
The Zinc-BED Transcription Factor Bedwarfed Promotes Proportional Dendritic Growth and Branching through Transcriptional and Translational Regulation in Drosophila

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
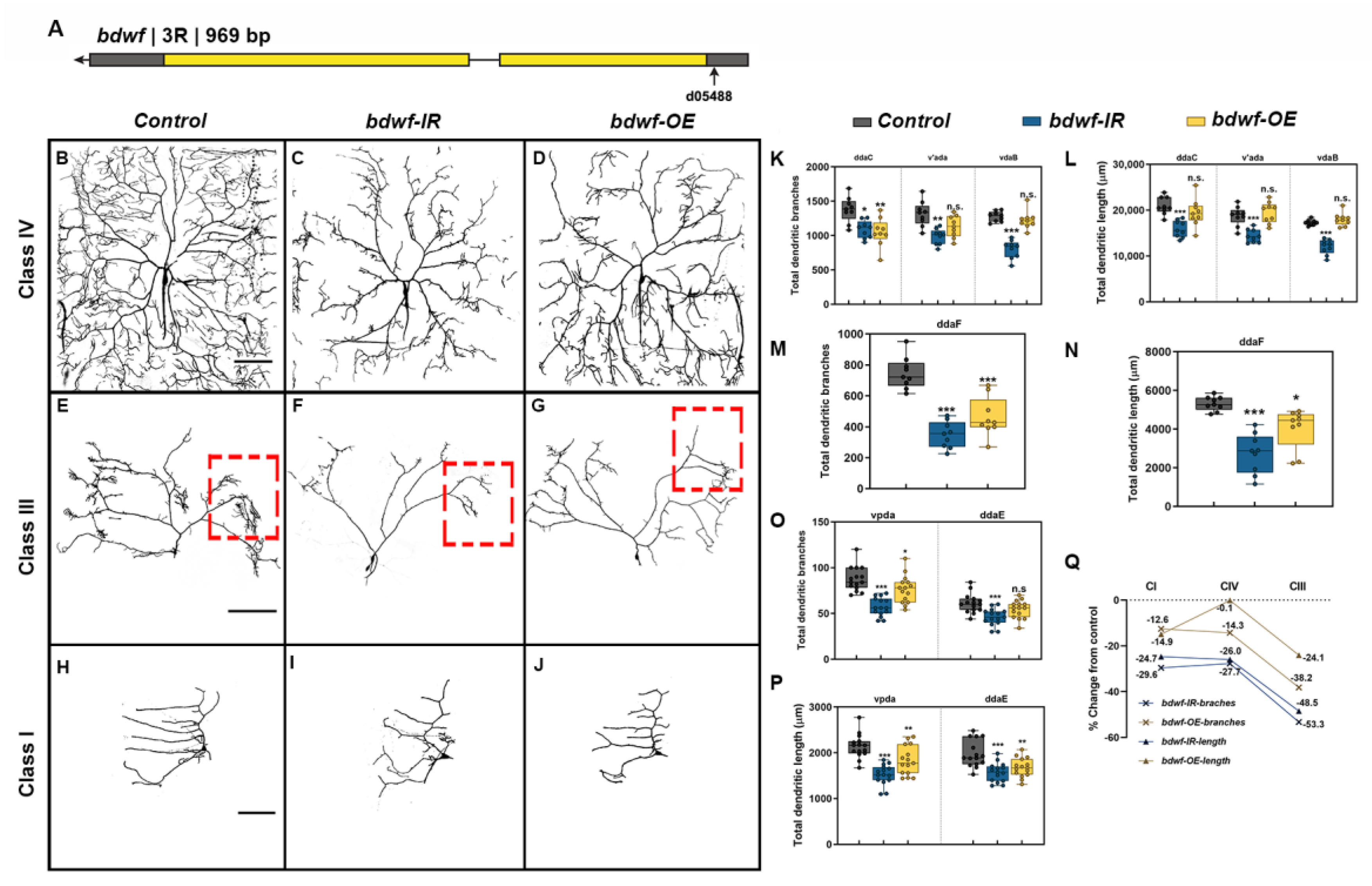
2.1. Bdwf Dysregulation Results in Dendritic Hypotrophy
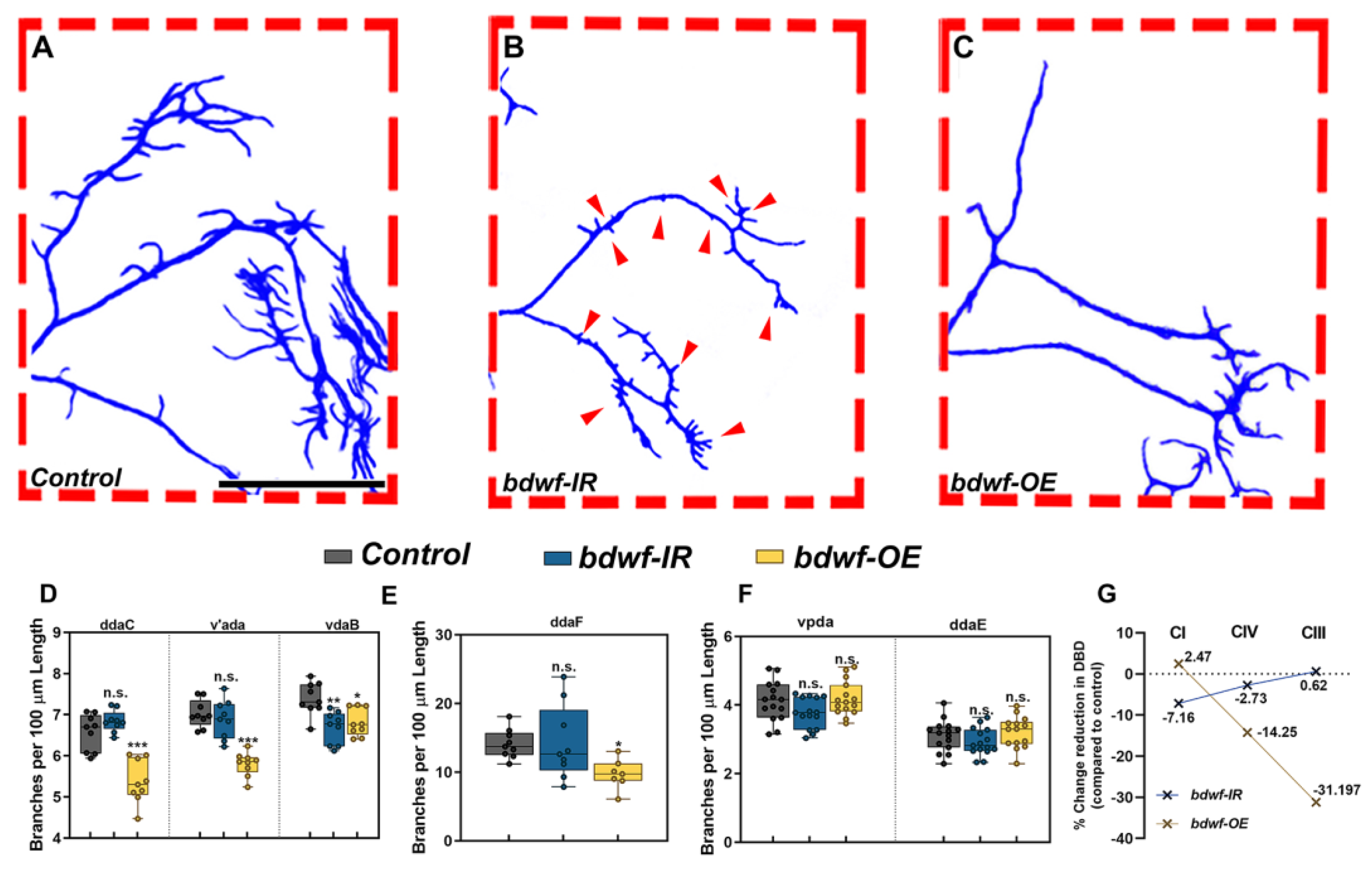
2.2. Bdwf Regulates Proportional Dendritic Growth and Branching in Morphologically Divergent Neurons

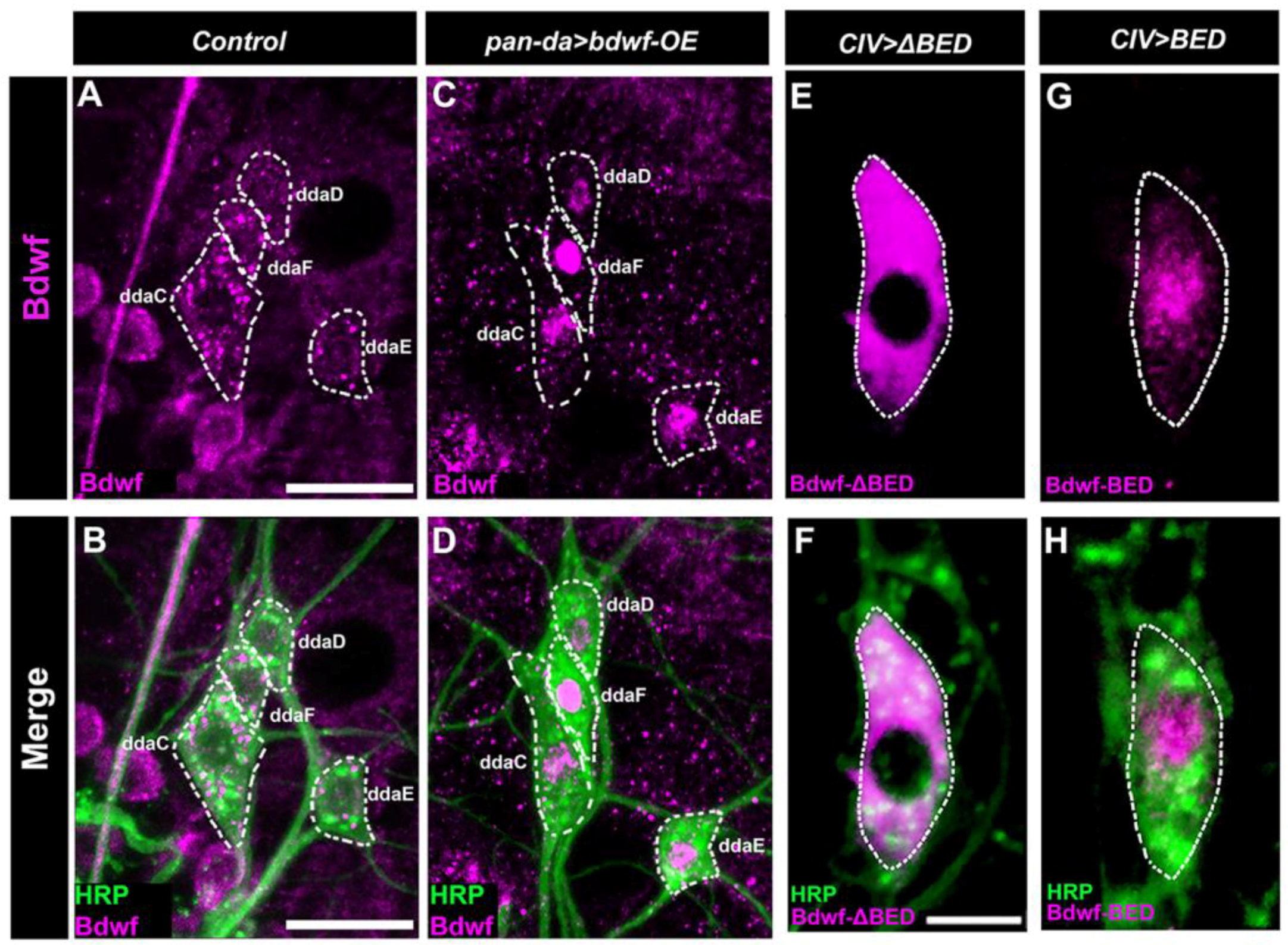
2.3. Bdwf Exhibits Nucleocytoplasmic Localization in Md Neurons
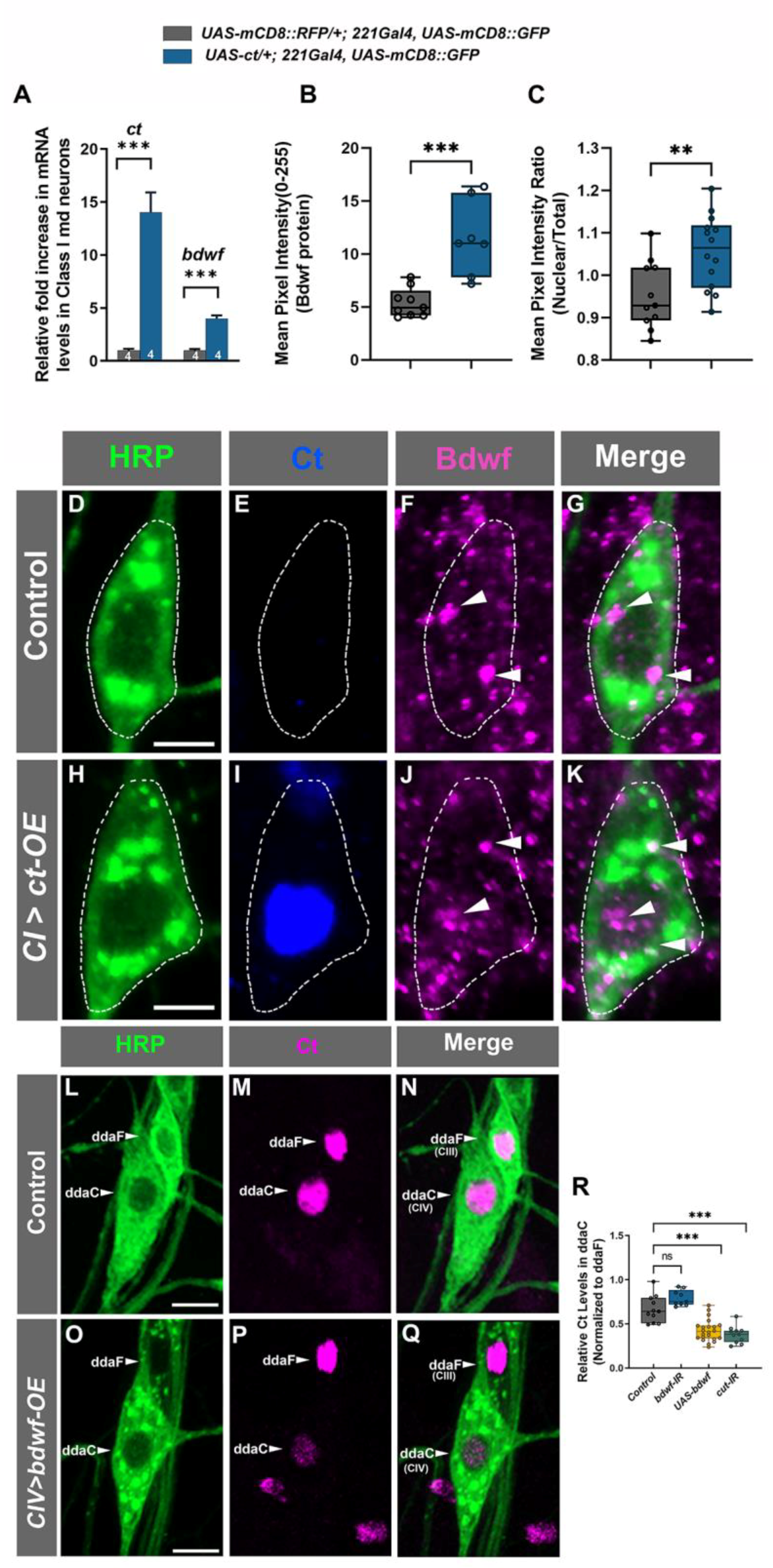
2.4. Cut and Bdwf Exhibit a Reciprocal Regulatory Relationship with Respect to Expression
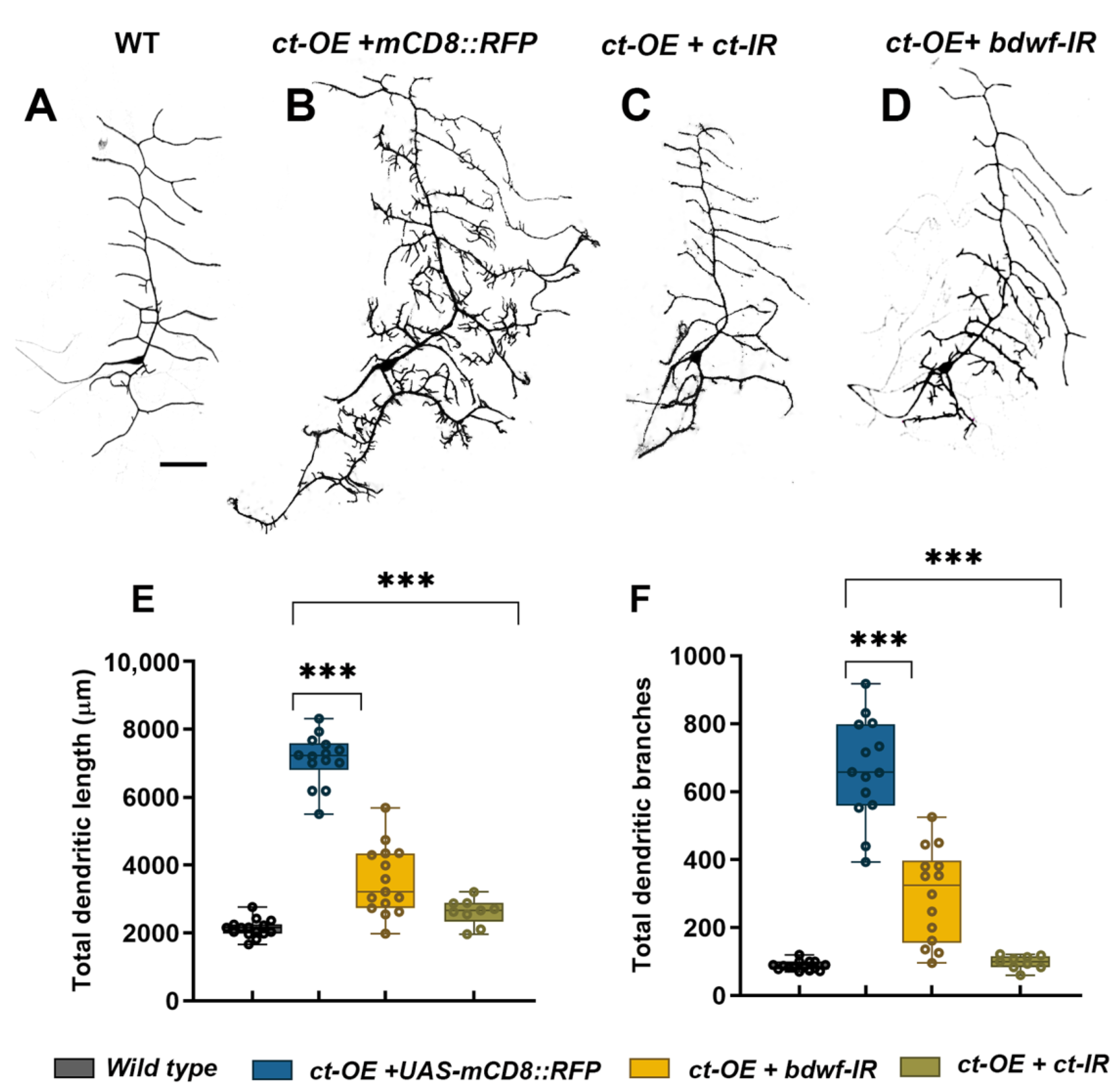
2.5. Bdwf Functions as a Downstream Effector of Cut-Mediated Dendritic Arborization
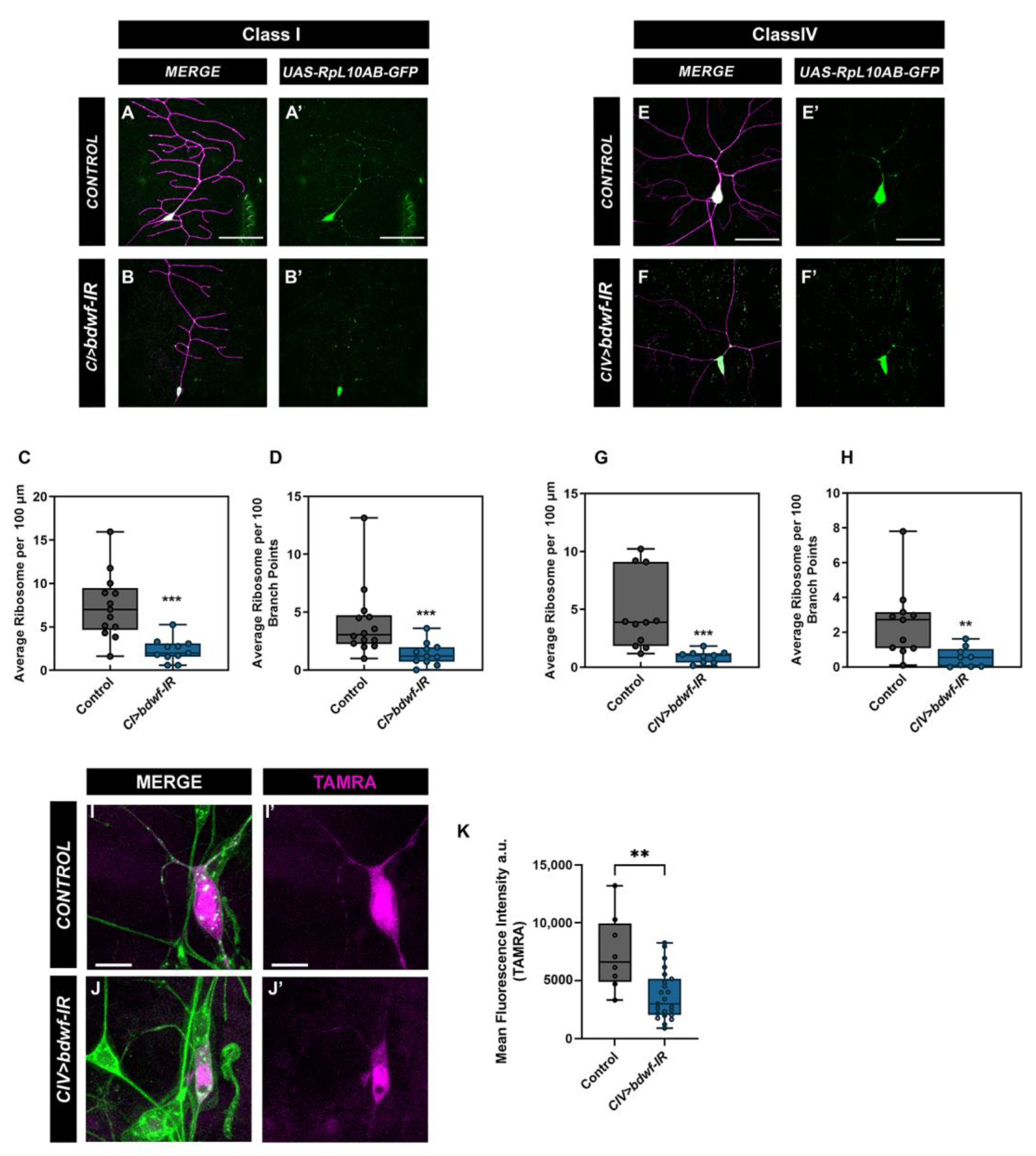
2.6. Bdwf Interacts and Colocalizes with Ribosomal Proteins
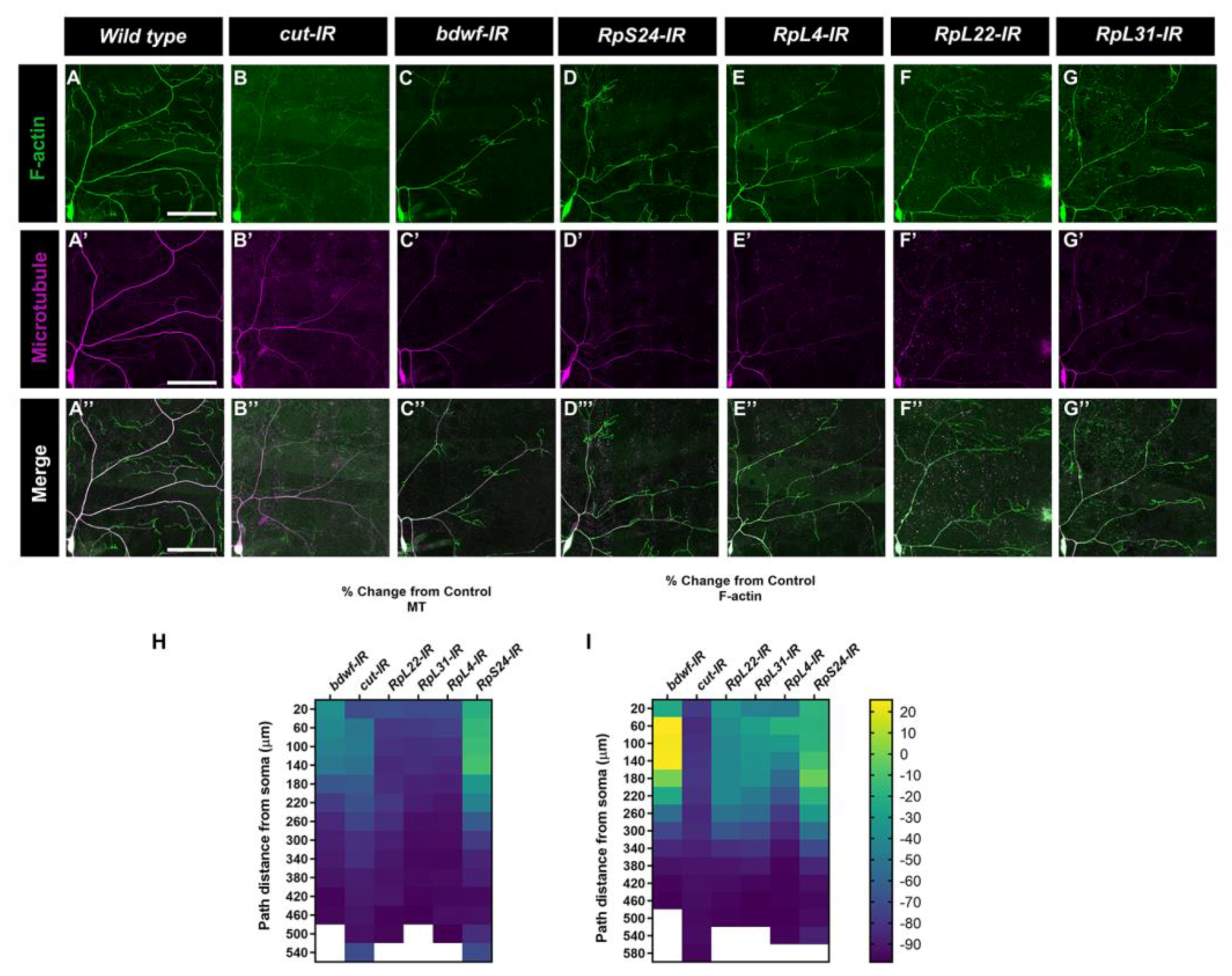
2.7. Bdwf Is Required for Ribosomal Trafficking and Protein Translation along theDendritic Arbor
3. Discussion
4. Materials and Methods
4.1. Drosophila Strains
4.2. Transgenic Strain Generation
4.3. Cell Isolation
4.4. qRT–PCR Analysis
4.5. Phenotypic Screening, live Image Confocal Microscopy, and Morphometric Quantitation
4.6. Multichannel Reconstructions
4.7. Generation of Anti-Bdwf Antibodies
4.8. Immunohistochemistry
4.9. FUNCAT Analyses
4.10. Mass Spectrometry Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lefebvre, J.L.; Sanes, J.R.; Kay, J.N. Development of Dendritic Form and Function. Annu. Rev. Cell Dev. Biol. 2015, 31, 741–777. [Google Scholar] [CrossRef]
- Prigge, C.L.; Kay, J.N. Dendrite morphogenesis from birth to adulthood. Curr. Opin. Neurobiol. 2018, 53, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Tavosanis, G. Dendrite enlightenment. Curr. Opin. Neurobiol. 2021, 69, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, J.L. Molecular mechanisms that mediate dendrite morphogenesis. Curr. Top. Dev. Biol. 2021, 142, 233–282. [Google Scholar] [CrossRef]
- Jan, Y.-N.N.; Jan, L.Y. Branching out: Mechanisms of dendritic arborization. Nat. Rev. Neuorsci. 2010, 11, 316–328. [Google Scholar] [CrossRef] [PubMed]
- de la Torre-Ubieta, L.; Bonni, A. Transcriptional Regulation of Neuronal Polarity and Morphogenesis in the Mammalian Brain. Neuron 2011, 72, 22–40. [Google Scholar] [CrossRef]
