
Extracellular Vesicles and Their Zeta Potential as Future Markers Associated with Nutrition and Molecular Biomarkers in Breast Cancer


 , , , ,
, , , ,
Abstract
1. Introduction
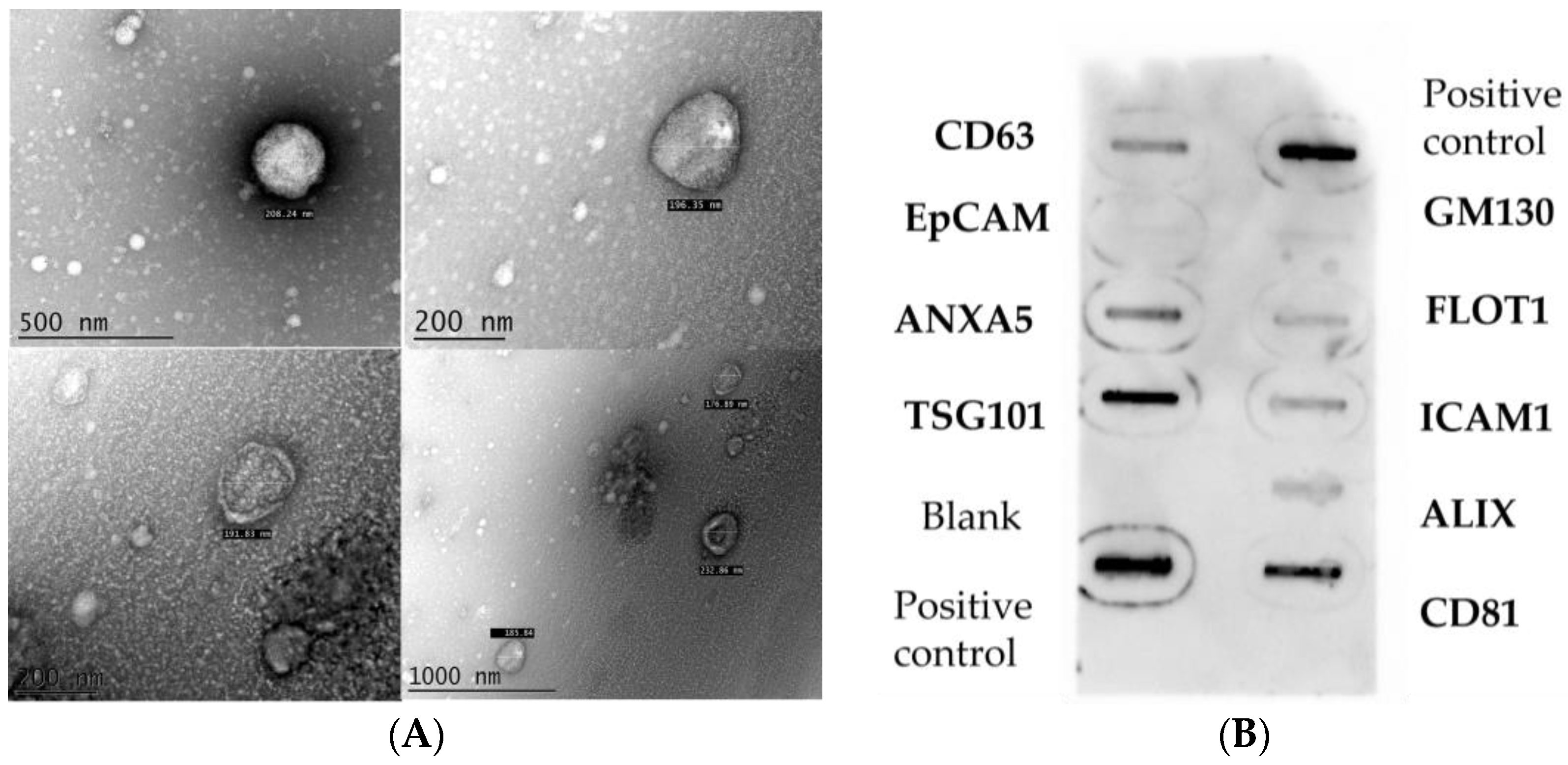
2. Results
3. Discussion
3.1. Different Populations of Extracellular Vesicles
3.2. Associations between the Zeta Potential and Patient Characteristics
3.3. Relevance of miRNA Expression-Containing Plasma EVs in Breast Cancer Patients
3.3.1. Expression of miRNA-145 in EVs
3.3.2. Expression of miRNA-155 in EVs Was Associated with Ki67 and Visceral Fat
3.3.3. miRNA-30 Expression Was Associated with HDL and LDL Cholesterol
4. Materials and Methods
4.1. Subjects
4.2. Isolation and Analysis of Extracellular Vesicles
4.2.1. Isolation of Plasma Extracellular Vesicles
4.2.2. Size and Zeta Potential Analyses
4.2.3. Analysis of miRNA in Extracellular Vesicles
4.2.4. Negative Staining Electron Microscopy Analysis
4.2.5. Specific Exosome Markers
4.2.6. Analysis of Biochemical Biomarkers
4.2.7. Statistical Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Combs, G.F.; Trumbo, P.R.; Mckinley, M.C.; Milner, J.; Studenski, S.; Kimura, T.; Watkins, S.M.; Raiten, D.J. Biomarkers in Nutrition: New Frontiers in Research and Application. Ann. N. Y. Acad. Sci. 2013, 1278, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Raiten, D.J.; Namasté, S.; Brabin, B.; Combs, G.; L’Abbe, M.R.; Wasantwisut, E.; Darnton-Hill, I. Executive Summary--Biomarkers of Nutrition for Development: Building a Consensus. Am. J. Clin. Nutr. 2011, 94, 633S–650S. [Google Scholar] [CrossRef] [PubMed]
- Custódio, I.D.D.; Marinho, E.D.C.; Gontijo, C.A.; Pereira, T.S.S.; Paiva, C.E.; De Maia, Y.C.P. Impact of Chemotherapy on Diet and Nutritional Status of Women with Breast Cancer: A Prospective Study. PLoS ONE 2016, 16, e0157113. [Google Scholar] [CrossRef]
- Agurs-Collins, T.; Ross, S.A.; Dunn, B.K. The Many Faces of Obesity and Its Influence on Breast Cancer Risk. Front. Oncol. 2019, 9, 765. [Google Scholar] [CrossRef]
- Aleixo, G.F.P.; Williams, G.R.; Nyrop, K.A.; Muss, H.B.; Shachar, S.S. Muscle Composition and Outcomes in Patients with Breast Cancer: Meta-Analysis and Systematic Review. Breast Cancer Res. Treat. 2019, 177, 569–579. [Google Scholar] [CrossRef]
- Aleixo, G.F.P.; Deal, A.M.; Nyrop, K.A.; Muss, H.B.; Damone, E.M.; Williams, G.R.; Yu, H.; Shachar, S.S. Association of Body Composition with Function in Women with Early Breast Cancer. Breast Cancer Res. Treat. 2020, 181, 411–421. [Google Scholar] [CrossRef]
- Ethun, C.G.; Bilen, M.A.; Jani, A.B.; Maithel, S.K.; Ogan, K.; Master, V.A. Frailty and Cancer: Implications for Oncology Surgery, Medical Oncology, and Radiation Oncology. CA Cancer J. Clin. 2017, 67, 362–377. [Google Scholar] [CrossRef]
- Guigni, B.A.; Callahan, D.M.; Tourville, T.W.; Miller, M.S.; Fiske, B.; Voigt, T.; Korwin-Mihavics, B.; Anathy, V.; Dittus, K.; Toth, M.J. Skeletal Muscle Atrophy and Dysfunction in Breast Cancer Patients: Role for Chemotherapy-Derived Oxidant Stress. Am. J. Physiol. Cell Physiol. 2018, 315, C744–C756. [Google Scholar] [CrossRef]
- Limon-Miro, A.T.; Valencia, M.E.; Lopez-Teros, V.; Alemán-Mateo, H.; Méndez-Estrada, R.O.; Pacheco-Moreno, B.I.; Astiazaran-Garcia, H. An Individualized Food-Based Nutrition Intervention Reduces Visceral and Total Body Fat While Preserving Skeletal Muscle Mass in Breast Cancer Patients under Antineoplastic Treatment. Clin. Nutr. 2021, 40, 4394–4403. [Google Scholar] [CrossRef]
- Limon-Miro, A.T.; Lopez-Teros, V.; Astiazaran-Garcia, H. Dynamic Macronutrient Meal-Equivalent Menu Method: Towards Individual Nutrition Intervention Programs. Methods Protoc. 2019, 2, 78. [Google Scholar] [CrossRef]
- World Cancer Research Fund/American Institute for Cancer Research. Continuous Update Project Expert Report 2018. Diet, Nutrition, Physical Activity and Breast Cancer. Available online: dietandcancerreport.org (accessed on 1 January 2022).
