
Tumor Necrosis Factor Superfamily 14 (LIGHT) Restricts Neovascularization by Decreasing Circulating Endothelial Progenitor Cells and Function

 ,
, 
Abstract
1. Introduction
2. Results
2.1. LIGHT Decreases the Recovery of Capillary Density in C57BL/6 Mice
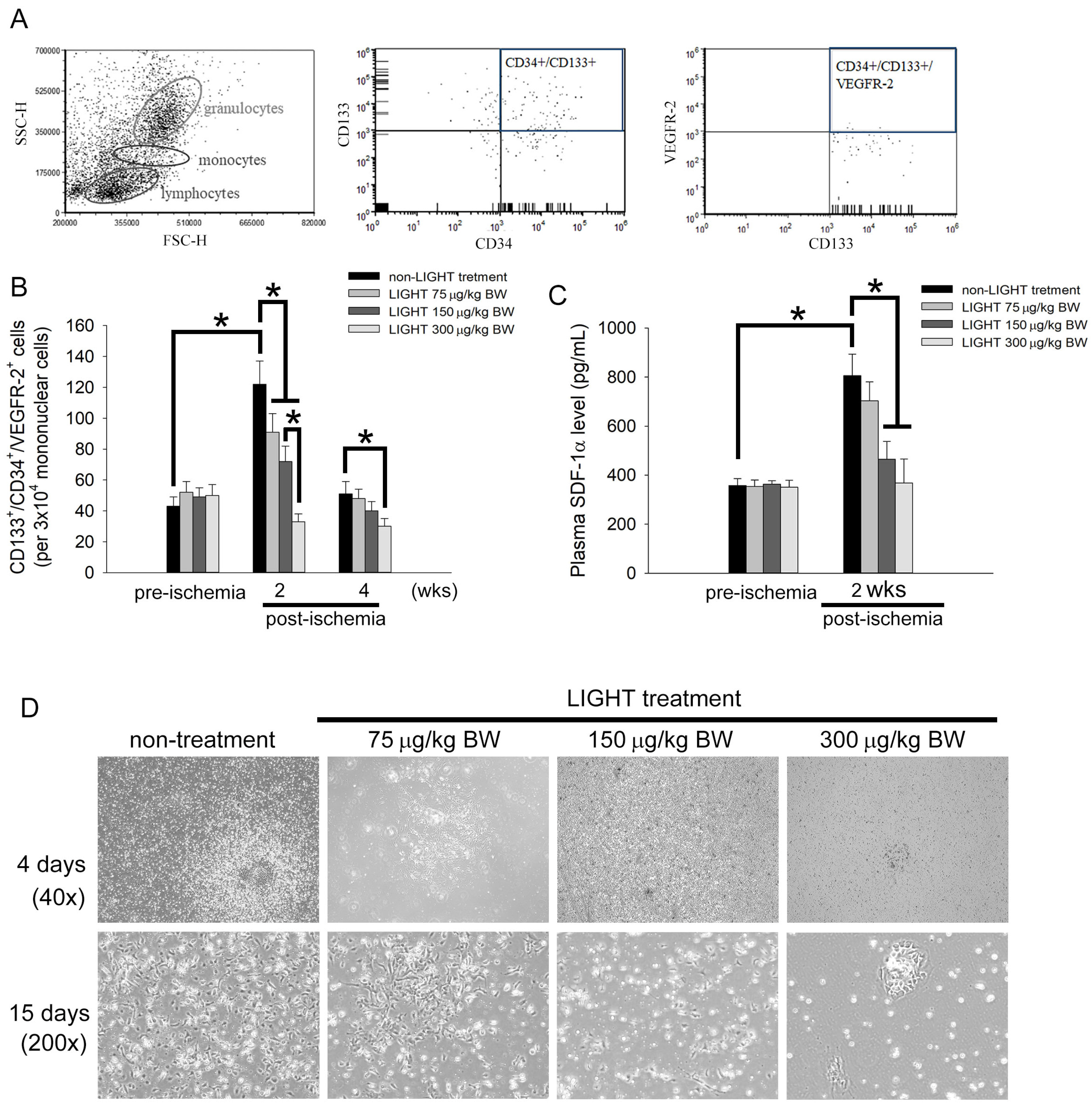
2.2. LIGHT Decreases the Number of Circulating EPCs In Vivo and Inhibits EPCs Differentiation Ex Vivo
2.3. LIGHT Induces Senescence and Mitochondrial Dysfunction in Human EPCs
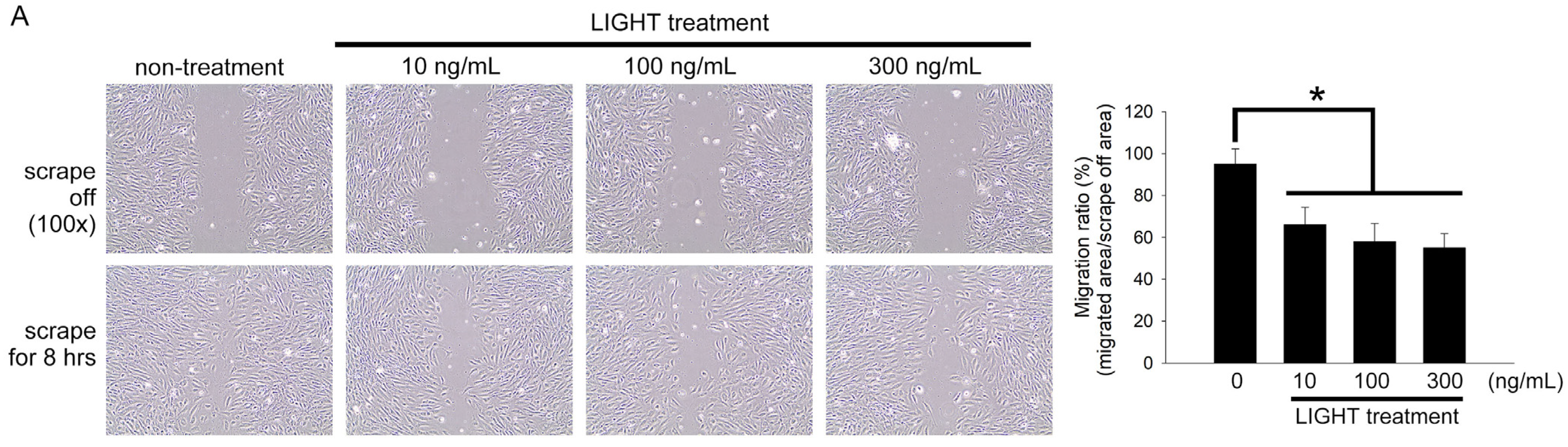
2.4. LIGHT Decreases Neovasculogenesis of Human EPCs
2.5. Akt- and eNOS-Related Pathways May Contribute to the Functional Disruption of Human EPCs by LIGHT
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animal Management and Grouping
4.3. Hind Limb Ischemia Surgery and Doppler Analysis
4.4. Immunohistochemical Staining
4.5. Western Blot Analysis
4.6. Flow Cytometry
4.7. Mouse Early EPCs Colony-Forming Assay
4.8. Isolation and Culture of Human EPCs
4.9. EPCs Senescent Assay
4.10. Succinate Dehydrogenase (SDH) Activity
4.11. Bioenergetic Analysis
4.12. EPC Migration Assay
4.13. EPC Tube Formation Assay
4.14. Real-Time Polymerase Chain Reaction (PCR)
4.15. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cannon, B. Cardiovascular disease: Biochemistry to behaviour. Nature 2013, 493, S2–S3. [Google Scholar] [CrossRef]
- Hage, F.G.; Venkataraman, R.; Zoghbi, G.J.; Perry, G.J.; DeMattos, A.M.; Iskandrian, A.E. The scope of coronary heart disease in patients with chronic kidney disease. J. Am. Coll. Cardiol. 2009, 53, 2129–2140. [Google Scholar] [CrossRef]
- Zhu, M.; Lei, L.; Zhu, Z.; Li, Q.; Guo, D.; Xu, J.; Chen, J.; Sha, H.; Zhang, X.; Yang, X.; et al. Excess TNF-alpha in the blood activates monocytes with the potential to directly form cholesteryl ester-laden cells. Acta Biochim. Biophys. Sin. 2015, 47, 899–907. [Google Scholar] [CrossRef]
- Dostert, C.; Grusdat, M.; Letellier, E.; Brenner, D. The TNF Family of Ligands and Receptors: Communication Modules in the Immune System and Beyond. Physiol. Rev. 2019, 99, 115–160. [Google Scholar] [CrossRef] [PubMed]
- Schneider, K.; Potter, K.G.; Ware, C.F. Lymphotoxin and LIGHT signaling pathways and target genes. Immunol. Rev. 2004, 202, 49–66. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, Y.; Chen, J.; Chen, C. Dual Roles of Tumor Necrosis Factor Superfamily 14 in Antiviral Immunity. Viral. Immunol. 2022, 35, 579–585. [Google Scholar] [CrossRef]
