
WAX INDUCER 1 Regulates β-Diketone Biosynthesis by Mediating Expression of the Cer-cqu Gene Cluster in Barley

 , , , , , and
, , , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Mutations in the WIN1 Gene Cause a Glossy Leaf Sheath Phenotype
2.2. The win1 KO-Associated Glossy Leaf Sheath Phenotype Is Recessive
2.3. WIN1 Is Allelic to Cer-x
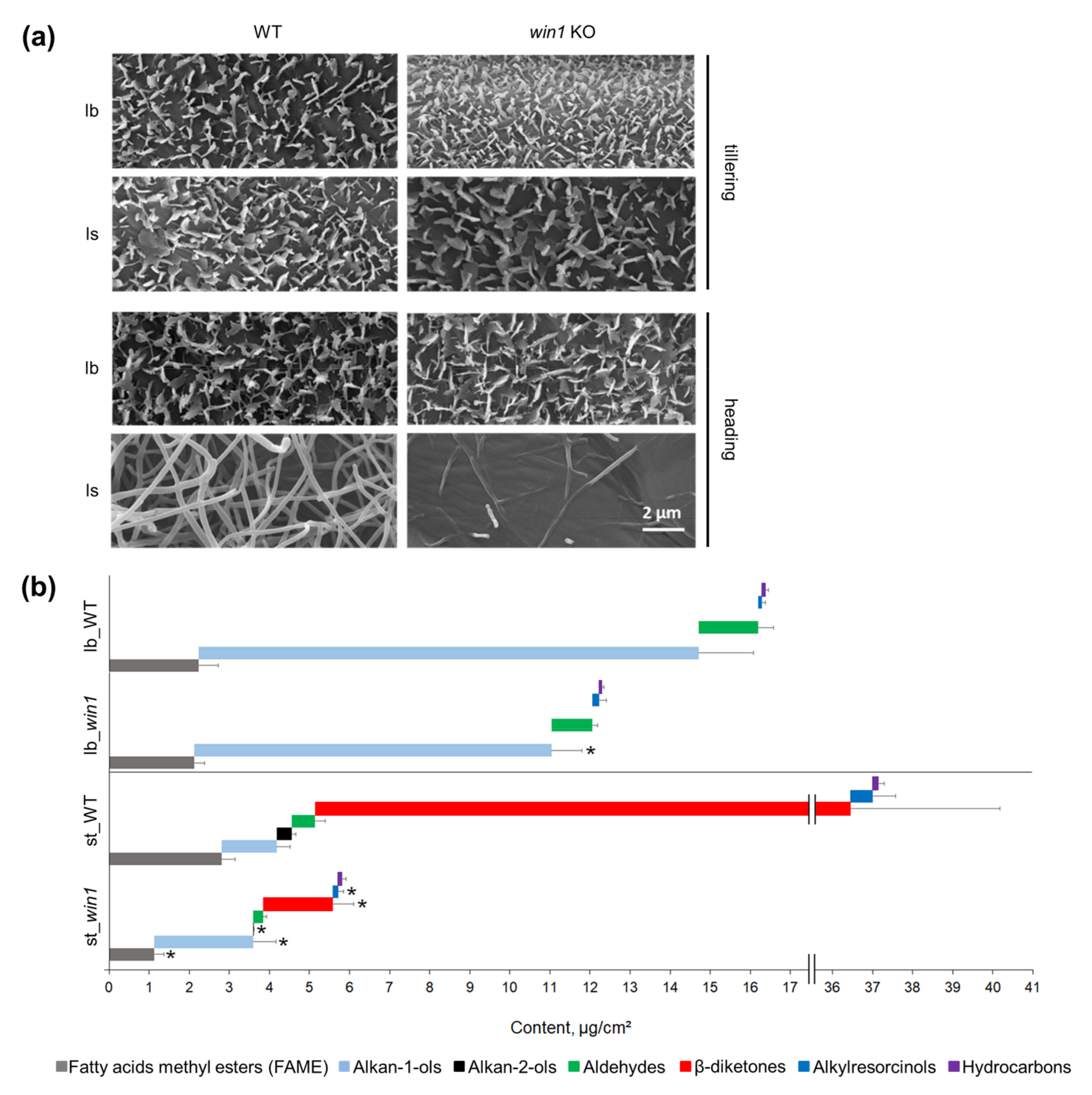
2.4. win1 Mutants Are Deficient in Tubule-Shaped Microscopic Cuticle Structures
2.5. win1 KO Plants Showed Reduced β-Diketone and Related Wax Compound Coverage
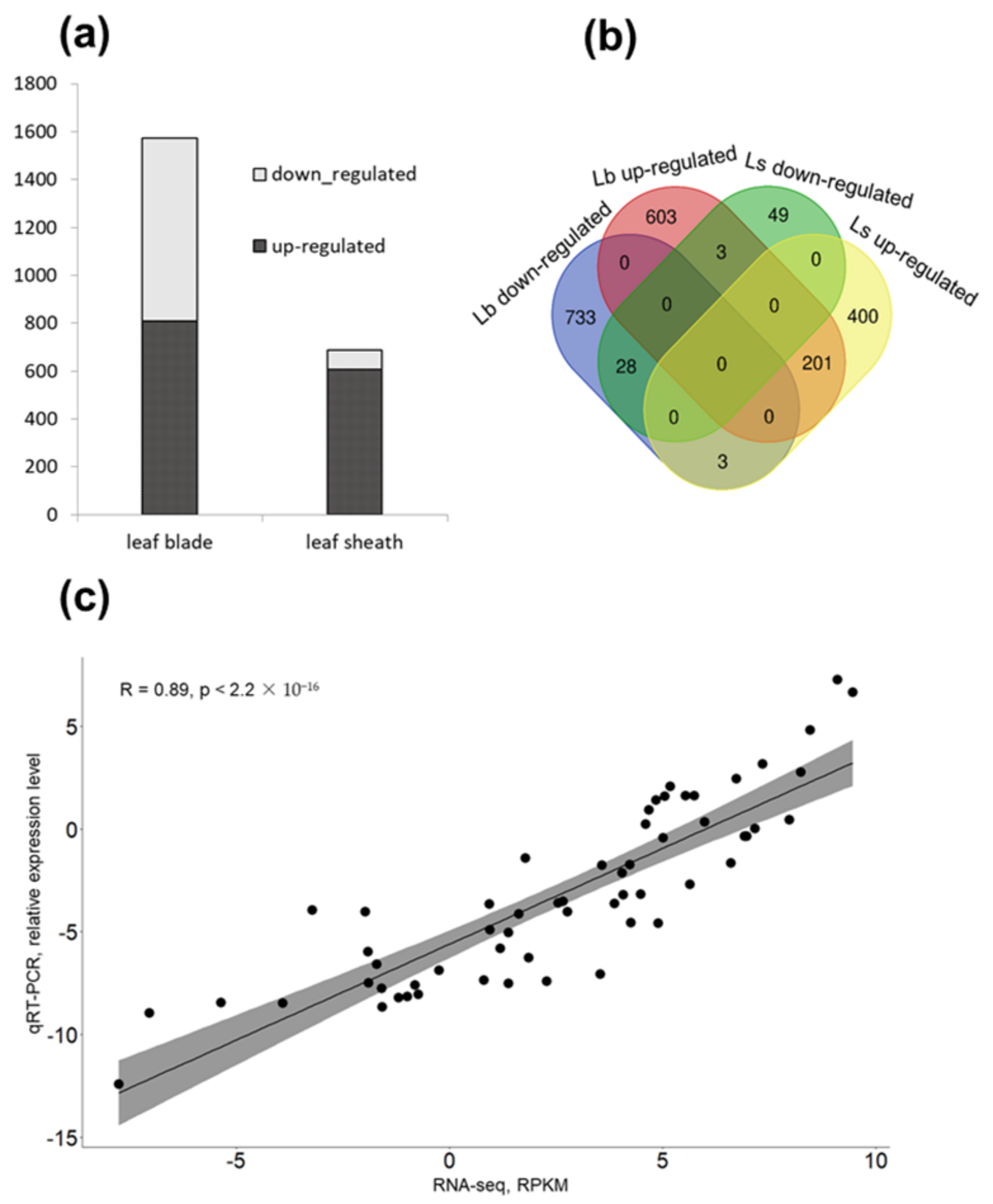
2.6. Transcriptome Analysis Reveals a High Number of WIN1-Dependent Genes
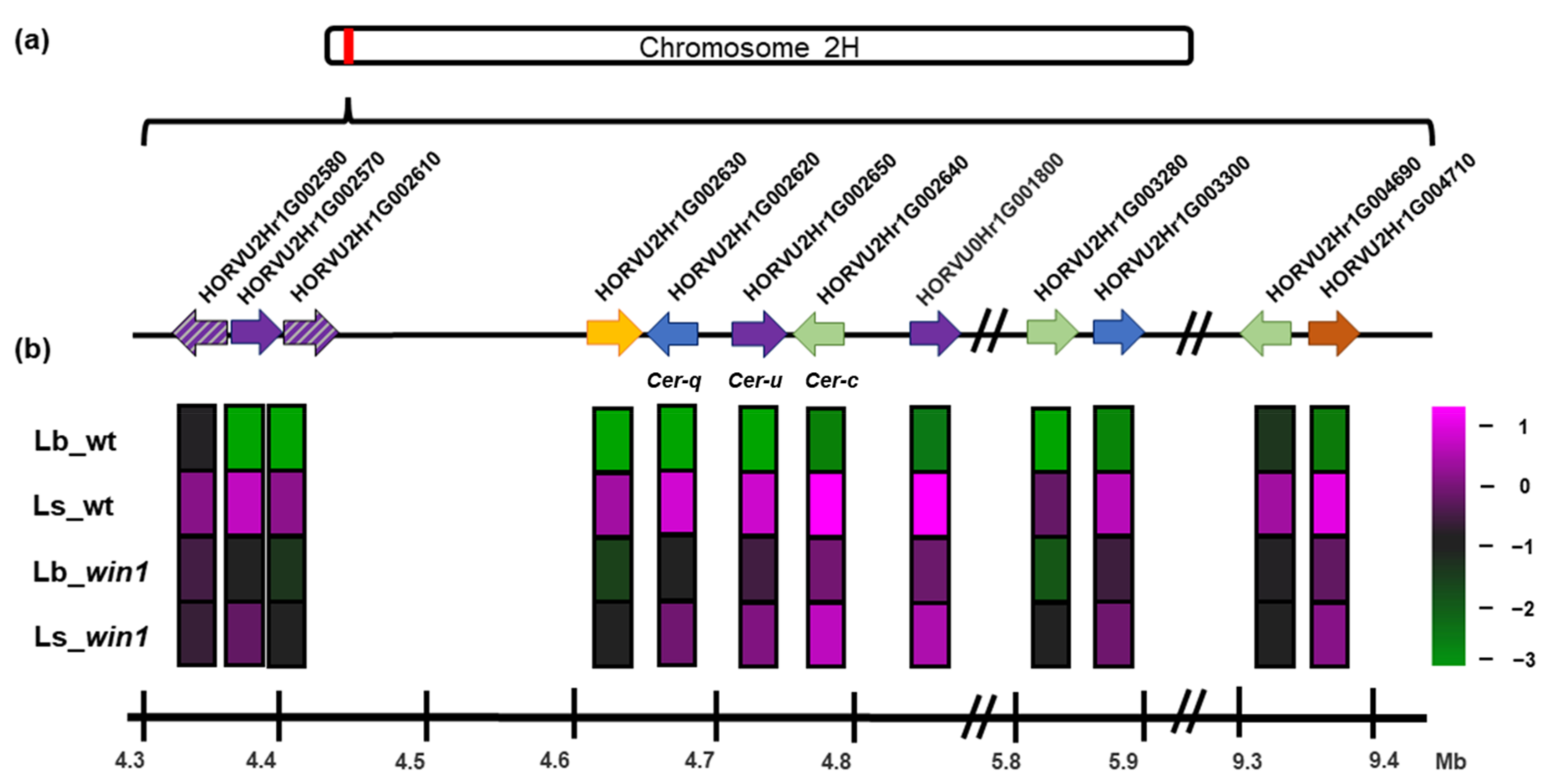
2.7. A WIN1-Dependent Set of Enzyme-Encoding Genes Is Contained in a Five-Mb Region of Chromosome 2H
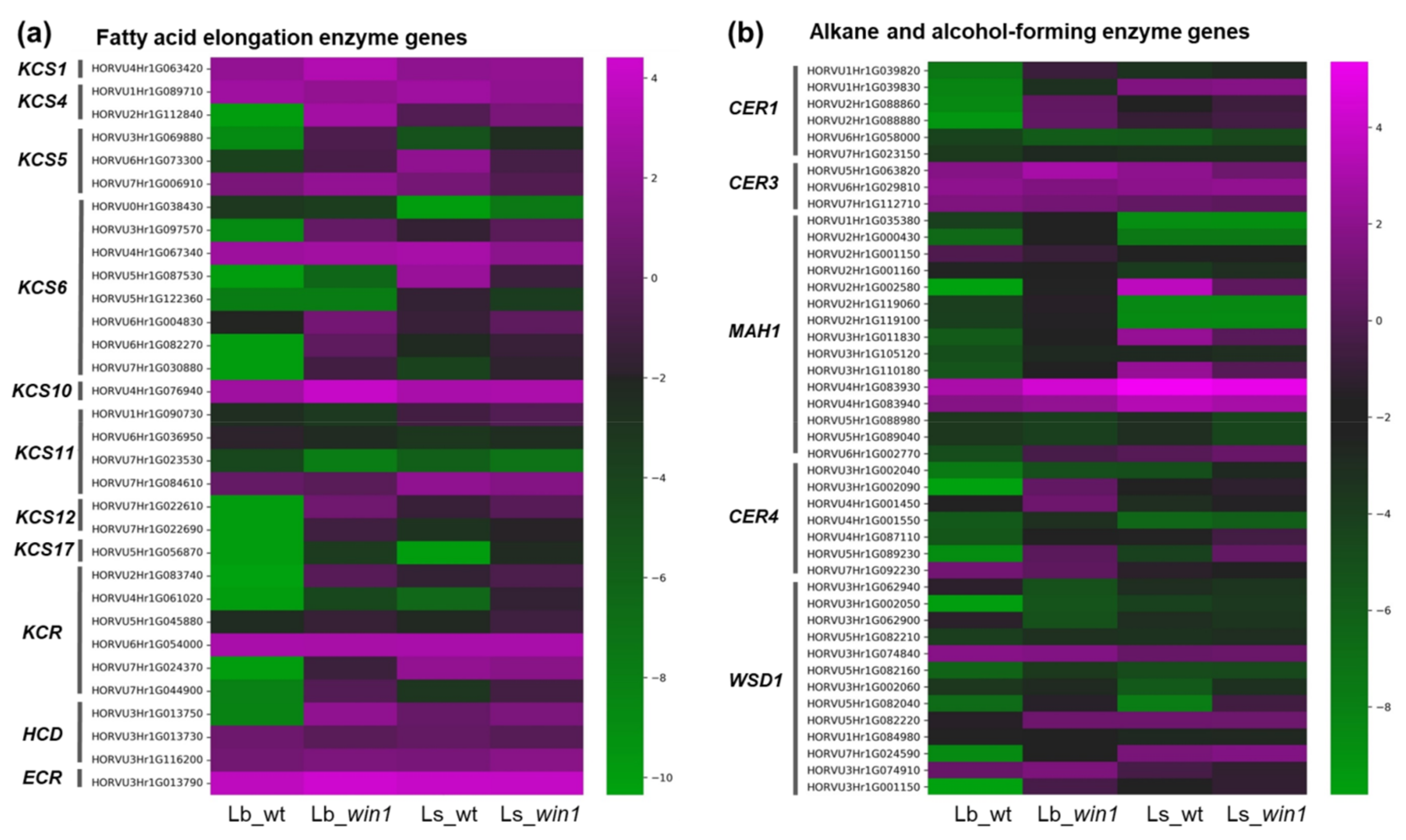
2.8. Barley Wax-Related Genes Exhibit Tissue-Specific Expression Patterns
3. Discussion
3.1. WIN1/win1 Isogenic Lines as Experimental Model to Study Cuticle Wax Formation in Barley
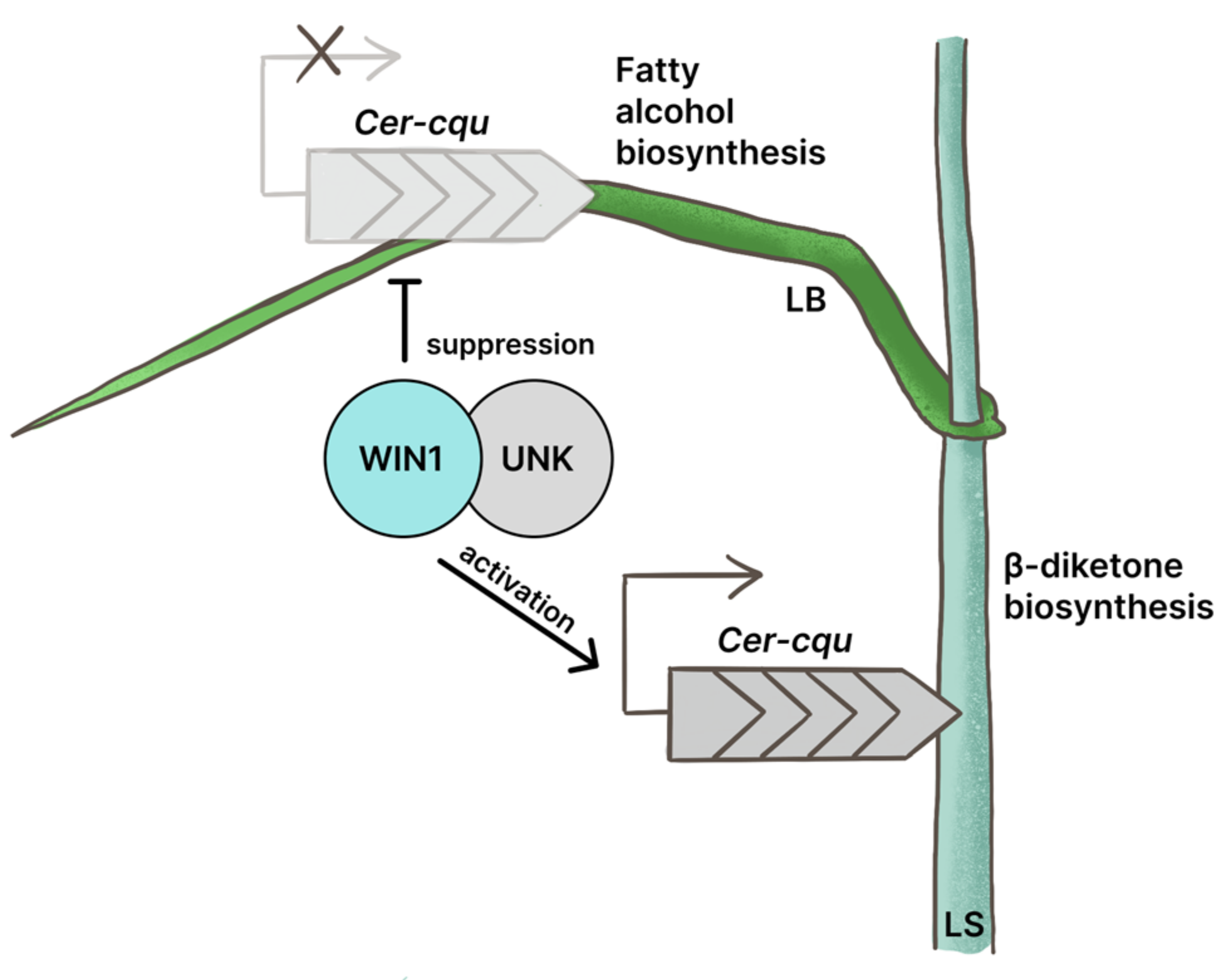
3.2. The WIN1 Transcription Factor Specifically Regulates the β-Diketone Biosynthesis Pathway
3.3. The Extended Cer-cqu Cluster Comprises Twelve Genes and Pseudogenes Regulated by the WIN1 Transcription Factor
3.4. The WIN1 Transcription Factor Is Involved in Complex Regulatory Mechanisms
4. Materials and Methods
4.1. Plant Material
4.2. Generation of Homozygous win1 KO Lines
4.3. General Phenotypic Analysis of the win1 KO Lines
4.4. Analysis of Mutant Phenotype Inheritance and Complementation Test
4.5. Scanning Electron Microscopy
4.6. Wax Analysis
4.7. RNA-seq Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petit, J.; Bres, C.; Mauxion, J.-P.; Bakan, B.; Rothan, C. Breeding for Cuticle-Associated Traits in Crop Species: Traits, Targets, and Strategies. J. Exp. Bot. 2017, 68, 5369–5387. [Google Scholar] [CrossRef] [PubMed]
- Lundqvist, U.; Lundqvist, A. Mutagen Specificity in Barley for 1580 Eceriferum Mutants Localized to 79 Loci. Hereditas 1988, 108, 1–12. [Google Scholar] [CrossRef]