- Santiago, C.; Bashaw, G.J. Transcription factors and effectors that regulate neuronal morphology. Development 2014, 141, 4667–4680. [Google Scholar] [CrossRef]
- Valnegri, P.; Puram, S.V.; Bonni, A. Regulation of dendrite morphogenesis by extrinsic cues. Trends Neurosci. 2015, 38, 439–447. [Google Scholar] [CrossRef]
- Nanda, S.; Das, R.; Cox, D.N.; Ascoli, G.A. Structural Plasticity in Dendrites: Developmental Neurogenetics, Morphological Reconstructions, and Computational Modeling. In Neurobiological and Psychological Aspects of Brain Recovery; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–34. [Google Scholar] [CrossRef]
- Pai, Y.J.; Moore, A.W. Transcription factor encoding of neuron subtype: Strategies that specify arbor pattern. Curr. Opin. Neurobiol. 2021, 69, 149–158. [Google Scholar] [CrossRef]
- Arlotta, P.; Molyneaux, B.J.; Chen, J.; Inoue, J.; Kominami, R.; Macklis, J.D. Neuronal subtype-specific genes that control corticospinal motor neuron development in vivo. Neuron 2005, 45, 207–221. [Google Scholar] [CrossRef]
- Molyneaux, B.J.; Arlotta, P.; Menezes, J.R.L.; Macklis, J.D. Neuronal subtype specification in the cerebral cortex. Nat. Rev. Neuorsci. 2007, 8, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Puram, S.V.; Bonni, A. Cell-intrinsic drivers of dendrite morphogenesis. Development 2013, 140, 4657–4671. [Google Scholar] [CrossRef] [PubMed]
- Yalgin, C.; Ebrahimi, S.; Delandre, C.; Yoong, L.F.; Akimoto, S.; Tran, H.; Amikura, R.; Spokony, R.; Torben-Nielsen, B.; White, K.P.; et al. Centrosomin represses dendrite branching by orienting microtubule nucleation. Nat. Neurosci. 2015, 18, 1437–1445. [Google Scholar] [CrossRef]
- Sears, J.C.; Broihier, H.T. FoxO regulates microtubule dynamics and polarity to promote dendrite branching in Drosophila sensory neurons. Dev. Biol. 2016, 418, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Grueber, W.B.; Jan, L.Y.; Jan, Y.N. Tiling of the Drosophila epidermis by multidendritic sensory neurons. Development 2002, 129, 2867–2878. [Google Scholar] [CrossRef] [PubMed]
- Jinushi-Nakao, S.; Arvind, R.; Amikura, R.; Kinameri, E.; Liu, A.W.; Moore, A.W. Knot/Collier and Cut Control Different Aspects of Dendrite Cytoskeleton and Synergize to Define Final Arbor Shape. Neuron 2007, 56, 963–978. [Google Scholar] [CrossRef] [PubMed]