- Brunetti, G.; Belisario, D.C.; Bortolotti, S.; Storlino, G.; Colaianni, G.; Faienza, M.F.; Sanesi, L.; Alliod, V.; Buffoni, L.; Centini, E.; et al. LIGHT/TNFSF14 Promotes Osteolytic Bone Metastases in Non-small Cell Lung Cancer Patients. J. Bone Miner. Res. 2020, 35, 671–680. [Google Scholar] [CrossRef]
- Heo, S.K.; Ju, S.A.; Lee, S.C.; Park, S.M.; Choe, S.Y.; Kwon, B.; Kwon, B.S.; Kim, B.S. LIGHT enhances the bactericidal activity of human monocytes and neutrophils via HVEM. J. Leukoc. Biol. 2006, 79, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Johansson-Percival, A.; Li, Z.J.; Lakhiani, D.D.; He, B.; Wang, X.; Hamzah, J.; Ganss, R. Intratumoral LIGHT Restores Pericyte Contractile Properties and Vessel Integrity. Cell Rep. 2015, 13, 2687–2698. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhang, B.; Li, Y.; Yang, F.; Liu, Y.; Xu, Z.; Wang, G. Dysregulated long non-coding RNAs involved in regulation of matrix degradation during type-B aortic dissection pathogenesis. Gen. Thorac. Cardiovasc. Surg. 2021, 69, 238–245. [Google Scholar] [CrossRef]
- Simonini, A.; Moscucci, M.; Muller, D.W.; Bates, E.R.; Pagani, F.D.; Burdick, M.D.; Strieter, R.M. IL-8 is an angiogenic factor in human coronary atherectomy tissue. Circulation 2000, 101, 1519–1526. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zhang, Y.; Guo, Z.; Wang, M. Cardiovascular Biology of Prostanoids and Drug Discovery. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 1454–1463. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, F.; Finn, A.V.; Yazdani, S.K.; Nakano, M.; Kolodgie, F.D.; Virmani, R. The importance of the endothelium in atherothrombosis and coronary stenting. Nat. Rev. Cardiol. 2012, 9, 439–453. [Google Scholar] [CrossRef] [PubMed]
- Forstermann, U.; Closs, E.I.; Pollock, J.S.; Nakane, M.; Schwarz, P.; Gath, I.; Kleinert, H. Nitric oxide synthase isozymes. Characterization, purification, molecular cloning, and functions. Hypertension 1994, 23, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Aicher, A.; Zeiher, A.M.; Dimmeler, S. Mobilizing endothelial progenitor cells. Hypertension 2005, 45, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Kirton, J.P.; Xu, Q. Endothelial precursors in vascular repair. Microvasc. Res. 2010, 79, 193–199. [Google Scholar] [CrossRef]