- Limon-Miro, A.T.; Lopez-Teros, V.; Astiazaran-Garcia, H. Dietary Guidelines for Breast Cancer Patients: A Critical Review. Adv. Nutr. 2017, 8, 613–623. [Google Scholar] [CrossRef]
- Valencia, M.E. Recomendaciones de Nutrimentos para la Poblacion Mexicana; Bourges, H., Casanueva, E., Rosado, J., Eds.; Editorial Médica Panamericana: México, Mexico, 2008. [Google Scholar]
- FDA-NIH Biomarker Working Group. BEST (Biomarkers, EndpointS, and Other Tools) Resource. Available online: https://www.ncbi.nlm.nih.gov/books/NBK326791/ (accessed on 12 December 2021).
- Fougère, B.; Vellas, B.; Van Kan, G.A.; Cesari, M. Identification of Biological Markers for Better Characterization of Older Subjects with Physical Frailty and Sarcopenia. Transl. Neurosci. 2015, 6, 103–110. [Google Scholar] [CrossRef]
- Califf, R.M. Biomarker Definitions and Their Applications. Exp. Biol. Med. 2018, 243, 213–221. [Google Scholar] [CrossRef]
- Mendivil-Alvarado, H.; Sosa-León, L.A.; Carvajal-Millan, E.; Astiazaran-Garcia, H. Malnutrition and Biomarkers: A Journey through Extracellular Vesicles. Nutrients 2022, 14, 1002. [Google Scholar] [CrossRef]
- Oliveira Rodríguez, M.; Serrano Pertierra, E.; García Costa, A.; Soraya López, M.; Yáñez Mo, M.; Cernuda Morollón, E.; Blanco López, M.C. Point of Care Detection of Extracellular Vesicles: Sensitivity Optimization and Multiple Target Detection. Biosens. Bioelectron. 2017, 87, 38–45. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Witwer, K.W.; Goberdhan, D.C.I.; O’Driscoll, L.; Théry, C.; Welsh, J.A.; Blenkiron, C.; Buzás, E.I.; Di Vizio, D.; Erdbrügger, U.; Falcón-Pérez, J.M.; et al. Updating MISEV: Evolving the Minimal Requirements for Studies of Extracellular Vesicles. J. Extracell. Vesicles 2021, 10, e12182. [Google Scholar] [CrossRef]
- Dimassi, S.; Karkeni, E.; Laurant, P.; Tabka, Z.; Landrier, J.F.; Riva, C. Microparticle MiRNAs as Biomarkers of Vascular Function and Inflammation Response to Aerobic Exercise in Obesity? Obesity 2018, 26, 1584–1593. [Google Scholar] [CrossRef]
- Holvoet, P.; Vanhaverbeke, M.; Bloch, K.; Baatsen, P.; Sinnaeve, P.; Janssens, S. Low MT-CO1 in Monocytes and Microvesicles Is Associated with Outcome in Patients with Coronary Artery Disease. J. Am. Heart Assoc. 2016, 5, e004207. [Google Scholar] [CrossRef]
- Hu, W.; Ru, Z.; Zhou, Y.; Xiao, W.; Sun, R.; Zhang, S.; Gao, Y.; Li, X.; Zhang, X.; Yang, H. Lung Cancer-Derived Extracellular Vesicles Induced Myotube Atrophy and Adipocyte Lipolysis via the Extracellular IL-6-Mediated STAT3 Pathway. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2019, 1864, 1091–1102. [Google Scholar] [CrossRef]
- Eitan, E.; Tosti, V.; Suire, C.N.; Cava, E.; Berkowitz, S.; Bertozzi, B.; Raefsky, S.M.; Veronese, N.; Spangler, R.; Spelta, F.; et al. In a Randomized Trial in Prostate Cancer Patients, Dietary Protein Restriction Modifies Markers of Leptin and Insulin Signaling in Plasma Extracellular Vesicles. Aging Cell 2017, 16, 1430–1433. [Google Scholar] [CrossRef] [PubMed]
- Foord, R.; Jakeman, E.; Oliver, C.J.; Pike, E.R.; Blagrove, R.J.; Wood, E.; Peacocke, A.R. Determination of Diffusion Coefficients of Haemocyanin at Low Concentration by Intensity Fluctuation Spectroscopy of Scattered Laser Light. Nature 1970, 227, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Cuadros-Moreno, A.; Casañas Pimentel, R.; San Martín-Martínez, E.; Yañes Fernandez, J. Dispersión de Luz Dinámica En La Determinación de Tamaño de Nanopartículas Poliméricas. Lat.-Am. J. Phys. Educ. 2014, 8, 14. [Google Scholar]
- Martínez, M.C.; Andriantsitohaina, R. Extracellular Vesicles in Metabolic Syndrome. Circ. Res. 2017, 120, 1674–1686. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.C.; Tual-Chalot, S.; Leonetti, D.; Andriantsitohaina, R. Microparticles: Targets and Tools in Cardiovascular Disease. Trends Pharmacol. Sci. 2011, 32, 659–665. [Google Scholar] [CrossRef]
- Lewin, B. Benjamin Lewin; McGraw-Hill: New Yourk, NY, USA, 2002; Volume 16. [Google Scholar]
- Gallo, A.; Tandon, M.; Alevizos, I.; Illei, G.G. The Majority of MicroRNAs Detectable in Serum and Saliva Is Concentrated in Exosomes. PLoS ONE 2012, 7, e30679. [Google Scholar] [CrossRef]
- Huntzinger, E.; Izaurralde, E. Gene Silencing by MicroRNAs: Contributions of Translational Repression and MRNA Decay. Nat. Rev. Genet. 2011, 12, 99–110. [Google Scholar] [CrossRef]
- Zou, C.; Xu, Q.; Mao, F.; Li, D.; Bian, C.; Liu, L.-Z.; Jiang, Y.; Chen, X.; Qi, Y.; Zhang, X.; et al. MiR-145 Inhibits Tumor Angiogenesis and Growth by N-RAS and VEGF. Cell Cycle 2012, 11, 2137–2145. [Google Scholar] [CrossRef]
- Jiang, K.; Yang, J.; Guo, S.; Zhao, G.; Wu, H.; Deng, G. Peripheral Circulating Exosome-Mediated Delivery of MiR-155 as a Novel Mechanism for Acute Lung Inflammation. Mol. Ther. 2019, 27, 1758–1771. [Google Scholar] [CrossRef]
- Katsuda, T.; Kosaka, N.; Ochiya, T. The Roles of Extracellular Vesicles in Cancer Biology: Toward the Development of Novel Cancer Biomarkers. Proteomics 2014, 14, 412–425. [Google Scholar] [CrossRef]
- Han, M.; Wang, Y.; Guo, G.; Li, L.; Dou, D.; Ge, X.; Lv, P.; Wang, F.; Gu, Y. MicroRNA-30d Mediated Breast Cancer Invasion, Migration, and EMT by Targeting KLF11 and Activating STAT3 Pathway. J. Cell Biochem. 2018, 119, 8138–8145. [Google Scholar] [CrossRef]
- Sayyed, A.A.; Gondaliya, P.; Mali, M.; Pawar, A.; Bhat, P.; Khairnar, A.; Arya, N.; Kalia, K. MiR-155 Inhibitor-Laden Exosomes Reverse Resistance to Cisplatin in a 3D Tumor Spheroid and Xenograft Model of Oral Cancer. Mol. Pharm. 2021, 18, 3010–3025. [Google Scholar] [CrossRef]
- Mendivil Alvarado, H.; Chavez Munguia, B.; Carvajal Millan, E.; Hernandez Hernandez, M.; Lopez Teros, V.; Pacheco Moreno, B.; Rascon Duran, L.; Astiazaran Garcia, H. Morphometric Characterization of Extracellular Vesicles in Adults with Obesity. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Grimaldi, A.M.; Nuzzo, S.; Condorelli, G.; Salvatore, M.; Incoronato, M. Prognostic and Clinicopathological Significance of MiR-155 in Breast Cancer: A Systematic Review. Int. J. Mol. Sci. 2020, 21, 5834. [Google Scholar] [CrossRef]