- Li, C.; Haslam, T.M.; Krger, A.; Schneider, L.M.; Mishina, K.; Samuels, L.; Yang, H.; Kunst, L.; Schaffrath, U.; Nawrath, C.; et al. The β-Ketoacyl-CoA Synthase Hv KCS1, Encoded by Cer-Zh, Plays a Key Role in Synthesis of Barley Leaf Wax and Germination of Barley Powdery Mildew. Plant Cell Physiol. 2018, 59, 806–822. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Jose, S.B.; Campoli, C.; Bayer, M.M.; Sánchez-Diaz, M.A.; McAllister, T.; Zhou, Y.; Eskan, M.; Milne, L.; Schreiber, M.; et al. Conserved Signalling Components Coordinate Epidermal Patterning and Cuticle Deposition in Barley. Nat. Commun. 2022, 13, 6050. [Google Scholar] [CrossRef]
- Schneider, L.M.; Adamski, N.M.; Christensen, C.E.; Stuart, D.B.; Vautrin, S.; Hansson, M.; Uauy, C.; Von Wettstein-Knowles, P. The Cer-Cqu Gene Cluster Determines Three Key Players in a β-Diketone Synthase Polyketide Pathway Synthesizing Aliphatics in Epicuticular Waxes. J. Exp. Bot. 2016, 67, 2715–2730. [Google Scholar] [CrossRef]
- Li, C.; Wang, A.; Ma, X.; Pourkheirandish, M.; Sakuma, S.; Wang, N.; Ning, S.; Nevo, E.; Nawrath, C.; Komatsuda, T.; et al. An Eceriferum Locus, Cer-Zv, Is Associated with a Defect in Cutin Responsible for Water Retention in Barley (Hordeum vulgare) Leaves. Theor. Appl. Genet. 2013, 126, 637–646. [Google Scholar] [CrossRef]
- Li, C.; Liu, C.; Ma, X.; Wang, A.; Duan, R.; Nawrath, C.; Komatsuda, T.; Chen, G. Characterization and Genetic Mapping of Eceriferum-Ym (Cer-Ym), A Cutin Deficient Barley Mutant with Impaired Leaf Water Retention Capacity. Breed. Sci. 2015, 65, 327–332. [Google Scholar] [CrossRef]
- Zhou, Q.; Li, C.; Mishina, K.; Zhao, J.; Zhang, J.; Duan, R.; Ma, X.; Wang, A.; Meng, Q.; Komatsuda, T.; et al. Characterization and Genetic Mapping of the β-Diketone Deficient Eceriferum-b Barley Mutant. Theor. Appl. Genet. 2017, 130, 1169–1178. [Google Scholar] [CrossRef]
- Tulloch, A.P. Composition of Leaf Surface Waxes of Triticum Species: Variation with Age and Tissue. Phytochemistry 1973, 12, 2225–2232. [Google Scholar] [CrossRef]