- Asahara, T.; Masuda, H.; Takahashi, T.; Kalka, C.; Pastore, C.; Silver, M.; Kearne, M.; Magner, M.; Isner, J.M. Bone marrow origin of endothelial progenitor cells responsible for postnatal vasculogenesis in physiological and pathological neovascularization. Circ. Res. 1999, 85, 221–228. [Google Scholar] [CrossRef]
- Lin, C.P.; Lin, F.Y.; Huang, P.H.; Chen, Y.L.; Chen, W.C.; Chen, H.Y.; Huang, Y.C.; Liao, W.L.; Huang, H.C.; Liu, P.L.; et al. Endothelial progenitor cell dysfunction in cardiovascular diseases: Role of reactive oxygen species and inflammation. Biomed. Res. Int. 2013, 2013, 845037. [Google Scholar] [CrossRef]
- Hill, J.M.; Zalos, G.; Halcox, J.P.; Schenke, W.H.; Waclawiw, M.A.; Quyyumi, A.A.; Finkel, T. Circulating endothelial progenitor cells, vascular function, and cardiovascular risk. N. Engl. J. Med. 2003, 348, 593–600. [Google Scholar] [CrossRef]
- Urbich, C.; Dimmeler, S. Endothelial progenitor cells: Characterization and role in vascular biology. Circ. Res. 2004, 95, 343–353. [Google Scholar] [CrossRef]
- Sandberg, W.J.; Halvorsen, B.; Yndestad, A.; Smith, C.; Otterdal, K.; Brosstad, F.R.; Froland, S.S.; Olofsson, P.S.; Damas, J.K.; Gullestad, L.; et al. Inflammatory interaction between LIGHT and proteinase-activated receptor-2 in endothelial cells: Potential role in atherogenesis. Circ. Res. 2009, 104, 60–68. [Google Scholar] [CrossRef]
- Lee, W.H.; Kim, S.H.; Lee, Y.; Lee, B.B.; Kwon, B.; Song, H.; Kwon, B.S.; Park, J.E. Tumor necrosis factor receptor superfamily 14 is involved in atherogenesis by inducing proinflammatory cytokines and matrix metalloproteinases. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 2004–2010. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.Y.; Tseng, W.K.; Wu, Y.W.; Lin, T.H.; Yeh, H.I.; Chang, K.C.; Wang, J.H.; Chou, R.H.; Huang, C.Y.; Huang, P.H.; et al. Circulating TNFSF14 (Tumor Necrosis Factor Superfamily 14) Predicts Clinical Outcome in Patients With Stable Coronary Artery Disease. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1240–1252. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.Y.; Hsu, C.Y.; Huang, P.H.; Chiang, C.H.; Leu, H.B.; Huang, C.C.; Chen, J.W.; Lin, S.J. Usefulness of Circulating Decoy Receptor 3 in Predicting Coronary Artery Disease Severity and Future Major Adverse Cardiovascular Events in Patients with Multivessel Coronary Artery Disease. Am. J. Cardiol. 2015, 116, 1028–1033. [Google Scholar] [CrossRef]