- Simental-Mendía, L.E.; Rodríguez-Morán, M.; Guerrero-Romero, F. The Product of Fasting Glucose and Triglycerides as Surrogate for Identifying Insulin Resistance in Apparently Healthy Subjects. Metab. Syndr. Relat. Disord. 2008, 6, 299–304. [Google Scholar] [CrossRef]
- Hood, J.L.; Wickline, S.A. A Systematic Approach to Exosome-Based Translational Nanomedicine. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2012, 4, 458–467. [Google Scholar] [CrossRef]
- Hofmann, L.; Ludwig, S.; Vahl, J.M.; Brunner, C.; Hoffmann, T.K.; Theodoraki, M.N. The Emerging Role of Exosomes in Diagnosis, Prognosis, and Therapy in Head and Neck Cancer. Int. J. Mol. Sci. 2020, 21, 4072. [Google Scholar] [CrossRef]
- Dini, L.; Tacconi, S.; Carata, E.; Tata, A.M.; Vergallo, C.; Panzarini, E. Microvesicles and Exosomes in Metabolic Diseases and Inflammation. Cytokine Growth Factor Rev. 2020, 51, 27–39. [Google Scholar] [CrossRef]
- Neven, K.Y.; Nawrot, T.S.; Bollati, V. Extracellular Vesicles: How the External and Internal Environment Can Shape Cell-To-Cell Communication. Curr. Environ. Health Rep. 2017, 4, 30–37. [Google Scholar] [CrossRef]
- Turturici, G.; Tinnirello, R.; Sconzo, G.; Geraci, F. Extracellular Membrane Vesicles as a Mechanism of Cell-to-Cell Communication: Advantages and Disadvantages. Am. J. Physiol. Cell Physiol. 2014, 306, C621–C633. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Sun, W.; Zhang, Q.; Gu, T.; Li, G. Plasma Exosomes as Novel Biomarker for the Early Diagnosis of Gastric Cancer. Cancer Biomark. 2018, 21, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, X.; Tao, W.A. Characterization and Applications of Extracellular Vesicle Proteome with Post-Translational Modifications. TrAC Trends Anal. Chem. 2018, 107, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Gündüz, Z.; Dursun, İ.; Tülpar, S.; Baştuğ, F.; Baykan, A.; Yikilmaz, A.; Patıroğlu, T.; Poyrazoglu, H.M.; Akin, L.; Yel, S.; et al. Increased Endothelial Microparticles in Obese and Overweight Children. J. Pediatr. Endocrinol. Metab. 2012, 25, 1111–1117. [Google Scholar] [CrossRef] [PubMed]
- Kakarla, R.; Hur, J.; Kim, Y.J.; Kim, J.; Chwae, Y.-J. Apoptotic Cell-Derived Exosomes: Messages from Dying Cells. Exp. Mol. Med. 2020, 52, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Wang, X.; Liu, X.; Du, H.; Sun, C.; Shao, X.; Tian, J.; Gu, X.; Wang, H.; Tian, J.; et al. Adipose-Derived Exosomes Exert Proatherogenic Effects by Regulating Macrophage Foam Cell Formation and Polarization. J. Am. Heart Assoc. 2018, 7, e007442. [Google Scholar] [CrossRef] [PubMed]
- Gillet, C.; Rasschaert, J. Phenotype of Bone Marrow Adipose Tissue: Specificities of the Anatomical Distribution, Secretory Profile, Lipid Content, and Response to Nutritional Status. Curr. Mol. Biol. Rep. 2018, 4, 8–15. [Google Scholar] [CrossRef]
- Rega-kaun, G.; Ritzel, D.; Kaun, C.; Ebenbauer, B.; Thaler, B.; Prager, M.; Demyanets, S.; Wojta, J.; Hohensinner, P.J. Changes of Circulating Extracellular Vesicles from the Liver after Roux-En-Y Bariatric Surgery. Int. J. Mol. Sci. 2019, 20, 2153. [Google Scholar] [CrossRef]
- Eguchi, A.; Lazic, M.; Armando, A.M.; Phillips, S.A.; Katebian, R.; Maraka, S.; Quehenberger, O.; Sears, D.D.; Feldstein, A.E. Circulating Adipocyte-Derived Extracellular Vesicles Are Novel Markers of Metabolic Stress. J. Mol. Med. 2016, 94, 1241–1253. [Google Scholar] [CrossRef]
- Gao, J.; Li, X.; Wang, Y.; Cao, Y.; Yao, D.; Sun, L.; Qin, L.; Qiu, H.; Zhan, X. Adipocyte-Derived Extracellular Vesicles Modulate Appetite and Weight through MTOR Signalling in the Hypothalamus. Acta Physiol. 2020, 228, 1–16. [Google Scholar] [CrossRef]
- Campello, E.; Zabeo, E.; Radu, C.M.; Spiezia, L.; Foletto, M.; Prevedello, L.; Gavasso, S.; Bulato, C.; Vettor, R.; Simioni, P. Dynamics of Circulating Microparticles in Obesity after Weight Loss. Intern. Emerg. Med. 2016, 11, 695–702. [Google Scholar] [CrossRef]
- Ciardiello, C.; Migliorino, R.; Leone, A.; Budillon, A. Large Extracellular Vesicles: Size Matters in Tumor Progression. Cytokine Growth Factor Rev. 2020, 51, 69–74. [Google Scholar] [CrossRef]
- Melo, S.A.; Sugimoto, H.; O’Connell, J.T.; Kato, N.; Villanueva, A.; Vidal, A.; Qiu, L.; Vitkin, E.; Perelman, L.T.; Melo, C.A.; et al. Cancer Exosomes Perform Cell-Independent MicroRNA Biogenesis and Promote Tumorigenesis. Cancer Cell 2014, 26, 707–721. [Google Scholar] [CrossRef]
- Joyce, D.P.; Kerin, M.J.; Dwyer, R.M. Exosome-Encapsulated MicroRNAs as Circulating Biomarkers for Breast Cancer. Int. J. Cancer 2016, 139, 1443–1448. [Google Scholar] [CrossRef]
- Fuentes, P.; Sesé, M.; Guijarro, P.J.; Emperador, M.; Sánchez-Redondo, S.; Peinado, H.; Hümmer, S.; Ramón y Cajal, S. ITGB3-Mediated Uptake of Small Extracellular Vesicles Facilitates Intercellular Communication in Breast Cancer Cells. Nat. Commun. 2020, 11, 4261. [Google Scholar] [CrossRef]
- Hallal, S.; Tűzesi, Á.; Grau, G.E.; Buckland, M.E.; Alexander, K.L. Understanding the Extracellular Vesicle Surface for Clinical Molecular Biology. J. Extracell. Vesicles 2022, 11, e12260. [Google Scholar] [CrossRef]
- Beit-yannai, E.; Tabak, S.; Stamer, W.D. Physical Exosome: Exosome Interactions. J. Cell. Mol. Med. 2018, 22, 2001–2006. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Rupert, D.L.M.; Claudio, V.; Lässer, C.; Bally, M. Methods for the Physical Characterization and Quantification of Extracellular Vesicles in Biological Samples. Biochim. Biophys. Acta BBA Gen. Subj. 2017, 1861, 3164–3179. [Google Scholar] [CrossRef]
- Woo, J.W.; Chung, Y.R.; Ahn, S.; Kang, E.; Kim, E.K.; Kim, S.H.; Kim, J.H.; Kim, I.A.; Park, S.Y. Changes in Biomarker Status in Metastatic Breast Cancer and Their Prognostic Value. J. Breast Cancer 2019, 22, 439–452. [Google Scholar] [CrossRef]
