosci. 1999, 11, 1685–1697. [Google Scholar] [CrossRef]
- Nusser, Z.; Sieghart, W.; Somogyi, P. Segregation of Different GABAA Receptors to Synaptic and Extrasynaptic Membranes of Cerebellar Granule Cells. J. Neurosci. 1998, 18, 1693–1703. [Google Scholar] [CrossRef] [PubMed]
- Pöltl, A.; Hauer, B.; Fuchs, K.; Tretter, V.; Sieghart, W. Subunit Composition and Quantitative Importance of GABAA Receptor Subtypes in the Cerebellum of Mouse and Rat. J. Neurochem. 2003, 87, 1444–1455. [Google Scholar] [CrossRef]
- Barnard, E.A.; Skolnick, P.; Olsen, R.W.; Mohler, H.; Sieghart, W.; Biggio, G.; Braestrup, C.; Bateson, A.N.; Langer, S.Z. International Union of Pharmacology. XV. Subtypes of γ-Aminobutyric acidA receptors: Classification on the basis of subunit structure and receptor function. Pharmacol. Rev. 1998, 50, 291–314. [Google Scholar] [PubMed]
- Treven, M.; Siebert, D.C.B.; Holzinger, R.; Bampali, K.; Fabjan, J.; Varagic, Z.; Wimmer, L.; Steudle, F.; Scholze, P.; Schnürch, M.; et al. Towards Functional Selectivity for A6β3γ2 GABA(A) Receptors: A Series of Novel Pyrazoloquinolinones. Br. J. Pharmacol. 2018, 175, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Varagic, Z.; Wimmer, L.; Schnürch, M.; Mihovilovic, M.D.; Huang, S.; Rallapalli, S.; Cook, J.M.; Mirheydari, P.; Ecker, G.F.; Sieghart, W.; et al. Identification of Novel Positive Allosteric Modulators and Null Modulators at the GABAA Receptor A+β- Interface. Br. J. Pharmacol. 2013, 169, 371–383. [Google Scholar] [CrossRef]
- Rudolph, U.; Möhler, H. Analysis of GABAA Receptor Function and Dissection of the Pharmacology of Benzodiazepines and General Anesthetics through Mouse Genetics. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 475–498. [Google Scholar] [CrossRef]
- Chen, Z.-W.; Olsen, R.W. GABAA Receptor Associated Proteins: A Key Factor Regulating GABAA Receptor Function. J. Neurochem. 2007, 100, 279–294. [Google Scholar] [CrossRef]
- Lüscher, B.; Keller, C.A. Regulation of GABAA Receptor Trafficking, Channel Activity, and Functional Plasticity of Inhibitory Synapses. Pharmacol. Ther. 2004, 102, 195–221. [Google Scholar] [CrossRef] [PubMed]
- Poulopoulos, A.; Aramuni, G.; Meyer, G.; Soykan, T.; Hoon, M.; Papadopoulos, T.; Zhang, M.; Paarmann, I.; Fuchs, C.; Harvey, K.; et al. Neuroligin 2 Drives Postsynaptic Assembly at Perisomatic Inhibitory Synapses through Gephyrin and Collybistin. Neuron 2009, 63, 628–642. [Google Scholar] [CrossRef] [PubMed]
- Krueger, D.D.; Tuffy, L.P.; Papadopoulos, T.; Brose, N. The Role of Neurexins and Neuroligins in the Formation, Maturation, and Function of Vertebrate Synapses. Curr. Opin. Neurobiol. 2012, 22, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Tretter, V.; Mukherjee, J.; Maric, H.-M.; Schindelin, H.; Sieghart, W.; Moss, S.J. Gephyrin, the Enigmatic Organizer at GABAergic Synapses. Front. Cell. Neurosci. 2012, 6, 23. [Google Scholar] [CrossRef] [PubMed]
- Kneussel, M.; Brandstätter, J.H.; Laube, B.; Stahl, S.; Müller, U.; Betz, H. Loss of Postsynaptic GABA(A) Receptor Clustering in Gephyrin-Deficient Mice. J. Neurosci. 1999, 19, 9289–9297. [Google Scholar] [CrossRef] [PubMed]
- Lévi, S.; Logan, S.M.; Tovar, K.R.; Craig, A.M. Gephyrin Is Critical for Glycine Receptor Clustering but Not for the Formation of Functional GABAergic Synapses in Hippocampal Neurons. J. Neurosci. 2004, 24, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Fritschy, J.-M.; Harvey, R.J.; Schwarz, G. Gephyrin: Where Do We Stand, Where Do We Go? Trends Neurosci. 2008, 31, 257–264. [Google Scholar] [CrossRef]
- Schägger, H.; Cramer, W.A.; von Jagow, G. Analysis of Molecular Masses and Oligomeric States of Protein Complexes by Blue Native Electrophoresis and Isolation of Membrane Protein Complexes by Two-Dimensional Native Electrophoresis. Anal. Biochem. 1994, 217, 220–230. [Google Scholar] [CrossRef]
- He, E.; Lozano, M.A.G.; Stringer, S.; Watanabe, K.; Sakamoto, K.; den Oudsten, F.; Koopmans, F.; Giamberardino, S.N.; Hammerschlag, A.; Cornelisse, L.N.; et al. MIR137 Schizophrenia-Associated Locus Controls Synaptic Function by Regulating Synaptogenesis, Synapse Maturation and Synaptic Transmission. Hum. Mol. Genet. 2018, 27, 1879–1891. [Google Scholar] [CrossRef]