- Leu, H.B.; Lin, C.P.; Lin, W.T.; Wu, T.C.; Chen, J.W. Risk stratification and prognostic implication of plasma biomarkers in nondiabetic patients with stable coronary artery disease: The role of high-sensitivity C-reactive protein. Chest 2004, 126, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.M.; Danielson, E.; Fonseca, F.A.; Genest, J.; Gotto, A.M., Jr.; Kastelein, J.J.; Koenig, W.; Libby, P.; Lorenzatti, A.J.; MacFadyen, J.G.; et al. Rosuvastatin to prevent vascular events in men and women with elevated C-reactive protein. N. Engl. J. Med. 2008, 359, 2195–2207. [Google Scholar] [CrossRef]
- Park, D.W.; Yun, S.C.; Lee, J.Y.; Kim, W.J.; Kang, S.J.; Lee, S.W.; Kim, Y.H.; Lee, C.W.; Kim, J.J.; Park, S.W.; et al. C-reactive protein and the risk of stent thrombosis and cardiovascular events after drug-eluting stent implantation. Circulation 2009, 120, 1987–1995. [Google Scholar] [CrossRef]
- Rittersma, S.Z.; de Winter, R.J.; Koch, K.T.; Schotborgh, C.E.; Bax, M.; Heyde, G.S.; van Straalen, J.P.; Mulder, K.J.; Tijssen, J.G.; Sanders, G.T.; et al. Preprocedural C-reactive protein is not associated with angiographic restenosis or target lesion revascularization after coronary artery stent placement. Clin. Chem. 2004, 50, 1589–1596. [Google Scholar] [CrossRef]
- Seino, Y.; Ogata, K.; Takano, T.; Ishii, J.; Hishida, H.; Morita, H.; Takeshita, H.; Takagi, Y.; Sugiyama, H.; Tanaka, T.; et al. Use of a whole blood rapid panel test for heart-type fatty acid-binding protein in patients with acute chest pain: Comparison with rapid troponin T and myoglobin tests. Am. J. Med. 2003, 115, 185–190. [Google Scholar] [CrossRef]
- Scholz, H.; Sandberg, W.; Damås, J.K.; Smith, C.; Andreassen, A.K.; Gullestad, L.; Frøland, S.S.; Yndestad, A.; Aukrust, P.; Halvorsen, B. Enhanced plasma levels of LIGHT in unstable angina: Possible pathogenic role in foam cell formation and thrombosis. Circulation 2005, 112, 2121–2129. [Google Scholar] [CrossRef]
- Halvorsen, B.; Santilli, F.; Scholz, H.; Sahraoui, A.; Gulseth, H.L.; Wium, C.; Lattanzio, S.; Formoso, G.; Di Fulvio, P.; Otterdal, K.; et al. LIGHT/TNFSF14 is increased in patients with type 2 diabetes mellitus and promotes islet cell dysfunction and endothelial cell inflammation in vitro. Diabetologia 2016, 59, 2134–2144. [Google Scholar] [CrossRef]
- Otterdal, K.; Smith, C.; Oie, E.; Pedersen, T.M.; Yndestad, A.; Stang, E.; Endresen, K.; Solum, N.O.; Aukrust, P.; Damas, J.K. Platelet-derived LIGHT induces inflammatory responses in endothelial cells and monocytes. Blood 2006, 108, 928–935. [Google Scholar] [CrossRef] [PubMed]