- Ye, B.; Zhang, Y.; Song, W.; Younger, S.H.; Jan, L.Y.; Jan, Y.N. Growing Dendrites and Axons Differ in Their Reliance on the Secretory Pathway. Cell 2007, 130, 717–729. [Google Scholar] [CrossRef] [PubMed]
- Sulkowski, M.J.; Iyer, S.C.; Kurosawa, M.S.; Iyer, E.P.R.; Cox, D.N. Turtle Functions Downstream of Cut in Differentially Regulating Class Specific Dendrite Morphogenesis in Drosophila. PLoS ONE 2011, 6, e22611. [Google Scholar] [CrossRef]
- Ye, B.; Kim, J.H.; Yang, L.; McLachlan, I.; Younger, S.; Jan, L.Y.; Jan, Y.N. Differential regulation of dendritic and axonal development by the novel Krüppel-like factor dar1. J. Neurosci. 2011, 31, 3309–3319. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.C.; Wang, D.; Iyer, E.P.R.; Trunnell, S.A.; Meduri, R.; Shinwari, R.; Sulkowski, M.J.; Cox, D.N. The RhoGEF Trio Functions in Sculpting Class Specific Dendrite Morphogenesis in Drosophila Sensory Neurons. PLoS ONE 2012, 7, e33634. [Google Scholar] [CrossRef]
- Nagel, J.; Delandre, C.; Zhang, Y.; Förstner, F.; Moore, A.W.; Tavosanis, G. Fascin controls neuronal class-specific dendrite arbor morphology. Development 2012, 139, 2999–3009. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.C.; Iyer, E.P.R.; Meduri, R.; Rubaharan, M.; Kuntimaddi, A.; Karamsetty, M.; Cox, D.N. Cut, via CrebA, transcriptionally regulates the COPII secretory pathway to direct dendrite development in Drosophila. J. Cell Sci. 2013, 126, 4732–4745. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Usui, T.; Satoh, D.; Moriyama, S.; Shimono, K.; Itoh, T.; Shirahige, K.; Uemura, T. Sensory-Neuron Subtype-Specific Transcriptional Programs Controlling Dendrite Morphogenesis: Genome-wide Analysis of Abrupt and Knot/Collier. Dev. Cell 2013, 27, 530–544. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, T.; Ou, Y.; Li, S.; Giniger, E.; van Meyel, D.J. Dendrite architecture organized by transcriptional control of the F-actin nucleator Spire. Development 2014, 141, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Das, R.; Bhattacharjee, S.; Patel, A.A.; Harris, J.M.; Bhattacharya, S.; Letcher, J.M.; Clark, S.G.; Nanda, S.; Iyer, E.P.R.; Ascoli, G.A.; et al. Dendritic Cytoskeletal Architecture Is Modulated by Combinatorial Transcriptional Regulation in Drosophila melanogaster. Genetics 2017, 207, 1401–1421. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.G.; Graybeal, L.L.; Bhattacharjee, S.; Thomas, C.; Bhattacharya, S.; Cox, D.N. Basal autophagy is required for promoting dendritic terminal branching in Drosophila sensory neurons. PLoS ONE 2018, 13, e0206743. [Google Scholar] [CrossRef]