- Sina, A.A.I.; Vaidyanathan, R.; Dey, S.; Carrascosa, L.G.; Shiddiky, M.J.A.; Trau, M. Real Time and Label Free Profiling of Clinically Relevant Exosomes. Sci. Rep. 2016, 6, 30460. [Google Scholar] [CrossRef]
- Tian, F.; Zhang, S.; Liu, C.; Han, Z.; Liu, Y.; Deng, J.; Li, Y.; Wu, X.; Cai, L.; Qin, L.; et al. Protein Analysis of Extracellular Vesicles to Monitor and Predict Therapeutic Response in Metastatic Breast Cancer. Nat. Commun. 2021, 12, 2536. [Google Scholar] [CrossRef] [PubMed]
- Nanou, A.; Zeune, L.L.; Bidard, F.C.; Pierga, J.Y.; Terstappen, L.W.M.M. HER2 Expression on Tumor-Derived Extracellular Vesicles and Circulating Tumor Cells in Metastatic Breast Cancer. Breast Cancer Res. 2020, 22, 86. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Woo, H.-K.; Lee, C.; Min, Y.; Kumar, S.; Sunkara, V.; Jo, H.-G.; Lee, Y.J.; Kim, J.; Ha, H.K.; et al. EV-Ident: Identifying Tumor-Specific Extracellular Vesicles by Size Fractionation and Single-Vesicle Analysis. Anal. Chem. 2020, 92, 6010–6018. [Google Scholar] [CrossRef] [PubMed]
- Goberdhan, D.C.I. Large Tumour-Derived Extracellular Vesicles as Prognostic Indicators of Metastatic Cancer Patient Survival. Br. J. Cancer 2022, 128, 471–473. [Google Scholar] [CrossRef]
- Botha, J.; Nielsen, M.H.; Christensen, M.H.; Vestergaard, H.; Handberg, A. Bariatric Surgery Reduces CD36-Bearing Microvesicles of Endothelial and Monocyte Origin. Nutr. Metab. 2018, 15, 76. [Google Scholar] [CrossRef]
- Clement, E.; Lazar, I.; Attané, C.; Carrié, L.; Dauvillier, S.; Ducoux-petit, M.; Esteve, D.; Menneteau, T.; Moutahir, M.; Le Gonidec, S.; et al. Adipocyte Extracellular Vesicles Carry Enzymes and Fatty Acids That Stimulate Mitochondrial Metabolism and Remodeling in Tumor Cells. EMBO J. 2020, 39, e102525. [Google Scholar] [CrossRef]
- Jayabalan, N.; Nair, S.; Nuzhat, Z.; Al, E. Cross Talk between Adipose Tissue and Placenta in Obese and Gestational Diabetes Mellitus Pregnancies via Exosomes. Front. Endocrinol. 2017, 8, 239. [Google Scholar] [CrossRef]
- Pardo, F.; Villalobos-Labra, R.; Sobrevia, B.; Toledo, F.; Sobrevia, L. Extracellular Vesicles in Obesity and Diabetes Mellitus. Mol. Asp. Med. 2018, 60, 81–91. [Google Scholar] [CrossRef]
- Barrachina, M.N.; Sueiro, A.M.; Casas, V.; Izquierdo, I.; Hermida-Nogueira, L.; Guitián, E.; Casanueva, F.F.; Abián, J.; Carrascal, M.; Pardo, M.; et al. A Combination of Proteomic Approaches Identifies A Panel of Circulating Extracellular Vesicle Proteins Related to the Risk of Suffering Cardiovascular Disease in Obese Patients. Proteomics 2018, 19, e1800248. [Google Scholar] [CrossRef]
- Matsumura, S.; Minamisawa, T.; Suga, K.; Kishita, H.; Akagi, T.; Ichiki, T.; Ichikawa, Y.; Shiba, K. Subtypes of Tumour Cell-Derived Small Extracellular Vesicles Having Differently Externalized Phosphatidylserine. J. Extracell. Vesicles 2019, 8, 1579541. [Google Scholar] [CrossRef]
- Hood, J.L.; San, R.S.; Wickline, S.A. Exosomes Released by Melanoma Cells Prepare Sentinel Lymph Nodes for Tumor Metastasis. Cancer Res. 2011, 71, 3792–3801. [Google Scholar] [CrossRef]
- Mathivanan, S.; Simpson, R.J. ExoCarta: A Compendium of Exosomal Proteins and RNA. Proteomics 2009, 9, 4997–5000. [Google Scholar] [CrossRef]
- Wubbolts, R.; Leckie, R.S.; Veenhuizen, P.T.M.; Schwarzmann, G.; Möbius, W.; Hoernschemeyer, J.; Slot, J.W.; Geuze, H.J.; Stoorvogel, W. Proteomic and Biochemical Analyses of Human B Cell-Derived Exosomes: Potential Implications for Their Function and Multivesicular Body Formation. J. Biol. Chem. 2003, 278, 10963–10972. [Google Scholar] [CrossRef]
- Surman, M.; Hoja-Łukowicz, D.; Szwed, S.; Drożdż, A.; Stępień, E.; Przybyło, M. Human Melanoma-Derived Ectosomes Are Enriched with Specific Glycan Epitopes. Life Sci. 2018, 207, 395–411. [Google Scholar] [CrossRef]
- Surman, M.; Stępień, E.; Hoja-Łukowicz, D.; Przybyło, M. Deciphering the Role of Ectosomes in Cancer Development and Progression: Focus on the Proteome. Clin. Exp. Metastasis 2017, 34, 273–289. [Google Scholar] [CrossRef]
- Kalra, H.; Drummen, G.P.C.; Mathivanan, S. Focus on Extracellular Vesicles: Introducing the next Small Big Thing. Int. J. Mol. Sci. 2016, 17, 170. [Google Scholar] [CrossRef]
- Eguchi, A.; Mulya, A.; Lazic, M.; Radhakrishnan, D.; Berk, M.P. Microparticles Release by Adipocytes Act as “ Find-Me ” Signals to Promote Macrophage Migration. PLoS ONE 2015, 10, e0123110. [Google Scholar] [CrossRef]
- Akagi, T.; Kato, K.; Kobayashi, M.; Kosaka, N.; Ochiya, T.; Ichiki, T. On-Chip Immunoelectrophoresis of Extracellular Vesicles Released from Human Breast Cancer Cells. PLoS ONE 2015, 10, e0123603. [Google Scholar] [CrossRef]
- Martins, Á.M.; Ramos, C.C.; Freitas, D.; Reis, C.A. Glycosylation of Cancer Extracellular Vesicles: Capture Strategies, Functional Roles and Potential Clinical Applications. Cells 2021, 10, 109. [Google Scholar] [CrossRef]
- Macedo-da-Silva, J.; Santiago, V.F.; Rosa-Fernandes, L.; Marinho, C.R.F.; Palmisano, G. Protein Glycosylation in Extracellular Vesicles: Structural Characterization and Biological Functions. Mol. Immunol. 2021, 135, 226–246. [Google Scholar] [CrossRef]
- Menck, K.; Scharf, C.; Bleckmann, A.; Dyck, L.; Rost, U.; Wenzel, D.; Dhople, V.M.; Siam, L.; Pukrop, T.; Binder, C.; et al. Microvesicles Mediate Breast Cancer Invasion through Glycosylated EMMPRIN. J. Mol. Cell Biol. 2015, 7, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Nishida-Aoki, N.; Tominaga, N.; Kosaka, N.; Ochiya, T. Altered Biodistribution of Deglycosylated Extracellular Vesicles through Enhanced Cellular Uptake. J. Extracell. Vesicles 2020, 9, 1713527. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Cao, L.; Wu, Y.; Wang, B.; Song, Z.; Yang, J.; Cheng, L.; Yang, X.; Zhou, X.; Dai, Z.; et al. Bisecting GlcNAc Modification Diminishes the Pro-Metastatic Functions of Small Extracellular Vesicles from Breast Cancer Cells. J. Extracell. Vesicles 2020, 10, e12005. [Google Scholar] [CrossRef] [PubMed]