- Koopmans, F.; Pandya, N.J.; Franke, S.K.; Phillippens, I.H.C.M.H.; Paliukhovich, I.; Li, K.W.; Smit, A.B. Comparative Hippocampal Synaptic Proteomes of Rodents and Primates: Differences in Neuroplasticity-Related Proteins. Front. Mol. Neurosci. 2018, 11, 364. [Google Scholar] [CrossRef]
- Meier, F.; Brunner, A.-D.; Frank, M.; Ha, A.; Bludau, I.; Voytik, E.; Kaspar-Schoenefeld, S.; Lubeck, M.; Raether, O.; Bache, N.; et al. DiaPASEF: Parallel Accumulation–Serial Fragmentation Combined with Data-Independent Acquisition. Nat. Methods 2020, 17, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Mann, M. MaxQuant Enables High Peptide Identification Rates, Individualized p.p.b.-Range Mass Accuracies and Proteome-Wide Protein Quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Demichev, V.; Messner, C.B.; Vernardis, S.I.; Lilley, K.S.; Ralser, M. DIA-NN: Neural Networks and Interference Correction Enable Deep Proteome Coverage in High Throughput. Nat. Methods 2020, 17, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Bolte, S.; Cordelières, F.P. A Guided Tour into Subcellular Colocalization Analysis in Light Microscopy. J. Microsc. 2006, 224, 213–232. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Names | Batch 1 | Batch 2 | |||||
|---|---|---|---|---|---|---|---|
| IPs on Cerebellum Lysate | IPs on Hippocampal Lysate | IPs on Cerebellum Lysate | IPs on Hippocampal Lysate | ||||
| Gabra1 | Gabra6a | Gabra6b | Gabra6b | Gabra6a | Gabra6b | Gabra6b | |
| Gabra1 | 180,790 ± 20,398|100% | 2487 ± 385|8.9% | 18,008 ± 4424|19.1% | 6 ± 12|NA | 6248 ± 1285|6.7% | 11,343 ± 2100|14.4% | 0|NA |
| Gabra6 | 56,914 ± 8010|31.5% | 27,938 ± 1797|100% | 94,089 ± 18,976|100% | 0 ± 0|NA | 92,982 ± 15,344|100% | 79,072 ± 12,676|100% | 0|NA |
| Gabrb2 | 255,366 ± 20,281|141.3% | 25,306 ± 2234|90.6% | 79,766 ± 13,765|84.8% | 0 ± 0|NA | 76,551 ± 15,043|82.2% | 88,527 ± 21,864|111.2% | 0|NA |
| Gabrb3 | 37,074 ± 4005|20.5% | 4748 ± 602|17% | 19,323 ± 5003|20.5% | 0 ± 0|NA | 21,835 ± 4511|23.4% | 18,904 ± 4756|23.7% | 0|NA |
| Gabrd | 31,971 ± 3319|17.7% | 9724 ± 1041|34.8% | 35,276 ± 5229|37.5% | 0 ± 0|NA | 39,517 ± 4543|43.4% | 47,333 ± 1218|60.8% | 0|NA |
| Gabrg2 | 81,516 ± 13,238|45.1% | 1487 ± 492|5.3% | 7641 ± 831|8.1% | 0 ± 0|NA | 7621 ± 456|8.3% | 8403 ± 297|10.8% | 0|NA |
| Gphn | 171 ± 51|0.1% | 7 ± 9|0% | 0 ± 0|0% | 4 ± 8|NA | 113 ± 33|0.1% | 9 ± 16|0% | 7 ± 1|NA |
| Nlgn2 | 58,868 ± 8423|32.6% | 695 ± 132|2.5% | 4571 ± 934|4.9% | 0 ± 0|NA | 234 ± 119|0.3% | 87 ± 79|0.1% | 0|NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, M.; Koopmans, F.; Paliukhovich, I.; van der Spek, S.J.F.; Dong, J.; Smit, A.B.; Li, K.W. Blue Native PAGE–Antibody Shift in Conjunction with Mass Spectrometry to Reveal Protein Subcomplexes: Detection of a Cerebellar α1/α6-Subunits Containing γ-Aminobutyric Acid Type A Receptor Subtype. Int. J. Mol. Sci. 2023, 24, 7632. https://doi.org/10.3390/ijms24087632
Chen M, Koopmans F, Paliukhovich I, van der Spek SJF, Dong J, Smit AB, Li KW. Blue Native PAGE–Antibody Shift in Conjunction with Mass Spectrometry to Reveal Protein Subcomplexes: Detection of a Cerebellar α1/α6-Subunits Containing γ-Aminobutyric Acid Type A Receptor Subtype. International Journal of Molecular Sciences. 2023; 24(8):7632. https://doi.org/10.3390/ijms24087632
Chicago/Turabian StyleChen, Miao, Frank Koopmans, Iryna Paliukhovich, Sophie J. F. van der Spek, Jian Dong, August B. Smit, and Ka Wan Li. 2023. "Blue Native PAGE–Antibody Shift in Conjunction with Mass Spectrometry to Reveal Protein Subcomplexes: Detection of a Cerebellar α1/α6-Subunits Containing γ-Aminobutyric Acid Type A Receptor Subtype" International Journal of Molecular Sciences 24, no. 8: 7632. https://doi.org/10.3390/ijms24087632
APA StyleChen, M., Koopmans, F., Paliukhovich, I., van der Spek, S. J. F., Dong, J., Smit, A. B., & Li, K. W. (2023). Blue Native PAGE–Antibody Shift in Conjunction with Mass Spectrometry to Reveal Protein Subcomplexes: Detection of a Cerebellar α1/α6-Subunits Containing γ-Aminobutyric Acid Type A Receptor Subtype. International Journal of Molecular Sciences, 24(8), 7632. https://doi.org/10.3390/ijms24087632