- Yoong, L.F.; Lim, H.-K.; Tran, H.; Lackner, S.; Zheng, Z.; Hong, P.; Moore, A.W. Atypical Myosin Tunes Dendrite Arbor Subdivision. Neuron 2020, 106, 452–467. [Google Scholar] [CrossRef]
- Bhattacharjee, S.; Lottes, E.N.; Nanda, S.; Golshir, A.; Patel, A.A.; Ascoli, G.A.; Cox, D.N. PP2A phosphatase regulates cell-type specific cytoskeletal organization to drive dendrite diversity. Front. Mol. Neurosci. 2022, 15, 926567. [Google Scholar] [CrossRef]
- Corty, M.M.; Tam, J.; Grueber, W.B. Dendritic diversification through transcription factor-mediated suppression of alternative morphologies. Development 2016, 143, 1351–1362. [Google Scholar] [CrossRef]
- Iyer, E.P.R.; Iyer, S.C.; Sullivan, L.; Wang, D.; Meduri, R.; Graybeal, L.L.; Cox, D.N. Functional Genomic Analyses of Two Morphologically Distinct Classes of Drosophila Sensory Neurons: Post-Mitotic Roles of Transcription Factors in Dendritic Patterning. PLoS ONE 2013, 8, e72434. [Google Scholar] [CrossRef]
- Aravind, L. The BED finger, a novel DNA-binding domain in chromatin-boundary-element-binding proteins and transposases. Trends Biochem. Sci. 2000, 25, 421–423. [Google Scholar] [CrossRef]
- Grueber, W.B.; Jan, L.Y.; Jan, Y.N. Different Levels of the Homeodomain Protein Cut Regulate Distinct Dendrite Branching Patterns of Drosophila Multidendritic Neurons. Cell 2003, 112, 805–818. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.M.; Gillette, T.A.; Ascoli, G.A. Quantifying neuronal size: Summing up trees and splitting the branch difference. Semin. Cell Dev. Biol. 2008, 19, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Das, R.; Bhattacharjee, S.; Letcher, J.M.; Harris, J.M.; Nanda, S.; Foldi, I.; Lottes, E.N.; Bobo, H.M.; Grantier, B.D.; Mihály, J.; et al. Formin 3 directs dendritic architecture via microtubule regulation and is required for somatosensory nociceptive behavior. Development 2021, 148, dev187609. [Google Scholar] [CrossRef]
- Kilo, L.; Stürner, T.; Tavosanis, G.; Ziegler, A.B. Drosophila Dendritic Arborisation Neurons: Fantastic Actin Dynamics and Where to Find Them. Cells 2021, 10, 2777. [Google Scholar] [CrossRef]