- Yeats, T.H.; Rose, J.K.C. The Formation and Function of Plant Cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef]
- Lee, S.B.; Suh, M.C. Advances in the Understanding of Cuticular Waxes in Arabidopsis Thaliana and Crop Species. Plant Cell Rep. 2015, 34, 557–572. [Google Scholar] [CrossRef]
- Von Wettstein-Knowles, P. The Polyketide Components of Waxes and the Cer-Cqu Gene Cluster Encoding a Novel Polyketide Synthase, the Β-Diketone Synthase, DKS. Plants 2017, 6, 28. [Google Scholar] [CrossRef]
- Hen-Avivi, S.; Savin, O.; Racovita, R.C.; Lee, W.S.; Adamski, N.M.; Malitsky, S.; Almekias-Siegl, E.; Levy, M.; Vautrin, S.; Bergès, H.; et al. A Metabolic Gene Cluster in the Wheat W1 and the Barley Cer-Cqu Loci Determines β-Diketone Biosynthesis and Glaucousness. Plant Cell 2016, 28, 1440–1460. [Google Scholar] [CrossRef]
- Nützmann, H.W.; Huang, A.; Osbourn, A. Plant Metabolic Clusters—From Genetics to Genomics. New Phytol. 2016, 211, 771–789. [Google Scholar] [CrossRef]
- Kannangara, R.; Branigan, C.; Liu, Y.; Penfield, T.; Rao, V.; Mouille, G.; Höfte, H.; Pauly, M.; Riechmann, J.L.; Broun, P. The Transcription Factor WIN1/SHN1 Regulates Cutin Biosynthesis in Arabidopsis Thaliana. Plant Cell 2007, 19, 1278–1294. [Google Scholar] [CrossRef]
- Aharoni, A.; Dixit, S.; Jetter, R.; Thoenes, E.; van Arkel, G.; Pereira, A. The SHINE Clade of AP2 Domain Transcription Factors Activates Wax Biosynthesis, Alters Cuticle Properties, and Confers Drought Tolerance When Overexpressed in Arabidopsis. Plant Cell 2004, 16, 2463–2480. [Google Scholar] [CrossRef]
- Bi, H.; Shi, J.; Kovalchuk, N.; Luang, S.; Bazanova, N.; Chirkova, L.; Zhang, D.; Shavrukov, Y.; Stepanenko, A.; Tricker, P.; et al. Overexpression of the TaSHN1 Transcription Factor in Bread Wheat Leads to Leaf Surface Modifications, Improved Drought Tolerance, and No Yield Penalty under Controlled Growth Conditions. Plant Cell Environ. 2018, 41, 2549–2566. [Google Scholar] [CrossRef]