- Temoche-Diaz, M.M.; Shurtleff, M.J.; Nottingham, R.M.; Yao, J.; Fadadu, R.P.; Lambowitz, A.M.; Schekman, R. Distinct Mechanisms of MicroRNA Sorting into Cancer Cell-Derived Extracellular Vesicle Subtypes. Elife 2019, 8, e47544. [Google Scholar] [CrossRef] [PubMed]
- Groot, M.; Lee, H. Sorting Mechanisms for MicroRNAs into Extracellular Vesicles and Their Associated Diseases. Cells 2020, 9, 1044. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Chen, Q.; Wang, W.; Ling, Y.; Yan, Y.; Xia, P. Hepatocyte-Derived Extracellular Vesicles Promote Endothelial Inflammation and Atherogenesis via MicroRNA-1. J. Hepatol. 2020, 72, 156–166. [Google Scholar] [CrossRef]
- Shehzad, A.; Islam, S.U.; Shahzad, R.; Khan, S.; Lee, Y.S. Extracellular Vesicles in Cancer Diagnostics and Therapeutics. Pharmacol. Ther. 2021, 223, 107806. [Google Scholar] [CrossRef]
- Ozawa, P.M.M.; Vieira, E.; Lemos, D.S.; Souza, I.L.M.; Zanata, S.M.; Pankievicz, V.C.; Tuleski, T.R.; Souza, E.M.; Wowk, P.F.; Urban, C.D.A.; et al. Identification of MiRNAs Enriched in Extracellular Vesicles Derived from Serum Samples of Breast Cancer Patients. Biomolecules 2020, 10, 150. [Google Scholar] [CrossRef]
- Kang, M.H.; Zhang, L.H.; Wijesekara, N.; De Haan, W.; Butland, S.; Bhattacharjee, A.; Hayden, M.R. Regulation of ABCA1 Protein Expression and Function in Hepatic and Pancreatic Islet Cells by MiR-145. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2724–2732. [Google Scholar] [CrossRef]
- Sheykhhasan, M.; Kalhor, N.; Sheikholeslami, A.; Dolati, M.; Amini, E.; Fazaeli, H. Exosomes of Mesenchymal Stem Cells as a Proper Vehicle for Transfecting MiR-145 into the Breast Cancer Cell Line and Its Effect on Metastasis. Biomed Res. Int. 2021, 2021, 5516078. [Google Scholar] [CrossRef]
- Zhang, P.; Lim, S.; Jiang, K.; Chew, T.; Low, B.; Lim, C. Distinct MRNAs in Cancer Extracellular Vesicles Activate Angiogenesis and Alter Transcriptome of Vascular Endothelial Cells. Cancers 2021, 13, 2009. [Google Scholar] [CrossRef]
- Pan, S.; Zhao, X.; Shao, C.; Fu, B.; Huang, Y.; Zhang, N.; Dou, X.; Zhang, Z.; Qiu, Y.; Wang, R.; et al. STIM1 Promotes Angiogenesis by Reducing Exosomal MiR-145 in Breast Cancer MDA-MB-231 Cells. Cell Death Dis. 2021, 12, 38. [Google Scholar] [CrossRef]
- Dong, M.; Xu, T.; Li, H.; Li, X. LINC00052 Promotes Breast Cancer Cell Progression and Metastasis by Sponging MiR-145-5p to Modulate TGFBR2 Expression. Oncol. Lett. 2021, 21, 368. [Google Scholar] [CrossRef]
- Wang, Y.; Dan, L.; Li, Q.; Li, L.; Zhong, L.; Shao, B.; Yu, F.; He, S.; Tian, S.; He, J.; et al. ZMYND10, an Epigenetically Regulated Tumor Suppressor, Exerts Tumor-Suppressive Functions via MiR145-5p/NEDD9 Axis in Breast Cancer. Clin. Epigenetics 2019, 11, 184. [Google Scholar] [CrossRef]
- Hutcheson, R.; Terry, R.; Chaplin, J.; Smith, E.; Musiyenko, A.; Russell, J.C.; Lincoln, T.; Rocic, P. MicroRNA-145 Restores Contractile Vascular Smooth Muscle Phenotype and Coronary Collateral Growth in the Metabolic Syndrome. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 727–736. [Google Scholar] [CrossRef]
- Greenlee, H.; Iribarren, C.; Rana, J.S.; Cheng, R.; Nguyen-Huynh, M.; Rillamas-Sun, E.; Shi, Z.; Laurent, C.A.; Lee, V.S.; Roh, J.M.; et al. Risk of Cardiovascular Disease in Women With and Without Breast Cancer: The Pathways Heart Study. J. Clin. Oncol. 2022, 40, 1647–1658. [Google Scholar] [CrossRef]
- Bradshaw, P.T.; Stevens, J.; Khankari, N.; Teitelbaum, S.L.; Neugut, A.I.; Gammon, M.D. Cardiovascular Disease Mortality Among Breast Cancer Survivors. Epidemiology 2016, 27, 6–13. [Google Scholar] [CrossRef]
- Cercato, C.; Fonseca, F.A. Cardiovascular Risk and Obesity. Diabetol. Metab. Syndr. 2019, 11, 74. [Google Scholar] [CrossRef]
- Mehta, L.S.; Watson, K.E.; Barac, A.; Beckie, T.M.; Bittner, V.; Cruz-Flores, S.; Dent, S.; Kondapalli, L.; Ky, B.; Okwuosa, T.; et al. Cardiovascular Disease and Breast Cancer: Where These Entities Intersect: A Scientific Statement From the American Heart Association. Circulation 2018, 137, e30–e66. [Google Scholar] [CrossRef]
- Hang, W.; Feng, Y.; Sang, Z.; Yang, Y.; Zhu, Y.; Huang, Q.; Xi, X. Downregulation of MiR-145-5p in Cancer Cells and Their Derived Exosomes May Contribute to the Development of Ovarian Cancer by Targeting CT. Int. J. Mol. Med. 2019, 43, 256–266. [Google Scholar] [CrossRef]
- Kim, S.; Choi, M.C.; Jeong, J.Y.; Hwang, S.; Jung, S.G.; Joo, W.D.; Park, H.; Song, S.H.; Lee, C.; Kim, T.H.; et al. Serum Exosomal MiRNA-145 and MiRNA-200c as Promising Biomarkers for Preoperative Diagnosis of Ovarian Carcinomas. J. Cancer 2019, 10, 1958–1967. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Qin, S.; An, T.; Tang, Y.; Huang, Y.; Zheng, L. MiR-145 Detection in Urinary Extracellular Vesicles Increase Diagnostic Efficiency of Prostate Cancer Based on Hydrostatic Filtration Dialysis Method. Prostate 2017, 77, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Tianyuan, S.; Minzhi, Z.; We, L.; Lin, Z.; Jianping, W.; Mouming, Z. Tripeptide Leu-Ser-Trp Regulates the Vascular Endothelial Cells Phenotype Switching by Mediating the Vascular Smooth Muscle Cells-Derived Small Extracellular Vesicles Packaging of MiR-145. Molecules 2022, 27, 7025. [Google Scholar] [CrossRef]
- Shinohara, H.; Kuranaga, Y.; Kumazaki, M.; Sugito, N.; Yoshikawa, Y.; Takai, T.; Taniguchi, K.; Ito, Y.; Akao, Y. Regulated Polarization of Tumor-Associated Macrophages by MiR-145 via Colorectal Cancer-Derived Extracellular Vesicles. J. Immunol. 2017, 199, 1505–1515. [Google Scholar] [CrossRef]
- Gai, X.; Tang, B.; Liu, F.; Wu, Y.; Wang, F.; Jing, Y.; Huang, F.; Jin, D.; Wang, L.; Zhang, H. MTOR/MiR-145-Regulated Exosomal GOLM1 Promotes Hepatocellular Carcinoma through Augmented GSK-3β/MMPs. J. Genet. Genom. 2019, 46, 235–245. [Google Scholar] [CrossRef]