- Nanda, S.; Bhattacharjee, S.; Cox, D.N.; Ascoli, G.A. Distinct relations of microtubules and actin filaments with dendritic architecture. iScience 2020, 23, 101865. [Google Scholar] [CrossRef]
- Nanda, S.; Bhattacharjee, S.; Cox, D.N.; Ascoli, G.A. An imaging analysis protocol to trace, quantify, and model multi-signal neuron morphology. STAR Protoc. 2021, 2, 100567. [Google Scholar] [CrossRef]
- Noma, K.; Goncharov, A.; Ellisman, M.H.; Jin, Y. Microtubule-dependent ribosome localization in C. elegans neurons. eLife 2017, 6, e26376. [Google Scholar] [CrossRef]
- Thomas, A.; Lee, P.; Dalton, J.E.; Nomie, K.J.; Stoica, L.; Costa-Mattioli, M.; Chang, P.; Nuzhdin, S.; Arbeitman, M.N.; Dierick, H.A. A Versatile Method for Cell-Specific Profiling of Translated mRNAs in Drosophila. PLoS ONE 2012, 7, e40276. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, I.; Marter, K.; Kobler, O.; Niehues, S.; Abele, J.; Müller, A.; Bussmann, J.; Storkebaum, E.; Ziv, T.; Thomas, U.; et al. Cell-selective labelling of proteomes in Drosophila melanogaster. Nat. Commun. 2015, 6, 7521. [Google Scholar] [CrossRef]
- Nyegaard, M.; Severinsen, J.E.; Als, T.D.; Hedemand, A.; Straarup, S.; Nordentoft, M.; McQuillin, A.; Bass, N.; Lawrence, J.; Thirumalai, S.; et al. Support of association between BRD1 and both schizophrenia and bipolar affective disorder. Am. J. Med. Genet. B Neuropsychiartr. Genet. 2010, 153, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Mitz, A.R.; Philyaw, T.J.; Boccuto, L.; Shcheglovitov, A.; Sarasua, S.M.; Kaufmann, W.E.; Thurm, A. Identification of 22q13 genes most likely to contribute to Phelan McDermid syndrome. Eur. J. Hum. Genet. 2018, 26, 293. [Google Scholar] [CrossRef] [PubMed]
- Schenkel, L.C.; Aref-Eshghi, E.; Rooney, K.; Kerkhof, J.; Levy, M.A.; McConkey, H.; Rogers, R.C.; Phelan, K.; Sarasua, S.M.; Jain, L.; et al. DNA methylation epi-signature is associated with two molecularly and phenotypically distinct clinical subtypes of Phelan-McDermid syndrome. Clin. Epigenet. 2021, 13, 2. [Google Scholar] [CrossRef] [PubMed]
- Seefelder, M.; Kochanek, S. A meta-analysis of transcriptomic profiles of Huntington’s disease patients. PLoS ONE 2021, 16, e0253037. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, D.; Sano, Y.; Adachi, Y.; Okamoto, Y.; Osada, H.; Takahashi, T.; Yamaguchi, T.; Osumi, T.; Hirose, F. hDREF regulates cell proliferation and expression of ribosomal protein genes. Mol. Cell. Biol. 2007, 27, 2003–2013. [Google Scholar] [CrossRef]