- Li, X.; Liu, N.; Sun, Y.; Wang, P.; Ge, X.; Pei, Y.; Liu, D.; Ma, X.; Li, F.; Hou, Y. The Cotton GhWIN2 Gene Activates the Cuticle Biosynthesis Pathway and Influences the Salicylic and Jasmonic Acid Biosynthesis Pathways. BMC Plant Biol. 2019, 19, 379. [Google Scholar] [CrossRef]
- Taketa, S.; Amano, S.; Tsujino, Y.; Sato, T.; Saisho, D.; Kakeda, K.; Nomura, M.; Suzuki, T.; Matsumoto, T.; Sato, K.; et al. Barley Grain with Adhering Hulls Is Controlled by an ERF Family Transcription Factor Gene Regulating a Lipid Biosynthesis Pathway. Proc. Natl. Acad. Sci. USA 2008, 105, 4062–4067. [Google Scholar] [CrossRef]
- Gerasimova, S.V.; Hertig, C.; Korotkova, A.M.; Kolosovskaya, E.V.; Otto, I.; Hiekel, S.; Kochetov, A.V.; Khlestkina, E.K.; Kumlehn, J. Conversion of Hulled into Naked Barley by Cas Endonuclease-Mediated Knockout of the NUD Gene. BMC Plant Biol. 2020, 20, 255. [Google Scholar] [CrossRef]
- McAllister, T.; Campoli, C.; Eskan, M.; Liu, L.; McKim, S.M. A Gene Encoding a SHINE1/WAX INDUCER1 Transcription Factor Controls Cuticular Wax in Barley. Agronomy 2022, 12, 1088. [Google Scholar] [CrossRef]
- Kumar, A.; Yogendra, K.N.; Karre, S.; Kushalappa, A.C.; Dion, Y.; Choo, T.M. WAX INDUCER1 (HvWIN1) Transcription Factor Regulates Free Fatty Acid Biosynthetic Genes to Reinforce Cuticle to Resist Fusarium Head Blight in Barley Spikelets. J. Exp. Bot. 2016, 67, 4127–4139. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, J.D. Structure and Biosynthesis of β-Diketones in Barley Spike Epicuticular Wax. Carlsberg Res. Commun. 1979, 44, 133–147. [Google Scholar] [CrossRef]
- Mascher, M. Pseudomolecules and Annotation of the Second Version of the Reference Genome Sequence Assembly of Barley Cv. Morex [Morex V2]; IPK Gatersleben: Gatersleben, Germany, 2019. [Google Scholar]
- Sun, Y.; Yao, R.; Ji, X.; Wu, H.; Luna, A.; Wang, Z.; Jetter, R. Characterization of an Alkylresorcinol Synthase That Forms Phenolics Accumulating in the Cuticular Wax on Various Organs of Rye (Secale cereale). Plant J. 2020, 102, 1294–1312. [Google Scholar] [CrossRef] [PubMed]