- Lo, J.C.; Wang, Y.; Tumanov, A.V.; Bamji, M.; Yao, Z.; Reardon, C.A.; Getz, G.S.; Fu, Y.X. Lymphotoxin beta receptor-dependent control of lipid homeostasis. Science 2007, 316, 285–288. [Google Scholar] [CrossRef]
- Tamada, K.; Shimozaki, K.; Chapoval, A.I.; Zhu, G.; Sica, G.; Flies, D.; Boone, T.; Hsu, H.; Fu, Y.X.; Nagata, S.; et al. Modulation of T-cell-mediated immunity in tumor and graft-versus-host disease models through the LIGHT co-stimulatory pathway. Nat. Med. 2000, 6, 283–289. [Google Scholar] [CrossRef]
- Tamada, K.; Shimozaki, K.; Chapoval, A.I.; Zhai, Y.; Su, J.; Chen, S.F.; Hsieh, S.L.; Nagata, S.; Ni, J.; Chen, L. LIGHT, a TNF-like molecule, costimulates T cell proliferation and is required for dendritic cell-mediated allogeneic T cell response. J. Immunol. 2000, 164, 4105–4110. [Google Scholar] [CrossRef]
- Zheng, Q.Y.; Cao, Z.H.; Hu, X.B.; Li, G.Q.; Dong, S.F.; Xu, G.L.; Zhang, K.Q. LIGHT/IFN-γ triggers β cells apoptosis via NF-κB/Bcl2-dependent mitochondrial pathway. J. Cell Mol. Med. 2016, 20, 1861–1871. [Google Scholar] [CrossRef]
- Steinberg, M.W.; Shui, J.W.; Ware, C.F.; Kronenberg, M. Regulating the mucosal immune system: The contrasting roles of LIGHT, HVEM, and their various partners. Semin. Immunopathol. 2009, 31, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Šedý, J.; Bekiaris, V.; Ware, C.F. Tumor necrosis factor superfamily in innate immunity and inflammation. Cold Spring Harb. Perspect. Biol. 2014, 7, a016279. [Google Scholar] [CrossRef]
- Riffelmacher, T.; Giles, D.A.; Zahner, S.; Dicker, M.; Andreyev, A.Y.; McArdle, S.; Perez-Jeldres, T.; van der Gracht, E.; Murray, M.P.; Hartmann, N.; et al. Metabolic activation and colitis pathogenesis is prevented by lymphotoxin β receptor expression in neutrophils. Mucosal. Immunol. 2021, 14, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Pessayre, D.; Mansouri, A.; Fromenty, B. Nonalcoholic steatosis and steatohepatitis. V. Mitochondrial dysfunction in steatohepatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 282, G193–G199. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Kaiyala, K.J.; Lam, J.; Agrawal, N.; Nguyen, L.; Ogimoto, K.; Spencer, D.; Morton, G.J.; Schwartz, M.W.; Dichek, H.L. In vivo structure-function studies of human hepatic lipase: The catalytic function rescues the lean phenotype of HL-deficient (hl-/-) mice. Physiol. Rep. 2015, 3, e12365. [Google Scholar] [CrossRef] [PubMed]