- Zhou, H.; Zhou, J.; Teng, H.; Yang, H.; Qiu, J.; Li, X. MiR-145 Enriched Exosomes Derived from Bone Marrow-Derived Mesenchymal Stem Cells Protects against Cerebral Ischemia-Reperfusion Injury through Downregulation of FOXO1. Biochem. Biophys. Res. Commun. 2022, 632, 92–99. [Google Scholar] [CrossRef]
- Cho, K.A.; Choi, D.W.; Kim, Y.H.; Kim, J.; Ryu, K.H.; Woo, S.Y. Mesenchymal Stem Cell-Derived Exosomes Protect Muscle Loss by MiR-145-5p Activity Targeting Activin A Receptors. Cells 2021, 10, 2169. [Google Scholar] [CrossRef]
- Zhao, C.; Chen, J.Y.; Peng, W.M.; Yuan, B.; Bi, Q.; Xu, Y.J. Exosomes from Adipose-Derived Stem Cells Promote Chondrogenesis and Suppress Inflammation by Upregulating MiR-145 and MiR-221. Mol. Med. Rep. 2020, 21, 1881. [Google Scholar] [CrossRef]
- Ding, Y.; Cao, F.; Sun, H.; Wang, Y.; Liu, S.; Wu, Y.; Cui, Q.; Mei, W.T.; Li, F. Exosomes Derived from Human Umbilical Cord Mesenchymal Stromal Cells Deliver Exogenous MiR-145-5p to Inhibit Pancreatic Ductal Adenocarcinoma Progression. Cancer Lett. 2019, 442, 351–361. [Google Scholar] [CrossRef]
- Zhang, F.; Isak, A.N.; Yang, S.; Song, Y.; Ren, L.; Feng, C.; Chen, G. Smartly Responsive DNA–MiRNA Hybrids Packaged in Exosomes for Synergistic Enhancement of Cancer Cell Apoptosis. Nanoscale 2022, 14, 6612–6619. [Google Scholar] [CrossRef]
- Gonzalez-Villasana, V.; Rashed, M.H.; Gonzalez-Cantú, Y.; Bayraktar, R.; Luis Menchaca-Arredondo, J.; Manuel Vazquez-Guillen, J.; Rodriguez-Padilla, C.; Lopez-Berestein, G.; Resendez-Perez, D. Research Article Presence of Circulating MiR-145, MiR-155. Dis. Markers 2019, 2019, 6852917. [Google Scholar] [CrossRef]
- Hemmatzadeh, M.; Mohammadi, H.; Jadidi-Niaragh, F.; Asghari, F.; Yousefi, M. The Role of Oncomirs in the Pathogenesis and Treatment of Breast Cancer. Biomed. Pharmacother. 2016, 78, 129–139. [Google Scholar] [CrossRef]
- Kia, V.; Paryan, M.; Mortazavi, Y.; Biglari, A.; Mohammadi-Yeganeh, S. Evaluation of Exosomal MiR-9 and MiR-155 Targeting PTEN and DUSP14 in Highly Metastatic Breast Cancer and Their Effect on Low Metastatic Cells. J. Cell Biochem. 2019, 120, 5666–5676. [Google Scholar] [CrossRef]
- Conley-LaComb, M.K.; Saliganan, A.; Kandagatla, P.; Chen, Y.Q.; Cher, M.L.; Chinni, S.R. PTEN Loss Mediated Akt Activation Promotes Prostate Tumor Growth and Metastasis via CXCL12/CXCR4 Signaling. Mol. Cancer 2013, 12, 85. [Google Scholar] [CrossRef]
- Santos, J.C.; Lima, N.D.S.; Sarian, L.O.; Matheu, A.; Ribeiro, M.L.; Derchain, S.F.M. Exosome-Mediated Breast Cancer Chemoresistance via MiR-155 Transfer. Sci. Rep. 2018, 8, 829. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, L.; Yu, G.; Sun, Z.; Wang, T.; Tian, X.; Duan, X.; Zhang, C. Exosomal MiR-1246 and MiR-155 as Predictive and Prognostic Biomarkers for Trastuzumab-Based Therapy Resistance in HER2-Positive Breast Cancer. Cancer Chemother. Pharmacol. 2020, 86, 761–772. [Google Scholar] [CrossRef]
- Wu, Q.; Sun, S.; Li, Z.; Yang, Q.; Li, B.; Zhu, S.; Wang, L.; Wu, J.; Yuan, J.; Yang, C.; et al. Tumour-Originated Exosomal MiR-155 Triggers Cancer-Associated Cachexia to Promote Tumour Progression 11 Medical and Health Sciences 1112 Oncology and Carcinogenesis 06 Biological Sciences 0601 Biochemistry and Cell Biology. Mol. Cancer 2018, 17, 155. [Google Scholar] [CrossRef]
- Sharma, S.; Opyrchal, M.; Lu, X. Harnessing Tumorous Flaws for Immune Supremacy: Is MiRNA-155 the Weak Link in Breast Cancer Progression? J. Clin. Investig. 2022, 132, e163010. [Google Scholar] [CrossRef]
- Meng, L.; Xing, Z.; Guo, Z.; Qiu, Y.; Liu, Z. Hypoxia-Induced MicroRNA-155 Overexpression in Extracellular Vesicles Promotes Renal Cell Carcinoma Progression by Targeting FOXO3. Aging 2021, 13, 9613. [Google Scholar] [CrossRef]
- Yang, Z.; Shi, X.; Gao, Z.; Chu, L. MiR-155-5p in Extracellular Vesicles Derived from Choroid Plexus Epithelial Cells Promotes Autophagy and Inflammation to Aggravate Ischemic Brain Injury in Mice. Oxidative Med. Cell. Longev. 2022, 2022, 8603427. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, G.; Ding, Y.-Y.; Zhang, Y.-X. Expression of MiR-155 in Serum Exosomes in Children with Epilepsy and Its Diagnostic Value. Dis. Markers 2022, 2022, 7979500. [Google Scholar] [CrossRef] [PubMed]
- Stevic, I.; Buescher, G.; Ricklefs, F.L. Monitoring Therapy Efficiency in Cancer through Extracellular Vesicles. Cells 2020, 9, 130. [Google Scholar] [CrossRef] [PubMed]
- Silveira, E.A.; Da Silva Filho, R.R.; Claudia, M.; Spexoto, B.; Haghighatdoost, F.; Sarrafzadegan, N.; De Oliveira, C. Molecular Sciences The Role of Sarcopenic Obesity in Cancer and Cardiovascular Disease: A Synthesis of the Evidence on Pathophysiological Aspects and Clinical Implications. Int. J. Mol. Sci. 2021, 22, 4339. [Google Scholar] [CrossRef] [PubMed]
- Bulut, G.; Atci, N. Sarcopenic Obesity in Early Breast Cancer Patients with Metabolic Syndrome: A Cross-Sectional Study. Futur. Oncol. 2022, 18, 2489–2498. [Google Scholar] [CrossRef]
- Sun, S.; Wu, Q.; Li, J.; Li, Z.; Sun, S.; Zhu, S.; Wang, L.; Wu, J.; Yuan, J.; Zhang, Y.; et al. Exosomes from the Tumour-Adipocyte Interplay Stimulate Beige/Brown Differentiation and Reprogram Metabolism in Stromal Adipocytes to Promote Tumour Progression. J. Exp. Clin. Cancer Res. 2019, 38, 223. [Google Scholar] [CrossRef]
- Uddin, M.S.; Kabir, M.T.; Jakaria, M.; Mamun, A.A.; Niaz, K.; Amran, M.S.; Barreto, G.E.; Ashraf, G.M. Endothelial PPARγ Is Crucial for Averting Age-Related Vascular Dysfunction by Stalling Oxidative Stress and ROCK. Neurotox. Res. 2019, 36, 583–601. [Google Scholar] [CrossRef]
- Verma, S.; Szmitko, P.E. The Vascular Biology of Peroxisome Proliferator-Activated Receptors: Modulation of Atherosclerosis. Can. J. Cardiol. 2006, 22, 12B–17B. [Google Scholar] [CrossRef]