- Farber, D.B.; Theendakara, V.P.; Akhmedov, N.B.; Saghizadeh, M. ZBED4, a novel retinal protein expressed in cones and Müller cells. Adv. Exp. Med. Biol. 2010, 664, 79–87. [Google Scholar] [CrossRef]
- Saghizadeh, M.; Gribanova, Y.; Akhmedov, N.B.; Farber, D.B. ZBED4, a cone and Müller cell protein in human retina, has a different cellular expression in mouse. Mol. Vis. 2011, 17, 2011–2018. [Google Scholar]
- Wittenberg, G.M.; Wang, S.S.-H. Evolution and scaling of dendrites. Dendrites 2007, 43–67. [Google Scholar] [CrossRef]
- Shimono, K.; Fujishima, K.; Nomura, T.; Ohashi, M.; Usui, T.; Kengaku, M.; Toyoda, A.; Uemura, T. An evolutionarily conserved protein CHORD regulates scaling of dendritic arbors with body size. Sci. Rep. 2014, 4, 4415. [Google Scholar] [CrossRef]
- Parrish, J.Z.; Xu, P.; Kim, C.C.; Jan, L.Y.; Jan, Y.N. The microRNA bantam Functions in Epithelial Cells to Regulate Scaling Growth of Dendrite Arbors in Drosophila Sensory Neurons. Neuron 2009, 63, 788–802. [Google Scholar] [CrossRef]
- Seto, H.; Hayashi, Y.; Kwon, E.; Taguchi, O.; Yamaguchi, M. Antagonistic regulation of the Drosophila PCNA gene promoter by DREF and Cut. Genes Cells. 2006, 11, 499–512. [Google Scholar] [CrossRef] [PubMed]
- Marygold, S.J.; Roote, J.; Reuter, G.; Lambertsson, A.; Ashburner, M.; Millburn, G.H.; Harrison, P.M.; Yu, Z.; Kenmochi, N.; Kaufman, T.C.; et al. The ribosomal protein genes and Minute loci of Drosophila melanogaster. Genome Biol. 2007, 8, R216. [Google Scholar] [CrossRef] [PubMed]
- Boutros, M.; Kiger, A.A.; Armknecht, S.; Kerr, K.; Hild, M.; Koch, B.; Haas, S.A.; Paro, R.; Perrimon, N. Genome-wide RNAi analysis of growth and viability in Drosophila cells. Science 2004, 303, 832–835. [Google Scholar] [CrossRef] [PubMed]
- Slomnicki, L.P.; Pietrzak, M.; Vashishta, A.; Jones, J.; Lynch, N.; Elliot, S.; Poulos, E.; Malicote, D.; Morris, B.E.; Hallgren, J.; et al. Requirement of Neuronal Ribosome Synthesis for Growth and Maintenance of the Dendritic Tree. J. Biol. Chem. 2016, 291, 5721. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.Y.; Williams, C.; Yan, C.; Koledachkina, T.; Luedke, K.; Dalton, J.; Bloomsburg, S.; Morrison, N.; Duncan, K.E.; Kim, C.C.; et al. The SLC36 transporter Pathetic is required for extreme dendrite growth in Drosophila sensory neurons. Genes Dev. 2015, 29, 1120–1135. [Google Scholar] [CrossRef]