- Oshima, Y.; Shikata, M.; Koyama, T.; Ohtsubo, N.; Mitsuda, N.; Ohme-Takagi, M. MIXTA-Like Transcription Factors and WAX INDUCER1/SHINE1 Coordinately Regulate Cuticle Development in Arabidopsis and Torenia Fournieri. Plant Cell 2013, 25, 1609–1624. [Google Scholar] [CrossRef]
- Tong, T.; Fang, Y.X.; Zhang, Z.; Zheng, J.; Zhang, X.; Li, J.; Niu, C.; Xue, D.; Zhang, X. Genome-Wide Identification and Expression Pattern Analysis of the KCS Gene Family in Barley. Plant Growth Regul. 2020, 93, 89–103. [Google Scholar] [CrossRef]
- Weidenbach, D.; Jansen, M.; Franke, R.B.; Hensel, G.; Weissgerber, W.; Ulferts, S.; Jansen, I.; Schreiber, L.; Korzun, V.; Pontzen, R.; et al. Evolutionary Conserved Function of Barley and Arabidopsis 3-KETOACYL-CoA SYNTHASES in Providing Wax Signals for Germination of Powdery Mildew Fungi. Plant Physiol. 2014, 166, 1621–1633. [Google Scholar] [CrossRef]
- Von Wettstein-Knowles, P. Biosynthetic Relationships between β-Diketones and Esterified Alkan-2-Ols Deduced from Epicuticular Wax of Barley Mutants. Mol. Gen. Genet. 1976, 144, 43–48. [Google Scholar] [CrossRef]
- Bernard, A.; Joubès, J. Arabidopsis Cuticular Waxes: Advances in Synthesis, Export and Regulation. Prog. Lipid Res. 2013, 52, 110–129. [Google Scholar] [CrossRef]
- Richardson, A.; Franke, R.; Kerstiens, G.; Jarvis, M.; Schreiber, L.; Fricke, W. Cuticular Wax Deposition in Growing Barley (Hordeum vulgare) Leaves Commences in Relation to the Point of Emergence of Epidermal Cells from the Sheaths of Older Leaves. Planta 2005, 222, 472–483. [Google Scholar] [CrossRef]
- Karpichev, I.V.; Cornivelli, L.; Small, G.M. Multiple Regulatory Roles of a Novel Saccharomyces Cerevisiae Protein, Encoded by YOL002c, in Lipid and Phosphate Metabolism. J. Biol. Chem. 2002, 277, 19609–19617. [Google Scholar] [CrossRef]
- Von Wettstein-Knowles, P. Plant Waxes; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2012; ISBN 9780470015902. [Google Scholar]