- Zheng, B.; Yin, W.-N.; Suzuki, T.; Zhang, X.-H.; Zhang, Y.; Song, L.-L.; Jin, L.-S.; Zhan, H.; Zhang, H.; Li, J.-S.; et al. Exosome-Mediated MiR-155 Transfer from Smooth Muscle Cells to Endothelial Cells Induces Endothelial Injury and Promotes Atherosclerosis. Mol. Ther. 2017, 25, 1279–1294. [Google Scholar] [CrossRef]
- Fitzsimons, S.; Oggero, S.; Bruen, R.; McCarthy, C.; Strowitzki, M.J.; Mahon, N.G.; Ryan, N.; Brennan, E.P.; Barry, M.; Perretti, M.; et al. MicroRNA-155 Is Decreased During Atherosclerosis Regression and Is Increased in Urinary Extracellular Vesicles During Atherosclerosis Progression. Front. Immunol. 2020, 11, 576516. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, C.; Liu, L.; Xi, A.; Chen, B.; Li, Y.; Du, J. Macrophage-Derived Mir-155-Containing Exosomes Suppress Fibroblast Proliferation and Promote Fibroblast Inflammation during Cardiac Injury. Mol. Ther. 2017, 25, 192–204. [Google Scholar] [CrossRef]
- Roberts, T.C.; Blomberg, K.E.M.; McClorey, G.; EL Andaloussi, S.; Godfrey, C.; Betts, C.; Coursindel, T.; Gait, M.J.; Smith, C.E.; Wood, M.J. Expression Analysis in Multiple Muscle Groups and Serum Reveals Complexity in the MicroRNA Transcriptome of the Mdx Mouse with Implications for Therapy. Mol. Ther.-Nucleic Acids 2012, 1, e39. [Google Scholar] [CrossRef]
- Guess, M.G.; Barthel, K.K.B.; Harrison, B.C.; Leinwand, L.A. MiR-30 Family MicroRNAs Regulate Myogenic Differentiation and Provide Negative Feedback on the MicroRNA Pathway. PLoS ONE 2015, 10, e0118229. [Google Scholar] [CrossRef]
- Veitch, S.; Njock, M.-S.; Chandy, M.; Siraj, M.A.; Chi, L.; Mak, H.; Yu, K.; Rathnakumar, K.; Perez-Romero, C.A.; Chen, Z.; et al. MiR-30 Promotes Fatty Acid Beta-Oxidation and Endothelial Cell Dysfunction and Is a Circulating Biomarker of Coronary Microvascular Dysfunction in Pre-Clinical Models of Diabetes. Cardiovasc. Diabetol. 2022, 21, 31. [Google Scholar] [CrossRef]
- Zhang, S.; Guo, L.J.; Zhang, G.; Wang, L.L.; Hao, S.; Gao, B.; Jiang, Y.; Tian, W.G.; Cao, X.E.; Luo, D.L. Roles of MicroRNA-124a and MicroRNA-30d in Breast Cancer Patients with Type 2 Diabetes Mellitus. Tumor Biol. 2016, 37, 11057–11063. [Google Scholar] [CrossRef]
- Huang, J.P.; Chang, C.C.; Kuo, C.Y.; Huang, K.J.; Sokal, E.M.; Chen, K.H.; Hung, L.M. Exosomal MicroRNAs MiR-30d-5p and MiR-126a-5p Are Associated with Heart Failure with Preserved Ejection Fraction in STZ-Induced Type 1 Diabetic Rats. Int. J. Mol. Sci. 2022, 23, 7514. [Google Scholar] [CrossRef]
- Fullerton, J.L.; Cosgrove, C.C.; Rooney, R.A.; Work, L.M. Extracellular Vesicles and Their MicroRNA Cargo in Ischaemic Stroke. J. Physiol. 2022. [Google Scholar] [CrossRef]
- Li, J.; Salvador, A.M.; Li, G.; Valkov, N.; Ziegler, O.; Yeri, A.; Xiao, C.Y.; Meechoovet, B.; Alsop, E.; Rodosthenous, R.S.; et al. Mir-30d Regulates Cardiac Remodeling by Intracellular and Paracrine Signaling. Circ. Res. 2021, 128, e1–e23. [Google Scholar] [CrossRef]
- Yin, H.; Wang, Y.; Wu, Y.; Zhang, X.; Zhang, X.; Liu, J.; Wang, T.; Fan, J.; Sun, J.; Yang, A.; et al. EZH2-Mediated Epigenetic Silencing of MiR-29/MiR-30 Targets LOXL4 and Contributes to Tumorigenesis, Metastasis, and Immune Microenvironment Remodeling in Breast Cancer. Theranostics 2020, 10, 8494. [Google Scholar] [CrossRef]
- Jamshidi, M.; Fagerholm, R.; Muranen, T.A.; Kaur, S.; Potdar, S.; Khan, S.; Netti, E.; Mpindi, J.P.; Yadav, B.; Kiiski, J.I.; et al. High MiR-30 Expression Associates with Improved Breast Cancer Patient Survival and Treatment Outcome. Cancers 2021, 13, 2907. [Google Scholar] [CrossRef]
- Elhelbawy, N.G.; Zaid, I.F.; Khalifa, A.A.; Gohar, S.F.; Fouda, E.A. MiRNA-148a and MiRNA-30c Expressions as Potential Biomarkers in Breast Cancer Patients. Biochem. Biophys. Rep. 2021, 27, 101060. [Google Scholar] [CrossRef]
- Bao, S.; Wang, X.; Wang, Z.; Yang, J.; Liu, F.; Yin, C. MicroRNA-30 Mediates Cell Invasion and Metastasis in Breast Cancer. Biochem. Cell Biol. 2018, 96, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Croset, M.; Pantano, F.; Kan, C.W.S.; Bonnelye, E.; Descotes, F.; Alix-Panabieres, C.; Lecellier, C.H.; Bachelier, R.; Allioli, N.; Hong, S.S.; et al. MiRNA-30 Family Members Inhibit Breast Cancer Invasion, Osteomimicry, and Bone Destruction by Directly Targeting Multiple Bone Metastasis-Associated Genes. Cancer Res. 2018, 78, 5259–5273. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.J.; Yang, S.Y.; Wang, D.D.; Chen, X.; Shen, H.Y.; Zhang, X.H.; Zhong, S.L.; Tang, J.H.; Zhao, J.H. The MiR-30 Family: Versatile Players in Breast Cancer. Tumor Biol. 2017, 39, 1010428317692204. [Google Scholar] [CrossRef] [PubMed]
- Kunc, M.; Popęda, M.; Szałkowska, A.; Niemira, M.; Bieńkowski, M.; Pęksa, R.; Łacko, A.; Radecka, B.S.; Braun, M.; Pikiel, J.; et al. MicroRNA Expression Profile in Single Hormone Receptor-Positive Breast Cancers Is Mainly Dependent on HER2 Status—A Pilot Study. Diagnostics 2020, 10, 617. [Google Scholar] [CrossRef]
- Council for International Organizations of Medical Sciences. International Ethical Guidelines for Health-Related Research Involving Humans; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Chen, C.; Ridzon, D.A.; Broomer, A.J.; Zhou, Z.; Lee, D.H.; Nguyen, J.T.; Barbisin, M.; Xu, N.L.; Mahuvakar, V.R.; Andersen, M.R.; et al. Real-Time Quantification of MicroRNAs by Stem-Loop RT-PCR. Nucleic Acids Res. 2005, 33, e179. [Google Scholar] [CrossRef]
- Kramer, M.F. Stem-Loop RT-QPCR for MiRNAs. Curr. Protoc. Mol. Biol. 2011, 95, 1–15. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Taylor, S.C.; Nadeau, K.; Abbasi, M.; Lachance, C.; Nguyen, M.; Fenrich, J. The Ultimate QPCR Experiment: Producing Publication Quality, Reproducible Data the First Time. Trends Biotechnol. 2019, 37, 761–774. [Google Scholar] [CrossRef]
- Maldonado, G.; Greenland, S. Simulation Study of Confounder-Selection Strategies. Am. J. Epidemiol. 1993, 138, 923–936. [Google Scholar] [CrossRef]