- Gazda, H.T.; Grabowska, A.; Merida-Long, L.B.; Latawiec, E.; Schneider, H.E.; Lipton, J.M.; Vlachos, A.; Atsidaftos, E.; Ball, S.E.; Orfali, K.A.; et al. Ribosomal Protein S24 Gene Is Mutated in Diamond-Blackfan Anemia. Am. J. Hum. Genet. 2006, 79, 1110–1118. [Google Scholar] [CrossRef]
- Deliu, L.P.; Turingan, M.; Jadir, D.; Lee, B.; Ghosh, A.; Grewal, S.S. Serotonergic neuron ribosomal proteins regulate the neuroendocrine control of Drosophila development. PLoS Genet. 2022, 18, e1010371. [Google Scholar] [CrossRef]
- Holt, C.E.; Martin, K.C.; Schuman, E.M. Local translation in neurons: Visualization and function. Nat. Struct. Mol. Biol. 2019, 26, 557–566. [Google Scholar] [CrossRef]
- Singh, A.K.; Abdullahi, A.; Soller, M.; David, A.; Brogna, S. Visualisation of ribosomes in Drosophila axons using Ribo-BiFC. Biol. Open 2019, 8, bio047233. [Google Scholar] [CrossRef]
- Lottes, E.N.; Cox, D.N. Homeostatic Roles of the Proteostasis Network in Dendrites. Front. Cell. Neurosci. 2020, 14, 264. [Google Scholar] [CrossRef]
- Nakahata, Y.; Yasuda, R. Plasticity of spine structure: Local signaling, translation and cytoskeletal reorganization. Front. Synaptic. Neurosci. 2018, 10, 29. [Google Scholar] [CrossRef] [PubMed]
- Thibault, S.T.; Singer, M.A.; Miyazaki, W.Y.; Milash, B.; Dompe, N.A.; Singh, C.M.; Buchholz, R.; Demsky, M.; Fawcett, R.; Francis-Lang, H.L.; et al. A complementary transposon tool kit for Drosophila melanogaster using P and piggyBac. Nat. Genet. 2004, 36, 283–287. [Google Scholar] [CrossRef]
- Hu, Y.; Roesel, C.; Flockhart, I.; Perkins, L.; Perrimon, N.; Mohr, S. UP-TORR: Online Tool for Accurate and Up-to-Date Annotation of RNAi Reagents. Genetics 2013, 195, 37–45. [Google Scholar] [CrossRef]
- Iyer, E.P.R.; Iyer, S.C.; Sulkowski, M.J.; Cox, D.N. Isolation and purification of Drosophila peripheral neurons by magnetic bead sorting. J. Vis. Exp. 2009, 34, e1599. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Nanda, S.; Chen, H.; Das, R.; Bhattacharjee, S.; Cuntz, H.; Torben-Nielsen, B.; Peng, H.; Cox, D.N.; De Schutter, E.; Ascoli, G.A. Design and implementation of multi-signal and time-varying neural reconstructions. Sci. Data 2018, 5, 170207. [Google Scholar] [CrossRef] [PubMed]