- Cárdenas, P.D.; Sonawane, P.D.; Pollier, J.; Vanden Bossche, R.; Dewangan, V.; Weithorn, E.; Tal, L.; Meir, S.; Rogachev, I.; Malitsky, S.; et al. GAME9 Regulates the Biosynthesis of Steroidal Alkaloids and Upstream Isoprenoids in the Plant Mevalonate Pathway. Nat. Commun. 2016, 7, 10654. [Google Scholar] [CrossRef]
- Gerasimova, S.V.; Korotkova, A.M.; Hertig, C.; Hiekel, S.; Hofe, R.; Budhagatapalli, N.; Otto, I.; Hensel, G.; Shumny, V.K.; Kochetov, A.V.; et al. Targeted Genome Modifcation in Protoplasts of a Highly Regenerable Siberian Barley Cultivar Using RNA-Guided Cas9 Endonuclease. Vavilov J. Genet. Breed. 2018, 22, 1033–1039. [Google Scholar] [CrossRef]
- Marthe, C.; Kumlehn, J.; Hensel, G. Barley (Hordeum vulgare L.) Transformation Using Immature Embryos. In Methods in Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2015; Volume 1224, pp. 71–83. ISBN 9781493916955. [Google Scholar]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A Decimal Code for the Growth Stages of Cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Koukos, D.; Meletiou-Christou, M.-S.; Rhizopoulou, S. Leaf Surface Wettability and Fatty Acid Composition of Arbutus Unedo and Arbutus Andrachne Grown under Ambient Conditions in a Natural Macchia. Acta Bot. Gall. 2015, 162, 225–232. [Google Scholar] [CrossRef]
- Dove, H.; Mayes, R.W. Protocol for the Analysis of N-Alkanes and Other Plant-Wax Compounds and for Their Use as Markers for Quantifying the Nutrient Supply of Large Mammalian Herbivores. Nat. Protoc. 2006, 1, 1680–1697. [Google Scholar] [CrossRef]
- Laskoś, K.; Czyczyło-Mysza, I.M.; Dziurka, M.; Noga, A.; Góralska, M.; Bartyzel, J.; Myśków, B. Correlation between Leaf Epicuticular Wax Composition and Structure, Physio-biochemical Traits and Drought Resistance in Glaucous and Non-glaucous Near-isogenic Lines of Rye. Plant J. 2021, 108, 93–119. [Google Scholar] [CrossRef]
- Kutz, W.M.; Adkins, H. The Cleavage of Certain Unsymmetrical 1,3-Diketones. J. Am. Chem. Soc. 1930, 52, 4036–4042. [Google Scholar] [CrossRef]