- Gonzalvez, F.; D’Aurelio, M.; Boutant, M.; Moustapha, A.; Puech, J.P.; Landes, T.; Arnauné-Pelloquin, L.; Vial, G.; Taleux, N.; Slomianny, C.; et al. Barth syndrome: Cellular compensation of mitochondrial dysfunction and apoptosis inhibition due to changes in cardiolipin remodeling linked to tafazzin (TAZ) gene mutation. Biochim. Biophys. Acta 2013, 1832, 1194–1206. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence |
|---|---|
| integrin β1 | forward: 5′-ctg gtg tgg ttg ctg gaa ttg ttc-3′ |
| reverse: 5′-cct cat act tcg gat tga cca cag-3′ | |
| integrin β3 | forward: 5′-cct gct cat ctg gaa act cct ca-3′ |
| reverse: 5′-cgg tac gtg ata ttg gtg aag gta g-3′ | |
| E-selectin | forward: 5′-ttg gta gct gga ctt tct gct gc-3′ |
| reverse: 5′-gta aga agg ctt ttg gta gct tcc-3′ | |
| GAPDH | forward: 5′-tgc ccc ctc tgc tga tgc c-3′ |
| reverse: 5′-cct ccg acg cct gct tca cca c-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, C.-Y.; Huang, C.-Y.; Shih, C.-M.; Lin, Y.-W.; Huang, P.-H.; Lin, S.-J.; Liu, C.-W.; Lin, C.-Y.; Lin, F.-Y. Tumor Necrosis Factor Superfamily 14 (LIGHT) Restricts Neovascularization by Decreasing Circulating Endothelial Progenitor Cells and Function. Int. J. Mol. Sci. 2023, 24, 6997. https://doi.org/10.3390/ijms24086997
Hsu C-Y, Huang C-Y, Shih C-M, Lin Y-W, Huang P-H, Lin S-J, Liu C-W, Lin C-Y, Lin F-Y. Tumor Necrosis Factor Superfamily 14 (LIGHT) Restricts Neovascularization by Decreasing Circulating Endothelial Progenitor Cells and Function. International Journal of Molecular Sciences. 2023; 24(8):6997. https://doi.org/10.3390/ijms24086997
Chicago/Turabian StyleHsu, Chien-Yi, Chun-Yao Huang, Chun-Ming Shih, Yi-Wen Lin, Po-Hsun Huang, Shing-Jong Lin, Chen-Wei Liu, Cheng-Yen Lin, and Feng-Yen Lin. 2023. "Tumor Necrosis Factor Superfamily 14 (LIGHT) Restricts Neovascularization by Decreasing Circulating Endothelial Progenitor Cells and Function" International Journal of Molecular Sciences 24, no. 8: 6997. https://doi.org/10.3390/ijms24086997
APA StyleHsu, C.-Y., Huang, C.-Y., Shih, C.-M., Lin, Y.-W., Huang, P.-H., Lin, S.-J., Liu, C.-W., Lin, C.-Y., & Lin, F.-Y. (2023). Tumor Necrosis Factor Superfamily 14 (LIGHT) Restricts Neovascularization by Decreasing Circulating Endothelial Progenitor Cells and Function. International Journal of Molecular Sciences, 24(8), 6997. https://doi.org/10.3390/ijms24086997