- Akram, M.A.; Nanda, S.; Maraver, P.; Armañanzas, R.; Ascoli, G.A. An open repository for single-cell reconstructions of the brain forest. Sci. Data 2018, 5, 180006. [Google Scholar] [CrossRef]
- Erdmann, I.; Marter, K.; Kobler, O.; Niehues, S.; Bussmann, J.; Müller, A.; Abele, J.; Storkebaum, E.; Thomas, U.; Dieterich, D.C. Cell Type-specific Metabolic Labeling of Proteins with Azidonorleucine in Drosophila. Bio-Protocol 2017, 7, e2397. [Google Scholar] [CrossRef] [PubMed]
- Marter, K.; Kobler, O.; Erdmann, I.; Soleimanpour, E.; Landgraf, P.; Müller, A.; Abele, J.; Thomas, U.; Dieterich, D.C. Click Chemistry (CuAAC) and Detection of Tagged de novo Synthesized Proteins in Drosophila. Bio-Protocol 2019, 9, e3142. [Google Scholar] [CrossRef] [PubMed]
- Dennis, G.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 2003, 4, R60. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhattacharjee, S.; Iyer, E.P.R.; Iyer, S.C.; Nanda, S.; Rubaharan, M.; Ascoli, G.A.; Cox, D.N. The Zinc-BED Transcription Factor Bedwarfed Promotes Proportional Dendritic Growth and Branching through Transcriptional and Translational Regulation in Drosophila. Int. J. Mol. Sci. 2023, 24, 6344. https://doi.org/10.3390/ijms24076344
Bhattacharjee S, Iyer EPR, Iyer SC, Nanda S, Rubaharan M, Ascoli GA, Cox DN. The Zinc-BED Transcription Factor Bedwarfed Promotes Proportional Dendritic Growth and Branching through Transcriptional and Translational Regulation in Drosophila. International Journal of Molecular Sciences. 2023; 24(7):6344. https://doi.org/10.3390/ijms24076344
Chicago/Turabian StyleBhattacharjee, Shatabdi, Eswar Prasad R. Iyer, Srividya Chandramouli Iyer, Sumit Nanda, Myurajan Rubaharan, Giorgio A. Ascoli, and Daniel N. Cox. 2023. "The Zinc-BED Transcription Factor Bedwarfed Promotes Proportional Dendritic Growth and Branching through Transcriptional and Translational Regulation in Drosophila" International Journal of Molecular Sciences 24, no. 7: 6344. https://doi.org/10.3390/ijms24076344
APA StyleBhattacharjee, S., Iyer, E. P. R., Iyer, S. C., Nanda, S., Rubaharan, M., Ascoli, G. A., & Cox, D. N. (2023). The Zinc-BED Transcription Factor Bedwarfed Promotes Proportional Dendritic Growth and Branching through Transcriptional and Translational Regulation in Drosophila. International Journal of Molecular Sciences, 24(7), 6344. https://doi.org/10.3390/ijms24076344