- Connor, R.; Adkins, H. Alcoholysis and Hydrolysis of 1,3-Diketones and Beta-Keto Esters. J. Am. Chem. Soc. 1932, 54, 3420–3427. [Google Scholar] [CrossRef]
- Harrabi, S.; Ferchichi, A.; Bacheli, A.; Fellah, H. Policosanol Composition, Antioxidant and Anti-Arthritic Activities of Milk Thistle (Silybium marianum L.) Oil at Different Seed Maturity Stages. Lipids Health Dis. 2018, 17, 82. [Google Scholar] [CrossRef]
- Von Mühlen, C.; Marriott, P.J. Retention Indices in Comprehensive Two-Dimensional Gas Chromatography. Anal. Bioanal. Chem. 2011, 401, 2351–2360. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Li, Y.; Yan, J.; Cao, D.; Liang, Y. Analysis of Essential Oils and Fatty Acids from Platycodi Radix Using Chemometric Methods and Retention Indices. J. Chromatogr. Sci. 2013, 51, 318–330. [Google Scholar] [CrossRef] [PubMed]
- Ogihara, S. GC/MS Analysis of Long-Chain Aldehydes from Recent Coral. Am. J. Anal. Chem. 2018, 09, 46–51. [Google Scholar] [CrossRef]
- Lorenz, P.; Heinrich, M.; Conrad, J.; Heller, A.; Stintzing, F.C.; Kammerer, D.R. Comprehensive Characterisation of n -Alkylresorcinols and Other Lipid Constituents of Mercurialis Tomentosa L. from Alicante, Spain. Chem. Biodivers. 2017, 14, e1600255. [Google Scholar] [CrossRef]
- Đorđević, M.R.; Zlatković, D.B.; Radulović, N.S. Scilla Bifolia Wax as a Source of Diverse Long-Chain Resorcinols and Alkane-1,3-diols. Chem. Biodivers. 2021, 18, e2000811. [Google Scholar] [CrossRef]
- Racovita, R.C.; Hen-Avivi, S.; Fernandez-Moreno, J.-P.; Granell, A.; Aharoni, A.; Jetter, R. Composition of Cuticular Waxes Coating Flag Leaf Blades and Peduncles of Triticum Aestivum Cv. Bethlehem. Phytochemistry 2016, 130, 182–192. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Lin, H.-N.; Hsu, W.-L. DART: A Fast and Accurate RNA-Seq Mapper with a Partitioning Strategy. Bioinformatics 2018, 34, 190–197. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Von Zitzewitz, J.; Szűcs, P.; Dubcovsky, J.; Yan, L.; Francia, E.; Pecchioni, N.; Casas, A.; Chen, T.H.H.; Hayes, P.M.; Skinner, J.S. Molecular and Structural Characterization of Barley Vernalization Genes. Plant Mol. Biol. 2005, 59, 449–467. [Google Scholar] [CrossRef]