- Vittinghoff, E.; Glidden, D.V.; Shiboski, S.C.; McCulloch, C.E. Regression Methods in Biostatistics: Linear, Logistic, Survival, and Repeated Measures Models; Gail, K., Krickeberg, K., Samet, J., Tsiatis, A., Wong, W., Eds.; Springer Science + Business Media, Inc.: New York, NY, USA, 2010; Volume 53. [Google Scholar] [CrossRef]


{kind=link}
| Variables | Nutritional Intervention | p-Value | |
|---|---|---|---|
| Baseline | 6 Months | ||
| Weight (kg) | 72.5 (21.5) | 69.6 (19.5) | 0.002 |
| Body mass index (kg/m2) | 26.4 (8.1) | 26.2 (7.3) | 0.002 |
| Fat mass (kg) | 30 (16.9) | 26.3 (16.2) | 0.002 |
| Fat mass (%) | 41.6 (7.95) | 39.3 (7.4) | 0.03 |
| Fat mass index (kg/m2) | 11.1 (5.3) | 10.1 (5.7) | 0.002 |
| Visceral fat (g) | 697 (530.7) | 590 (582.2) | 0.01 |
| Fat-free mass (kg) | 40.3 (8.9) | 41.1 (9.73) | 0.08 |
| Appendicular mass index (kg/m2) | 5.1 (1.6) | 5.7 (1.48) | 0.06 |
| Outcomes | miRNA-30 | miRNA-155 | Zeta Potential | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Explanatory Variables | Regression Coefficient | p-Value | 95%CI | Regression Coefficient | p-Value | 95%CI | Regression Coefficient | p-Value | 95%CI | |||
| TyG index 1 | −0.375 | 0.344 | −1.151 | 0.401 | 0.009 | 0.97 | −0.493 | 0.512 | 0.231 | 0.706 | −0.9696 | −1.432 |
| Glucose (mg/dL) | −0.006 | 0.394 | −0.020 | 0.008 | −0.0001 | 0.977 | −0.009 | 0.008 | −0.017 | 0.092 * | −0.038 | 0.002 |
| VLDL (mg/dL) | −0.030 | 0.267 * | −0.084 | 0.023 | −0.003 | 0.835 | −0.036 | 0.029 | 0.057 | 0.125 * | −0.015 | 0.13 |
| LDL (mg/dL) | −0.012 | 0.003 * | −0.020 | −0.004 | −0.004 | 0.204 ** | −0.010 | 0.002 | −0.008 | 0.239 ** | −0.023 | 0.005 |
| HDL (mg/dL) | 0.024 | 0.121 * | −0.006 | 0.054 | 0.015 | 0.118 * | −0.003 | 0.033 | −0.015 | 0.564 | −0.066 | 0.036 |
| Triglycerides (mg/dL) | −0.001 | 0.597 | −0.009 | 0.005 | 0.0008 | 0.708 | −0.003 | 0.005 | 0.007 | 0.124 * | −0.002 | 0.017 |
| Total cholesterol (mg/dL) | −0.009 | 0.021 * | −0.016 | −0.001 | −0.002 | 0.461 | −0.007 | 0.003 | −0.005 | 0.374 | −0.018 | 0.007 |
| Lean mass index (kg/m2) | −0.094 | 0.576 | −0.428 | 0.238 | −0.103 | 0.326 | −0.310 | 0.103 | 0.013 | 0.957 | −0.492 | 0.520 |
| Visceral fat (g) | −0.0006 | 0.289 * | −0.001 | 0.0005 | −0.0007 | 0.087 * | −0.001 | 0.0001 | 0.001 | 0.21 ** | −0.0007 | 0.003 |
| Fat mass index (kg/m2) | −0.028 | 0.698 | −0.171 | 0.114 | −0.081 | 0.062 * | −0.167 | 0.004 | 0.235 | 0.029 * | 0.024 | 0.446 |
| Physical activity (minutes) | −0.0001 | 0.898 | −0.002 | 0.002 | 0.0001 | 0.84 | −0.001 | 0.001 | −0.002 | 0.192 * | −0.005 | 0.001 |
| Weight (kg) | −0.015 | 0.298 * | −0.043 | 0.013 | −0.017 | 0.05 * | −0.035 | 0.00003 | 0.026 | 0.291 ** | −0.022 | 0.075 |
| Ki67 | 0.408 | 0.334 | −0.419 | 1.235 | 0.462 | 0.086 * | −0.065 | 0.989 | −0.937 | 0.199 * | −2.368 | 0.493 |
| ER-positive | −0.004 | 0.993 | −0.966 | 0.958 | −0.378 | 0.242 ** | −1.013 | 0.255 | 1.643 | 0.032 * | 0.138 | 3.147 |
| HER 2/NEU (+) | 0.852 | 0.048 * | 0.006 | 1.698 | 0.066 | 0.834 | −0.553 | 0.686 | 1.374 | 0.063 * | −0.075 | 2.825 |
| Luminal A | −0.515 | 0.575 | −2.315 | 1.284 | −0.292 | 0.63 | −1.486 | 0.900 | −1.206 | 0.456 | −4.383 | 1.969 |
| Luminal B | −0.808 | 0.384 | −2.626 | 1.010 | −0.707 | 0.25 ** | −1.912 | 0.498 | −0.128 | 0.937 | −3.338 | 3.080 |
| BC Grade II | 0.102 | 0.876 | −1.191 | 1.397 | −0.077 | 0.862 | −0.950 | 0.795 | −0.947 | 0.423 | −3.266 | 1.370 |
| BC Grade III | 0.124 | 0.886 | −1.570 | 1.819 | 0.412 | 0.479 | −0.730 | 1.555 | −1.082 | 0.485 | −4.118 | 1.953 |
| Outcomes | Explanatory Variables | Regression Coefficient | p-Value | 95%CI | AIC | |
|---|---|---|---|---|---|---|
| miRNA-155 | Visceral fat (g) | −0.0007 | 0.050 | −0.001 | −7.04 × 10−7 | 87.731 |
| Ki67 | 0.478 | 0.049 | 0.002 | 0.953 | ||
| Zeta potential | ER-positive | 1.572 | 0.010 | 0.370 | 2.775 | 132.326 |
| HER2/NEU(+) | 2.159 | 0.000 | 1.111 | 3.206 | ||
| Ki67 | −1.391 | 0.007 | −2.401 | −0.381 | ||
| Weight (kg) | −0.094 | 0.005 | −0.160 | −0.028 | ||
| Visceral fat (g) | 0.004 | 0.000 | 0.002 | 0.0075 | ||
| miRNA-30 | HDL (mg/dL) | 0.026 | 0.053 | −0.0003 | 0.053 | 116.137 |
| LDL (mg/dL) | −0.012 | 0.001 | −0.020 | −0.004 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendivil-Alvarado, H.; Limon-Miro, A.T.; Carvajal-Millan, E.; Lizardi-Mendoza, J.; Mercado-Lara, A.; Coronado-Alvarado, C.D.; Rascón-Durán, M.L.; Anduro-Corona, I.; Talamás-Lara, D.; Rascón-Careaga, A.; et al. Extracellular Vesicles and Their Zeta Potential as Future Markers Associated with Nutrition and Molecular Biomarkers in Breast Cancer. Int. J. Mol. Sci. 2023, 24, 6810. https://doi.org/10.3390/ijms24076810
Mendivil-Alvarado H, Limon-Miro AT, Carvajal-Millan E, Lizardi-Mendoza J, Mercado-Lara A, Coronado-Alvarado CD, Rascón-Durán ML, Anduro-Corona I, Talamás-Lara D, Rascón-Careaga A, et al. Extracellular Vesicles and Their Zeta Potential as Future Markers Associated with Nutrition and Molecular Biomarkers in Breast Cancer. International Journal of Molecular Sciences. 2023; 24(7):6810. https://doi.org/10.3390/ijms24076810
Chicago/Turabian StyleMendivil-Alvarado, Herminia, Ana Teresa Limon-Miro, Elizabeth Carvajal-Millan, Jaime Lizardi-Mendoza, Araceli Mercado-Lara, Carlos D. Coronado-Alvarado, María L. Rascón-Durán, Iván Anduro-Corona, Daniel Talamás-Lara, Antonio Rascón-Careaga, and et al. 2023. "Extracellular Vesicles and Their Zeta Potential as Future Markers Associated with Nutrition and Molecular Biomarkers in Breast Cancer" International Journal of Molecular Sciences 24, no. 7: 6810. https://doi.org/10.3390/ijms24076810
APA StyleMendivil-Alvarado, H., Limon-Miro, A. T., Carvajal-Millan, E., Lizardi-Mendoza, J., Mercado-Lara, A., Coronado-Alvarado, C. D., Rascón-Durán, M. L., Anduro-Corona, I., Talamás-Lara, D., Rascón-Careaga, A., & Astiazarán-García, H. (2023). Extracellular Vesicles and Their Zeta Potential as Future Markers Associated with Nutrition and Molecular Biomarkers in Breast Cancer. International Journal of Molecular Sciences, 24(7), 6810. https://doi.org/10.3390/ijms24076810