- Himi, E.; Nisar, A.; Noda, K. Colour Genes (R and Rc) for Grain and Coleoptile Upregulate Flavonoid Biosynthesis Genes in Wheat. Genome 2005, 48, 747–754. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerasimova, S.V.; Kolosovskaya, E.V.; Vikhorev, A.V.; Korotkova, A.M.; Hertig, C.W.; Genaev, M.A.; Domrachev, D.V.; Morozov, S.V.; Chernyak, E.I.; Shmakov, N.A.; et al. WAX INDUCER 1 Regulates β-Diketone Biosynthesis by Mediating Expression of the Cer-cqu Gene Cluster in Barley. Int. J. Mol. Sci. 2023, 24, 6762. https://doi.org/10.3390/ijms24076762
Gerasimova SV, Kolosovskaya EV, Vikhorev AV, Korotkova AM, Hertig CW, Genaev MA, Domrachev DV, Morozov SV, Chernyak EI, Shmakov NA, et al. WAX INDUCER 1 Regulates β-Diketone Biosynthesis by Mediating Expression of the Cer-cqu Gene Cluster in Barley. International Journal of Molecular Sciences. 2023; 24(7):6762. https://doi.org/10.3390/ijms24076762
Chicago/Turabian StyleGerasimova, Sophia V., Ekaterina V. Kolosovskaya, Alexander V. Vikhorev, Anna M. Korotkova, Christian W. Hertig, Mikhail A. Genaev, Dmitry V. Domrachev, Sergey V. Morozov, Elena I. Chernyak, Nikolay A. Shmakov, and et al. 2023. "WAX INDUCER 1 Regulates β-Diketone Biosynthesis by Mediating Expression of the Cer-cqu Gene Cluster in Barley" International Journal of Molecular Sciences 24, no. 7: 6762. https://doi.org/10.3390/ijms24076762
APA StyleGerasimova, S. V., Kolosovskaya, E. V., Vikhorev, A. V., Korotkova, A. M., Hertig, C. W., Genaev, M. A., Domrachev, D. V., Morozov, S. V., Chernyak, E. I., Shmakov, N. A., Vasiliev, G. V., Kochetov, A. V., Kumlehn, J., & Khlestkina, E. K. (2023). WAX INDUCER 1 Regulates β-Diketone Biosynthesis by Mediating Expression of the Cer-cqu Gene Cluster in Barley. International Journal of Molecular Sciences, 24(7), 6762. https://doi.org/10.3390/ijms24076762